

Палеонтологический журнал, 2020, № 2, стр. 40-49
Новые гриллоновые насекомые (Insecta: Gryllones) из местонахождения Бабий Камень (верхняя пермь России). 2. Отряд Reculida и Gryllones incertае ordinis
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Череповецкий государственный университет
Череповец, Россия
* E-mail: danil_aristov@mail.ru
Поступила в редакцию 07.03.2019
После доработки 05.04.2019
Принята к публикации 05.04.2019
Аннотация
Из верхнепермского местонахождения Бабий Камень (вятский ярус России) описаны новые таксоны отряда Reculida (Insecta: Gryllones): имаго Chauliodites babiy sp. nov. (семейство Chaulioditidae), Stegopterum kabalum sp. nov., S. pashka sp. nov., Permoshurabia tshasha sp. nov., Permuliercula chimaera gen. et sp. nov. (семейство Geinitziidae), Khosara ultima sp. nov. (семейство Liomopteridae), Permotoma aba gen. et sp. nov. (семейство Skaliciidae) и нимфа Tarakanikha satura gen. et sp. nov. (Gryllones incertае ordinis). Shurabia annosa Aristov, 2004 переописан и отнесен к роду Geinitzia.
Данная статья является продолжением работы по описанию гриллоновых насекомых из местонахождения Бабий Камень (тараканихинские и кедровские слои мальцевской свиты). Это местонахождение расположено в Кемеровской обл. России и относится к вятскому ярусу верхней перми (Садовников, 2016). По мнению других авторов (Щербаков, 2008), Бабий Камень имеет нижнетриасовый возраст. В первой части данной работы (Аристов, 2020) были рассмотрены эоблаттидовые насекомые. Ниже рассматриваются представители отряда Reculida, принадлежащего к надотряду Blattidea инфракласса Gryllones, и нимфа неясного положения, относящаяся к этому же инфраклассу. Кроме того, рассмотрена фауна всех гриллоновых насекомых местонахождения Бабий Камень и проведено ее сравнение с другими позднепермскими, ранне- и среднетриасовыми фаунами.
Инфракласс Gryllones представлен в Бабьем Камне прямокрылообразными и тараканообразными гриллоновыми насекомыми. К первым относятся прямокрылые Mesoedischiidae (Горохов, 2005). Тараканообразные (состав надотряда дан по: Aristov, 2015a) представлены эоблаттидовыми (Аристов, 2020) и их потомками: тараканами (Blattida: Subioblattidae: Rasnitsyn et al., 2013), рекулидовыми (см. ниже) и эмбиями (Embiida: Alexarasniidae: Shcherbakov, 2015).
Фауна рекулидовых насекомых (Insecta: Reculida) кедровских слоев Бабьего Камня до настоящего момента включала два рода и четыре вида семейства Chaulioditidae (Аристов, 2003) и один род с одним видом семейства Geinitziidae (Аристов, 2004). Ниже описаны еще один вид Chaulioditidae, четыре вида, относящихся к трем родам семейства Geinitziidae и по одному представителю Liomopteridae и Skaliciidae. Из перечисленных семейств Chaulioditidae и Geinitziidae известны как из перми, так и из триаса, а Skaliciidae и Liomopteridae – только из перми (Rasnitsyn et al., 2013). Кроме того, описана нимфа неясного положения, отнесенная нами к гриллоновым насекомым, – Tarakanikha satura gen. et sp. nov., которая была найдена в тараканихинских слоях мальцевской свиты. Эти слои древнее кедровских, но также относятся к местонахождению Бабий Камень. Кроме этой нимфы, в тараканихинских слоях найдено переднее крыло хаулиодитиды Chauliodites, не отличающееся от Ch. cancellatus (Aristov, 2003), описанного из кедровских слоев (Аристов, 2003). По количеству экземпляров доминируют Chaulioditidae – 68% (два рода, пять видов), меньше Geinitziidae – 27% (четыре рода, шесть видов), Skaliciidae и Liomopteridae единичны – 3 и 2%, соответственно (по одному роду и виду).
По составу с Бабьим Камнем сходны одновозрастные фауны рекулидовых Тунгусского бассейна (вятский ярус Кемеровской обл. России: Садовников, 2016). В местонахождениях Нирунгдакан-1, Нижняя Люлюикта-1, Анакит-1 и Анакит-3 найдены Chaulioditidae и Geinitziidae, представленные одним родом с двумя видами каждое (Аристов, 2011). Отсутствие Skaliciidae и Liomopteridae может объясняться меньшим объемом сборов – 62 экз. в Бабьем Камне и 11 экз. в местонахождениях Тунгуски. В Тунгуске доминируют хаулиодитиды, составляющие 82%.
Также Бабий Камень сходен по составу рекулидовых с верхнепермским (северодвинским) местонахождением Исады в Вологодской обл. России (Aristov, 2013b). В этом местонахождении найдены доминирующие Chaulioditidae, составляющие 52% остатков рекулидовых (четыре рода, семь видов), Geinitziidae – 36% (три рода, три вида) и Liomopteridae – 12% (четыре рода, четыре вида). Следует отметить, что доля эоблаттидовых и рекулидовых от всех насекомых в Бабьем Камне заметно выше, чем в Исадах.
Фауна рекулидовых вятских местонахождений Соковка и Балымотиха во Владимирской обл. и Недуброво в Вологодской обл. представлена только Chaulioditidae (Aristov, 2013a). Тот же состав характерен и для нижнетриасовой (оленекской) фауны Петропавловки в Оренбургской обл. России (неопубликованные данные автора) и среднетриасовых (анизийских) местонахождений Германии (Bashkuev et al., 2012) и Франции (Аристов и др., 2011). Бабий Камень выделяется на фоне вятско–анизийских фаун рекулидовых своим относительно высоким разнообразием (наличием Skaliciidae и Liomopteridae). За этим исключением, фауны рекулидовых конца перми и начала триаса существенных различий не имеют.
Из найденных в Бабьем Камне эоблаттидовых Babalidae являются эндемиком (Аристов, 2020), Megakhosaridae и Blattogryllidae достаточно широко распространены как в верхней перми, так и в среднем триасе (Storozhenko, Aristov, 2014). Раннетриасовые эоблаттидовые на сегодняшний день не известны. Комплекс эоблаттидовых Бабьего Камня существенно отличается от Исад по отсутствию палеозойских элементов. В Исадах зарегистрированы последние находки Permotermopsidae, Atactophlebiidae и Cacurgidae (Aristov et al., 2013). Cостав эоблаттид из Бабьего Камня, за исключением эндемичных Babalidae, характерен как для вятских Недуброво и местонахождений Тунгусского бассейна, так и для анизийских Вогез. В Недуброво найдены Megakhosaridae (Storozhenko, Aristov, 2014), в местонахождении Анакит-3 (Тунгусский Бассейн: Аристов, 2011) и Вогезы найдены и Megakhosaridae и Blattogryllidae, в Вогезах, кроме того, обнаружены Mesorthopteridae (Аристов и др., 2011).
Таким образом, за счет последних находок пермских Liomopteridae и Skaliciidae фауна Reculida Бабьего Камня выглядит скорее пермской. Фауна Eoblattida является характерной как для вятского века верхней перми, так и для среднего триаса, и отличается от остальных фаун этого времени наличием эндемичных Babalidae и отсутствием Mesorthopteridae. Остальные гриллоновые насекомые Бабьего Камня – прямокрылые Mesoedischiidae, тараканы Subioblattidae и эмбии Alexarasniidae – известны из мезозоя (Rasnitsyn et al., 2013).
Состав описанных в данной работе и ранее (Аристов, 2019) гриллоновых не противоречит отнесению мальцевской свиты Кузбасса к терминальной перми (Yan et al., 2019 и др.). Ранее эти отложения рассматривались как нижнетриасовые (Щербаков, 2008). Сообщество считалось кризисным во многом благодаря доминированию Chaulioditidae, рассматриваемых ранее как “таксон катастроф” (“disaster taxon”), т.е., таксон, характерный для резких смен биоты. Впоследствии было показано широкое распространение этого семейства во временных окрестностях границы перми и триаса, и Chaulioditidae перестали рассматриваться в таком качестве (Пономаренко, 2016).
Фауна гриллоновых насекомых Бабьего Камня содержит как палеозойские, так и постпалеозойские (появившиеся в палеозое и перешедшие в мезозой) элементы с преобладанием последних, представленных семью семействами. Палеозойских семейств только три, включая эндемичных Babalidae (см. выше). Учитывая, что четыре семейства Gryllones являются первыми находками мезозойских групп, по составу фауна представляется скорее мезозойской. Более или менее палеозойской группой выглядят только Reculida. Однако это не свидетельствует о триасовом возрасте местонахождения, поскольку высокая доля постпалеозойских семейств характерна и для вятских местонахождений (Rasnitsyn et al., 2013).
Несмотря на то, что отложения Бабьего Камня формировались во время вулканической активности в Сибири (аргиллиты этого местонахождения имеют туфогенную природу: Shcherbakov, 2015), фауна эоблаттидовых и рекулидовых не выглядит кризисной. В крупнейшем верхнепермском местонахождении Исады на 47 экз. эоблаттидовых приходится шесть семейств, на 50 экз. рекулидовых – три семейства. В Бабьем Камне на 14 экз. Eoblattida приходится три семейства, на 62 экз. Reculida – четыре семейства. То есть разнообразие этих двух местонахождений вполне сопоставимо. Еще один отряд тараканообразных, эмбии, впервые появляются в Бабьем Камне (семейство Alexarasniidae Shcherbakov, 2015). Для оставшихся тараканообразных, собственно Blattida, по сравнению с более древними и более молодыми местонахождениями, характерно резкое падение их доли в фауне (в Бабьем Камне доля тараканов составляет 3%). При этом в большинстве вятских и оленекских местонахождениях Европейской России тараканы составляют около половины остатков насекомых. Падает и наблюдаемое разнообразие – найденные немногочисленные тараканы относятся к семейству Subioblattidae и являются первой находкой этого семейства (Rasnitsyn et al., 2013). Тараканы Blattulidae из Бабьего Камня указаны в упомянутой работе ошибочно (П. Вшанский, личн. сообщ.). Палеозойско-мезозойские семейства Phylloblattidae, Argentinoblattidae и Caloblattinidae в Бабьем Камне не найдены (Rasnitsyn et al., 2013), что может быть связано с временным резким (на порядок) сокращением количества тараканов. Такая же низкая доля тараканов наблюдается в примерно одновозрастных с Бабьим Камнем местонахождениях Анакит, Недуброво и Залазна (Кемеровская, Вологодская и Кировская обл. России, соответственно).
На сегодняшний день единственная крупная раннетриасовая фауна насекомых – это оленекская Петропавловка в Оренбургской обл. России. Посткризисная фауна большинства гриллоновых этого местонахождения бедна и представлена только неопределенными прямокрылыми, тараканами и рекулидовыми Chaulioditidae (Щербаков и др., 2019).
По всей видимости, излияние сибирских траппов как таковое не имело катастрофических последствий для гриллоновых насекомых. Напротив, из девяти семейств Gryllones, найденных в Бабьем Камне, три представляют собой первые находки – Babalidae, Alexarasniidae и Mesoedischiidae. Для двух семейств (Liomopteridae и Skaliciidae) находки в Бабьем Камне являются последними, четыре семейства проходят из палеозоя в мезозой (Subioblattidae, Megakhosaridae, Blattogryllidae и Chaulioditidae).
Таким образом, изменения фауны гриллоновых насекомых на рубеже палеозоя и мезозоя имели место, но выразились они в росте разнообразия, а не в его сокращении. Причиной этому, как ни парадоксально, могла быть именно вулканическая активность. Перечисленные выше гриллоновые, кроме Mesoedischiidae, были более или менее тесно связаны с подстилкой и, соответственно, с растениями. Биологическая активность вулканических почв по сравнению с невулканическими зональными почвами повышена. Это создает благоприятные условия для растений и, опосредованно, для стратобионтов.
Наша работа дополнительно подтверждает достаточно высокое, некризисное, разнообразие фауны Бабьего Камня, что не противоречит пермскому возрасту этого местонахождения. Это же касается и одновозрастных с Бабьим Камнем фаун Тунгуски и местонахождения Недуброво (в некоторых работах относимых к триасу: Щербаков, 2008). Фауны гриллоновых этих местонахождений выглядят обедненным вариантом фауны Бабьего Камня.
Известно, что нормированное количество семейств (количество семейств из местонахождения, деленное на натуральный логарифм количества экземпляров из этого местонахождения) возрастает в конце перми (Бабий Камень и Тунгуска: Расницын и др., 2013). Наши новые данные этот рост подтверждают.
В перечисленных выше местонахождениях «кризис» выражен, прежде всего, в низкой доле тараканов (как следствие экологической депрессии), но не в сколько-нибудь заметном падении разнообразия. Учитывая долю новых семейств, это не конец пермского вымирания, а начало мезозойской диверсификации, что было продемонстрировано ранее (Расницын и др., 2013) и подтверждается нашими новыми данными.
Автор признателен А.П. Расницыну, А.Г. Пономаренко (ПИН РАН) и С.Ю. Стороженко (ФНЦ Биоразнообразия ДВО РАН) за замечания по рукописи. Работа поддержана Программой № 15 Президиума Российской Академии Наук “Проблемы происхождения жизни и становления биосферы” и грантом РФФИ № 18-04-00322.
НАДОТРЯД BLATTIDEA
ОТРЯД RECULIDA
СЕМЕЙСТВО CHAULIODITIDAE HANDLIRSCH, 1906
Род Chauliodites Heer, 1864
Chauliodites babiy Aristov, sp. nov.
Табл. VI , фиг. 1 (см. вклейку)
Рис. 1.
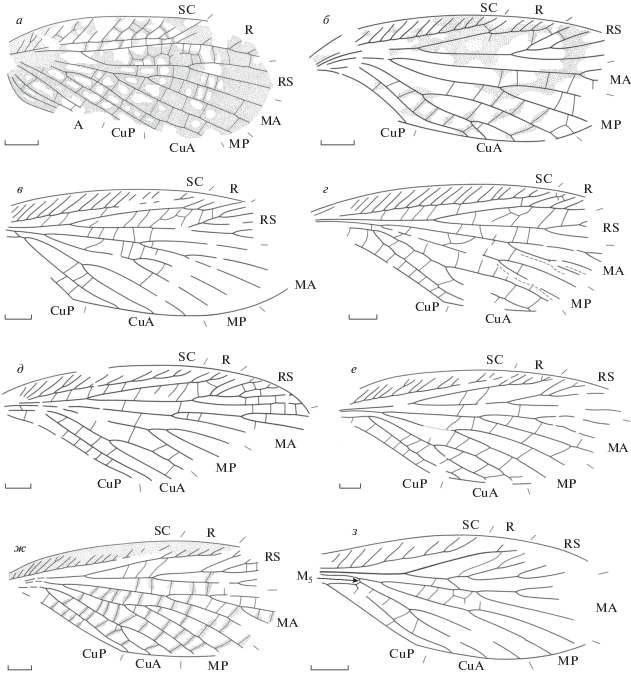
Представители отряда Reculida из Бабьего Камня (Кемеровская обл.; верхняя пермь, вятский ярус), передние крылья: а – Chauliodites babiy sp. nov., голотип ПИН, № 4887/212 (семейство Chaulioditidae); б – Geinitzia annosa (Aristov, 2004) comb. nov., голотип ПИН, № 4887/28; в –Stegopterum kabalum sp. nov., голотип ПИН, № 4887/520; г – S. pashka sp. nov., голотип ПИН, № 4887/533; д–ж – Permoshurabia tshasha sp. nov.: д – голотип ПИН, № 4887/529, е – паратип ПИН, № 4887/555, ж – паратип ПИН, № 4887/519; з – Permuliercula chimaera gen. et sp. nov., голотип ПИН, № 4887/223 (семейство Geinitziidae). Длина масштабной линейки на рис. 1 и 2 соответствует 1 мм.
Название вида от местонахождения Бабий Камень.
Голотип – ПИН, № 4887/212, прямой и обратный отпечатки переднего крыла; Кемеровская обл., Новокузнецкий р-н, правый берег р. Томь в 10 км ниже п. Усть-Нарык, местонахождение Бабий Камень; верхняя пермь, вятский ярус, верхневятский подъярус, мальцевская свита, кедровские слои.
Описание (рис. 1, а). Передний край переднего крыла выпуклый. Костальное поле у основания RS в 3.5 раза шире субкостального. SC с простыми и Y-образными передними ветвями, заканчивается в дистальной трети крыла. R с редкими передними ветвями, изогнут к переднему краю крыла до и после основания RS. Интеррадиальное поле резко расширяется в своем основании. Основание RS расположено в конце базальной четверти крыла, RS начинает ветвиться до дистальной трети крыла с двумя или более ветвями. М начинает ветвиться на некотором расстоянии от основания RS. МА начинает ветвиться вблизи основания, с двумя ветвями, МР простая. CuA разделяется на простые CuA1 и CuA2 непосредственно перед основанием RS. Вершины CuA1 и CuA2 не слиты, вершина CuA1 заканчивается в дистальной трети крыла. Интеркубитальное поле не расширено в основании. А1 простая, А2 с двумя ветвями, не образующими анастомозов. Поперечные жилки простые и образующие двойной ряд ячеек в интеррадиальном поле. Мембрана крыла темная, с небольшими светлыми пятнами.
Размеры в мм: длина переднего крыла 10.
Сравнение. По строению М новый вид наиболее сходен с Ch. issadensis Aristov, 2009 из верхнепермского местонахождения Исады в России, от которого отличается не слитыми вершинами ветвей CuA. У Ch. issadensis вершины CuA1 и CuA2 слиты (Aristov, 2013b).
Материал. Голотип.
СЕМЕЙСТВО GEINITZIIDAE HANDLIRSCH, 1906
Определительная таблица родов пермских Geinitziidae
1(2) CuA с четырьмя ветвями.................................. ...................................Geinitzia Handlirsch, 1906
2(1) CuA с тремя ветвями.
3(4) Вершина CuA2 изогнута параллельно заднему краю крыла...................Shurabia Martynov, 1937
4(3) Вершина CuA2 не изогнута параллельно заднему краю крыла.
5(6) RS гребенчатый назад.............Stegopterum Sharov, 1961
6(5) RS дихотомический или ветвящийся беспорядочно...................Permoshurabia Aristov, 2009
Род Geinitzia Handlirsch, 1906
Geinitzia annosa (Aristov, 2004) comb. nov.
Табл. VI , фиг. 2
Рис. 2.
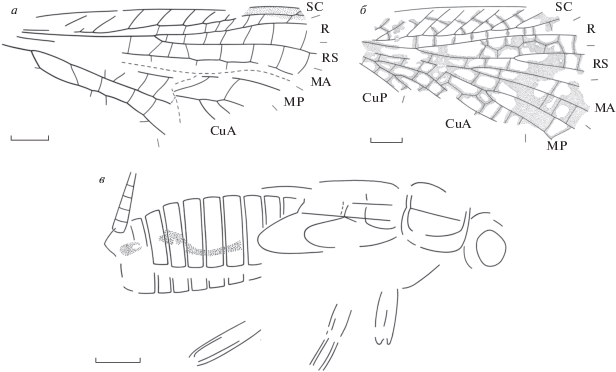
Представители отряда Reculida и Gryllones incertae ordinis из Бабьего Камня (Кемеровская обл.; верхняя пермь, вятский ярус): а – Khosara ultima sp. nov., голотип ПИН, № 4887/568 (семейство Liomopteridae), переднее крыло; б – Permotoma aba gen. et sp. nov., голотип ПИН, № 4887/193 (семейство Skaliciidae), переднее крыло; в – Tarakanikha satura gen. et sp. nov., голотип ПИН, № 4887/351 (Gryllones incertae ordinis), общий вид.
Shurabia annosa: Аристов, 2004, c. 51, рис. 2, б, 3, б.
Голотип – ПИН, № 4887/28, прямой и обратный отпечатки переднего крыла; местонахождение Бабий Камень; верхняя пермь, мальцевская свита, кедровские слои.
Описание (рис. 1, б). Передний край переднего крыла слабовыпуклый. Костальное поле у основания RS в 1.6 раза шире субкостального. SC с частыми простыми передними ветвями, заканчивается у середины крыла. Передние ветви R простые и частые. Четырехветвистый RS начинает ветвиться в базальной трети крыла, интеррадиальное поле расширено в середине. МА с коротким развилком у вершины крыла, МР начинает ветвиться в дистальной трети крыла, с двумя окончаниями. CuA1 начинает ветвиться в своей базальной трети, задняя ветвь CuA1 с коротким развилком. Вершина CuA2 изогнута параллельно заднему краю крыла. CuP дуговидно изогнута к переднему краю крыла. Окраска в виде пятен и полос вдоль поперечных жилок в задней половине крыла.
Размеры в мм: длина переднего крыла около 11.
Сравнение. От остальных представителей рода G. annosa отличается коротким развилком на задней ветви CuA1. У остальных гейниций задняя ветвь CuA1 ветвится у основания (Стороженко, 1998).
Замечание. При описании вид был ошибочно отнесен к роду Shurabia (Аристов, 2004). Для обсуждаемого вида характерна четырехветвистая CuA, типичная для рода Geinitzia, тогда как у Shurabia CuA имеет три ветви (см. определительную таблицу).
Материал. Голотип.
Род Stegopterum Sharov, 1961
Stegopterum kabalum Aristov, sp. nov.
Табл. VI , фиг. 3
Название вида от қабал шорск. – пепел.
Голотип – ПИН, № 4887/520, прямой и обратный отпечатки переднего крыла; местонахождение Бабий Камень; верхняя пермь, мальцевская свита, кедровские слои.
Описание (рис. 1, в). Передний край переднего крыла выпуклый. Костальное поле у основания RS в 2 раза шире субкостального. SC с частыми простыми передними ветвями, заканчивается у границы дистальной трети крыла. R с тремя передними ветвями. Основание RS расположено в базальной трети крыла, интеррадиальное поле расширено в середине. RS с пятью или более ветвями, гребенчатый назад, с дополнительным развилком на задней ветви. М с выраженной М5, разделяется на МА и МР примерно посередине между основанием RS и первым развилком CuA. МА и МР начинают ветвиться у середины крыла, с тремя и двумя ветвями соответственно. CuA1 разделяется на две простые ветви перед своей серединой, передняя ветвь заканчивается до дистальной трети крыла, CuA2 слабоизогнута у вершины. Поперечные жилки в основном простые, но в интеррадиальном поле образуют несколько ячеек.
Размеры в мм: длина переднего крыла около 16.
Сравнение. S. kabalum наиболее сходен с S. hirtum Sharov, 1961 из казанского местонахождения Калтан в Кемеровской обл. России. Новый вид отличается рано ветвящейся МА и короткой CuA. У S. hirtum МА простая, передняя ветвь CuA заканчивается в дистальной трети крыла (Аристов, 2004).
Материал. Голотип.
Stegopterum pashka Aristov, sp. nov.
Табл.VI, фиг. 4
Название вида от пашқа шорск. – иной (другой).
Голотип – ПИН, № 4887/533, прямой и обратный отпечатки переднего крыла; местонахождение Бабий Камень; верхняя пермь, мальцевская свита, кедровские слои.
Описание (рис. 1, г). Передний край переднего крыла выпуклый. Костальное поле у основания RS в 1.7 раза шире субкостального. В основании костального поля проходит тонкая выпуклая жилка, параллельная переднему краю крыла. SC с частыми простыми передними ветвями, заканчивается перед дистальной четвертью крыла. R с четырьмя или более передними ветвями. Основание RS расположено у границы базальной трети крыла, интеррадиальное поле расширено в середине. RS гребенчатый назад, с четырьмя или более ветвями. М5 редуцирована до тонкой поперечной жилки. Первые развилки М и CuA сближены, М разделяется на МА и МР на заметном расстоянии перед основанием RS. МА и МР начинают ветвиться за серединой крыла, МА, вероятно, с четырьмя ветвями, МР двуветвистая. CuA1 разделяется на две простые ветви у своей середины, передняя ветвь заканчивается до дистальной трети крыла, CuA2, вероятно, изогнута у вершины параллельно заднему краю крыла. Поперечные жилки в основном простые, H-образные в интеррадиальном и Y-образные в интермедиальном полях.
Размеры в мм: длина переднего крыла около 13.
Сравнение. Новый вид наиболее сходен с S. kabalum, от которого отличается более длинной SC, редукцией М5 и смещением первого развилка М к первому развилку CuA. У S. kabalum SC заканчивается перед дистальной третью крыла, М5 четко выражена, первые развилки М и CuA не сближены (см. выше).
Замечание. Жилка в основании костального поля у S. pashka, вероятно, является “ложной костой”, характерной для прямокрылообразных насекомых. У рекулид этот признак крайне редок, но не уникален. Такое же строение основания костального поля известно для Parachauliodites orthopteroides Aristov, 2013 из северодвинского местонахождения Исады в Вологодской обл. России (семейство Chaulioditidae: Аристов, 2013).
Материал. Голотип.
Род Permoshurabia Aristov, 2009
Permoshurabia tshasha Aristov, sp. nov.
Табл. VI , фиг. 5, 6
Название вида от чаш шорск. – молодой.
Голотип – ПИН, № 4887/529, прямой и обратный отпечатки переднего крыла; местонахождение Бабий Камень; верхняя пермь, мальцевская свита, кедровские слои.
Описание (рис. 1, д–ж). Передний край переднего крыла выпуклый. Костальное поле у основания RS примерно в 2 раза шире субкостального. SC с частыми простыми, реже дихотомирующими передними ветвями, заканчивается у середины крыла или перед его дистальной третью. R с тремя–шестью передними ветвями. Основание RS расположено на границе базальной трети крыла, интеррадиальное поле не расширено. RS с пятью–семью основными ветвями, у середины крыла или на границе его дистальной трети разделяется на две ветви – гребенчатую вперед переднюю и простую или дихотомирующую заднюю. М5 не выражена, М разделяется на МА и МР примерно посередине между основанием RS и первым развилком CuA. МА с тремя–четырьмя, МР с двумя–тремя ветвями. CuA1 делится на две простые ветви у своей середины или дистальнее, вершина CuA2 слабо S-образно изогнута. Поперечные жилки простые. Окраска (сохранилась только на паратипе № 4887/519) в виде темной полосы вдоль переднего края крыла и полосок вдоль поперечных жилок.
Размеры в мм: длина переднего крыла около 16.
Сравнение. От остальных представителей рода отличается гребенчатой вперед передней ветвью RS. У P. mesenensis Aristov, 2009 из Сояны (казанский ярус Архангельской обл. России) и P. komi Aristov, 2015 из Воркуты (кунгурский ярус Республики Коми, Россия) RS дихотомический, четырехветвистый (Аристов, 2009; Aristov, 2015b).
Материал. Голотип и паратипы ПИН, №№ 4887/519 и 555 из того же местонахождения.
Род Permuliercula Aristov, gen. nov.
Название рода от пермского периода и muliercula лат. – самка; ж. р.
Типовой вид – P. chimaera sp. nov.
Диагноз. Мелкие насекомые. В переднем крыле костальное поле у основания RS немного шире субкостального. SC заканчивается у середины крыла. RS дихотомический, начинается в конце базальной трети крыла, с первым развилком перед серединой крыла, задняя ветвь RS слита с МА. Интеррадиальное поле расширенное, пересечено длинной, мощной, S-образно изогнутой поперечной жилкой. М5 впадает в CuA после ее разделения на ветви, М разделяется на ветви на равном расстоянии как от первого развилка CuA, так и от основания RS. CuA1 двуветвистая, вершина CuA2 изогнута S-образно.
Видовой состав. Типовой вид.
Сравнение. Новый род наиболее сходен с родом Shurabia Martynov, 1937, от которого, как и от остальных родов гейнициид Permuliercula gen. nov. отличается М5, впадающей в CuA1. У остальных Geinitziidae, как и у всех Reculida, М5 впадает в CuA до ее разделения на ветви (Стороженко, 1998; Аристов и др., 2009; Aristov, 2015a, b).
Замечание. Впадение М5 в CuA после ее разделения на ветви является одной из важнейших синапоморфий надотряда Orthopteroidea (Горохов, 2004). Вне этого надотряда такой анастомоз у гриллоновых характерен только для описываемого рода и некоторых семейств кнемидолестидовых подотряда Parmapterina (Aristov, 2016).
Permuliercula chimaera Aristov, sp. nov.
Табл. VI , фиг. 7
Название вида от Chimaera греч. – Химера. В палеонтологии – никогда не существовавший организм, ошибочно реконструированный на основе остатков организмов различных видов.
Голотип – ПИН, № 4887/222, прямой и обратный отпечатки переднего крыла; местонахождение Бабий Камень; верхняя пермь, мальцевская свита, кедровские слои.
Описание (рис. 1, з). Передний край переднего крыла выпуклый. SC с простыми, слабо S‑образно изогнутыми передними ветвями. RS с четырьмя–пятью ветвями. МА с четырьмя ветвями, МР двуветвистая. Задняя ветвь CuA1 изогнута S‑образно. Интеркубитальное поле плавно сужается к заднему краю крыла, CuP прямая.
Размеры в мм: длина переднего крыла 10.
Материал. Голотип.
СЕМЕЙСТВО LIOMOPTERIDAE SELLARDS, 1909
Род Khosara Martynov, 1937
Khosara ultima Aristov, sp. nov.
Табл. VI , фиг. 8
Название вида ultima лат. – последняя.
Голотип – ПИН, № 4887/568, прямой и обратный отпечатки переднего крыла; местонахождение Бабий Камень; верхняя пермь, мальцевская свита, кедровские слои.
Описание (рис. 2, а). Передний край переднего крыла слабовыпуклый. Костальное поле у основания RS в 2 раза шире субкостального. SC с простыми передними ветвями, заканчивается в дистальной четверти крыла. RS начинается сразу за границей базальной трети крыла. RS, МА и МР до дистальной трети крыла простые. CuA1 прямая, начинает ветвиться у своей вершины, образуя задний гребень из трех или более ветвей. CuA2 простая, S-образно изогнутая. Ствол МА с изломами в местах впадения поперечных жилок. Окраска в виде пятна в области птеростигмы.
Размеры в мм: длина переднего крыла около 12.
Сравнение. Новый вид наиболее сходен с K. permiakovae Martynov, 1937 из уржумского местонахождения Каргала в Оренбургской обл. России. Новый вид отличается простой МА в середине крыла и гребенчатой CuA1. У K. permiakovae МА ветвится у середины крыла, CuA1 с двумя ветвями (Стороженко, 1998).
Материал. Голотип.
СЕМЕЙСТВО SKALICIIDAE KUKALOVÁ, 1964
Род Permotoma Aristov, gen. nov.
Название рода от пермского периода и р. Томь; ж. р.
Типовой вид – T. aba sp. nov.
Диагноз. Мелкие насекомые. В переднем крыле костальное поле у основания RS в 2 раза шире субкостального. SC заканчивается у начала дистальной трети крыла. RS отделяется в базальной четверти крыла и начинает ветвиться за его серединой. M разделяется на МА и МР за основанием RS. МА и МР начинают ветвиться у середины крыла. CuA1 простая, изогнута S-образно, CuA2 ветвится у своего основания.
Видовой состав. Типовой вид.
Сравнение. Новый род наиболее сходен с Afrogrylloblattus Aristov et Mostovsky, 2013 из верхнепермского местонахождения Лиджеттон в ЮАР наличием М, разделяющейся на МА и МР за основанием RS. Род Permotoma отличается узким костальным полем, простой CuA1 и рано ветвящейся CuA2. У Afrogrylloblattus костальное поле у основания RS примерно равно по ширине субкостальному, CuA1 ветвится, CuA2 начинает ветвиться у своей середины (Аристов, Мостовский, 2013).
Permotoma aba Aristov, sp. nov.
Табл. VI , фиг. 9
Название вида от абинской серии.
Голотип – ПИН, № 4887/193, прямой и обратный отпечатки переднего крыла; местонахождение Бабий Камень; верхняя пермь, мальцевская свита, кедровские слои.
Описание (рис. 2, б). Передний край переднего крыла слабовыпуклый. SC с простыми передними ветвями, соединенными поперечными жилками. Интеррадиальное поле широкое, но уже субкостального. RS и МА с двумя или более ветвями, МР двуветвистая. CuA2 с двумя или более ветвями. Основание интеркубитального поля не расширено. Поперечные жилки простые и образующие двойной ряд ячеек в интеррадиальном поле и посередине медиального поля. Окраска вдоль поперечных жилок и в виде небольших пятен у основания крыла, первого развилка М и крупного пятна в дистальной половине крыла.
Размеры в мм: длина переднего крыла около 12.
Материал. Кроме голотипа – экз. ПИН, № 4887/236 из того же местонахождения.
GRYLLONES INCERTAЕ ORDINIS
GRYLLONES INCERTAЕ FAMILIAЕ
Род Tarakanikha Aristov, gen. nov.
Название рода от тараканихинских слоев; ж. р.
Типовой вид – T. satura sp. nov.
Диагноз. Голова крупная, с крупными глазами. Пронотум вытянутый, сужающийся к своему переднему краю. Паранотальное кольцо квадратное, не закрывающее голову. Боковые лопасти параноталий сужаются к заднему краю пронотума. Мезонотум квадратный, короче пронотума, метанотум короче мезонотума. Передние ноги длинные, с широкими невооруженными голенями. Средние и задние голени длинные, направлены назад. Передние и задние крыловые зачатки с закругленными вершинами, расположены параллельно оси тела. Тергиты брюшка узкие, церки короткие, с короткими члениками, резко сужающиеся к вершине.
Видовой состав. Типовой вид.
Сравнение. Новый род наиболее сходен с Tshekardites Aristov, 2004 из нижнепермского местонахождения Чекарда (кунгурский ярус Пермского края, Россия). Описываемый род отличается крупными глазами, сужающимся к переднему краю пронотумом и толстыми голенями. У Tshekardites глаза небольшие, пронотум квадратный, голени стройные (Aristov, 2004; Аристов и др., 2006). От еще одного сходного рода Kaltanympha из местонахождения Калтан (казанский ярус Кемеровской обл., Россия) Tarakanikha отличается узким пронотумом и короткими церками со сравнительно длинными члениками. У Kaltanympha пронотум квадратный, церки длинные с очень короткими члениками (Стороженко, 1998).
Замечания. Общий план строения (наличие не закрывающих голову параноталий, параллельные передние и задние крыловые зачатки, не увеличенные задние ноги с направленными назад голенями и членистые церки) описываемой нимфы характерен для двух отрядов гриллоновых – Eoblattida и Reculida (состав этих отрядов см.: Aristov, 2015a). Такие признаки, как крупные голова и глаза, расширяющиеся к голове боковые лопасти параноталий и короткие церки характерны для эоблаттидовых семейства Megakhosaridae. Наиболее сходно с описываемой нимфой имаго мегахозариды Tshekhosara из нижней перми России (Novokshonov, 1998). Однако это сходство не представляется нам достаточно надежным для отнесения новой нимфы к эоблаттидовым, тем более к какому-либо их семейству.
На фоне брюшка сохранилось неструктурированное органическое содержимое кишечника, вероятно, растительного происхождения. Судя по крупным глазам, направленным назад голеням, характерным для стратобионтов (водным нимфам, наоборот, свойственны голени, направленные вперед: Аристов и др., 2006; Vršanský et al., 2019), длинным ногам и содержимому кишечника обсуждаемый вид был сапрофагом, ведущим более или менее открытый образ жизни.
Tarakanikha satura Aristov, sp. nov.
Табл. VI , фиг. 10
Название вида satura лат. – сытая.
Голотип – ПИН, № 4887/351, прямой и обратный отпечатки нимфы; местонахождение Бабий Камень; верхняя пермь, мальцевская свита, тараканихинские слои.
Описание (рис. 2, в). Длина пронотума в 1.3 раза больше его максимальной ширины. Передний и задний края паранотального кольца узкие. Задние крыловые зачатки заметно шире передних. Церки короткие, пяти- или шестичлениковые.
Размеры в мм: длина тела 10.
Материал. Голотип.
Список литературы
Аристов Д.С. Ревизия семейства Tomiidae // Палеонтол. журн. 2003. № 1. С. 32–39.
Аристов Д.С. Фауна гриллоблаттидовых насекомых (Grylloblattida) конца поздней перми и первой половины триаса // Палеонтол. журн. 2004. № 5. С. 45–52.
Аристов Д.С. Обзор стратиграфического распространения пермских Grylloblattida (Insecta) с описанием новых таксонов // Палеонтол. журн. 2009. № 6. С. 37–45.
Аристов Д.С. Новые и малоизвестные Grylloblattida (Insecta) из межтрапповых отложений Тунгусского бассейна Сибири // Палеонтол. журн. 2011. № 5. С. 61–68.
Аристов Д.С. Новые Grylloblattida (Insecta) из верхней перми и нижнего триаса Европейской России и верхней перми Казахстана // Палеонтол. журн. 2013. № 1. С. 48–56.
Аристов Д.С. Новые гриллоновые насекомые (Insecta: Gryllones) из местонахождения Бабий Камень (верхняя пермь России). Часть 1. Отряд Eoblattida // Палеонтол. журн. 2020. № 1. С. 44–49.
Аристов Д.С., Вапплер Т., Расницын А.П. Новые и малоизвестные Geinitziidae (Insecta; Grylloblattida) из триаса и юры Европы, Средней Азии, Монголии и Южной Африки // Палеонтол. журн. 2009. № 4. С. 59–65.
Аристов Д.С., Гравожель-Стамм Л., Маршал-Папье Ф. Новые гриллоблаттиды (Insecta; Grylloblattida) из вольциевого песчаника Вогез (средний триас Франции) // Палеонтол. журн. 2011. № 2. С. 39–45.
Аристов Д.С., Мостовский М.Б. Гриллоблаттиды (Insecta; Grylloblattida) из верхней перми Южной Африки // Палеонтол. журн. 2013. № 5. С. 37–43.
Аристов Д.С., Новокшонов В.Г., Паньков Н.Н. Систематика нимф гриллоблаттидовых насекомых (Insecta; Grylloblattida) // Палеонтол. журн. 2006. № 1. С. 75–85.
Горохов А.В. Примитивные Titanoptera и ранняя эволюция Polyneoptera // Чтения памяти Н.А. Холодковского. Вып. 57. № 1. СПб.: ЗИН РАН, 2004. С. 1–54.
Горохов А.В. Обзор триасовых прямокрылых (Orthoptera) с описанием новых и малоизвестных таксонов. Сообщение 2 // Палеонтол. журн. 2005. № 3. С. 46–54.
Пономаренко А.Г. Насекомые во временных окрестностях пермо-триасового кризиса // Палеонтол. журн. 2016. № 2. С. 68–81.
Расницын А.П., Аристов Д.С., Расницын Д.А. Насекомые у рубежа перми и раннего триаса (уржумский–оленекский века) и проблема пермотриасового кризиса биоразнообразия // Журн. общ. биол. 2013. Т. 74. № 1. С. 30–52.
Садовников Г.Н. Трапповый вулканизм Сибири и «пермо–триасовое вымирание» // Геол. и разведка. 2016. № 2. С. 8–14.
Стороженко С.Ю. Систематика, филогения и эволюция гриллоблаттидовых насекомых (Insecta: Grylloblattida). Владивосток: Дальнаука, 1998. 207 с.
Щербаков Д.Е. О пермских и триасовых энтомофаунах в связи с биогеографией и пермо-триасовым кризисом // Палеонтол. журн. 2008. № 1. С. 15–33.
Щербаков Д.Е., Башкуев А.С., Василенко Д.В. и др. Новое местонахождение раннетриасовых насекомых – Петропавловка // Палеострат-2019. Годичн. собрание (науч. конф.) секции палеонтол. МОИП и Моск. отделения Палеонтол. об-ва при РАН. Москва, 28–30 января 2019 г. М.: ПИН РАН, 2019. С. 68–69.
Aristov D.S. The fauna of grylloblattid insects (Grylloblattida) of the Lower Permian locality of Tshekarda // Paleontol. J. 2004. V. 38. Suppl. 2. P. 80–145.
Aristov D.S. 1.3. Vyatkian–Olenekian localities // Fossil Insects of the Middle and Upper Permian of European Russia / Ed. Aristov D.S. N.Y., 2013a. P. 669–675 (Paleontol. J. V. 47. № 7).
Aristov D.S. 2.6. New grylloblattids (Insecta: Grylloblattida) from the Upper Permian of the Vologda region // Fossil Insects of the Middle and Upper Permian of European Russia / Ed. Aristov D.S. N.Y., 2013b. P. 751–766 (Paleontol. J. V. 47. № 7).
Aristov D.S. Classification of order Eoblattida (Insecta; Blattidea) with description of new taxa // Far Eastern Entomol. 2015a. № 301. P. 1–56.
Aristov D.S. A new gryllones insects (Insecta: Gryllones) from the Permian of the Russia // Paleontol. J. 2015b. V. 49. № 12. P. 1310–1333.
Aristov D.S. Classification of the Parmapterina subord. n. (Insecta: Cnemidolestida) // Far Eastern Entomol. 2016. № 311. P. 13–22.
Aristov D.S., Bashkuev A.S., Vassilenko D.V. Review of the entomofauna // Fossil Insects of the Middle and Upper Permian of European Russia / Ed. Aristov D.S. N.Y., 2013. P. 658–667 (Paleontol. J. V. 47. № 7).
Bashkuev A., Sell J., Aristov D. et al. Insects from the Buntsandstein of Lower Franconia and Thuringia // Paläontol. Z. 2012. Bd 86. P. 175–185.
Novokshonov V.G. Tshekhosara improvida n. gen., n. sp. (Insecta; Grylloblattida: Megakhosaridae) aus dem unteren Perm des Urals // Paläontol. Z. 1998. Bd 72. S. 65–66.
Rasnitsyn A.P., Aristov D.S., Rasnitsyn D.A. Insects of the Permian and Early Triassic (Urzhumian–Olenekian ages) and the problem of the Permian–Triassic biodiversity crisis // Fossil Insects of the Middle and Upper Permian of European Russia / Ed. Aristov D.S. N.Y., 2013. P. 793–823 (Paleontol. J. V. 47. № 7).
Shcherbakov D.E. Permian and Triassic ancestors of webspinners (Embiodea) // Russ. Entomol. J. 2015. V. 24. № 3. P. 187–200.
Storozhenko S.Yu., Aristov D.S. Review of the Paleozoic and Mesozoic families Megakhosaridae and Blattogryllidae (Insecta: Grylloblattida) // Far Eastern Entomol. 2014. № 271. P. 1–28.
Vršanský P., Sendi H., Aristov D. et al. Ancient roaches further exemplify ‘no land return’ in aquatic insects // Gondwana Res. 2019. V. 68. P. 22–33. https://doi.org/10.1016/j.gr.2018.10.020
Yan E.V., Beutel R.G., Lawrence J.F. et al. Archaeomalthus – (Coleoptera, Archostemata) a ‘ghost adult’ of Micromalthidae from Upper Permian deposits of Siberia? // Histor. Biol. 2019. https://doi.org/10.1080/08912963.2018.1561672
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал