

Палеонтологический журнал, 2020, № 4, стр. 54-63
Новые ручейники (Insecta: Trichoptera) семейств Vitimotauliidae и Philopotamidae из местонахождения Хасурты (нижний мел России), с кратким обзором ископаемой мировой фауны
И. Д. Сукачева a, Д. С. Аристов a, b, *
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Череповецкий государственный университет
Череповец, Россия
* E-mail: danil_aristov@mail.ru
Поступила в редакцию 23.07.2019
После доработки 17.09.2019
Принята к публикации 28.10.2019
Аннотация
Описаны новые представители семейства Vitimotauliidae: Multimodus amplus sp. nov., Purbimodus parvulus sp. nov., Burimodus novus gen. et sp. nov., и новый вид семейства Philopotamidae Juraphilopotamus callidus sp. nov. из нижнего мела России (Республика Бурятия, местонахождение Хасурты). Приведены определительные таблицы родов семейства Vitimotauliidae и видов родов Purbimodus и Juraphilopotamus. Дается геологическое и географическое распространение ископаемых форм из семейств Vitimotauliidae и Philopotamidae. Анализ стратиграфического распространения ручейников указывает на возраст отложений Хасурты не моложе начала раннего мела.
Новые представители семейств Vitimotauliidae и Philopotamidae, описываемые в настоящей работе, собраны экспедиционными отрядами ПИН РАН 2004, 2009 и 2014 гг. из нижнемелового местонахождения Хасурты (Россия, Республика Бурятия, Закаменский р-н, окрестности хут. Хасурты, в 25 км восточнее г. Закаменска). Обзор фауны насекомых из этого местонахождения был опубликован ранее (Сукачева, Василенко, 2019). Отложения относятся к гусиноозерской серии и датируются ранним мелом (Копылов, Расницын, 2017).
Материал, описанный в настоящей работе, хранится в колл. лаборатории артропод Палеонтологического ин-та им. А.А. Борисяка (ПИН РАН).
Авторы выражают глубокую благодарность А.П. Расницыну, А.Г. Пономаренко (ПИН РАН), В.Д. Иванову и С.И. Мельницкому (СПбГУ) за ценные советы и консультации, а также Н.С. Гороховой (ООО “Издательство АСТ”) за техническую помощь. Работа поддержана Программой Президиума РАН “Эволюция органического мира. Роль и влияние планетарных процессов” и грантом РФФИ № 18-04-00322.
ОТРЯД TRICHOPTERA
СЕМЕЙСТВО VITIMOTAULIIDAE SUKATSHEVA, 1968
Типовой род – Vitimotaulius Sukatsheva, 1968.
Диагноз. Vitimotauliidae – в основном, крупные насекомые (длина крыльев от 6 до 15 мм). Передние крылья удлиненные, с более или менее выступающими вершиной и торнусом. Прямая Sc и R со слабым вершинным изгибом впадают в передний край крыла дистальнее середины длины крыла. Ствол RS довольно короткий. RS ветвится проксимальнее ветвления M. Ячеи DC, MC и TC всегда закрыты. Поперечные жилки прямые или слегка вогнутые с дистальной стороны. Имеются все пять апикальных развилков. Ветви развилков F1 и F2 параллельны. Субкостальное поле часто уже костального. CuA разветвляется раньше или на одном уровне с развилком F1. CuP и A1 впадают в задний край крыла в одной точке или близко друг от друга, дистальнее или на уровне середины длины крыла. Ячея TC средней длины, чаще длинная, длиннее ячей DC и MC. A2 различной длины. Иногда на крыльях имеется темный рисунок.
В задних крыльях развилок F4 отсутствует. Поперечные жилки rs2–rs3, rs4–m1+2 и m3+4–cua имеются. Антенны тонкие, небольшие, немного короче или равны передним крыльям, основания антенн утолщены. На передних ногах имеются две апикальные шпоры. Все тело покрыто густыми волосками (Wang et al., 2009a).
Состав. Семейство насчитывает пять родов с 27 видами.
Замечания. Семейство Vitimotauliidae известно, главным образом, из нижнего мела Евразии (Юго-Восточная Сибирь, Забайкалье, Монголия, Китай). Кроме того, оно найдено в нижнем мелу Южной Англии (табл. 1).
Таблица 1.
Стратиграфическое и географическое распространение представителей cемейства Vitimotauliidae
| Мел | Верхний | Сеноман | Бурея, Россия | Multimodus bureensis Sukatsheva, 1982 | Сукачева, 1982 |
| Нижний | Апт | Бон-Цаган, Монголия | Multimodus parcus Sukatsheva, 1992 | Сукачева, 1992 | |
| Байса, Россия | Multimodus incompletus Sukatsheva, 1968 | Сукачева, 1968 | |||
| Байса, Россия | Multimodus insperatus Sukatsheva, 1968 | Сукачева, 1968 | |||
| Байса, Россия | Multimodus longirameus Sukatsheva, 1968 | Сукачева, 1968 | |||
| Байса, Россия | Multimodus maculatus Sukatsheva, 1968 | Сукачева, 1968 | |||
| Байса, Россия | Multimodus martynovae Sukatsheva, 1968 | Сукачева, 1968 | |||
| Байса, Россия | Multimodus obscurus Sukatsheva, 1968 | Сукачева, 1968 | |||
| Байса, Россия | Multimodus pedunculatus Sukatsheva, 1968 | Сукачева, 1968 | |||
| Байса, Россия | Multimodus picturatus Sukatsheva, 1968 | Сукачева, 1968 | |||
| Байса, Россия | Vitimotaulius legibilis Sukatsheva, 1968 | Сукачева, 1968 | |||
| Шар-Тологой, Монголия | Multimodus sp. | ориг. | |||
| Исянь, Китай | Multimodus dissithus Ren, 1995 | Ren, 1995 | |||
| Исянь, Китай | Multimodus stigmaeus Ren, 1995 | Ren, 1995 | |||
| Исянь, Китай | Parachorista (=Multimodus) miris Lin, 1976 | Lin, 1976 | |||
| Исянь, Китай | Sinomodus macilentus Wang et al., 2009 | Wang et al., 2009a | |||
| Исянь, Китай | Sinomodus peltatus Wang et al., 2009 | Wang et al., 2009a | |||
| Исянь, Китай | Sinomodus spatiosus Wang et al., 2009 | Wang et al., 2009a | |||
| Исянь, Китай | Vitimotauliidae? gen.? sp.? | Davis et al., 2010 | |||
| Ундурга, Россия | Vitimotaulius magnus Sukatsheva, 1990 | Сукачева, 1990 | |||
| Семен, Россия | gen. sp. | ориг. | |||
| Романовка, Россия | gen. sp. | ориг. | |||
| Баррем- готтерив | Гурван-Эрэн, Монголия | gen. sp. | ориг. | ||
| Манлай, Монголия | gen. sp. | ориг. | |||
| Берриас | Пурбек, Англия | Purbimodus minor Sukatsheva et Jarzembowski, 2001 | Sukatsheva, Jarzembowski, 2001 | ||
| Пурбек, Англия | Purbimodus medius Sukatsheva et Jarzembowski, 2001 | Sukatsheva, Jarzembowski, 2001 | |||
| Пурбек, Англия | Purbimodus rasnitsyni Sukatsheva et Jarzembowski, 2001 | Sukatsheva, Jarzembowski, 2001 | |||
| Пурбек, Англия | Purbimodus saxosus Sukatsheva et Jarzembowski, 2001 | Sukatsheva, Jarzembowski, 2001 | |||
| Н. мел | Хасурты, Россия | Burimodus novus Sukatsheva et Aristov, sp. nov. | ориг. | ||
| Хасурты, Россия | Multimodus amplus Sukatsheva et Aristov, sp. nov. | ориг. | |||
| Хасурты, Россия | Purbimodus parvulus Sukatsheva et Aristov, sp. nov. | ориг. | |||
| Юра | Верхняя | В. юра | Бичектуй, Россия | Multimodus perillustris Sukatsheva, 1990 | Сукачева, 1990 |
| Унда, Россия | Vitimotaulius secundus Sukatsheva, 1990 | Сукачева, 1990 | |||
| Змеиный, Россия | gen. sp. | ориг. | |||
| Савина, Россия | gen. sp. | ориг. | |||
| Хоотын-Хотгор, Монголия | gen. sp. | ориг. |
Первоначально семейство было описано только по передним крыльям (Сукачева, 1968). Однако в 2009 г. из местонахождения Исянь (Китай) был описан новый род Sinomodus Wang, Liang, Ren et Shin, 2009 с тремя видами (Wang et al., 2009a). Благодаря хорошей сохранности отпечатков из Китая, в описание были добавлены морфология головы и груди, которые мы приводим в нашей работе.
Определительная таблица родов семейства Vitimotauliidae
1(2) Развилок F5 расположен на одном уровне с развилком F1....................................................... ...........Sinomodus Wang, Liang, Ren et Shin, 2009
2(1) Развилок F5 расположен проксимальнее развилка F1
3(8) Ячея MC заметно короче ячеи DC
4(5) Передние крылья не более 10 мм. CuP и A1 впадают в задний край крыла на середине или вблизи середины длины крыла.......................... .....Purbimodus Sukatsheva et Jarzembowski, 2001
5(4) Передние крылья более 10 мм
6(7) Субкостальное поле равной ширины с костальным. Ячея TC короткая (в 1.3 раза короче ячеи MC)............................................................ ..............Burimodus Sukatsheva et Aristov, sp. nov.
7(6) Субкостальное поле уже костального. Ячея TC длинная (в 1.5 раза длиннее ячеи MC)......... ...............................Multimodus Sukatsheva, 1968
8(3) Ячея MC длиннее или почти равна ячее DC... .............................Vitimotaulius Sukatsheva, 1968
Род Multimodus Sukatsheva, 1968
Multimodus: Сукачева, 1968, с. 63; 1982, с. 90; 1990, с. 98; 1992, с. 116; Ren, 1995, с. 95–97.
Типовой вид – M. martynovae Sukatsheva, 1968.
Диагноз. RS примерно в полтора раза короче ячеи DC, отходит от R в базальной четверти крыла. RS1+2 и RS3+4 (как и M1+2 и M3+4) разветвляются в конце или на середине третьей четверти крыла. Ячея MC короткая, составляет примерно 2/3 длины ячеи DC. Ствол M длинный, длиннее ячеи MC. CuP и A1 оканчиваются дистальнее или немного проксимальнее середины длины крыла, в одной точке или близко друг от друга. Субкостальное поле уже, чем костальное. Часто на крыле имеется темный рисунок. Размеры передних крыльев 10 мм и более.
Видовой состав. 15 видов (см. табл. 1).
Сравнение. См. определительную таблицу родов Vitimotauliidae.
Multimodus amplus Sukatsheva et Aristov, sp. nov.
Табл. VI , фиг. 1 (см. вклейку)
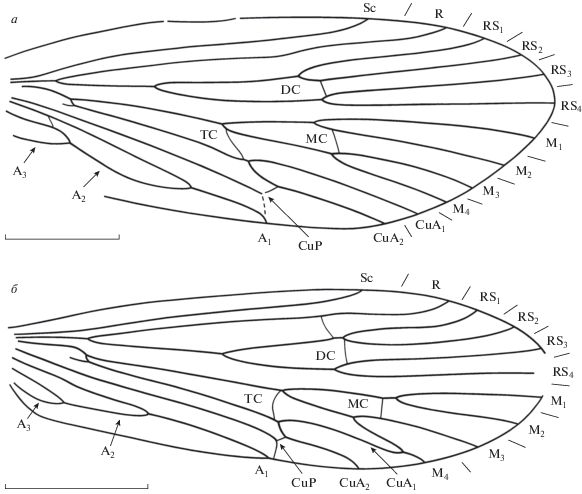
Рис. 1.
Представители семейства Vitimotauliidae из местонахождения Хасурты, передние крылья: а – Multimodus amplus sp. nov., голотип ПИН, № 5026/579; б – Purbimodus parvulus sp. nov., голотип ПИН, № 5026/1498. Длина масштабной линейки на а – 3 мм, на б – 2 мм.
Название вида amplus лат. – крупный.
Голотип – ПИН, №5026/579, прямой и обратный отпечатки полного переднего крыла; Россия, Республика Бурятия, Закаменский р-н, окрестности хут. Хасурты, в 25 км восточнее г. Закаменска, местонахождение Хасурты; нижний мел, гусиноозерская серия.
Описание (рис. 1, а). Крыло довольно широкое, длина больше ширины в 2.6 раза. Передний край крыла слабовыпуклый, вершина закруглена и расположена напротив окончания RS4. Торнус обозначен слабо. Наибольшая ширина крыла на уровне вершины развилка F4. Sc длинная, прямая, заканчивается чуть дистальнее середины третьей четверти крыла на уровне окончания CuA2. R длинный, со слабым изгибом на уровне начала F1 и слабым загибом у вершины. Субкостальное поле в два раза уже костального. Имеется поперечная жилка r–rs1. Ячея DC в 1.5 раза длиннее ячеи MC и равна трети длины крыла. F1 начинается заметно проксимальнее развилка F2. Поперечная rs2–rs3, закрывающая ячею DC, прямая. F3 длинный, начинается несколько проксимальнее F4. Поперечная m2–m3 слабо косая. Ячея TC длинная. Поперечная жилка cua2–cup очень короткая. Окончание CuP десклеротизовано. Анальная петля длинная. A1 впадает в задний край крыла немного проксимальнее середины его длины. A2 очень длинная, в 2.4 раза длиннее A3.
Размеры в мм: длина крыла 15.0, ширина крыла 5.0.
Сравнение. Описываемый вид наиболее сходен с M. pedunculatus Sukatsheva, 1968 из неокома Забайкалья (местонахождение Байса) по расположению вершин апикальных развилков и длине A2. Отличается большими размерами и несколько изогнутым R. У остальных видов рода R прямой, кроме M. bureensis Sukatsheva, 1982 из сеномана Хабаровского края (местонахождение Бурея) и M. stigmaeus Ren, 1995 из апта Китая (местонахождение Исянь) (табл. 1). У последних также имеется небольшой изгиб на R.
Материал. Голотип.
Род Purbimodus Sukatsheva et Jarzembowski, 2001
Purbimodus: Sukatsheva, Jarzembowski, 2001, c. 688.
Типовой вид – P. minor Sukatsheva et Jar-zembowski, 2001.
Диагноз. RS короткий, отходит от R в базальной части крыла. RS1+2 и RS3+4 разветвляются в начале третьей четверти крыла. Костальное поле широкое, субкостальное поле узкое. Ячея DC длиннее ячеи MC. CuP и A1 впадают в задний край крыла в районе середины длины крыла в одной точке или довольно далеко друг от друга.
Размеры в мм: 6.0–10.
Состав. Пять видов (табл. 1).
Сравнение. См. определительную таблицу родов Vitimotauliidae.
Определительная таблица видов рода Purbimodus Sukatsheva et Jarzembowski, 2001
1(4) CuP и A1 впадают в задний край крыла в разных точках
2(3) RS в 1.2 раза короче ячеи DC. Ячея DC немного длиннее ячеи MC..................................... ......P. rasnitsyni Sukatsheva et Jarzembowski, 2001
3(2) RS более чем на половину короче ячеи DC. Ячея DC равна по длине ячее MC...................... ..........P. medius Sukatsheva et Jarzembowski, 2001
4(1) CuP и A1 впадают в задний край крыла в одной точке
5(8) Ячея DC длиннее ячеи MC. Sc впадает в передний край крыла чуть дистальнее вершин развилков F1 и F2
6(7) Поперечная жилка cua2–cup длинная. CuA1 прямая, не соединяется с M3+4........................... .........P. saxosus Sukatsheva et Jarzembowski, 2001
7(6) Поперечная жилка сua2–cup очень короткая. CuA1 иногда искривлена и соединяется с M3+4, не доходя до края крыла..................................... ..............P. parvulus Sukatsheva et Aristov, sp. nov.
8(5) Ячея DC почти равна по длине или чуть больше ячеи MC. Sc впадает в передний край крыла значительно дистальнее вершин развилков F1 и F2... ...........P. minor Sukatsheva et Jarzembowski, 2001
Purbimodus parvulus Sukatsheva et Aristov, sp. nov.
Табл. VI , фиг. 2
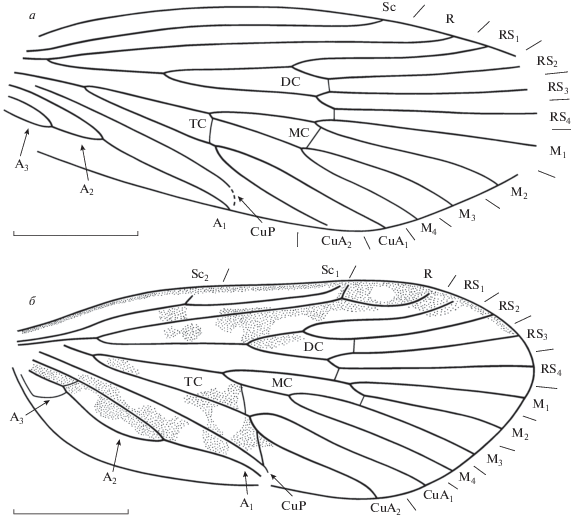
Рис. 2.
Представители семейств Vitimotauliidae и Philopotamidae из местонахождения Хасурты, передние крылья: а – Burimodus novus gen. et sp. nov., голотип ПИН, № 5340/1456; б – Juraphilopotamus callidus sp. nov., голотип ПИН, № 5026/595. Длина масштабной линейки – 3 мм.
Название вида parvulus лат. – мелкий.
Голотип – ПИН, № 5026/1498, прямой и обратный отпечатки полного переднего крыла; Республика Бурятия, местонахождение Хасурты; нижний мел; гусиноозерская серия.
Описание (рис. 1, б). Крыло довольно узкое, длина больше ширины в 2.7 раза. Передний край крыла прямой, вершина закруглена. Наибольшая ширина крыла на уровне впадения CuA2 в задний край крыла. Sc средней длины, прямая, заканчивается на уровне середины третьей четверти крыла, чуть дистальнее вершин развилков F1 и F2. Субкостальное поле узкое, в два раза уже костального. R со слабым длинным вершинным ковшеобразным изгибом. Имеется слабо косая поперечная жилка r–rs1+2, расположенная немного проксимальнее вершины F1. Ячея DC почти равна по длине своему стволу и в 1.3 раза длиннее ячеи MC. F1 начинается чуть дистальнее развилка F2. Поперечная rs1+2–rs3 слабо изогнута. F3 короткий, стебельчатый, начинается значительно дистальнее F4. Поперечная m1+2–m3 прямая. Ячея TC длинная. Ячеи DC, MC и TC закрыты. Поперечная m–cua дугообразно изогнута. Основание CuA частично редуцированное. Развилок F4 тоже короткий, почти равен по длине F3, широкий. CuA1 сливается с M4, не доходя до края крыла. CuP и A1 впадают в задний край крыла почти в одной точке на середине длины крыла. A2 довольно короткая, составляет 1.8 длины A1. Поперечная cua2–cup короткая. CuP резко загибается к точке впадения в задний край. Анальное поле узкое.
Размеры в мм: длина крыла 8.0, ширина 3.0.
Сравнение. См. определительную таблицу видов Purbimodus.
Замечание. Слияние CuA1 с M4, вероятно, является индивидуальной особенностью этого экземпляра.
Материал. Голотип.
Род Burimodus Sukatsheva et Aristov, gen. nov.
Название рода от Бурятии и modus лат. – форма.
Типовой вид – B. novus sp. nov.
Диагноз. Крупные насекомые с несколько вытянутыми крыльями (длина превышает ширину более чем в три раза). Жилки четкие, сильные. Костальное и субкостальное поля одинаковой ширины. Sc и R прямые, параллельные. Имеются все пять апикальных развилков. Ячея DC в 1.6 раза длиннее ячеи MC, начинается значительно проксимальнее ее начала. Ствол ячеи MC почти равен по длине самой ячее. Ячея TC короткая, только в 1.1 раза длиннее ячеи MC. Развилок F5 начинается значительно проксимальнее всех остальных развилков. CuP и A1 короткие, впадают в задний край крыла в одной точке заметно проксимальнее середины длины крыла. Окончание CuP десклеротизовано. A2 короткая.
Cостав. Типовой вид.
Сравнение. См. определительную таблицу родов Vitimotauliidae.
Burimodus novus Sukatsheva et Aristov, sp. nov.
Табл. VI , фиг. 3
Название вида novus лат. – новый.
Голотип – ПИН, № 5340/1456, прямой и обратный отпечатки полного переднего крыла; Республика Бурятия, местонахождение Хасурты; нижний мел; гусиноозерская серия.
Описание (рис. 2, а). Крыло узкое, длина превышает ширину в 3.3 раза. Передний край крыла слабовыпуклый. Вершина крыла, вероятно, вытянута (сохранность не позволяет сказать точно). Sc длинная, оканчивается на уровне середины третьей четверти длины крыла. Вершина развилка F1 расположена заметно проксимальнее вершины F2. Развилки F1–F4 длинные. Поперечная жилка rs2–rs3, закрывающая ячею DC, прямая. Вершина F3 расположена чуть дистальнее вершины F4 и на одном уровне с вершиной F2. Развилок F4 намного короче F3. F5 начинается проксимальнее всех остальных развилков. Поперечная m2–m3, закрывающая ячею MC, слабо косая. Поперечная m3+4–cua1+2, закрывающая ячею TC, слабо дугообразная. A2 короткая. Анальное поле широкое.
Размеры в мм: длина крыла 13.1, ширина крыла 5.5.
Материал. Кроме голотипа, паратип ПИН, № 5026/588 из того же местонахождения.
СЕМЕЙСТВО PHILOPOTAMIDAE STEPHENS, 1836
Типовой род – Philopotamus Stephens, 1829.
Диагноз. Ручейники в основном средних размеров (4–9 мм). Передние крылья вытянуто-яйцевидные, закругленные на вершине, обычно с широким костальным полем, всегда пересеченным косой срединной жилкой. Имеются все пять апикальных развилков, редко отсутствуют развилки F4 и F1. Ячеи DC и ТС закрытые и довольно короткие. Ячея MC закрытая (кроме рода Wormaldia MacLachlan, 1865). Особенно необычно короткая ячея TC, которая равна или короче развилка F5, равна или чуть длиннее ячеи DC. A2 и A3 короткие. CuP и анальные жилки впадают в задний край крыла в одной точке или близко друг от друга. Анальное поле широкое. Антенны сильные, но не длинные, с короткими члениками. Первый членик нижнечелюстных щупиков короткий, второй длиннее, снабжен щетинками; четвертый равен второму, третий длиннее их; пятый кольчатый, равен по длине третьему и четвертому, вместе взятым. Личинки строят длинные трубки, обитают в горных потоках, родниках или реках. Всего известно более 1000 видов современных филопотамид, распространенных по всему миру.
Состав. 25 родов, из них 12 ископаемых. Стратиграфическое и географическое распространение ископаемых представителей этого семейства, описанных до 2009 г., см.: Wang et al., 2009b. Список таксонов, описанных позже, а также дополнение к ранее описанным, см. в табл. 2.
Таблица 2.
Стратиграфическое и географическое распространение ископаемых представителей семейства Philopotamidae, не учтенных в публикации Wang et al., 2009b
| Неоген | Миоцен | Саксонский янтарь | Wormaldia adventaria Mey, 1988 | Mey, 1988 |
|---|---|---|---|---|
| Палеоген | Эоцен | Балтийский янтарь | Holocentropus cavus Mey, 1985 | Mey, 1985 |
| Ровенский янтарь | Holocentropus flexiflagrum Melnitskiy et Ivanov, 2010 | Мельницкий, Иванов, 2010 | ||
| Wormaldia nastientia Melnitskiy et Ivanov, 2010 | Мельницкий, Иванов, 2010 | |||
| Wormaldia pobeda Melnitskiy et Ivanov, 2010 | Мельницкий, Иванов, 2010 | |||
| Electrocyrnus perpusillus Melnitskiy et Ivanov, 2010 | Мельницкий, Иванов, 2010 | |||
| Мел | В. мел | Бирманский янтарь | Wormaldia resina Wichard et Wang, 2016 | Wichard, Wang, 2016 |
| Wormaldia cretacea Wichard et Wang, 2016 | Wichard, Wang, 2016 | |||
| Wormaldia myanmar Wichard et al., 2005 | Wichard et al., 2005 | |||
| Кзыл-Джар, Казахстан | Kulickiella roziczkae Sukatsheva, 2001 | Sukatsheva, 2001 | ||
| Н. мел | Хасурты, Россия | Juraphilopotamus callidus Sukatsheva et Aristov, sp. nov. | ориг. | |
| Н. мел–в. юра | Кемпендяй, Россиия | Kempia piotri Sukatsheva, 2018 | Аристов, Сукачева, 2018 | |
| Mesoviatrix paradoxa Sukatsheva, 2018 | Аристов, Сукачева, 2018 | |||
| Юра | Ср. юра | Кубеково, Россия | Juraphilopotamus funeralis Sukatsheva et Vasilenko, 2020 | Сукачева, Василенко, 2020 |
| Archphilopotamus kubekovensis Sukatsheva et Vasilenko, 2020 | Сукачева, Василенко, 2020 |
Замечания. Philopotamidae – одно из наиболее примитивных семейств ручейников, находящихся в корне их филогенетического древа. Самые древние из них были найдены в среднем или верхнем триасе Киргизии в местонахождении Мадыген – Philopotamus asiaticus Sukatsheva, 1973 (Сукачева, 1973; Дмитриев и др., 2018).
Род Juraphilopotamus Wang, Zhao et Ren, 2009
Определительная таблица видов рода Juraphilopotamus Wang, Zhao et Ren, 2009
1(4) Довольно крупные насекомые, длина переднего крыла не менее 11 мм. Развилок на R тупоконечный. Ячея DC короче ячеи MC в 1.3 раза
2(3) Имеется поперечная жилка cua2–cup. Длина переднего крыла 14 мм. Окраска в виде темных пятен................................................................... ................J. callidus Sukatsheva et Aristov, sp. nov.
3(2) Поперечная жилка cua2–cup отсутствует. Длина передних крыльев 11–12 мм. Окраски в виде пятен нет.................................................... ......................J. lubricus Wang, Zhao et Ren, 2009
4(1) Мелкие насекомые, длина передних крыльев 5.0 мм. Развилок на R остроконечный. Ячея DC короче ячеи MC в четыре раза..................... ..............J. funeralis Sukatsheva et Vasilenko, 2020
Замечания. Род Juraphilopotamus был описан (Wang et al., 2009b) из средней юры Китая (Внутренняя Монголия, местонахождение Даохугоу). Интересно, что этот род сходен, с одной стороны, с родом Burimodus gen. nov. из местонахождения Хасурты (см. выше), а с другой стороны, с родом Mesoviatrix Sukatsheva, 2018 из верхней юры–нижнего мела Якутии (местонахождение Кемпендяй: Аристов, Сукачева, 2018) и с Kulickiella roziczkae Sukatsheva, 2001 из верхнего мела Казахстана (местонахождение Кзыл-Джар; Sukatsheva, 2001). Все указанные роды близки по наличию развилка на R; первые два – еще и по довольно крупным размерам (14–15 мм), а вторые два, наоборот, по мелким размерам (6–7 мм). Отличаются все роды по взаимному расположению развилков, длинам Sc, A1 и CuP и длинам ячей DC, MC и TC. На основании жилкования передних крыльев мы относим род Juraphilopotamus к семейству Philopotamidae, несмотря на предложение Гао и др. (Gao et al., 2013) поместить этот род в семейство Hydrobiosidae. С нашей точки зрения, жилкование всех видов рода обладает достаточно важными систематическими признаками, типичными для семейства Philopotamidae: широким костальным полем с дополнительной ветвью Sc2, закрытыми ячеями DC, MC и TC, более короткой ячеей DC по сравнению с MC, впадением CuP и A1 в одной точке в задний край крыла, широким анальным полем и яйцевидно закругленной формой крыльев. Кроме того, на А3 имеется излом, обычно соответствующий месту впадения поперечной жилки, соединяющей А3 с задним краем крыла. Такая поперечная жилка является важнейшим диагностическим признаком Philopotamidae. Мы считаем, что отнесение Juraphilopotamus к Hydrobiosidae на основании того, что первый и второй членики максиллярных пальп Juraphilopotamus более сходны по форме с таковыми у Hydrobiosidae, недостаточно обоснованно. Кроме того, на помещенных в работе Гао с соавт. фотографиях нет четкого изображения максиллярных щупиков, и быть уверенными в такой интерпретации затруднительно.
Juraphilopotamus callidus Sukatsheva et Aristov, sp. nov.
Табл. VI , фиг. 4
Название вида callidus лат. – искусный.
Голотип – ПИН, № 5026/595, прямой и обратный отпечатки полного переднего крыла; Республика Бурятия, местонахождение Хасурты; нижний мел; гусиноозерская серия.
Описание (рис. 2, б). Крыло широкое, длина превышает ширину в 2.2 раза. Вершина закруглена, расположена напротив окончания RS4. Наибольшая ширина крыла на уровне окончания CuA2. Sc перед дистальной третью крыла плавно изгибается вперед. Имеется косая жилка c–sc, расположенная чуть проксимальнее середины второй четверти длины крыла на уровне впадения A2 в A1. R длинный, с ковшеобразным развилком у вершины. Костальное поле на уровне разветвления RS в два раза шире субкостального. Ячея DC короткая, в 1.3 раза короче ячеи MC и в 1.5 раза короче своего ствола. Поперечная жилка rs2–rs3, закрывающая ячею DC, прямая, расположена значительно дистальнее вершин развилков F1 и F2. F1 начинается немного проксимальнее вершины развилка F2. Имеется прямая поперечная жилка rs4–m1+2. Ствол ячеи MC в 1.2 раза длиннее самой ячеи. Развилок F3 длиннее своего ствола в 1.3 раза. F2 длиннее своего ствола в 2.7 раза. Ячея MC закрыта прямой поперечной жилкой m2–m3. RS разветвляется в конце второй четверти длины крыла. M разветвляется чуть дистальнее середины второй четверти длины крыла и заметно проксимальнее разветвления RS. Ячея TC короткая, закрыта косой длинной поперечной m3+4–cua1+2. Развилок F5 мощный, длинный. Поле между CuA1 и CuA2 сужено в середине. CuP и A1 довольно короткие, оканчиваются на заднем крае крыла немного проксимальнее середины длины крыла в одной точке. Вершины CuP и А1 десклеротизованы. Имеется слабо косая длинная жилка cua2–cup. A1 и A2 средней величины. A2 в 1.6 раза короче A1. A3 очень короткая. Имеется четкая cua2–a1. Окраска в виде разбросанных по крылу небольших темных пятен.
Размеры в мм: длина крыла 14.0, ширина 5.0.
Сравнение. См. определительную таблицу рода Juraphilopotamus.
Материал. Голотип.
ОБСУЖДЕНИЕ
Анализ описанных в данной работе имаго ручейников позволяет несколько уточнить геологический возраст местонахождения Хасурты. Род Juraphilopotamus (семейство Philopotamidae) описан по виду J. lubricus Wang, Zhao et Ren, 2009 из средней юры Китая (местонахождение Даохугоу: Wang et al., 2009b). Второй вид этого рода J. funeralis Sukatsheva et Vasilenko, 2020 описан из средней юры Красноярского края (местонахождение Кубеково: Сукачева, Василенко, 2020). Это могло бы указывать на юрский и даже на среднеюрский характер фауны. С другой стороны, высокое разнообразие семейства Vitimotauliidae, как уже говорилось выше, характерно для раннего мела, включая китайскую фауну Жехэ (Исянь, Дабейгоу и др.), которую часто сопоставляют с окрестностями границы баррема и апта (около 125 млн лет: Chang et al., 2017) и с которой хорошо соотносятся фауны Байсы и других отложений в Забайкалье (фауна Ephemeropsis–Coptoclava, по: Жерихин, 1978). Однако Purbimodus parvulus sp. nov. принадлежит к роду, все остальные виды которого описаны из пурбека Англии (табл. 1), т.е., из надежно датированного берриаса (самое основание нижнего мела). Помимо описанных выше ручейников семейств Vitimotauliidae и Philopotamidae, из Хасурты ранее были описаны представители семейства Dysoneuridae. Этот комплекс ручейников близок к комплексам местонахождений Унда и Утан (верхняя юра–нижний мел Забайкалья, Россия: Сукачева, Василенко, 2019) и не позволяет датировать его более поздним временем, чем начало раннего мела.
Список литературы
Аристов Д.С., Сукачева И.Д. Новые насекомые (Insecta: Trichoptera, Reculida, Eoblattida) из мезозоя Азии // Палеонтол. журн. 2018. № 4. С. 53–61.
Дмитриев В.Ю., Аристов Д.С., Башкуев А.С. и др. Разнообразие насекомых от карбона до современности // Палеонтол. журн. 2018. № 6. С. 21–31.
Жерихин В.В. Развитие и смена меловых и кайнозойских фаунистических комплексов (трахейные и хелицеровые). М.: Наука, 1978. 197 с. (Тр. Палеонтол. ин-та АН СССР. Т. 165).
Копылов Д.С., Расницын А.П. Новые сепульки (Hymenoptera: Sepulcidae) из нижнего мела Азии. I: Parapamphiliinae и Xyelulinae // Палеонтол. журн. 2017. № 1. С. 66–72.
Мельницкий С.И., Иванов В.Д. Новые виды ручейников (Insecta: Trichoptera) из ровенского янтаря (эоцен Украины) // Палеонтол. журн. 2010. № 3. С. 57–64.
Сукачева И.Д. Мезозойские ручейники (Trichoptera) Забайкалья // Палеонтол. журн. 1968. № 2. С. 59–75.
Сукачева И.Д. Новые ручейники (Trichoptera) из мезозоя Средней Азии // Палеонтол. журн. 1973. № 5. С. 100–107.
Сукачева И.Д. Историческое развитие ручейников (Trichoptera) // Тр. Палеонтол. ин-та АН СССР. 1982. Т. 197. 112 с.
Сукачева И.Д. Ручейники. Phryganeida // Позднемезозойские насекомые Восточного Забайкалья. М.: Наука, 1990. С. 94–123 (Тр. Палеонтол. ин-та АН СССР. Т. 239).
Сукачева И.Д. Новые ископаемые представители отряда ручейников (Phryganeida) из Монголии // Новые таксоны ископаемых беспозвоночных Монголии. М.: Наука, 1992. С. 111–117 (Тр. Совм. Росс.-Монгол. палеонтол. экспед. Вып. 41).
Сукачева И.Д., Василенко Д.В. Новые ручейники семейства Dysoneuridae (Insecta, Trichoptera) из мезозоя Забайкалья (местонахождение Хасурты) // Палеонтол. журн. 2019. № 5. С. 63–68.
Сукачева И.Д., Василенко Д.В. Новые ручейники семейства Philopotamidae (Insecta, Trichoptera) из средней юры Сибири (местонахождение Кубеково) // Палеонтол. журн. 2020. № 1. С. 50–55.
Chang Su-Chin, Gao Ke-Qin, Zhou Chang-Fu, Jourdan F. New chronostratigraphic constraints of the Yixian Formation with implications for the Jehol Biota // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2017. № 487. P. 399–406.
Davis S.R., Engel M.S., Ren D. A pupal caddisfly from the Early Cretaceous of China (Trichoptera) // Cret. Res. 2010. V. 31. P. 396–399.
Gao Y., Yao Y., Ren D. A new Middle Jurassic caddisfly (Trichoptera, Hydrobiosidae) from China // Fossil Rec. 2013. V. 16. № 1. P. 111–116.
Mey W. Die Köcherfliegen des Sächsischen Bernsteins (I) // Dtsch. entomol. Z. 1985. Bd 32. H. 4–5. S. 275–278.
Mey W. The caddisflies of the Saxonian Amber (III) (Trichoptera) // Dtsch. entomol. Z. 1988. Bd 35. H. 4–5. S. 299–309.
Lin Q. The Jurassic fossil insects from Western Liaoning // Acta Palaeontol. Sin. 1976. V. 15. № 1. P. 98–117.
Ren D. Insect: Trichoptera // Faunae and Stratigraphy of the Jurassic–Cretaceous in Beijing and the Adjacent Area / Eds. Ren D., Lu L.W., Gao Z.G., Ji S.A. Beijing: Seismic Publ. House, 1995. P. 95–97 (in Chinese).
Sukatsheva I.D. New representative of Philopotamidae (Trichoptera) family from the Upper Cretaceous of Kazakhstan // Prace Museum Ziemi. 2001. № 46. P. 65–66.
Sukatsheva I.D., Jarzembowski E.A. Fossil caddisflies (Insecta: Trichoptera) from the Early Cretaceous of South England II // Cret. Res. 2001. V. 22. P. 685–694.
Wang M., Liang J., Ren D. New fossil Vitimotauliidae (Insecta: Trichoptera) from the Jehol Biota of Liaoning Province, China // Cret. Res. 2009a. V. 30. P. 592–598.
Wang M., Zhao Y., Ren D. New fossil caddisfly from Middle Jurassic of Daohugou, Inner Mongolia, China (Trichoptera: Philopotamidae) // Science Direct Progress in Natural Science. 2009b. V. 19. P. 1427–1431.
Wichard W., Poinar G.O., Corvallis J. Köcherfliegen aus dem Birma Bernstein der oberen Kreide von Myanmar (Insecta, Trichoptera) // Mitt. Geol. Paläontol. Inst. Univ. Hamburg. 2005. H. 89. S. 129–136.
Wichard W., Wang M.X. New Cretaceous caddisflies from Burmese amber (Insecta, Trichoptera) // Cret. Res. 2016. V. 61. P. 129–135.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал