

Палеонтологический журнал, 2020, № 6, стр. 43-54
Редкие длинноусые двукрылые (Insecta: Diptera) из местонахождения Хасурты, Забайкалье
Е. Д. Лукашевич *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: elukashevich@hotmail.com
Поступила в редакцию 28.12.2019
После доработки 16.01.2020
Принята к публикации 16.01.2020
Аннотация
Из нижнемелового местонахождения Хасурты (Западное Забайкалье, Россия) описаны новые таксоны длинноусых двукрылых Praearchitipula ribeiroi sp. nov., P. kaluginae sp. nov., P. podenasi sp. nov. (Pediciidae), ?Protanyderus madrizi sp. nov. (Tanyderidae), Eoptychoptera fasbenderi sp. nov. (Ptychopteridae), Gilkania transbaikalica gen. et sp. nov. (Chironomidae), Amorimyia robusta gen. et sp. nov. (Anisopodidae).
В 2003 г. в Западном Забайкалье геологами “Бурятгеоцентра” (Улан-Удэ) было обнаружено местонахождение мезозойских насекомых Хасурты, а собранные оттуда насекомые были переданы О.Р. Мининой в Палеонтологический ин-т РАН (ПИН). За прошедшие годы лабораторией артропод ПИН было проведено несколько экспедиций (в 2004, 2009, 2014 и 2019 гг.). Местонахождение оказалось очень богатым, был собран большой материал, доминирующим отрядом насекомых оказались двукрылые – в коллекциях ПИН замаркировано около 2500 отпечатков.
Описание двукрылых из этого местонахождения началось недавно. Пока описаны представители лишь немногих редких семейств Nematocera, а именно Axymyiidae (Blagoderov, Lukashevich, 2013), Bolitophilidae (Greenwalt, Blagoderov, 2019), Kaluginamyiidae (Lukashevich et al., 2020), некоторые Ptychopteridae (Lukashevich, 2019) и Perissommatidae (Lukashevich, Blagoderov, 2020). Данная статья продолжает серию систематических описаний длинноусых двукрылых из Хасурты, редко встречающихся в палеонтологической летописи мезозоя.
Все описанные ниже экземпляры происходят из местонахождения Хасурты, расположенного на левом берегу р. Хасурты в ее среднем течении в Закаменском р-не Бурятии (50°21′ N; 103°38′ E); отложения относятся к нижней части гусиноозерской серии, нижний мел. Типовые материалы хранятся в Палеонтологическом ин-те им. А.А. Борисяка РАН, Москва.
В работе используется номенклатура жилкования и система Nematocera, принятая нами ранее (Shcherbakov et al., 1995; Lukashevich, Ribeiro, 2019). Сканирующие электронные фотографии отпечатков без напыления были сделаны с помощью микроскопа Tescan Vega XMU с использованием обратного (BSE) и вторичного электронного (SE) детекторов.
Автор выражает сердечную благодарность Р. Ракитову (ПИН) за помощь при работе на сканирующем электронном микроскопе и А. Башкуеву (ПИН) за тщательное препарирование многих экземпляров. Работа выполнена при частичной поддержке РФФИ, проект № 18-04-00322 и Программы I.21П “Биоразнообразие природных систем”.
ИНФРАОТРЯД TIPULOMORPHA
СЕМЕЙСТВО PEDICIIDAE OSTEN-SACKEN, 1860
ПОДСЕМЕЙСТВО PEDICIINAE OSTEN-SACKEN, 1860
Замечания. К подсемейству принадлежит подавляющее большинство ныне живущих родов этого небольшого семейства, 11 из 12. Характеризуется голой, лишенной макротрихий крыловой пластинкой. Как и для семейства в целом, для подсемейства характерно наличие шпор на голенях всех ног и положение sc-r проксимальнее основания радиального сектора Rs (Савченко, 1989).
Личинки Pediciinae – преимущественно водные и полуводные, обитают в переувлажненной или болотистой почве, часто среди водных и полуводных мхов, хотя известен род, развивающийся в толще разлагающейся древесины лежащих на земле стволов (Кривошеина, 2011). При таком разнообразии освоенных биотопов о местообитании личинок мезозойского рода, обсуждаемого ниже, остается только гадать. Все известные гидробионтные личинки Pediciinae являются зоофагами.
Из ныне живущих родов педициид в ископаемом состоянии известен лишь один, Tricyphona Zetterstedt, 1837 (кайнозой Европы, эоценовый балтийский янтарь; Alexander, 1931; Podenas, 1999, 2001). Кроме того, описано два вымерших рода из Азии: кайнозойский монотипический Fragisternella Krzemiński, 2000 по единственному неполному отпечатку крыла из Приморья России (Krzemiński, 2000) и мезозойский Praearchitipula из трех юрских местонахождений России и Китая (Калугина, Ковалев, 1985; Lukashevich, 2009; Gao et al., 2015). Этот же род Praearchitipula обнаружен в Хасурты. Как и у других представителей подсемейства, у Praearchitipula макротрихии на крыле располагаются только по жилкам.
Род Praearchitipula Kalugina in Kalugina et Kovalev, 1985
Замечания. Род первоначально был описан в Limoniidae, но оказался сборным – только один вид является лимониидой, а типовой и остальные виды были переведены в педицииды (Lukashevich, 2009). До сих пор было описано шесть видов; все, несомненно, юрские: P. notabilis Kalugina, 1985, P. trinervis Kalugina, 1985 (оба – Кубеково), P. spasskia Kalugina, 1985 (Новоспасское), P. abnormis (Hao et Ren, 2009), P. apprima Gao et al., 2015 и P. mirabilis Gao et al., 2015 (все – Даохугоу).
Уточненный диагноз рода был опубликован ранее (Gao et al., 2015). У новых видов, как у большинства видов рода, Sc длинная, оканчивается дистальнее ответвления Rs, sc-r расположен в проксимальной половине крыла, R1 длинная, R2 поперечная, у края R1, ствол Rs разветвляется на длинный R2 + 3 и короткий R4 + 5 (иногда отсутствует), положение r-m варьирует (упирается в R5 или R4 + 5), m-cu упирается в середину M3 + 4 (иногда дистальнее).
Ниже впервые описываются терминалии самца Praearchitipula (терминология по Савченко, 1989); кроме того, впервые обнаружены очень крупные представители рода (длина крыла 17.0–17.6 мм), а на крыле у основания Rs – излом, иногда со шпорой. Подобные крупные комары известны среди ныне живущих родов Pedicia и Tricyphona (Alexander, Byers, 1981; Савченко, 1989), в обоих родах встречается и относительно длинный радиальный сектор, начинающийся в проксимальной части крыла, и излом у основания Rs (иногда со шпорой), и одна терминальная пара черпаковидых гоностилей. Кроме того, для Tricyphona, как и для Praearchitipula, характерно ветвление Rs на R2 + 3 и R4 + 5 и отмечена значительная изменчивость в ветвлении радиального сектора: внутри вида может варьировать характер ветвления Rs (на R2 + 3 + 4 и R5 или на R2 + 3 и R4 + 5), длина ствола R4 + 5 и его наличие (отсутствует, если R2 + 3, R4 и R5 расходятся из одной точки: Alexander, 1962), наличие шпоры при изломе Rs (Podenas, 2001) и положение r-m (упирается в ствол или развилок R4 + 5: Alexander, 1953). Но между Tricyphona и Praearchitipula есть, как минимум, одно важное различие в опушении глаз.
Педицииды – единственное семейство Tipuloidea с опушенными глазами, что обычно считается симплезиоморфией (Oosterbroek, Theowald, 1991). Как и у другого семейства типуломорф, Trichoceridae, у современных Pediciidae опушение глаз известно не у всех родов, но у подавляющего их большинства (Савченко, 1989). У всех современных Tricyphona глаза опушены, причем короткие редкие волоски между фасетками были обнаружены и в ископаемом состоянии у некоторых кайнозойских видов этого рода (Podenas, 2001). У Praearchitipula, описываемых ниже, опушение глаз не обнаружено, несмотря на специально предпринятое изучение на сканирующем электронном микроскопе нескольких хорошо сохранившихся голов (табл. VII , фиг. 1; см. вклейку). Отсутствие опушения глаз уже отмечалось у китайского вида этого рода P. apprima (Gao et al., 2015). Так что о синонимизации Praearchitipula речь не идет, но вопрос о взаимоотношениях Praearchitipula и Tricyphona, несомненно, заслуживает дальнейшего изучения.
Рис. 1.
Представители рода Praearchitipula (Pediciidae) из нижнего мела Хасурты: а, б – P. ribeiroi sp. nov., голотип ПИН, № 5340/1100, крыло (а) и терминалии самца (б); в, г – P. kaluginae sp. nov., голотип ПИН, № 5026/1633, рисунок и фото крыла; д, е – P. podenasi sp. nov., голотип ПИН, № 5340/1104, рисунок и фото крыла. Стрелки маркируют поперечную жилку sc-r.
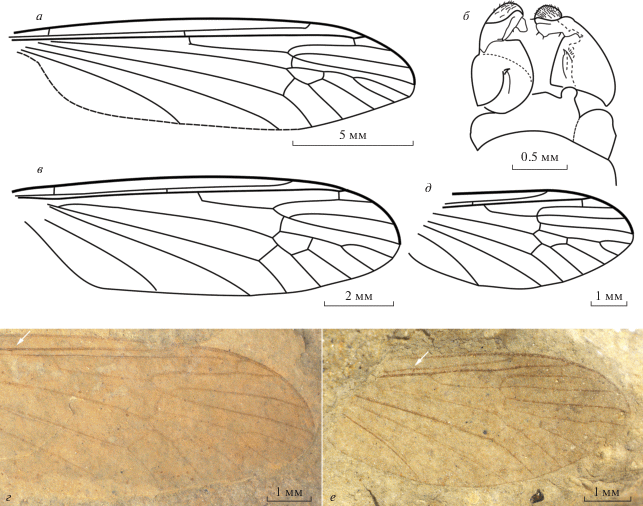
Praearchitipula ribeiroi Lukashevich, sp. nov.
Табл. VI , фиг. 1; табл. VII , фиг. 1 (см. вклейку)
Название вида – в честь бразильского диптеролога Г. Рибейро (G. Ribeiro).
Голотип – ПИН, № 5340/1100±, позитивный и негативный отпечатки самца хорошей сохранности с отчлененной головой, лежащей рядом; Хасурты; нижний мел.
Описание (рис. 1, а, б). Крупные комары. Тело и его придатки темные, ноги окрашены равномерно. Волоски на границе фасеток не видны. Крыловая пластинка прозрачная, без пятен, кроме очень бледной птеростигмы. Поперечная sc-r прямо перед ответвлением Rs или удалена от него на заметное расстояние (в 1.5 раза короче М3 + 4), Rs длинный, начинается в проксимальной части крыла, ствол Rs с изломом в основании и со шпорой, в 1.3–1.5 раза длиннее R2 + 3. Ствол R4 + 5 короткий, не длиннее поперечной r-m, которая упирается прямо в развилок R4 + 5. Вилка М1 + 2 в два раза длиннее dM1 + 2 и в 1.5 раза длиннее дискоидальной ячейки; поперечная m-cu расположена в проксимальной половине короткой дискоидальной ячейки, bM3 + 4 в 1.5 раза короче dM3 + 4. Дополнительных поперечных жилок нет. Гипопигий самца компактный, не шире предыдущих сегментов, строение до конца не ясно: вероятно, IX тергит поперечный, на вершине с широкими округлыми боковыми выростами, гонококситы массивные, при основании с небольшими узкими интербазами, заостренными на вершине, дистально с одной парой неявственно дифференцированных гоностилей, гоностили черпаковидные, на вершине с многочисленными щетинками, мезально, возможно, вытянуты в широкий, тупой ростральный выступ с несколькими мелкими шипиками.
Размеры в мм: голотип: длина головы 1.5, груди 2.9, брюшка 12.5, бедра переднего 8.5, среднего 8.0, заднего 9.5, задней голени 9.8, лапки 11.2; длина крыла 17.2 (голотип) – 17.6.
Сравнение. Гораздо крупнее всех ранее описанных видов; по жилкованию близок к P. abnormis, отличаясь более длинным стволом Rs (у китайского вида ствол Rs соразмерен R2 + 3), отсутствием темной птеростигмы, наличием шпоры в основании Rs.
Материал. Кроме голотипа, паратип ПИН, № 5026/1635 (изолированное крыло).
Praearchitipula kaluginae Lukashevich, sp. nov.
Название вида – в память российского диптеролога Н.С. Калугиной.
Голотип – ПИН, № 5026/1633±, позитивный и негативный отпечаток самки; Хасурты; нижний мел.
Описание (рис. 1, в, г). Крупная самка. Тело темное, яйцеклад светлый. Крыловая пластинка прозрачная, без пятен. Поперечная sc-r заметно удалена от ответвления Rs (расстояние соразмерно длине М3 + 4); Rs длинный, начинается в проксимальной половине, близко к середине крыла, ствол Rs с изломом в основании, в 1.4 раза длиннее R2 + 3. Ствол R4 + 5 короткий, соразмерен поперечной r-m, которая в него упирается. Вилка М1 + 2 в два раза длиннее dM1 + 2 и в 1.5 раза длиннее дискоидальной ячейки, m-cu упирается в проксимальную половину короткой дискоидальной ячейки, bM3 + 4 в два раза короче dM3 + 4. Дополнительных поперечных жилок нет.
Размеры в мм: длина груди 2.3, брюшка (с яйцекладом) 10.3, крыла 12.0.
Сравнение. По положению r-m, упирающейся в ствол R4 + 5, новый вид близок только к P. apprima, отличаясь проксимальным положением m-cu (у китайского вида bM3 + 4 в два раза длиннее dM3 + 4) и отсутствием видимой птеростигмы.
Замечания. Жилкование P. kaluginaе sp. nov. очень близко к жилкованию P. ribeiroi sp. nov., отличаясь лишь положением r-m и отсутствием шпоры у основания Rs, что вполне может быть проявлением индивидуальной изменчивости (см. выше). Но поскольку подобное положение r-m у Praearchitipula было зафиксировано лишь однажды (являясь обычным, например, у современной Tricyphona), а самки обычно крупнее самцов (тогда как самец P. ribeiroi sp. nov. заметно крупнее обсуждаемой самки), целесообразно выделить новый вид.
Материал. Голотип.
Praearchitipula podenasi Lukashevich, sp. nov.
Название вида – в честь литовского диптеролога С. Поденаса (S. Podenas).
Голотип – ПИН, № 5340/1104±, позитивный и негативный отпечаток изолированного крыла; Хасурты; нижний мел.
Описание (рис. 1, д, е). Комары средних размеров. Крыловая пластинка прозрачная, без пятен, кроме бледной птеростигмы. Поперечная sc-r удалена от ответвления Rs (расстояние в 1.1–2.0 раза короче М3 + 4); Rs относительно короткий, начинается в проксимальной части или прямо посередине крыла, ствол Rs плавно изгибается, соразмерен R2 + 3. Ствол R4 + 5 очень короткий, короче r-m, или отсутствует и R2 + 3, R4 и R5 расходятся из одной точки; поперечная r-m упирается в R5 и соразмерна bR5. Вилка М1 + 2 не длиннее dM1 + 2 и в 1.2–1.4 раза короче дискоидальной ячейки, bM3 + 4 равен dM3 + 4. Дополнительных поперечных жилок нет.
Размеры в мм: длина крыла 7.0–7.5 (голотип).
Сравнение. По жилкованию с практически отсутствующим R4 + 5 близок к P. trinervis (Lukashevich, 2009, рис. 3C ), но крупнее, и отличается более коротким стволом Rs и более длинным базальным отрезком R5.
Материал. Кроме голотипа, паратипы: изолированные крылья ПИН, №№ 5026/297±, 1648±.
* * *
Помимо описанного выше материала, к роду Praearchitipula относятся еще пять отпечатков имаго из местонахождения Хасурты. Жилкование крыльев имаго плохой сохранности ПИН, №№ 5026/1634±, 1645± и ПИН, №№ 5340/805±, 1648±, 2753± напоминает P. ribeiroi sp. nov., но они мельче (длина крыла 7.6–11.5 мм), и у некоторых из них не видно шпоры у основания Rs. Не исключено, что среди этих экземпляров содержатся и другие виды.
ИНФРАОТРЯД PSYCHODOMORPHA
СЕМЕЙСТВО TANYDERIDAE OSTEN-SACKEN, 1880
Замечания. Реликтовое семейство комаров, объединяющее всего десяток современных и несколько вымерших родов, ныне обитающих преимущественно в умеренном климате обоих полушарий (Eskov, Lukashevich, 2015). Личинки танидерид известны из двух биотопов: некоторые роды (в том числе Protanyderus) обитают на песчаном или галечном дне разнообразных водотоков, другие – в погруженных в воду мертвых стволах и ветках на начальной стадии разложения, являясь полуводными ксилофагами (Lukashevich, Shcherbakov, 2014).
Род Protanyderus Handlirsch, 1909
Замечания. Современный род Protanyderus оказался широко распространенным в юрских отложениях Азии (Lukashevich, Krzemiński, 2009; Dong et al., 2015). К настоящему времени описано восемь юрских видов Protanyderus из России (Сибирь), Казахстана, Монголии, Китая, причем самый древний – из средней юры (Lukashevich, Krzemiński, 2009). Уже высказывались сомнения в правильности отнесения всех этих юрских танидерид к современному роду, поскольку большинство танидерид описаны по изолированным крыльям (Lukashevich, 2020). Единственный известный из юрских отложений самец P. nebulosus Lukashevich, Krzemiński, 2009 имеет плохо интерпретируемые, но, несомненно, неинвертированные терминалии, в отличие от современных представителей рода, что только подкрепляет сомнения. Правда, в пределах юрского рода танидерид Praemacrochile Kalugina, 1985 также отмечена вариабельность по наличию или отсутствию поворота терминалий (Lukashevich, 2018).
?Protanyderus madrizi Lukashevich, sp. nov.
Табл. VI , фиг. 2, 3
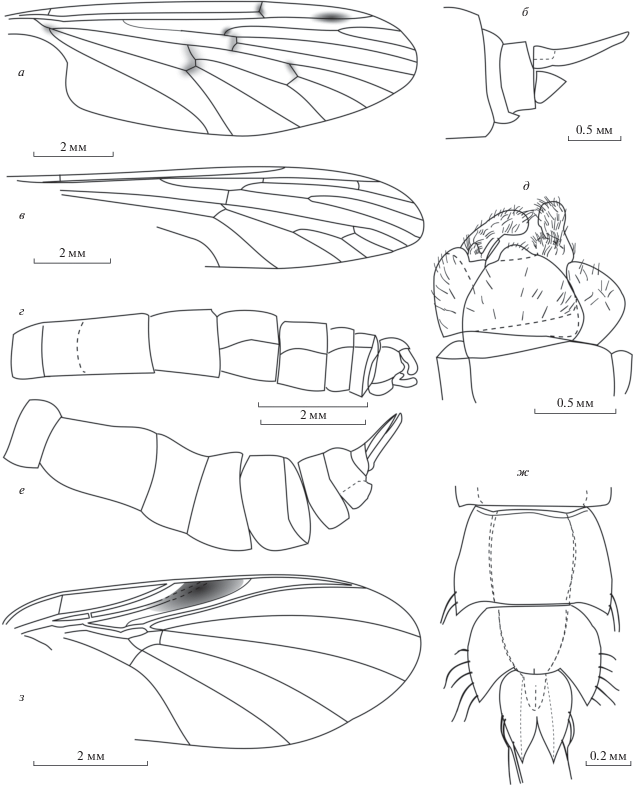
Рис. 2.
Представители Nematocera из нижнего мела Хасурты: а, б – ?Protanyderus madrizi sp. nov. (Tanyderidae), голотип ПИН, № 5340/2733, изолированное крыло (а) и паратип ПИН, № 5340/1499, терминалии самки (б); в–е – Eoptychoptera fasbenderi sp. nov. (Ptychopteridae), голотип ПИН, № 5340/2735, крыло (в), брюшко (г) и терминалии самца (д) и паратип ПИН, № 5340/2744, брюшко самки (е); ж – Gilkania transbaikalica gen. et sp. nov. (Chironomidae), голотип ПИН, № 5340/2952, вершина брюшка куколки; з – Amorimyia robusta gen. et. sp. nov. (Anisopodidae), голотип ПИН, № 5340/2978, крыло.
Название вида – в честь американского диптеролога Р. Мадрица (R.I. Madriz).
Голотип – ПИН, № 5340/2733±, позитивный и негативный отпечаток изолированного крыла; Хасурты; нижний мел.
Описание (рис. 2, а, б). Крупные комары. Крыло длиннее брюшка, в 3 раза длиннее своей ширины, с бледной птеростигмой и небольшими бледными пятнами на всех поперечных жилках и почти на всех развилках (не видно на развилке R2 + 3). Sc оканчивается сразу за проксимальной третью R2 + 3; R1 достигает середины R2; R2 относительно длинная, R2 + 3 в 1.7 раза длиннее R2; R5 в 1.7 раза длиннее ствола Rs, чья проксимальная половина заметно тоньше дистальной; M1 соразмерна M1 + 2; ячейка m2 в 1.6–2.0 раза шире ячейки m3 у края крыла. Церки самки удлиненные.
Размеры в мм: длина крыла голотипа 11.1, длина брюшка 8.5.
Сравнение. Новый вид близок к P. nebulosus, от которого отличается относительно более длинной М1 (у P.nebulosus она заметно короче ствола М1 + 2) и отсутствием пятен на жилках R5 и M4.
Замечания. Самцы опять не найдены, так что ни подтвердить, ни опровергнуть правильность отнесения этих танидерид к современному роду пока не удалось. У самок современных видов этого рода церки округлые, а не удлиненные, поэтому вид отнесен к роду с сомнением. Подобные удлиненные узкие церки, насколько известно, вообще впервые обнаружены среди танидерид.
Материал. Кроме голотипа, неполная самка с частично сохранившимся телом и крылом, экз. ПИН, № 5340/1499±.
СЕМЕЙСТВО PTYCHOPTERIDAE SCHINER, 1864
Замечания. Реликтовое семейство комаров, объединяющее всего три современных рода, было гораздо разнообразнее в мезозое, откуда известно в 4 раза больше вымерших родов (Лукашевич, 2008). Личинки птихоптерид – детритофаги, ведут полуводный образ жизни, обитая в прибрежной зоне разнообразных водоемов, включая ручьи и сфагновые болота.
Из местонахождения Хасурты уже описаны представители ныне живущего подсемейства Bittacomorphinae (Lukashevich, 2019), которые сосуществовали с мезозойскими Eoptychopterinae, описываемыми в этой статье. Подобное сосуществование в раннем мелу в одном местонахождении представителей вымерших и ныне живущих подсемейств птихоптерид уже отмечалось ранее (Калугина, 1989; Lukashevich, 1995; Lukashevich et al., 2001).
ПОДСЕМЕЙСТВО EOPTYCHOPTERINAE HANDLIRSCH, 1906
Род Eoptychoptera Handlirsch, 1906
Eoptychoptera: Handlirsch, 1906.
Proptychoptera: Handlirsch, 1906, с. 489; Carpenter, 1992, с. 406.
?Eolimnobia: Handlirsch, 1906, с. 489; Lukashevich, 2008, с. 68.
Metaxybittacus (part.): Bode, 1953, с. 287; Lukashevich et al., 1998, с. 318.
Acritorhyphus: Bode, 1953, с. 299; Shcherbakov et al., 1995, с. 97.
Palaeolimnobia: Bode, 1953, с. 301; Shcherbakov et al., 1995, с. 97.
?Prodocidia: Whalley, 1985, с. 157; Lukashevich, 2000, с. 352.
Типовой вид – Eoptychoptera simplex Handlirsch, 1906, нижняя юра Германии.
Диагноз. Комары средних и крупных размеров. Крылья с пятнами или без них; Sc и R1 длинные, поперечная r-m обычно сближена с развилком Rs и расположена проксимальнее или дистальнее его, 4 медиальных жилки, поперечная im соединяет M2 и M3. Первый сегмент брюшка нормально развит, второй удлинен, седьмой – самый узкий. Гипопигий самца обычно не шире предыдущих сегментов брюшка, гоностили соразмерны гонококситам.
Видовой состав. Кроме нового вида, 21 вид из мезозоя Евразии и Южной Америки.
Замечания. Этот род – самый разнообразный и широко распространенный среди вымерших птихоптерид, известен c ранней юры до раннего мела, обитал не только в Евразии, но и в южном полушарии, на территории современной Бразилии (Krzemiński et al., 2015; Lukashevich, Arillo, 2016). Уже в ранней юре среди древнейших Eoptychoptera крылья различались по ширине и наличию пятен, причем обычно более широкие крылья были с пятнами на большинстве развилков и поперечных жилках, а на удлиненных крыльях обнаруживается лишь птеростигма. Разница крыльев привела к тому, что и А. Хандлирш, и А. Боде (Handlirsch, 1906–1908; Bode, 1953) описывали их как два разных рода, но после описания многочисленных мезозойских Eoptychoptera формально разделить эти два типа хотя бы на уровне подродов не удалось (Lukashevich et al., 1998; Лукашевич, 2012б). Несмотря на обычность Eoptychoptera в мезозое, гениталии самца до сих пор описывались лишь у юрских E. asiatica Lukashevich, 1993 и E. aucta Lukashevich, 1993 из Каратау (Lukashevich, 1993, рис. 3b ). У этих видов, относящихся к “пятнистому типу”, гениталии не шире предыдущих сегментов, а гоностили, полностью видные лишь у E. asiatica, простые. У самца E. cantabrica Lukashevich et Arillo, 2016, относящегося к “удлиненному типу”, гениталии сохранились не полностью, однако можно предположить, что они заметно шире предыдущих сегментов брюшка. У описываемого ниже самца нового вида того же “удлиненного типа” брюшко не расширяется к концу, а гениталии явно не шире предыдущих сегментов, при этом их интерпретация затруднена (см. ниже). Это опять не позволяет разделить многоликий род на подроды.
Eoptychoptera fasbenderi Lukashevich, sp. nov.
Табл. VI , фиг. 4
Название вида – в честь американского диптеролога А. Фасбендера (A. Fasbender).
Голотип – ПИН, № 5340/2735±, позитивный и негативный отпечаток неполного самца; Хасурты; нижний мел.
Описание (рис. 2, в–е). Крупные комары. Крыло длиннее брюшка, в 4 раза длиннее своей ширины, жилки слабо затемнены, на крыловой мембране имеются небольшие макротрихии, особенно густые в передней части крыла. Sc оканчивается далеко за серединой крыла. R1 длинный, занимает 87.5% от общей длины крыла, Rs недлинный, занимает 62.5% от общей длины крыла, ветвится несимметрично, его направление продолжает R4 + 5; R2 + 3 в 2.0–2.2 раза длиннее R3; вилка R4 + 5 в 1.7–2.0 раза длиннее своей рукоятки; поперечная r-m расположена заметно проксимальнее развилка Rs. Дополнительная поперечная жилка r-rs соединяет R1 и R2 + 3 недалеко от места ответвления последней. Отрезок bas M3 + 4 гораздо короче, чем поперечные r-m и m-cu, поперечная r-m, bas M3 + 4 и m-cu образуют корду.
Брюшко довольно тонкое и длинное, не расширяется к концу, с нормально развитым сегментом I, сильно удлиненным сегментом II (равен по длине сегментам III и IV вместе взятым) и, вероятно, с разделенным стернитом II; сегменты III–V постепенно укорачиваются, сегменты VI–VII заметно уже, сегмент VII самый узкий, в виде кольца. Гонококситы, видимо, простые, гоностили пальцевидные, немного расширяющиеся на вершине, направлены внутрь, опушены длинными щетинками, видимо, сложные – с коротким базальным выростом, густо опушенным длинными щетинками; никаких зубчиков не видно. Базальный вырост, вероятно, вентральный, в несколько раз короче основного дорсального. Гипандрий массивный. Склеротизованный яйцеклад самки массивный, удлиненный (равен по длине сегментам VII + VIII), с почти прямыми церками.
Размеры в мм: длина груди 2.0–2.5 (голотип 2.0), длина брюшка7.8–8.5 (голотип 7.8), длина крыла 10.1–11.8 (голотип 11.2), ширина крыла 2.8 (голотип), длина гипопигия 0.9, яйцеклада 1.3.
Сравнение. Дополнительная поперечная жилка r-rs отличает новый вид от всех остальных видов рода. По жилкованию крыла ближе всего к E. cretaceous Kalugina, 1989 из нижнемеловой Байсы, кроме дополнительной поперечной жилки, которая отчетливо видна лишь на половине экземпляров; отличается мелкими деталями: более длинной вилкой R4 + 5 (в два раза длиннее рукоятки, а не в полтора), проксимальным положением r-m (расположена до развилка Rs, а не прямо под ним) и очень коротким bas M3 + 4.
Замечания. В ключе для видов рода, опубликованном недавно (Lukashevich, Arillo, 2016), попадает на пункт 33 – E.cretaceous (крыло без пятен, вилка R4 + 5 лишь в два раза длиннее своей рукоятки, короткий bas M3 + 4, im впадает в M2), однако bas M3 + 4 заметно короче m-cu. Дополнительная поперечная жилка в радиальном поле обнаружена впервые в пределах семейства, но дополнительная поперечная с другой локализацией уже была описана: у позднемелового Neuseptychoptera Szadziewski et al., 2018 обнаружены две r‑m, одна упирается в ствол Rs, другая – в R4 + 5 (Szadziewski et al., 2018). Дополнительная поперечная жилка r-rs в том же месте известна у типуломорф: триасового Psychotipa Shcherbakov, 1995 (Vladipteridae) и у многих видов современного рода Dicranota Zetterstedt, 1838 (Pediciidae) (Савченко, 1989; Shcherbakov et al., 1995).
Как и у ранее описанных видов рода Proptychopterina Kalugina, 1985 (Лин, Лукашевич, 2006), интерпретация гипопигия затруднена. Хотя сурстили являются неотъемлемой частью гипопигия всех современных птихоптерид, у описываемого здесь самца невозможно точно определить, наблюдается ли сочетание гоностилей с сурстилями или сложные гоностили при полном отсутствии сурстилей, т.к. голотип захоронился дорсовентрально. Как и у Proptychopterina, более короткие выросты, вероятно, занимают более вентральное положение, поэтому мы вновь считаем их не сурстилями, а вентральными выростами гоностилей. Вентральные выросты гоностиля были описаны и в другом роде, Leptychoptera Lukashevich et Azar, 2003, в подроде Burmaptychoptera Lukashevich, 2004, но там они соразмерны дорсальному выросту гоностиля (Lukashevich, Grimaldi, 2004). Однако нельзя исключить, что обсуждаемые опушенные лопасти являются выростами не гоностилей, а гонококситов, которые были обнаружены у номинативного подрода Leptychoptera Lukashevich et Azar, 2003 (Lukashevich, Azar, 2003).
Материал. Кроме голотипа, паратипы: неполные самки ПИН, №№ 5340/794±, 2744± и изолированное крыло ПИН, № 5340/1495±.
ИНФРАОТРЯД CULICOMORPHA
СЕМЕЙСТВО CHIRONOMIDAE NEWMAN, 1834
ПОДСЕМЕЙСТВО PODONOMINAE THIENEMANN ET EDWARDS, 1937
Замечания. Это небольшое подсемейство, ныне включающее всего 13 родов (Makarchenko, Endo, 2009; Cranston et al., 2010), было широко распространено уже в юре, причем обычно в юрских местонахождениях Азии наиболее многочисленны преимагинальные водные стадии (Калугина, Ковалев, 1985; Калугина, 1993). Многие современные подономины реофильны и тяготеют к водоемам с холодной, чистой и богатой кислородом водой: например, Trichotanypus Kieffer, 1906 – современный род, вероятно, наиболее близкий к описываемому здесь роду и широко распространенному в мезозое Oryctochlus Kalugina, 1985. Личинки Trichotanypus – зоофаги, развиваются в холодных предгорных, горных и тундровых водотоках, часто в моховых подушках на камнях и песчаном дне, т.е., род является кренофилом, обычно ассоциированным со мхами (Макарченко, 1985; Lindergaard, 1995). Однако не все современные подономины кренофилы, а в мезозое недавно описан меловой род подономин из бирманского янтаря (Giłka et al., 2020), для которого реконструируют совершенно другую палеообстановку. Поэтому пока преждевременно делать выводы о биологии нового рода, известного по одной куколке из одного местонахождения.
Для юрских отложений Сибири показано доминирование представителей подсемейства среди хирономид и по разнообразию, и по обилию находок (Калугина, Ковалев, 1985), но в Хасурты ситуация другая – найден только один экзувий куколки, относящийся к Podonominae, который описывается ниже.
Н.С. Калугина (Калугина, Ковалев, 1985) обнаружила значительное сходство между юрскими Podonominae и Tanypodinae и заключила, что эти два близких подсемейства были еще слабо разграничены, что позже было подтверждено нами (Lukashevich, Przhiboro, 2012, 2018). Новый род, по-новому сочетающий признаки обоих подсемейств, демонстрирует еще одно свидетельство этой близости.
Род Gilkania gen. nov.
Название рода – в честь польского диптеролога В. Гилки (W. Giłlka).
Типовой вид – Gilkania transbaikalica sp. nov.
Диагноз. Куколка среднего размера. Торакальные рога небольшие, узкие, пластрон, вероятно, маленький. Ножные чехлы встречаются у вершины крыловых чехлов; дистальные части чехлов передних и средних ног практически прямые, положение задних чехлов неясно. Брюшные сегменты I–VI прямоугольные, VII – трапециевидный, со слабо вытянутыми назад задними углами, самый массивный, VIII–IX округлые, VIII с сильно вытянутыми назад лопастевидными задними углами и сильно выгнутым задним краем и выпуклыми боковыми краями. Апофизы присутствуют на сегментах I–VII. Анальный плавник удлиненный, дистально разделен на две лопасти, без коротких щетинок на концах лопастей, анальные лопасти на вершине заострены. Латеральные щетинки LS на VII–VIII сегменте длинные, лишь слабо изогнутые, на VII сегменте их не меньше 2 пар, на VIII – 5 пар. Латеральных щетинок LS на IX сегменте три пары, две сближенные щетинки расположены в срединной части и одна – гораздо дистальнее; все они длинные, по крайней мере, две сближенные щетинки изогнуты. Все латеральные LS щетинки на VII–IX сегментах затемнены у основания.
Видовой состав. Типовой вид.
Сравнение. Новый род близок к мезозойскому Oryctochlus по форме и хетотаксии VIII и IX брюшных сегментов, но отличается от всех до сих пор известных подономин наличием мощных LS щетинок на сегменте VII.
Замечания. Наличие мощных LS щетинок на сегменте VII характерно для другого подсемейства, Tanypodinae, но там их не меньше трех пар, а обычно – больше. Пока нельзя исключить, что и у описываемого экзувия их больше, но с обеих сторон отчетливо видны лишь две пары. Наличие этих щетинок, несомненно, в данном случае является признаком родового уровня, поэтому, несмотря на неполную сохранность единственного экзувия, было решено выделить новый род. У современного Trichotanypus на задних углах сегмента VII есть пара щетинок, но они гораздо слабее и короче боковых щетинок на сегменте VIII и расположены по-другому (Brundin, 1986).
Систематическое положение некоторых мезозойских родов, известных только по куколкам (Jurochlus Kalugina, 1985, Langtonius Lukashevich et Przhiboro, 2018), остается непонятным, в связи с обнаруженной у них комбинацией подономинных и таниподинных черт (Lukashevich, Przhiboro, 2012, 2018). Однако куколка нового рода не похожа ни на один из этих родов, а своеобразная форма и хетотаксия ее VIII и IX сегментов практически неотличимы от подономины Oryctochlus, известной по всем жизненным стадиям (Калугина, Ковалев, 1985; Lukashevich, 2012). У Oryctochlus не отмечено никаких таниподинных черт, а длинные изогнутые латеральные щетинки с темными основаниями (т.н. черными зонами по Л. Брундину), обнаруженные на анальном плавнике нового рода, известны только у некоторых современных подономин, включая Trichotanypus, и у мезозойского Oryctochlus (Brundin, 1966; Lukashevich, 2012; Lukashevich, Przhiboro, 2012). На основании этого уникального признака, подкрепленного своеобразной формой и хетотаксией VIII и IX сегментов, новый род отнесен к подономинам.
Gilkania transbaikalica Lukashevich, sp. nov.
Табл. VII , фиг. 2
Название вида – от Забайкалья.
Голотип – ПИН, № 5340/2952±, позитивный и негативный отпечаток экзувия куколки хорошей сохранности; Хасурты; нижний мел.
Описание (рис. 2, ж). Сброшенный экзувий очень светлый, торакальные рога коричневые, в 3 раза длиннее своей ширины. Крыловые чехлы широкие, к вершине не сужающиеся, тупо закругленные, достигают заднего края брюшного сегмента II. Латеральные щетинки LS на сегменте VII расположены у заднего края и, вероятно, несколько тоньше щетинок сегмента VIII. Заднебоковые лопасти сегмента VIII занимают больше трети длины сегмента и достигают трети длины анального плавника. Латеральные щетинки LS занимают заднюю половину сегмента VIII, передняя щетинка расположена чуть раньше середины сегмента, все щетинки на равном расстоянии друг от друга. Анальный плавник вдвое уже сегмента VIII, с сильно расходящимися внутренними краями, без мелких зубчиков снаружи. Генитальные чехлы самки короче половины длины плавника, оканчиваются на уровне двух сближенных щетинок.
Размеры в мм: общая длина тела куколки 4.5, длина торакального рога 0.27, длина анальной лопасти 0.4, ширина 0.15.
Материал. Голотип.
ИНФРАОТРЯД BIBIONOMORPHA
СЕМЕЙСТВО ANISOPODIDAE KNAB, 1912
ПОДСЕМЕЙСТВО TEGINAE LUKASHEVICH, 2012
Замечания. Семейство Anisopodidae очень небольшое, включает всего 15 ныне живущих родов (Pape et al., 2011). Вымершее подсемейство Teginae было выделено как монотипное. Два близких вида своеобразного рода Tega Blagoderov et al., 1993 были описаны из юрских отложений Казахстана и Монголии и отнесены к современному семейству Cramptonomyiidae (Blagoderov et al., 1993). После переизучения типового материала Tega karatavica Blagoderov et al., 1993 из Каратау, T. penna Blagoderov et al., 1993 из Шар-Тэга (известно всего три отпечатка) и неописанного материала из других юрских отложений Азии, род был перенесен в Anisopodidae, в собственное подсемейство (Лукашевич, 2012а). Ниже описывается первый представитель подсемейства из меловых отложений. Как и в типовом роде, у него отмечены диагностические признаки подсемейства: R2 + 3 постепенно сближается с С, без сильного изгиба в дистальной трети, ветви Rs сильно сближены, впадают в С, медиана с четырьмя ветвями, дискоидальная ячейка очень маленькая, im в проксимальной половине крыла. Сперматеки не видны (вероятно, из-за сильной склеротизации брюшка).
Образ жизни личинок мезозойского подсемейства неизвестен, но во всех ныне живущих подсемействах Anisopodidae личинки – наземные сапрофаги, обитающие в разлагающейся органике (древесина, бродящий сок в натеках на стволах деревьев, плодовые тела древесных грибов).
Род Amorimyia Lukashevich, gen. nov.
Название рода – в честь бразильского диптеролога Д. Аморима (D. Amorim) и от myia греч. – муха; ж.р.
Типовой вид – Amorimyia robusta sp. nov.
Диагноз. Коренастые анизоподоиды среднего размера. Глаза самки широко расставлены, глазки не видны, задняя и нижняя часть головы опушена длинными щетинками. Антенны короткие, заметно короче грудного отдела и не длиннее ширины головы, скапус длиннее педицеллюса, жгутик 14-члениковый, все членики густо и равномерно усажены короткими щетинками. Грудь массивная, сильновыпуклая. Крыло широкое, заметно длиннее брюшка, крыловая мембрана густо и равномерно усажена микротрихиями, макротрихии расположены в несколько рядов на костальной жилке и в один ряд – на радиальных. Костальный край выпуклый, костальное поле расширено в районе плечевой жилки, sc-r постсекторальная. Костальная жилка С исчезает сразу за окончанием R4 + 5. Жилки Sc и R1 короткие, оканчиваются в проксимальной половине крыла, R2 + 3 – сразу за серединой крыла. Sc и радиальные жилки толще медиальных, кроме очень тонкой R2 + 3. Все три радиальные жилки (R1, R2 + 3, R4 + 5) упираются в С задолго до вершины крыла.
Волоски на теле, кроме головы, не видны. Брюшко коротко-цилиндрическое, менее чем в 1.5 раза длиннее головы с грудью, с наибольшей шириной на уровне сегментов II–III.
Видовой состав. Типовой вид.
Сравнение. Новый род отличается от типового для подсемейства крупными размерами (у юрских видов длина крыла 3 мм), густо опушенной головой, телом без опушения и крылом с выпуклой широкой костальной ячейкой, постсекторальным положением sc-r и утолщенными радиальными жилками.
Замечания. Очень тонкая R2 + 3 была обнаружена нами при переизучении типового материала у паратипа Tega karatavica (Лукашевич, 2012а, табл. Х, фиг. 9 ), хотя у голотипа этого вида толщина ветвей Rs практически не отличается. Так что пока вариабельность и вес этого признака остаются непонятными.
Судя по всему, редкое и длинное опушение головы не затрагивает глаза, т.к. ни в одной зоне с сохранившимися фасетками макротрихии не найдены (табл. VII , фиг. 3 а). Опушение глаз длинными волосками известно у одного современного рода анизоподид – чилийского Lobogaster Philippi, 1965 (Kania et al., 2019, табл. 1 ).
Amorimyia robusta Lukashevich, sp. nov.
Табл. VI , фиг. 5 ; табл. VII , фиг. 3
Название вида от robustus лат. – крепкий, массивный.
Голотип – ПИН, № 5340/2978±, позитивный и негативный отпечатки целой самки; Хасурты; нижний мел.
Описание (рис. 2, з). Самка. Тело и его придатки темные. Голова слабо поперечная. Жгутик антенны почти не суживается к вершине, его членики цилиндрические, слабо поперечные. Ротовые части и максиллярные щупики не видны. Крыло примерно в 2.5 раза длиннее своей ширины. Мембрана без пятен, кроме крупной темной птеростигмы. Rs длинный, занимает 80% длины крыла. Ствол М слабее, чем слабо расходящиеся М1, М2, М3, М4, в вершину крыла упирается М1. Поперечная im упирается прямо в развилок М1 + 2 почти на уровне окончания Sc. Дискоидальная ячейка занимает 1/13 длины крыла, широкая, длина лишь немного больше ширины. Отрезок крыла между окончаниями М3 и М4 незначительно короче отрезка между окончаниями М2 и М3. Поперечная m-cu упирается прямо в развилок М3 + 4. Ноги сильные, бедра, по крайней мере, задние, равномерно расширены, достигают середины брюшка. Голени тоньше бедер, вершины голеней не видны. Церки очень короткие.
Размеры в мм: длина тела 7.4, крыла 7.0, груди 2.2, брюшка 4.2, головы 1.0; ширина головы 1.0; длина антенны 0.9, заднего бедра 1.5.
Материал. Голотип.
Список литературы
Калугина Н.С. Новые психодоморфные двукрылые насекомые мезозоя Сибири (Diptera: Eoptychopteridae, Ptychopteridae) // Палеонтол. журн. 1989. № 1. С. 65–77.
Калугина Н.С. Хаобориды и комары-звонцы из мезозоя Восточного Забайкалья (Diptera: Chaoboridae, Chironomidae) // Тр. Палеонтол. ин-та РАН. 1993. Т. 252. С. 117–139.
Калугина Н.С., Ковалев В.Г. Двукрылые насекомые юры Сибири. М.: Наука, 1985. 198 с.
Кривошеина Н.П. Определительная таблица родов ксило-микобионтных личинок Limoniidae и Pediciidae (Diptera) // Russ. Entomol. J. 2011. № 20 (1). С. 81–100.
Лин Ц., Лукашевич Е.Д. Proptychopterina (Diptera: Eoptychopteridae) из юры Северо-Восточного Китая // Палеонтол. журн. 2006. № 3. С. 56–60.
Лукашевич Е.Д. Ptychopteridae (Insecta: Diptera): история изучения и объем семейства // Палеонтол. журн. 2008. № 1. С. 68–77.
Лукашевич Е.Д. Новые бибиономорфные двукрылые юры Азии (Insecta: Diptera) // Палеонтол. журн. 2012а. № 3. С. 52–64.
Лукашевич Е.Д. Филогения Ptychopteroidea (Insecta: Diptera) // Палеонтол. журн. 2012б. № 5. С. 32–40.
Макарченко Е.А. Хирономиды Дальнего Востока СССР. Подсемейства Podonominae, Diamesinae и Prodiamesinae (Diptera, Chironomidae). Владивосток: ДВНЦ АН СССР, 1985. 200 с.
Савченко Е.Н. Комары-лимонииды фауны СССР: Определитель надвидовых таксонов с каталогизированным обзором видов. Киев: Наук. думка, 1989. 380 с.
Alexander C.P. Crane-flies of the Baltic amber (Diptera) // Bernstein-Forschungen. 1931. V. 2. P. 1–135.
Alexander C.P. New or insufficiently-known crane-flies from Chile (Family Tipulidae, Order Diptera). Part V // Agricultura Tecnica. 1953. V. 13. № 1. P. 1–23.
Alexander C.P. New or little known Tipuloidea (Diptera) from Argentina. II // Acta Zool. Lilloana. 1962. V. 18. P. 335–358.
Alexander C.P., Byers G.W. Tipulidae // Manual of Nearctic Diptera. Vol. 1 / Eds. McAlpine J.F. et al. Ottawa, 1981. P. 153–190 (Res. Branch Agric. Canada Monogr. № 27).
Bode A. Die Insektenfauna des ostniedersächsischen Oberen Lias // Palaeontogr. A. 1953. Bd 103. H. 1–4. S. 1–375.
Blagoderov V.A., Krzemińska E., Krzemiński W. Fossil and recent Anisopodomorpha (Diptera, Oligoneura): family Cramptonomyiidae // Acta Zool. Cracov. 1993. V. 35. P. 573–579.
Blagoderov V.A., Lukashevich E.D. New Axymyiidae (Insecta: Diptera) from the Mesozoic of east Siberia // Pol. Pis. Entomol. 2013. V. 82. P. 257–271.
Brundin L. Transantarctic relationships and their significance, as evidenced by chironomid midges with a monograph of the subfamilies Podonominae and Aphroteniinae and the austral Heptagyiae // Kungl. Sven. Vetenskapsakad. Handl., Fjarde Ser. 1966. V. 11. P. 1–472.
Brundin L. The pupae of Podonominae (Diptera: Chironomidae) of the Holarctic Region—Keys and diagnoses // Chironomidae of the Holarctic Region: Keys and Diagnoses: Part 2. Pupae / Ed. Wiederholm T. P. 19–30 (Entomol. Scand. 1986. Suppl. № 28).
Carpenter F.M. Treatise on Invertebrate Palaeontology. Part R. Arthropoda 4. V. 3 & 4: Superclass Hexapoda. Boulder, Lawrence: Geol. Soc. America; The Kansas Geol. Surv., 1992. 655 p.
Cranston P.S., Hardy N.B., Morse G.E. et al. When molecules and morphology concur: the ‘Gondwan’ midges (Diptera: Chironomidae) // Syst. Entomol. 2010. V. 35. P. 636–648.
Dong F., Shih C., Skibińska K. et al. New species of Tanyderidae (Diptera) from the Jiulongshan Formation of China // Alcheringa: Austral. J. Palaeontol. 2015. V. 39. № 4. P. 494–507.https://doi.org/10.1080/03115518.2015.1041308
Eskov K.Y., Lukashevich E.D. On the history of ranges of two relict nematoceran families, Ptychopteridae and Tanyderidae (Insecta: Diptera): a biogeographical puzzle // Russ. Entomol. J. 2015. V. 24. P. 257–270.
Handlirsch A. Die fossilen Insekten und die Phylogenie der rezenten Formen. Leipzig: Engelmann, 1906–1908. 1430 S.
Gao O., Shin C., Kopec K. et al. New species and revision of Pediciidae (Diptera) from the Middle Jurassic of northeastern China and Russia // Zootaxa. 2015. V. 3963. № 2. P. 240–249.
Giłka W., Zakrzewska M., Makarchenko E.A. Burmochlus gen. nov., the first Cretaceous member of Archaeochlus cluster (Diptera: Chironomidae: Podonominae) // Cret. Res. 2020 V. 106.https://doi.org/10.1016/j.cretres.2019.104261
Greenwalt D.E., Blagoderov V.A. Review of the fossil record of Bolitophilidae, with description of new taxa and discussion of position of Mangas Kovalev (Diptera: Sciaroidea) // Zootaxa. 2019. V. 4567. № 3. P. 546–460.
Kania I., Wojtoń M., Lukashevich E. et al. Anisopodidae (Insecta: Diptera) from Upper Cretaceous amber of northern Myanmar // Cret. Res. 2019. V. 94. P. 190–206. https://doi.org/10.1016/j.cretres.2018.10.013
Krzemiński W. The Oligocene Tipulomorpha (Diptera) from Bolshaya Svetlovodnaya (Eastern Asia, Russia) // Pol. Pis. Entomol. 2000. V. 69. P. 239–245.
Krzemiński W., Kania I., Lukashevich E. The first South American record of fossil Eoptychopterinae (Ptychopteridae, Diptera) from Lower Cretaceous Santana Formation // Cret. Res. 2015. V. 52. P. 548–555.
Lindegaard C. Chironomidae (Diptera) of European cold springs and factors influencing their distribution // J. Kansas Entomol. Soc. Suppl. 1995. V. 68. P. 108–131.
Lukashevich E.D. New Eoptychopteridae (Diptera: Psychodomorpha) from the Jurassic of Asia // Paleontol. J. 1993. V. 27. № 1A. P. 103–123.
Lukashevich E.D. First pupae of the Eoptychopteridae and Ptychopteridae from the Mesozoic of Siberia (Insecta: Diptera) // Paleontol. J. 1995. V. 29. № 4. P. 164–170.
Lukashevich E.D. On the systematic position of Prodocidia (Diptera) from the Lower Lias of England // Paleontol. J. 2000. V. 34. Suppl. № 3. P. 352–354.
Lukashevich E.D. Limoniidae (Diptera) in the Upper Jurassic of Shar Teg, Mongolia // Zoosymposia. 2009. V. 3. P. 131–154.
Lukashevich E.D. Pupae of Mesozoic Oryctochlus Kalugina, 1985 (Chironomidae: Podonominae), with description of two new species // Proc. of the 18th Intern. Symp. on Chironomidae. Fauna norvegica. 2012. V. 31. P. 159–165.
Lukashevich E.D. Male terminalia and their rotation in Tanyderidae (Diptera, Nematocera) since the Mesozoic // Histor. Biol. 2020. V. 32. № 4. P. 462–475. https://doi.org/10.1080./08912963.2018.1500568
Lukashevich E.D. Review of the fossil record of Bittacomorphinae (Diptera: Ptychopteridae) // Zootaxa. 2019. V. 4461. № 3. P. 566–578.https://doi.org/10.11646/zootaxa.4661.3.9
Lukashevich E.D., Ansorge J., Krzemiński W., Krzemińska E. Revision of Eoptychopterinae (Diptera: Eoptychopteridae) // Pol. Pis. Entomol. 1998. V. 67. P. 311–343.
Lukashevich E., Arillo A. New Eoptychoptera (Insecta: Diptera, Ptychopteridae) from the Lower Cretaceous of Spain // Cret. Res. 2016. V. 58. P. 254–264.
Lukashevich E.D., Azar D. First Eoptychopteridae (Insecta: Diptera) from Early Cretaceous Lebanese amber // Acta Zool. Cracov. 2003. V. 46 (Suppl.–Fossil Insects). P. 195–204.
Lukashevich E., Blagoderov V. Review of Mesozoic Perissommatidae // Zootaxa. 2020. V. 4718. № 4. P. 481–496. https://doi.org/10.11646/zootaxa.4718.4.3
Lukashevich E.D., Coram R.A., Jarzembowski E.A. New true flies (Insecta: Diptera) from the Lower Cretaceous of southern England // Cret. Res. 2001. V. 22. P. 451–460.
Lukashevich E.D., Grimaldi D. Eoptychopteridae (Insecta: Diptera) in Cretaceous amber from Myanmar // Stud. Dipterol. 2004. V. 10. № 2. P. 359–366.
Lukashevich E., Krzemiński W. New Jurassic Tanyderidae (Diptera) from Asia with first find of larvae // Zoosymposia. 2009. V. 3. P. 155–172.
Lukashevich E.D., Pepinelli M., Currie D.C. A new family and genus of Cretaceous nematocerous flies (Insecta: Diptera) // J. Syst. Palaeontol. 2020. V. 18. № 3. P. 213–231.https://doi.org/10.1080/14772019.2019.1588796
Lukashevich E.D., Przhiboro A.A. Pupae of Mesozoic Jurochlus Kalugina, 1985 (Diptera: Chironomidae), with description of four new species // Zootaxa. 2012. V. 3478. P. 434–452.
Lukashevich E.D., Przhiboro A.A. A new genus of Chironomidae (Insecta: Diptera) from the Lower Cretaceous of Mongolia // Paleontol. J. 2018. V. 52. № 12. P. 1401–1407.
Lukashevich E.D., Ribeiro G.C. Mesozoic fossils and the phylogeny of Tipulomorpha (Insecta: Diptera) // J. Syst. Palaeontol. 2019. V. 17. № 8. P. 635–652. https://doi.org/10.1080/14772019.2018.1448899
Lukashevich E.D., Shcherbakov D.E. First description of Tanyderidae (Diptera) larvae from South America // Russ. Entomol. J. 2014. V. 23. № 2. P. 121–138.
Oosterbroek P., Theowald B. Phylogeny of the Tipuloidea based on characters of larvae and pupae (Diptera, Nematocera) with an index to the literature except Tipulidae // Tijdshcr. Entomol. 1991. V. 134. P. 211–267.
Makarchenko E.A., Endo K. The description of immature stages of Linevitshia Makarchenko and Sasayusurika Makarchenko (Diptera: Chironomidae, Diamesinae), with some remarks on taxonomy and systematics of these genera // Euroasian Entomol. J. 2009. V. 8. P. 64 –70.
Pape T., Blagoderov V., Mostovski M.B. Order Diptera Linnaeus, 1758 // Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness / Ed. Zhang Z.-Q. Zootaxa. 2011. V. 3148. P. 222–229.
Podenas S. Paradelphomyia (Diptera, Limoniidae) from the Baltic amber // Acta Zool. Lituanica. Biodiversity. 1999. V. 9. № 3. P. 107–110.
Podenas S. New species of Tricyphona crane flies (Diptera: Pediciidae) from Baltic amber (Eocene) // Proc. Acad. Natur. Sci. Philad. 2001. V. 151. P. 31–39.
Shcherbakov D.E., Lukashevich E.D., Blagoderov V.A. Triassic Diptera and initial radiation of the order // Intern. J. Dipter. Res. 1995. V. 6. № 2. P. 75–115.
Szadziewski R., Krynicki V.E., Krzemiński W. The latest record of the extinct subfamily Eoptychopterinae (Diptera: Ptychopteridae) from Upper Cretaceous amber of North Carolina // Cret. Res. 2018. V. 82. P. 147–151.
Whalley P.E.S. The systematic and palaeogeography of the Lower Jurassic insects of Dorset, England // Bull. Brit. Museum Natur. Hist. (Geol.) 1985. V. 39. № 3. P. 107–189.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал