

Палеонтологический журнал, 2020, № 2, стр. 50-55
Olibrolitus, новый род семейства Phalacridae (Coleoptera, Cucujoidea) из саксонского и балтийского янтаря
Г. Ю. Любарский a, *, Е. Э. Перковский b, c, **
a Зоологический музей Московского государственного университета
Москва, Россия
b Институт зоологии им. И.И. Шмальгаузена НАН Украины
Киев, Украина
c Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: lgeorgy@rambler.ru
** E-mail: perkovsk@gmail.com
Поступила в редакцию 26.03.2019
После доработки 08.04.2019
Принята к публикации 08.04.2019
Аннотация
Описывается Olibrolitus katyae gen. et sp. nov. из позднеэоценового янтаря. Новый род не входит в современные группы родов Phalacridae. Из эоценового янтаря указаны находки нескольких современных родов, а также недавно описан род Neolitochropus, однако новый род Olibrolitus не близок к этому роду, и пока трудно указать его связь с какими-либо современными родами. Новый род отличается от других родов семейства по отсутствию пришовных линий на надкрыльях и строению лапок (первый членик задних лапок длиннее второго, сочленены свободно).
ВВЕДЕНИЕ
Небольшое семейство гладышей (Coleoptera: Cucujoidea: Phalacridae) содержит более 600 видов в мировой фауне (Gimmel, 2013: 52 рода, 635 видов), представлено во всех зоогеографических регионах. В Палеарктике насчитывается 146 видов (Švec, 2007). У жуков этого семейства тело гладкое, часто блестящее, не покрыто волосками, размер маленький, усики 11-члениковые с 3-члениковой булавой (изредка булава из 4 или 5 члеников), формула лапок 5-5-5 (изредка 5-5-4, 4-4-4, 4-5-4).
Наиболее древние ископаемые находки гладышей известны из верхнеальбского испанского янтаря (Peris et al., 2016). Помимо этого, Phalacridae известны из верхнеэоценовых балтийского и ровенского янтарей (Phalacrus Paykull, Olibrus Erichson и Stilbus Seidlitz: Klebs, 1910; Hieke, Pietrzeniuk, 1984; Lyubarsky, Perkovsky, 2011, 2016) и среднемиоценовых доминиканского и мексиканского янтарей (этот материал не определен до родового уровня; Spahr, 1981). A.Г. Кирейчук и A. Нель (Kirejtshuk, Nel, 2008) отметили находки Phalacridae в нижнеэоценовом янтаре Уазы. В основном из эоцена указывались современные роды, до того как Г.Ю. Любарский и Е.Э. Перковский (Lyubarsky, Perkovsky, 2016) описали из эоценового янтаря новый род Neolitochropus Lyubarsky et Perkovsky. Ископаемый род Neolitochropus связан с современными родами, распространенными в тропиках. Таким образом, из эоценового янтаря известны роды Phalacrus, Olibrus, Stilbus, Neolitochropus, а также новый род, описываемый в данной работе.
Новый род не близок ни к одному современному роду; он найден в коллекции янтаря К. и Х. Хоффейнсов (Christel and Hans Werner Hoffeins, Hamburg, Germany, CCHH). Материалы, описываемые в статье, будут храниться в коллекции янтаря Senckenberg Deutsches Entomologisches Institut, Müncheberg, Germany (SDEI).
Фотографии сделаны при помощи стереомикроскопа Leica M165C в центре коллективного использования научного оборудования “Animalia”, в Ин-те зоологии им. И.И. Шмальгаузена НАН Украины, Киев.
СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
ОТРЯД COLEOPTERA
НАДСЕМЕЙСТВО CUCUJOIDEA LATREILLE, 1802
СЕМЕЙСТВО PHALACRIDAE LEACH, 1815
Род Olibrolitus Lyubarsky et Perkovsky, gen. nov.
Название рода составлено из комбинации названий родов Olibrus и Ochrolitus Sharp, поскольку новый род похож на оба эти рода; м. р.
Типовой вид – Olibrolitus katyae sp. nov.
Диагноз. Тело голое, усики 11-члениковые, с 3-члениковой булавой, членики булавы поперечные. Формула лапок 5-5-5, членики лапок с лопастинками, 4-й членик с сильно расширенными лопастинками. Продольные линии на надкрыльях отсутствуют. Щиток небольшой, поперечный. Ктенидий на передних голенях отсутствует, передние голени без длинных шпор на наружных углах вершины. Средние голени с двумя шпорами на вершине. Вырост заднегруди широкий, шире, чем диаметр средних тазиков, т. е. средние тазики отчетливо разделены. Метавентральные линии не отделены от тазиковых впадин. 1-й членик задних лапок длиннее 2-го.
Сравнение. См. “Обсуждение” ниже.
Olibrolitus katyae Lyubarsky et Perkovsky, sp. nov.
Табл. VII , фиг. 1–3
Рис. 1.
Olibrolitus katyae gen. et sp. nov., голотип CCHH 1206-2, общий вид: а – вентрально; б – дорсально.
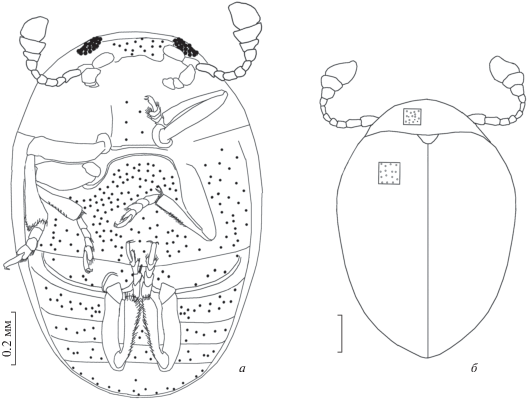
Голотип – CCHH 1206-2, хранится в коллекции янтаря SDEI; саксонский янтарь, верхний эоцен.
Вид назван в честь Екатерины (Кати) Сидорчук (1981–2019).
Описание (рис. 1). Тело короткое, овальное, не покрыто волосками, слабо блестящее, размер маленький; черный верх без светлых отметин, нижняя сторона блестящая (табл. VII , фиг. 1а, 1б). Длина тела в 1.5 раза больше ширины.
Голова. Сильно поперечная, несколько уплощенная. Пунктировка головы слабая, редкая, расстояние между точками составляет 1–2 диаметра точки. Глаза небольшого размера, не окантованы, тонко фасетированы, фасетки выпуклые. Фронтоклипеальная область выступает над местами прикрепления усиков, вершина клипеуса обрезана. Фронтоклипеальный шов отсутствует, усиковые бороздки отсутствуют. Последний членик нижнечелюстных щупиков длиннее предыдущего. Усики заходят за задний край переднеспинки. 3-члениковая булава усика слегка асимметрична, членик 9-й конический, членик 10‑й поперечный, членик 11-й овальный, не суженный. Членики усиков 4-8 примерно равной длины, каждый членик в 1.5 раза длиннее ширины (рис. 1). Членик усика 2 самый длинный, членик 7 немного длиннее членика 5.
Грудь. Переднеспинка сильно поперечная, слабо и редко пунктирована, с умеренно развитой прищитковой лопастью. Основание переднеспинки не окантовано. Щиток треугольный, со слегка округленными углами, очень слабо пунктированный. Задние углы переднеспинки налегают на основание надкрылий. Вырост переднегруди широкий, округленный при виде сбоку, обрезанный апикально, по бокам не окантованный, без крупных хет на вершине. Передние тазики широко разделены. Щиток маленький, треугольный. Средние тазики широко разделены, расстояние между ними больше одного диаметра средней тазиковой впадины. Вырост заднегруди не выдается за передний край средних тазиков, имеет равномерно выпуклый передний край. Заднегрудь сильно и редко пунктирована, расстояние между точками составляет 2–3 диаметра точки. Задние тазики соприкасаются. Вырост заднегруди заходит за передний край средних тазиков, на вершине образует равномерно выпуклую лопасть. Передний край задних тазиков без окантовки. Продольная заднегрудная линия отсутствует. Эпиплевры полностью развиты, сужены начиная с уровня заднегруди. Надкрылья без пришовных линий, пунктировка надкрыльев спутанная, точки слегка удлиненные, без поперечных складочек. Длина надкрыльев в 1.2 раза больше ширины.
Объяснения к таблице VII
Фиг. 1–3 . Olibrolitus katyae gen. et sp. nov.: 1 – голотип CCHH 1206-2, общий вид: 1а – дорсально; 1б – вентрально; 2 – паратип CCHH 1206-3, усики, передние и задние ноги; 3 – паратип CCHH 1223-3, задние лапки и голени с одной шпорой.
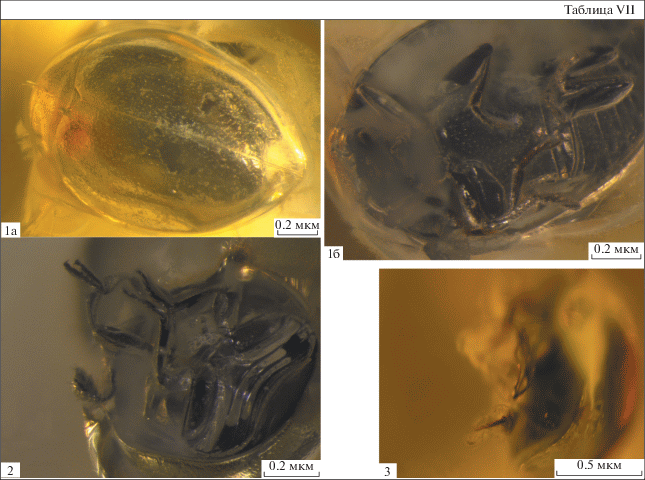
Ноги довольно стройные, бедра и голени не расширены (табл. VII , фиг. 1б). Передние голени стройные, без шпор (табл. VII , 1б). Ктенидий на передних голенях отсутствует, однако на них имеются длинные жесткие щетинки по всей длине голени. Средние голени с двумя шпорами, достигающими в длину приблизительно половины длины 1‑го членика лапок, длина шпор примерно равна ширине голени на вершине. Задние голени без апикального ктенидия, с одной шпорой (табл. VII , фиг. 2, 3 ). Членики лапок с длинными хетами в апикальной части, слегка лопастевидные, 4 членик задних лапок с сильно развитыми лопастинками, между которыми расположен 5-й членик. Формула лапок 5-5-5 (пол экземпляра неизвестен). Задние лапки по длине примерно равны другим лапкам. Соединение 1-го и 2-го члеников лапок свободное. 2-й членик задних лапок в 1.5 раза длиннее 3-го членика. 3-й членик задних лапок в 2 раза длиннее 4-го членика, последний с большими лопастинками. Формула шпор на голенях 0-2-1.
Брюшко. Состоит из 5 свободно сочлененных сегментов. Сегмент 1 в 1.5 раза длиннее сегмента 2 (рис. 1, б). Сегменты 2, 3 и 4 приблизительно равной длины. Бедренные линии на 1-м сегменте брюшка отсутствуют. Сегменты слабо и редко пунктированы; сегмент 5 без бороздок или гребней. 1–4 брюшные сегменты слегка пунктированы. Сегмент 5 выпуклый, с хеттами на заднем крае, сильно пунктированный.
Размеры в мм. Длина тела голотипа 1.4; длина надкрыльев голотипа 1.1, ширина – 0.9, длина переднеспинки 0.3, ширина 0.7. Паратипы: CCHH 1206-3 – длина тела 1.2, ширина 0.7; CCHH 1223-3 – длина тела 1.4, ширина 0.7.
Материал. Голотип и паратипы: CCHH 1206-3 и CCHH 1223-3, оба паратипа из балтийского янтаря, верхний эоцен (SDEI), из Янтарного (Palmnicken). Голотип и паратипы в хорошем состоянии.
ОБСУЖДЕНИЕ
Последняя ревизия семейства на уровне родов была выполнена М. Гиммелем (Gimmel, 2013). Он выделил 9 групп родов: Phaenocephalus, Stilbus, Pseudolibrus, Phalacrus, Olibroporus, Ochrolitus, Olibrus и Litochropus; в каждую группу входят 2–4 рода. Кроме этих групп родов, Гиммель оставил 9 родов вне групп, как группы incertae sedis, без соотнесения с какой-либо группой.
В разделе работы, посвященной филогении семейства, Гиммель пишет (Gimmel, 2013, с. 131): “The attempt at formulation of a higher classification in the family Phalacridae is beset with difficulties. Surveying the genera in a pre-phylogenetic context, one finds no overt structural syndromes upon which to divide up the family into mutually exclusive units. Instead, one is presented with an exasperating number of permutations of mostly binary characters. Even the would-be constructor of a purely phenetic classification would be stymied by the amount of convergent evolution that has apparently occurred within the group, since no one character system or systems in combination seem to emerge above the rest to aid in creation of stable subdivisions.”
С помощью кладистического анализа роды incertae sedis не могут быть помещены в группы: “The level of resolution from this phylogenetic analysis was inadequate to place the remaining genera convincingly into groups” (Gimmel, 2013, с. 141).
В такой ситуации, когда признаки родов довольно свободно комбинируются, многие родовые признаки конвергентны, нет возможности решить, к какой группе родов относится новый род. Поэтому нет возможности сделать заключение, в каком зоогеографическом регионе обитают гладыши, близкородственные новому роду.
При сравнении признаков современных родов можно заключить, что многие признаки комбинативны, свободно сочетаются, и новый род обладает новой комбинацией признаков, в ином сочетании свойственных современным родам. Современные роды показывают свободное сочетание нескольких групп признаков, важных при определении родового ранга в этом семействе. Эти свободно сочетающиеся признаки родового ранга можно объединить в несколько групп: 1) отношение длины 1 и 2 члеников задних лапок; 2) положение границы между средне- и заднегрудью и расположение средних тазиков; 3) отношение расстояния между средними тазиками и диаметра средних тазиков; 4) формула лапок, т.е. число члеников на передних, средних и задних лапках; 5) наличие или отсутствие ктенидия на передних голенях; 6) формула шпор, т.е. число и относительная длина шпор на передних, средних и задних голенях; 7) наличие метавентральных линий: эти линии либо отставлены от границы средних тазиков, либо сближены с этой границей; и другие признаки. Новый род представляет новый вариант комбинации родовых признаков. Новый род характеризуется отсутствием пришовных линий на надкрыльях, 1-й и 2-й членики задних лапок приблизительно равны по длине, 1-й членик немного длиннее, членики соединены свободно, не слиты.
Таким образом, новый род отличается от современных родов в основном отсутствием пришовных линий на надкрыльях и строением члеников задних лапок. Например, у нового рода нет пришовных линий на надкрыльях, как у Phaenocephalus Wollaston, Eulitrus Sharp или Phalacropsis Casey, однако по другим признакам новый род явно не схож с этими родами, особенно по строению лапок. У Phaenocephalus формула лапок 4-4-4; у Eulitrus 2-й членик задних лапок длиннее 1-го членика. По строению заднегруди новый род несколько похож на род Olibrus, но отличается уже по отсутствию пришовных линий на надкрыльях. По строению задних лапок новый род несколько напоминает Ochrolitus. 2-й членик задних лапок примерно равен длине 1-го членика, подобно Entomocnemus Guillebeau, но от этого рода имеются сильные отличия в строении выроста заднегруди.
Отличия от недавно описанного рода Neolitochropus следующие. У Neolitochropus лапки без лопастинок, а у нового рода все членики лапок, и особенно 4‑й членик, несут хорошо развитые лопасти; у Neolitochropus на каждом надкрылье имеется пришовная линия, а у нового рода линий на надкрыльях нет.
Новый род не может быть размещен в какой-либо группе родов, выделенной в ревизии (Gimmel, 2013), так что этот род относится, подобно многим другим родам, к Phalacridae incertae sedis.
Это второй род Phalacridae, известный только из эоценового янтаря. Среди близких семейств Cucujoidea мы можем наблюдать две типичные ситуации. Одна характерна для семейства Cryptophagidae. Почти вся меловая фауна Cryptophagidae состоит из недавно описанных родов из Китая, с Таймыра и из Испании, вымерших, как и первый меловой род Nganasania Zherikhin, 1977 (Lyubarsky, Perkovsky, 2017с), на границе мелового периода. Эоценовая фауна этого семейства до недавнего описания нового рода из балтийского янтаря (Lyubarsky, Perkovsky, 2019b) содержал только современные роды (несколько вымерших родов были перенесены в другие семейства: Lyubarsky, Perkovsky, 2018b), было описано несколько новых видов из современных родов (Lyubarsky, Perkovsky, 2019a и ссылки в этой работе). В результате выяснилось, что меловая фауна имела совсем иной состав родов, нежели эоценовая и современная, т.е. меловая фауна полностью вымерла (вероятно, на границе мела), и уже в эоцене стала появляться современная фауна. Современная фауна на родовом уровне не отличается от эоценовой.
Для семейства Erotylidae меловая фауна неизвестна. В то же время, ранее мы описали пять вымерших эоценовых родов (Lyubarsky, Perkovsky, 2012, 2017a, b, 2018a; Lyubarsky et al., 2016). Таким образом, вымирание многих родов Erotylidae произошло между эоценом и современностью, а не между мелом и эоценом, как у Cryptophagidae.
В семействе гладышей (Phalacridae) ситуация похожа на то, что можно видеть у Erotylidae. Для этого семейства известно лишь несколько находок, и они в основном относятся к эоцену. Описано два вымерших монотипических рода (Neolitochropus и теперь Olibrolitus. Первый из родов после подачи этой статьи перемещен в мезозойское семейство Cyclaxyridae, современное распространение которого ограничено Новой Зеландией; Gimmel et al., 2019). Очевидно, изменения, которые лежат в основе формирования современной фауны этого семейства, произошли позже эоценового времени.
Два из трех родов Phalacridae из янтарей с южного побережья Субпаратетиса (саксонский и ровенский янтарь) не известны из балтийского янтаря. Это отличается от распространения Erotylidae в янтарных лесах (три из четырех родов с южного берега Субпаратетиса неизвестны из балтийского янтаря: Lyubarsky, Perkovsky, 2018a и ссылки в этой работе), и распространения Hymenoptera (по крайней мере 16 родов из 74 неизвестны из балтийского янтаря: Perkovsky, 2018 и неопубликованные данные второго автора). Все это свидетельствует о существенных климатических отличиях между балтийским янтарным лесом и янтарными лесами южного побережья Субпаратетиса (Wolfe et al., 2016; Mänd et al., 2018). В то же время, лишь 15 и 65 описанных видов саксонских (биттерфельдских) муравьев (и 26 из 71 ровенских) неизвестны из балтийского янтаря (Dubovikoff et al., 2019, 2020; Radchenko, Perkovsky, 2020). Даже с учетом того, что многие ровенские и саксонские виды муравьев еще не описаны (Perkovsky, 2016), это говорит об одновозрастности балтийской, саксонской и ровенской фаун (Dlussky, Rasnitsyn, 2009), поэтому обнаружение нового вида фалакриды с широким ареалом не вызывает удивления.
* * *
Мы благодарны К. и Х. Хоффайнсам (Гамбург, ФРГ) за предоставление материала для изучения. Мы благодарим В.Ю. Назаренко (Ин-т зоологии им. И.И. Шмальгаузена НАН Украины, Киев) за сделанные фотографии типовых экземпляров.
Работа частично поддержана исследовательским грантом НИ Зоологического музея МГУ (АААА-А16-116021660077-3).
Список литературы
Dlussky G.M., Rasnitsyn A.P. Ants (Insecta: Vespida: Formicidae) in the Upper Eocene amber of Central and Eastern Europe // Paleontol. J. 2009. V. 43. № 9. P. 1024–1042.
Dubovikoff D.A., Dlussky G.M., Perkovsky E.E., Abakumov E.V. A new species of the genus Eldermyrmex Shattuck, 2011 (Hymenoptera, Formicidae) from Bitterfeld amber (Late Eocene) with species key of the genus // Paleontol. J. 2019. V. 53. № 10. P. 994–997.
Dubovikoff D.A., Dlussky G.M., Perkovsky E.E., Abakumov E.V. A new species of the genus Protaneuretus Wheeler, 1915 (Hymenoptera, Formicidae) from Bitterfeld amber (Late Eocene) with species key of the genus // Paleontol. J. 2020. B пeчaти.
Gimmel M.L. Genus-level revision of the family Phalacridae (Coleoptera: Cucujoidea) // Zootaxa. 2013. V. 3605. № 1. P. 1–147.
Gimmel M.L., Szawaryn K., Cai C., Leschen R.A.B. Mesozoic sooty mould beetles as living relicts in New Zealand // Proc. Roy. S. B. 2019. V. 286. Issue 1917. in press. https://doi.org/10.1098/rspb.2019.2176
Hieke F., Pietrzeniuk E. Die Bernstein-Käfer des Museums für Naturkunde, Berlin (Insecta, Coleoptera) // Mitt. Zool. Museum Berlin. 1984. V. 60. № 2. S. 297–326.
Kirejtshuk A.G., Nel A. New beetles of the suborder Polyphaga from the Lowermost Eocene French amber (Insecta: Coleoptera) // Ann. Soc. Entomol. France (n.s.). 2008. V. 44. № 4. P. 419–442.
Klebs R. Über Bernsteineinschlüsse im allgemeinen und die Coleopteren meiner Bernsteinsammlung // Schr. Phys.-Ökon. Ges. Königsberg. 1910. V. 51. S. 217–242.
Lyubarsky G.Yu., Perkovsky E.E. New species of Stilbus (Coleoptera, Clavicornia, Phalacridae) from the Late Eocene Rovno amber // Vestnik zool. 2011. V. 45. № 2. P. 187–190.
Lyubarsky G.Yu., Perkovsky E.E. A new genus of Erotylidae from Eocene amber (Coleoptera: Clavicornia) // Russ. Entomol. J. 2012. V. 21. № 1. P. 35–38.
Lyubarsky G.Yu., Perkovsky E.E. A new genus, Neolitochropus (Coleoptera: Cucujoidea: Phalacridae), from the Upper Eocene Bitterfeld amber // Russ. Entomol. J. 2016. V. 25. № 3. P. 249–253.
Lyubarsky G.Yu., Perkovsky E.E. Serramorphus, a new genus of Erotylidae from Eocene amber (Coleoptera: Clavicornia) from Late Eocene Bitterfeldian amber // Russ. Entomol. J. 2017a. V. 26. № 1. P. 37–40.
Lyubarsky G.Yu., Perkovsky E.E. Xenophagus, a new genus of pleasing fungus beetles (Coleoptera: Erotylidae) from Baltic amber // Russ. Entomol. J. 2017b. V. 26. № 2. P. 147–150.
Lyubarsky G.Yu., Perkovsky E.E. Re-description of the genus Nganasania Zherikhin, 1977 from Upper Cretaceous of Taimyr (Coleoptera: Cryptophagidae) // Russ. Entomol. J. 2017с. V. 26. № 3. P. 251–255.
Lyubarsky G.Yu., Perkovsky E.E. Microzavaljus, a second extinct genus of pleasing fungus beetles (Coleoptera: Erotylidae) from Bitterfeld amber // Russ. Entomol. J. 2018a. V. 20. № 3. P. 271–276.
Lyubarsky G.Yu., Perkovsky E.E. First species of Atomaria (Coleoptera: Clavicornia: Cryptophagidae) from Bitterfeld amber and several notes on Miocene and Cretaceous Cryptophagidae // Russ. Entomol. J. 2018b. V. 20. № 4. P. 381–385.
Lyubarsky G.Yu., Perkovsky E.E. First species of Ephistemus (Coleoptera: Clavicornia: Cryptophagidae) from Baltic amber // Russ. Entomol. J. 2019a. V. 28. № 1. P. 54–57.
Lyubarsky G.Yu., Perkovsky E.E. Spaniophagus, first new Eocene genus of silken fungus beetle from Baltic amber (Coleoptera: Clavicornia: Cryptophagidae) // Russ. Entomol. J. 2019b. V. 28. № 3. P. 263–268.
Lyubarsky G.Yu., Perkovsky E.E., Alekseev V.I. The first record of the subfamily Xenoscelinae (Coleoptera, Erotylidae) from the Baltic amber // Paleontol. J. 2016. V. 50. № 9. P. 963–969.
Mänd K., Muehlenbachs K., McKellar R.C. et al. Distinct origins for Rovno and Baltic ambers: Evidence from carbon and hydrogen stable isotopes // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2018. V. 505. P. 265–273.
Peris D., Ruzzier E., Perrichot V., Delclòs X. Evolutionary and paleobiological implications of Coleoptera (Insecta) from Tethyan-influenced Cretaceous ambers // Geosci. Frontiers. 2016. V. 7. № 4. P. 695–706.
Perkovsky E.E. Tropical and Holarctic ants in Late Eocene ambers // Vestnik zool. 2016. V. 50. № 2. P. 111–122.
Perkovsky E.E. Only half of Rovno Amber hymenopteran fauna is common with Baltic amber // Vestnik zool. 2018. V. 52. № 5. P. 353–360.
Radchenko A.G., Perkovsky E.E. New records of the fossil ant genus Prionomyrmex Mayr (Hymenoptera, Formicidae, Myrmeciinae) from Late Eocene European ambers // Paleontol. J. 2020. V. 54. B пeчaти.
Spahr U. Systematischer Katalog der Bernstein und Kopal–Käfer (Coleoptera) // Stuttg. Beitr. Naturk. Ser. B. Geol. Paläontol. 1981. V. 80. S. 1–107.
Švec Z. Phalacridae // Catalogue of Palaearctic Coleoptera. Vol. 4. Elateroidea–Derodontoidea–Bostrichoidea–Lymexyloidea–Cleroidea–Cucujoidea / Eds Löbl I., Smetana A. Stenstrup: Apollo Books, 2007. P. 64–65, 506–513.
Wolfe A.P., McKellar R.C., Tappert R. et al. Bitterfeld amber is not Baltic amber: Three geochemical tests and further constraints on the botanical affinities of succinite // Rev. Palaeobot. Palynol. 2016. V. 225. P. 21–32.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал