

Палеонтологический журнал, 2021, № 2, стр. 88-98
Морфология чешуйного покрова и стратиграфическое распространение Isadia arefievi A. Minich (Actinopterygii, Eurynotoidiformes)
А. С. Бакаев a, *, В. В. Буланов a, b, **
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Казанский (Приволжский) федеральный университет
Казань, Россия
* E-mail: alexandr.bakaev.1992@mail.ru
** E-mail: bulanov@paleo.ru
Поступила в редакцию 11.05.2020
После доработки 29.06.2020
Принята к публикации 29.06.2020
Аннотация
В верхнепермских отложениях Восточной Европы обнаружен новый морфотип чешуй лучеперых рыб отряда Eurynotoidiformes (Actinopterygii), относящийся к ранее описанному на основе изолированных зубов виду Isadia arefievi A. Minich, 2015. Чешуи, определяемые как I. arefievi, обнаружены в пяти местонахождениях (включая типовое), приуроченных к верхневятскому интервалу, в том числе к терминальнопермским отложениям жуковского горизонта (зона Archosaurus rossicus), охарактеризованных вязниковским фаунистическим комплексом тетрапод. Строение чешуйного покрова I. arefievi морфологически близко к таковому I. opokiensis A. Minich et Andrushkevich, 2017, что подтверждает предположение о принадлежности указанных видов одной филогенетической линии (I. opokiensis–I. arefievi). Новые находки I. arefievi свидетельствуют о широком распространении в водных сообществах вятского времени растительноядных рыб с соскребающим типом питания.
ВВЕДЕНИЕ
Eurynotoidiformes Minich et A. Minich, 1990 – эндемичный для Восточной Европы отряд лучеперых рыб, динамично эволюционировавший в пределах данного региона на протяжении средней–поздней перми, и исчезновение которого связано с кризисным пермо-триасовым событием. Интерес к данной группе продиктован трофической специализацией ее уржумско-вятских представителей, которые являются древнейшими известными облигатно-растительноядными рыбами, предположительно, уже к концу перми освоившими основные типы питания растительной биомассой, встречаемые у современных костистых рыб (Esin, 1997; Буланов, Миних, 2017; Бакаев, 2019; Pindakiewicz et al., 2020). Широкое распространение эвринотоидиформ в пределах Восточно-Европейской платформы (Миних, Миних, 1990; Есин, Машин, 1996), а также видоспецифичное строение их зубов и чешуй, легко идентифицируемых в комплексах изолированных ихтиолитов, определяют большой потенциал группы для ее использования в целях региональной стратиграфии.
Верхи северодвинского и вятский ярусы Восточной Европы сравнительно хорошо охарактеризованы ихтиофауной (Миних, Миних, 2009), в том числе остатками четырех видов эвринотоидиформ, относимых к роду Isadia: I. suchonensis A. Minich, 1986, I. aristoviensis A. Minich, 1990, I. opokiensis A. Minich et Andruskevich, 2017 и I. arefievi A. Minich, 2015. Для I. suchonensis и I. aristoviensis известно как строение зубов, так и чешуй (Миних, Миних, 2009). Isadia opokiensis описана по изолированным чешуям из местонахождения Мутовино (Миних, Андрушкевич, 2017), однако недавно из изохронного и географически близкого местонахождения Марьюшкина Слуда-С были описаны также и зубы, относимые к данному виду (Бакаев, 2020).
Вид I. arefievi описан только по изолированным зубам характерной формы (Миних и др., 2015), сочетающим мощный контрфорс с сильно расширенной коронкой, снабженной несколькими (до семи) высокими, широкими, полностью образованными акродином зубцами, некоторые из которых приобрели секировидную форму. Данные о строении чешуйного покрова вида до настоящего времени не публиковались.
В предлагаемой работе описывается комплекс чешуй, по нашему мнению, относящийся к I. arefievi и происходящий из пяти верхневятских местонахождений центра и северо-востока Восточно-Европейской платформы (рис. 1; табл. 1): Элеонора (типовое), Раша, Жуков Овраг-1-А, Сартаково и Лагерный Овраг-1. В трех первых местонахождениях чешуи нового морфотипа обнаружены вместе с зубами I. arefievi, имеющими хорошо узнаваемую форму (рис. 2). Эта совместная встречаемость, очевидное отличие от чешуй других описанных видов Isadia, а также наибольшее сходство с таковыми I. opokiensis, имеющей наиболее близкий (предположительно исходный для I. arefievi) тип строения зубов (Бакаев, 2020), убеждают в принадлежности описываемых ниже остатков к I. arefievi.
Рис. 1.
Расположение местонахождений с остатками Isadia arefievi А. Minich, 2015: 1 – Элеонора; 2 – Раша; 3 – Лагерный Овраг-1, Лагерный Овраг-3 и Сартаково; 4 – Жуков Овраг-1-А; 5 – Соковка.
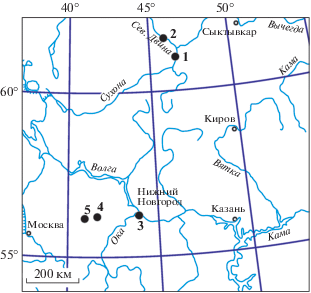
Таблица 1.
Список местонахождений с остатками Isadia arefievi A. Minich (данные по местонахождению Соковка из: Pindakiewicz et al., 2020).
| Местонахождение | Регион | Стратиграфическое положение | Чешуи | Зубы |
|---|---|---|---|---|
| Элеонора | Вологодская обл. | нефедовский горизонт | + | + |
| Раша | Архангельская обл. | нефедовский горизонт | + | + |
| Лагерный Овраг-3 | Нижегородская обл. | нефедовский горизонт | – | + |
| Сартаково | Нижегородская обл. | нефедовский горизонт | + | – |
| Жуков Овраг-1-А | Владимирская обл. | жуковский горизонт | + | + |
| Лагерный Овраг-1 | Нижегородская обл. | жуковский горизонт | + | – |
| Соковка | Владимирская обл. | жуковский горизонт | – | + |
Рис. 2.
Зубы Isadia arefievi А. Minich, 2015 из различных местонахождений: а – экз. ПИН, № 5652/400, лингвально; б – экз. ПИН, № 5157/401, латерально; в – экз. ПИН, № 5652/402, лингвально; г – экз. ПИН, № 5652/403, лабиально (Вологодская обл., Великоустюгский р-н, типовое местонахождение Элеонора; верхневятский подъярус, нефедовский горизонт, саларевская свита, комарицкая пачка); д – экз. ПИН, № 5652/300, лабиально (Владимирская обл., Гороховецкий р-н, мест. Жуков Овраг 1-А; верхневятский подъярус, нефeдовский горизонт, обнорская свита, гороховецкая пачка); е – экз. ПИН, № 5652/808, лингвально; ж – экз. ПИН, № 5157/809, лабиально; з – экз. ПИН, № 5652/810, лингвально (Архангельская обл., Карпогорский р-н, мест. Раша; верхневятский подъярус, саларевская свита); и – экз. ПИН, № 5652/604, лингвально; к – экз. ПИН, № 5652/606, лингвально (Нижний Новгород, мест. Лагерный Овраг-3; верхневятский подъярус, нефeдовский горизонт). Длина масштабной линейки 300 мкм.

В местонахождениях Сартаково и Лагерный Овраг-1 зубы I. arefievi пока не обнаружены, что, по нашему мнению, обусловлено либо ограниченной выборкой (Сартаково), либо фациальными особенностями захоронения (плохой сохранностью зубов в грубозернистых породах), а также относительно крупным размером просмотренных фракций, в которых мелкие ихтиолиты обычно не представлены (Лагерный Овраг-1).
Рассматривая вопрос о распространении вида, следует отметить, что ранее присутствие I. arefievi было установлено по находкам отдельных зубов в терминально-пермском местонахождении Соковка (Pindakievicz et al., 2020); кроме того, при первоописании вида к нему был отнесен также изолированный зуб из местонахождения Лагерный Овраг-3, хотя и отмечалось его не вполне стандартное строение (Миних и др., 2015). Зубы, найденные нами в том же местонахождении (рис. 2, и, к), в целом укладываются в диапазон морфологической изменчивости выборки зубов из Элеоноры, что подтверждает присутствие I. arefievi в средней части пермских отложений, представленных в разрезе Лагерный Овраг–Окский Съезд.
В большинстве местонахождений остатки I. arefievi происходят из грубозернистых отложений руслового аллювия (пески, гравелиты и песчаники), что в целом определяет лишь удовлетворительную сохранность чешуй, многие из которых достаточно сильно окатаны и сохранили ганоиновые структуры не на всем свободном поле. Местонахождения Элеонора и Раша приурочены к старично-русловым фациям (обогащенные органикой глины и алевролиты), большинство встреченных в них чешуй также сильно истерты при переносе или, не исключено, локальном переотложении.
Несмотря на это, новый материал в совокупности позволяет впервые и достаточно полно охарактеризовать чешуйный покров различных участков тела I. arefievi, что расширяет морфологическую характеристику вида и в дальнейшем позволит уверенно идентифицировать его по изолированным чешуям в других аллохтонных ориктоценозах. Полученные данные указывают на значительно более широкое распространение I. arefievi в вятское время на территории Восточно-Европейской платформы, а с учетом трофических адаптаций вида (Бакаев, 2020) – на распространенность растительноядных рыб с соскребающим типом питания в водных сообществах соответствующего временного интервала, а точнее – во второй половине поздневятского времени.
Действительно, для большинства перечисленных местонахождений ранее убедительно продемонстрирован поздневятский возраст: они хорошо позиционируются в разрезах центра и северо-востока Европейской России, в которых вскрыт комплекс пограничных отложений перми и триаса. Местонахождения Соковка, Жуков Овраг-1-А (Владимирская обл.) и Лагерный Овраг-1 (Нижний Новгород) приурочены к жуковскому горизонту (тетраподная комплексная зона Archosaurus rossicus) и охарактеризованы фауной тетрапод вязниковского фаунистического комплекса; помимо этого, их терминально-пермское положение убедительно продемонстрировано на основе других стратиграфически значимых групп (остракоды, конхостраки), литостратиграфических и палеомагнитных данных (Голубев, 2004; Сенников, Голубев, 2010; Голубев и др., 2012; Scholze et al., 2015, 2019; Балабанов и др., 2016; Наумчева, Голубев, 2019).
Недавно открытое местонахождение Элеонора (Вологодская обл.) расположено в верхней части разреза саларевской свиты (комарицкая пачка), вскрытой в бассейне р. М. Северная Двина, и в данном районе является наиболее молодым уровнем, охарактеризованным фауной пермских позвоночных (Арефьев и др., 2014; Миних и др., 2015; Миних, Миних, 2018). По количеству остатков тетрапод ориктоценоз местонахождения сравнительно беден, но содержит определимые микроостатки, на основе которых по отдельным зубам был описан наиболее специализированный вид рода Isadia – I. arefievi (Миних и др., 2015). Отсутствие среди тетрапод типичных видов-маркеров вязниковской фауны – Archosaurus rossicus (Proterosuchidae), Uralerpeton tverdochlebovae и Bystrowiana permira (Chroniosuchia), а также находка остатков крупноразмерных Pareiasauria, в целом не типичных для терминальной перми региона, но составлявших основу доминантного сообщества соколковского комплекса (Ивахненко, 1990, 2001; Ивахненко и др., 1997), позволило с оговорками отнести тетраподную ассоциацию из Элеоноры к соколковской фауне (зона Chroniosuchus paradoxus), отметив, таким образом, верхний репер стратиграфического распространения последней в данном разрезе (Миних и др., 2015).
В пользу этого мнения говорит значительная (ок. 25 м) толща перекрывающих Элеонору красноцветных отложений саларевской свиты, по биостратиграфическим и литологическим данным имеющей пермский возраст и бронируемой в разрезе Балебиха триасовыми конгломератами (Arefiev et al., 2015).
На более древний возраст Элеоноры по сравнению с известными местонахождениями жуковского горизонта указывают и новые данные по остракодам. Верхневятские отложения Московской синеклизы по остракодам подразделены на три толщи, последовательно сменяющие друг друга снизу вверх по разрезу: слои с Suchonella typica (слои с комплексом остракод I), слои с Suchonella clivosa (слои с комплексом остракод II) и слои с Suchonella rykovi (слои с комплексом остракод III) (Наумчева, Голубев, 2019; Голубев и др., 2020). Местонахождение Элеонора приурочено к слоям с Suchonella clivosa, в то время как жуковскому горизонту соответствуют слои с S. rykovi. Согласно указанным работам, к слоям с S. clivosa, охватывающим верхнюю часть нефедовского горизонта (Голубев и др., 2020), относится и местонахождение Лагерный Овраг-3 (обн. 1135, слои 23–29: Наумчева и др., 2017; Наумчева, Голубев, 2019). Однако это местонахождение приурочено к магнитозоне прямой полярности n1R3P (Балабанов и др., 2016) и является более древним, чем местонахождение Элеонора, которое расположено в магнитозоне обратной полярности r2R3P (Arefiev et al., 2015).
Местонахождение Раша не имеет точного позиционирования в разрезе саларевской свиты, однако содержит очень близкий к Элеоноре комплекс рыб (Isadia arefievi, I. aristoviensis, I. cf. suchonensis, Toyemia blumentalis, Geryonichthys sp., Elonichthyidae gen. indet., Palaeonisci indet.) и тетрапод (Lepospondyli ord. indet., Microphon cf. terminalis, Kotlassia prima, Karpinskiosaurus secundus, Pareiasauria gen. indet. и Chroniosuchus sp.). Более раннее определение в местонахождении на основе фрагментарного материала одной из форм вязниковского комплекса – Uralerpeton tverdochlebovae (Ивахненко и др., 1997; Голубев, 2000) – является ошибочным (В.К. Голубев, перс. сообщ.) и корректируется в настоящей работе. Имеющиеся фаунистические данные с учетом территориальной близости местонахождений, сходного типа вмещающих отложений и одинакового характера сохранности материала позволяют условно считать их одновозрастными до появления противоречащих данных.
Местонахождение Сартаково (Нижегородская обл.) открыто в 2003 г. (Манаева, Шумов, 2005). В 2004 г. здесь в глинисто-песчаных отложениях аллювиального генезиса, вскрывающихся в стенке оползня в средней части склона правого берега р. Ока, собраны многочисленные остатки остракод, двустворчатых моллюсков, а также остатки рыб и тетрапод. Собранный материал был передан И.С. Шумовым в Палеонтологический ин-т РАН (ПИН). Кроме I. arefievi, ихтиокомплекс местонахождения включает следующие виды (определения А.С. Бакаева): Toyemia blumentalis, Isadia aristoviensis, I. suchonensis, Mutovinia stella, Geryonichthys longus, Varialepis vitalii, Strelnia sp., Elonichthyidae gen. indet. Данный состав ихтиофауны соответствует вятскому возрасту, однако присутствие I. arefievi позволяет конкретизировать возраст местонахождения и отнести его к верхненефедовско–жуковскому интервалу, что подтверждается находками остракод вохминского горизонта нижнего триаса, которые были сделаны М.П. Арефьевым примерно в 21–22 м выше костеносных отложений (Сенников, Голубев, 2011).
Таким образом, по совокупности имеющихся данных диапазон стратиграфического распространения I. arefievi ограничен верхней частью верхневятского подъяруса (верхняя часть нефедовского и жуковский горизонты); исчезновение вида связано с экосистемными перестройками на рубеже перми и триаса.
МОРФОЛОГИЯ ЧЕШУЙНОГО ПОКРОВА ISADIA AREFIEVI A. MINICH
При описании чешуй Isadia arefievi была применена методика, предложенная Д.H. Есиным (1990), успешно использованная при изучении чешуйного покрова ряда ископаемых, в том числе базальных, лучеперых (Burrow, 1994; Trinajstic, 1999a, b; Choo, 2011; Chen et al., 2012; Qu et al., 2013; Choo et al., 2017; Cui et al., 2019; Bakaev, Kogan, 2020). Чешуйный покров большинства эвринотоидиформ хорошо охарактеризован, благодаря цельноскелетному материалу или значительным выборкам изолированных чешуй (Есин, 1995; Есин, Машин, 1996; Миних, Миних, 2009). Ниже дается общая характеристика строения чешуи эвринотоидиформ (применительно к боковым чешуям передней половины тела), отличающая их от других палеозойских лучеперых рыб.
Передний край прямой, нижний выпуклый, верхний симметрично вогнутый. Передневерхний угол высокий, заостренный. Передненижний угол сильно скошен, четко отделен от переднего края ступенькой, образующей заостренный выступ. Задненижний угол также скошен, отделен от заднего края ступенькой и несет зазубренность. Задневерхний угол округлый. Сочленовный шип и ямка хорошо развиты, медиальный гребень выражен слабо. Сочленовный шип был почти равен или равен по высоте передневерхнему углу. Киль всегда смещен вперед относительно шипа и ямки. Между килем и задним краем чешуи располагаются поры.
Языковидные выступы округлые или прямые. Ганоиновые гребни берут начало как от переднего, так и от верхнего края свободного поля. Верхние гребни (и борозды) направлены вентро-каудально; угол наклона постепенно выполаживается к нижнему краю чешуи, где они повторяют его контур. Свободное поле всегда несет вертикальные ряды зубцов (хорошо выраженных у самых передних чешуй и постепенно выполаживающихся при смещении назад).
Чешуи Isadia arefievi (табл. XI, XII , см. вклейку) толстые, свободное поле рельефное, сильно возвышается над погруженным, вдоль переднего и верхнего краев разделенное узкими, глубокими и сравнительно короткими бороздами на высокие гребни с каплевидно-округленными передними окончаниями. Вдоль переднего края, в бороздах между гребнями и зубцами, располагаются многочисленные поры, также изредка встречающиеся на гладком ганоиновом поле. Начиная с задней части участка “B” и далее в каудальном направлении, в центральной части чешуй появляется ровный, не рассеченный гребнями участок, гладкая поверхность которого может нести единичные ямки. В задней части ганоиновый слой распадается на множество отдельных зубцов, которые повреждены на всех чешуях выборки, но, судя по форме их оснований, имели стержневидную форму и довольно сильно выступали за пределы основания чешуи. На чешуях всех участков тела, за исключением, вероятно, наиболее задних чешуй участка “D”, располагавшихся в основании лопасти хвостового плавника, зубцы структурированы в каскадные образования.
Помимо очевидной топологической изменчивости чешуй, в изученной выборке проявляется онтогенетическая изменчивость. Так, более крупные чешуи отличаются от менее крупных чешуй из того же участка относительно менее широкими, но более многочисленными гребнями скульптуры и бóльшим количеством зубцов на заднем краю чешуи. Подобные различия укладываются в рамки онтогенетической изменчивости чешуйного покрова базальных актиноптеригий (Esin, 1995). В некоторых случаях чешуи с одного участка тела сходного размера могут существенно различаться (напр., табл. XI , фиг. 1, 2) по таким признакам, как число гребней скульптуры, число зубцов в задней части свободного поля. Подобные чешуи происходят из разных местонахождений, имеющих различный возраст, и различия могут показаться систематически значимыми и отражать филогенетическую историю таксона. Однако мы считаем, что подобные отличия объяснимы в рамках внутривидовой изменчивости. Так, хорошо известно, что многие современные лучеперые рыбы имеют выраженный, зависящий от среды обитания полиморфизм (Шаповалов, 2010; Senay et al., 2015; Nakayama et al., 2016), проявляющийся как в варьирующих размерах, так и в форме тела рыб, обитающих в различных условиях. Подобный полиморфизм чаще всего несет адаптивную функцию, и связан со скоростью течения, доступностью ресурсов, размерами водоема и так далее. Особенно хорошо изучено формирование быстро растущих пелагических, и медленно растущих литоральных форм (Senay et al., 2015). При одинаковом размере чешуя более молодых, но быстрее растущих рыб будет иметь меньшее число гребней и зубцов, чем чешуя более старых, но тугорослых рыб.
Ниже приводится составленное на изолированном материале описание чешуй различных участков тела I. arefievi. Топологическая изменчивость чешуйного покрова реконструирована на основе родственных таксонов, известных по целым скелетам (Kichkassia furkae, Lapkosubia uranensis, L. barbalepis, Isadia suchonensis, I. aristoviensis).
Участок “А” (табл. XII , фиг. 1, 2, 4 ). Чешуи этого участка в типовом местонахождении пока не обнаружены; в описании использованы образцы из других местонахождений. Высота чешуи примерно на 1/3 больше длины; длина погруженного поля почти равна длине свободного. У вентрально расположенных чешуй участка высота равна длине. Передний край прямой. Нижний сильновыпуклый, верхний сильновогнутый. Передневерхний угол высокий, округлый. Передненижний угол сильно скошен (занимает не менее половины высоты чешуи), почти прямой, переход в передний край в виде плавного закругления, граница находится немногим выше середины высоты свободного поля. Материал не позволяет реконструировать форму контура заднего края, а также форму задненижнего и задневерхнего углов у чешуй данного участка. Сочленовный шип и ямка высокие, расположены в середине длины чешуи. Сочленовный шип был почти равен или равен по высоте передневерхнему углу. Киль выпуклый, сильно смещен вперед из центрального положения. Между килем и задним краем чешуи располагаются поры.
Передний край свободного поля прямой. Ганоиновые гребни рельефно-выпуклые, имеют равномерно округленные каплевидные передние окончания и сужаются каудально. Ширина гребней вариабельна: самые широкие располагаются в передневерхнем углу поля. Борозды, разделяющие гребни, короткие, не более 1/3 длины свободного поля; каудально они слегка расширяются вследствие менее плотного смыкания гребней в центральной части чешуи, но не прорезают свободное поле насквозь, оканчиваясь вблизи оснований задних зубцов переднего ряда. Верхние гребни (и борозды) направлены вентро-каудально; угол наклона постепенно выполаживается к нижнему краю чешуи, где их ориентация почти горизонтальная. Каудальные зубцы организованы в несколько (не менее трех) каскадно расположенных рядов, основания которых находятся на разных уровнях и не сливаются с основаниями зубцов в соседних рядах. Основания зубцов переднего ряда выстроены вдоль вертикальной линии, проходящей по центру свободного поля и параллельной его переднему краю.
Участок “В” (табл. XI , фиг. 1–3; табл. XII , фиг. 3, 5 ). У передних чешуй участка высота на 20% больше длины; вблизи участка “C” длина чешуй равна высоте. Передний край прямой, реже – слабовыпуклый; задний – слабовыпуклый. Изгиб верхнего и нижнего краев варьирует: у передних чешуй участка они аналогичны таковым чешуй участка “А”; у чешуй, расположенных каудальнее, изгиб выполаживается. Передневерхний угол высокий, конец заострен. Передненижний угол скошен, прямой или немного закруглен, контакт с передним краем находится на уровне середины свободного поля или ниже, обычно округлeн. Задненижний край полого приподнят дорсо-каудально, часто вогнут. Киль резко очерченный, узкий, высокий. Сочленовный шип смещен из центрального положения к переднему краю чешуи, обычно ниже передневерхнего угла, рельефный у передних чешуй участка и плоский у задних; соответствующим образом меняется форма ямки.
Рис. 3.
Ультраскульптура ганоина чешуй тела Isadia arefievi А. Minich, 2015 на примере экз. ПИН, № 5652/404, чешуя участка “В”: а, б – участки ганоина под бóльшим увеличением. Длина масштабной линейки 0.3 мм

Погруженное поле занимает менее половины длины чешуи (у задних чешуй участка – 1/3 их длины). Передний край свободного поля почти прямой. Форма передних ганоиновых гребней варьирует в зависимости от размера чешуй (особей). У менее крупных экземпляров (табл. XII , фиг. 3) они менее многочисленны, почти плоские, более широкие, их форма и длина менее униморфны, контур передних окончаний неровный. У чешуй крупных особей гребни более многочисленные (до 10), резко выпуклые, относительно узкие, пальцевидные, четко отделенные бороздами, и имеют правильно-округленный передний край (табл. XI , фиг. 2). Гребни длиннее, чем на чешуях участка “А”, и занимают от 1/3 до 2/5 длины свободного поля. Наклон борозд и гребней в верхней части чешуи слабый, в средней и нижней частях – субгоризонтальный. Средняя треть свободного поля гладкая, не рассечена гребнями, содержит редкие ямки. Задние зубцы слабо наклонены вентрокаудально и, как видно на некоторых чешуях (табл. XI , фиг. 1), были длинными и имели стержневидную форму. На передних чешуях участка они образуют не менее трех каскадно расположенных рядов, на более задних чешуях – не менее двух.
Участок “С” (табл. XI , фиг. 4–7 ; табл. XII , фиг. 6–8 ). Пропорции чешуй меняются в зависимости от положения в пределах участка: в его начале длина чешуй равна высоте, ближе к участку “D” она в 1.5 раза больше высоты. Начиная с переднего края, высота чешуи равномерно увеличивается до линии оснований каудальных зубцов. Передневерхний угол невысокий, обычно заострeнный. Задненижний угол полого скошен, передненижний и задневерхний слабо изогнуты. Сочленовный шип и ямка маленькие или не выражены. Шип (при наличии) ниже передневерхнего угла. Киль выпуклый, у чешуй из задней части участка в виде вздутого бугра. Погруженное поле составляет около 1/3 длины чешуи. Передний край свободного поля прямой. Ганоиновые гребни рельефно выпуклые, ориентированы диагонально (в целом выстроены параллельно линии, идущей от передневерхнего угла к середине скошенного задненижнего фланга). Передние края гребней правильно округлые. Обычно гребни лежат плотно друг к другу, отделяясь узкими бороздами. На некоторых чешуях наблюдается слияние передних окончаний гребней и замыкание борозд в изолированные от внешнего края лакуны (табл. XI , фиг. 4 ); реже передний край свободного поля рассечен сильнее (табл. XII , фиг. 7 ). Длина борозд обычно составляет 1/3 длины свободного поля; заднюю четверть поля занимают зубцы, образующие от двух до четырех каскадных рядов.
Участок “D” (табл. XI , фиг. 8, 9 ; табл. XII , фиг. 9–11 ). Чешуи ромбической или прямоугольной формы. Длина превышает высоту в полтора–два раза. Погруженное поле составляет не более 1/5 длины чешуи. Передний и верхний края свободного поля очень ровные. Гребни резко выпуклые, сужаются каудально и разделены узкими, но глубокими щелями; по длине они занимают около трети свободного поля. Центральная часть свободного поля гладкая. У самых каудальных чешуй участка гребни почти полностью сливаются, и свободное поле представляет собой ровную поверхность (табл. XI , фиг. 9 ). Каудальные зубцы организованы в два ряда у всех чешуй участка, включая таковые в основании хвостового плавника; зона развития зубцов узкая. Шип и ямка не выражены, киль в виде валика.
Участок “F” (табл. XI , фиг. 11 ; табл. XII , фиг. 13, 14 ). Умеренно и сильно вытянутые чешуи (длина в 2–3 раза превышает высоту). Передневерхний угол очень низкий, в зависимости от положения чешуи на теле – короткий или вытянутый, заострeнный или округленный. Остальные углы сглажены. Сочленовный шип и ямка выражены слабо или отсутствуют. Погруженное поле хорошо развито у всех чешуй участка, обычно округленное на переднем конце (за исключением самых узких чешуй). Свободное поле занимает около половины длины чешуи. Скульптура из немногочисленных, коротких и мощных гребней, сильно приподнятых над остальными структурами свободного поля. Сглаженный участок в центральной части поля имеется. Каудальные зубцы расположены в два ряда, некоторые несут следы прижизненного стирания о грунт. Киль в виде вздутого бугра. Ямка широкая, но пологая (табл. XI , фиг. 14 ), расположена снизу и сзади от бугра.
Участок “G” (табл. XI , фиг. 10 ). Мелкие чешуи субквадратной формы с округленными углами. Передний край чешуи округлый, верхний – слегка выпуклый, нижний – слабо вогнутый. Задний край чешуи заужен. Погруженное поле очень широкое вдоль переднего края чешуи и едва развито вдоль верхнего. Шип и ямка отсутствуют. Свободное поле высоко приподнято над основанием чешуи; его поверхность гладкая, гребни на переднем краю отсутствуют или едва намечаются. Каудальные зубцы длинные, стержневидные, немногочисленные (пять–шесть), направлены назад и образуют только один ряд.
Чешуи у основания грудного плавника (табл. XII , фиг. 12 ). Небольшие чешуи округло-угловатой формы с хорошо развитым серповидно-изогнутым (симметричным или почти симметричным относительно длинной оси) погруженным полем. Свободное поле высокое, с округлым передним краем, рассеченным протяженными бороздами на немногочисленные рельефные гребни, которые по направлению назад не сливаются и переходят в каудальные зубцы.
Коньковые чешуи. Единственная обнаруженная коньковая чешуя (экз. ПИН, № 5677/2; табл. XII , фиг. 15 ), судя по ширине, наиболее вероятно, располагалась впереди от спинного плавника. Она максимально расширена в конце первой трети, имеет хорошо развитое, округло-вогнутое по переднему краю погруженное поле. Передний край свободного поля также вогнут. Передняя треть свободного поля рассечена изогнутыми бороздами на большое количество (13) гребней различной длины, ширина которых меняется по простиранию; гребни сравнительно невысокие. Центральная часть свободного поля гладкая. Заднебоковые края чешуи повреждены, но, вероятно, несли зазубренность того же типа, что и чешуи боковых сторон тела.
Ультраскульптура. Свободное поле чешуй морфотипа I. arefievi покрыто мелкими (менее 10 мкм в диаметре) бугорками (рис. 3). Бугорки на плоской поверхности в центральной части свободного поля имеют округлую форму (рис. 3, в), а расстояние между вершинами соседних бугорков составляет примерно 10 мкм. Однако на переднем крае гребней свободного поля бугорки имеют вытянутую форму (рис. 3, б), и тем длинней, чем на менее ровной поверхности они расположены. Так, наиболее удлинeнные бугорки расположены у самого переднего края ганоинового поля, а наиболее округлые – ближе к его центральной части. Это же наблюдается на чешуях других участков тела.
Тип ультраскульптуры ганоина I. arefievi (терминология по: Märss, 2006) характерен для базальных лучепeрых рыб (Reissner, 1859; Ermin et al., 1971; Schultze, 1966, 1968, 1977, 2016, 2018; Bakaev, Kogan, 2020), у которых бугорки соответствуют центрам клеток внутреннего слоя эпидермиса, и расстояние между которыми также составляет 10 мкм (т.е., соответствует размеру клеток; Märss, 2006). Различия в форме бугорков ультраскульптуры у I. arefievi отражают своеобразный процесс роста ганоина, отличный от роста настоящей эмали лопастепeрых рыб (Sire et al., 1987; Schultze, 2016). Вероятно, эпидермальные клетки были более растянуты на неровных частях скульптуры (на гребнях, около поровых отверстий и т.д.) и менее растянуты на ровных участках центральной части чешуи. Таким образом, форма бугорков отражает только форму эпидермальных клеток и их расположение, что позволяет сомневаться в значимости этого признака для систематики отряда.
Сравнение. Имеющаяся выборка изолированных чешуй из различных местонахождений в совокупности позволяет указать диагностические признаки Isadia arefievi в строении чешуйного покрова. От всех видов рода Isadia этот вид отличается: 1 – каскадным расположением зубцов на заднем краю чешуи на большинстве участков боковой стороны тела, до участка “D” включительно, а также (за исключением I. opokiensis): 2 – большей массивностью чешуй вследствие утолщения изопединовой и ганоиновой пластин, 3 – большей рельефностью киля, ямки и зубца, 4 – отсутствием килевидных образований вдоль осевой зоны ганоиновых гребней, и 5 – наличием гладкого поля в средней части чешуи на участках тела позади участка “А”. Дополнительно от I. opokiensis отличается: 1 – каплевидной (менее клиновидной), более выпуклой и резко очерченной формой передних окончаний ганоиновых гребней, 2 – меньшей длиной гребней, 3 – меньшим числом изолированных внутри ганоинового поля (не связанных с внешним краем) ямок и борозд, вследствие менее полного слияния в онтогенезе первичных гребней, и 4 – характером зазубренности заднего и нижнего краев чешуй, а именно – большим числом и меньшей упорядоченностью зубцов; дополнительно от I. aristoviensis отличается: 1 – меньшей длиной гребней, 2 – стержневидной формой и бóльшим количеством зубцов задне-нижнего обрамления чешуи, а от I. suchonensis: 1 – более выраженным разделением на гребни переднего края свободного поля, 2 – более суженными у основания каудальными зубцами.
По строению чешуй I. arefievi наиболее близок к I. opokiensis массивностью чешуй, рельефностью структур свободного поля и внутренней поверхности чешуи, формой ганоиновых гребней и зубцов, обрамляющих задний и нижний края чешуй. Полученные данные подтверждают точку зрения о филогенетической преемственности указанных видов, основанную на морфологическом сходстве челюстных зубов (Бакаев, 2020).
* * *
Авторы благодарны М.А. Наумчевой, И.С. Шумову и У.И. Карасeвой за сбор и подготовку (отмывку, просеивание и отбор из образцов породы) ценного сравнительного материала, В.К. Голубеву за стратиграфическую информацию. Данное исследование осуществлено при поддержке грантов РФФИ №№ 17-04-01937, 19‑34-90040 и 21-54-10003.
Список литературы
Арефьев М.П., Голубев В.К., Кухтинов Д.А. и др. Новое местонахождение остатков позднепермских животных и растений на Малой Северной Двине: вязниковский уровень? // Палеострат-2014. Тез. докл. годичн. собр. (науч. конф.) секции палеонтологии МОИП и московского отд. Палеонтол. об-ва при РАН. М.: ПИН РАН, 2014. С. 8–9.
Бакаев А.С. Новый морфотип зубов пермских рыб отряда Eurynotoidiformes // Палеострат-2019. Тез. докл. годичн. собр. (науч. конф.) секции палеонтологии МОИП и московского отд. Палеонтол. об-ва при РАН. М.: ПИН РАН, 2019. С. 11–12.
Бакаев А.С. Новый морфотип зубов рыб отряда Eurynotoidiformes (Actinopterygii) из верхнепермских отложений Европейской России // Палеонтол. журн. 2020. № 2. С. 78–86.
Балабанов Ю.П., Фетисова А.М., Голубев В.К., Сенников А.Г. Палеомагнитная и палеонтологическая характеристика пограничных отложений перми и триаса юга Московской синеклизы // Общая стратиграфическая шкала и методические проблемы разработки региональных стратиграфических шкал России. Матер. Межвед. рабоч. совещ. (С.-Петербург, 17–20 окт. 2016 г.). СПб.: Изд-во ВСЕГЕИ, 2016. С. 18–20.
Буланов В.В., Миних А.В. Конвергентный морфогенез зубной системы тетрапод и лучеперых рыб средней–поздней перми Восточной Европы // Тез конф. “Морфогенез в индивидуальном и историческом развитии: онтогенез и формирование биологического разнообразия”. М.: ПИН РАН, 2017. С. 9–11.
Голубев В.К. Пермские и триасовые хрониозухии и биостратиграфия верхнетатарских отложений Восточной Европы по тетраподам. М.: Наука, 2000. 174 с. (Тр. Палеонтол. ин-та РАН. Т. 276).
Голубев В.К. Граница перми и триаса на Восточно-Европейской платформе // Докл. Всеросс. совещ. “Структура и статус Восточно-Европейской стратиграфической шкалы пермской системы, усовершенствование ярусного расчленения верхнего отдела пермской системы общей стратиграфической шкалы”. Казань: КГУ, 2004. С. 19–21.
Голубев В.К., Миних А.В., Балабанов Ю.П. и др. Опорный разрез перми и триаса в Жуковом овраге у г. Гороховец, Владимирская область // Бюлл. региональной межведомственной стратиграфической комиссии по центру и югу Русской платформы. 2012. Вып. 5. С. 49–82.
Голубев В.К., Наумчева М.А., Фетисова А.М. Нижняя граница жуковского горизонта (верхняя пермь, Восточно-Европейская платформа) // Палеострат-2020. Тез. докл. годичн. собр. (науч. конф.) секции палеонтологии МОИП и московского отд. Палеонтол. об-ва при РАН. М.: ПИН РАН, 2020. С. 14–15.
Есин Д.Н. Чешуйный покров Amblypterina costata (Eichwald) и таксономия палеонисцид по изолированным чешуям // Палеонтол. журн. 1990. № 2. С. 89–96.
Есин Д.Н. Поднепермские палеонисциды Европейской части России. Дисс. … канд. геол.-мин. наук. М., 1995. 350 с.
Есин Д.Н., Машин В.Л. Ихтиолиты // Стратотипы и опорные разрезы верхней перми Поволжья и Прикамья / Ред. Есаулова Н.К., Лозовский В.Р. Казань: “Экоцентр”, 1996. С. 270–293.
Ивахненко М.Ф. Фаунистические комплексы тетрапод поздней перми Восточной Европы // Бюлл. МОИП. Отд. геол. 1990. Т. 65. Вып. 6. С. 55–60.
Ивахненко М.Ф. Тетраподы Восточно-Европейского плакката – позднепалеозойского территориально-природного комплекса. Пермь: “Книга”, 2001. 200 с.
Ивахненко М.Ф., Голубев В.К., Губин Ю.М. и др. Пермские и триасовые тетраподы Восточной Европы. М.: ГЕОС, 1997. 216 с. (Тр. Палеонтол. ин-та РАН. Т. 268).
Манаева Н.П., Шумов И.С. Новое Сартаковское местонахождение ископаемой позднепермской фауны // IV Междунар. конф. “Геология в школе и ВУЗе. Геология и цивилизация”. Тез. докл. (С.-Петербург, 25 июня–5 июля 2005 г) / Ред. Нестеров Е.М. СПб.: “Эпиграф”, 2005. С. 239–241.
Миних А.В., Андрушкевич С.О. Новый вид лучеперой рыбы из верхней перми Московской синеклизы // Изв. Сарат. ун-та. 2017. Т. 17. Сер. “Науки о Земле”. Вып. 2. С. 100–105.
Миних А.В., Арефьев М.П., Голубев В.К. Новый вид рыбы рода Isadia (Actinopterygii, Eurynotoidiformes) из нового местонахождения на р. Малая Северная Двина (терминальная пермь, Вологодская область) // Палеонтол. журн. 2015. № 6. С. 54–63.
Миних А.В., Миних М.Г. Ревизия некоторых палеонисков и новые рыбы из верхней перми Восточно-Европейской платформы и возможности использования их в стратиграфии // Вопросы геологии Южного Урала и Нижнего Поволжья. Сб. научн. трудов / Ред. Алешечкин О.И. Саратов: Изд-во Саратовск. ун-та, 1990. С. 84–104.
Миних А.В., Миних М.Г. Ихтиофауна перми Европейской России. Саратов: Изд. центр “Наука”, 2009. 244 с.
Миних А.В., Миних М.Г. Ихтиофауна в опорных разрезах средней и верхней перми Восточной Европы. Комплексы и зональная шкала. Ст. 1. Бассейн Северной Двины // Изв. Сарат. ун-та. 2018. Т. 18. Сер. “Науки о Земле”. Вып. 3. С. 184–193.
Наумчева М.А., Голубев В.К. Комплексы остракод пограничных отложений перми и триаса Московской синеклизы // Региональн. геол. и металлогения. 2019. № 80. С. 21–29.
Наумчева М.А., Голубев В.К., Балабанов Ю.П., Сенников А.Г. Остракоды из пограничных отложений перми и триаса разреза Окский Съезд, Нижний Новгород // Палеострат-2017. Тез. докл. годичн. собр. (науч. конф.) секции палеонтологии МОИП и московского отд. Палеонтол. об-ва при РАН. М.: ПИН РАН, 2017. С. 78.
Сенников А.Г., Голубев В.К. Пограничные отложения перми и триаса района гг. Вязники и Гороховец (Владимирская область) // Палеонтология и стратиграфия перми и триаса Северной Евразии: Матер. V междунар. конф., посв. 150-летию со дня рожд. В.П. Амалицкого (1860–1917). М.: ПИН РАН, 2010. С. 102–107.
Сенников А.Г., Голубев В.К. Открытие триаса в Нижнем Новгороде // Пермская система: стратиграфия, палеонтология, палеогеография, геодинамика и минеральные ресурсы. Сб. матер. Междунар. науч. конф. посвященной 170-летию со дня открытия пермской системы (5–9 сент. 2011 г.). Пермь: ПГУ, 2011. С. 307–312.
Шаповалов М.Е. Внутривидовая структура верхогляда Chanodichthys erythropterus озера Ханка // Изв. ТИНРО. 2010. Т. 162. С. 36–60.
Arefiev M.P., Golubev V.K., Balabanov Yu.P. et al. Type and reference sections of the Permian–Triassic continental sequences of the East European Platform: main isotope, magnetic, and biotic events. Sukhona and Severnaya Dvina Rivers field trip (XVIII Intern. Congr. on the Carbonif. and Permian). Moscow: PIN RAS, 2015. 104 p.
Bakaev A.S. New data on teeth of fish of the order Eurynotoidiformes (Pisces, Actinopterygii) from the Upper Permian of European Russia // Kazan Golovkinsky Stratigr. Meeting 2019: Late Paleozoic sedimentary Earth systems: stratigraphy, geochronology, petroleum resources. Abstr. vol. Kazan, 2019. P. 51–52.
Bakaev A.S., Kogan I. A new species of Burguklia (Pisces, Actinopterygii) from the Middle Permian of the Volga Region (European Russia) // Paläontol. Z. 2020. V. 94. № 1. P. 93–106. https://doi.org/10.1007/s12542-019-00487-6
Burrow C.J. Form and function in scales of Ligulalepis toombsi Schultze, a palaeoniscoid from the Early Devonian of Australia // Rec. S. Austr. Mus. 1994. V. 27. P. 175–185.
Chen D., Janvier P., Ahlberg P.E., Blom H. Scale morphology and squamation of the Late Silurian osteichthyan Andreolepis from Gotland, Sweden // Histor. Biol. 2012. V. 24. № 4. P. 411–423.
Choo B. Revision of the actinopterygian genus Mimipiscis (=Mimia) from the Upper Devonian Gogo Formation of Western Australia and the interrelationships of the early Actinopterygii // Earth and Environ. Sci. Trans. R. Soc. Edinb. 2011. V. 11. P. 1–28.
Choo B., Zhu M., Qu Q. et al. A new osteichthyan from the Late Silurian of Yunnan, China // PloS One. 2017 V. 12. № 3. e0170929, https://doi.org/10.1371/journal.pone.0170929
Cui X., Qiao T., Zhu M. Scale morphology and squamation pattern of Guiyu oneiros provide new insights into early osteichthyan body plan // Sci. Reports. 2019. V. 9. № 1. https://doi.org/10.1038/s41598-019-40845-7
Ermin R., Rau R., Reibedanz H. Der submikroskopische Aufbau der Ganoidschuppen von Polypterus im Vergleich zu den Zahngeweben der Säugetiere // Biomineralisation. 1971. V. 3. P. 12–21.
Esin D.N. Ontogenetic development of the squamation in some Palaeoniscoid fishes // Bull. Mus. Nat. Hist. Natur. Ser. 4. Sec. C. 1995. V. 17. P. 227–234.
Esin D.N. Peculiarities of trophic orientation changes in paleoniscoid assemblages from the Upper Permian of the European part of Russia // Modern Geol. 1997. V. 21. P. 185–195.
ärss T. Exoskeletal ultrastructure of early vertebrates // J. Vertebr. Paleontol. 2006. V. 26. № 2. P. 235–252
Nakayama S., Rapp T., Arlinghaus R. Fast-slow life history is correlated with individual differences in movements and prey selection in an aquatic predator in the wild // J. Animal Ecol. 2016. V. 86. № 2. P. 192–201.
Pindakiewicz M., Tałanda M., Sulej T. et al. New finds of teeth of the herbivorous actinopterygians from the latest Permian of East European Platform and feeding convergence among extinct and extant ray-finned fish // Acta Palaeontol. Pol. 2020. V. 65. № 1. P. 71–79.
Qu Q., Zhu M., Wang W. Scales and dermal skeletal histology of an early bony fish Psarolepis romeri and their bearing on the evolution of rhombic scales and hard tissues // PLoS One. 2013. V. 8. № 4. e61485.
Reissner E. Über die Schuppen von Polypterus und Lepidosteus // Arch. für Anat., Physiol. und Wiss. Medicin. 1859. P. 254–268.
Scholze F., Golubev V.K., Niedzwiedzki G. et al. Early Triassic conchostracans (Crustacea: Branchiopoda) from the terrestrial Permian-Triassic boundary sections in the Moscow syncline // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2015. V. 429. P. 22–40.
Scholze F., Golubev V.K., Niedźwiedzki G. et al. Late Permian conchostracans (Crustacea, Branchiopoda) from continental deposits in the Moscow Syneclise, Russia // J. Paleontol. 2019. V. 93. № 1. P. 72–97.
Schultze H.-P. Morphologische und histologische Untersuchungen an Schuppen mesozoischer Actinopterygier (Uebergang von Ganoidzu Rundschuppen) // N. Jb. Geol. Paläontol. Abh. 1966. V. 126. № 3. P. 232–314.
Schultze H.-P. Palaeoniscoidea-Schuppen aus dem Unterdevon Australiens und Kanadas und aus dem Mitteldevon Spitzbergens // Bull. Brit. Mus. Natur. Hist. Geol. 1968. V. 16. P. 342–368.
Schultze H.-P. Ausgangsform und Entwicklung der rhombischen Schuppen der Osteichthyes (Pisces) // Paläontol. Z. 1977. V. 51. P. 152–168.
Schultze H.-P. Scales, enamel, cosmine, ganoine, and early osteichthyans // C. R. Palevol. 2016. V. 15. № 1–2. P. 83–102.
Schultze H.-P. Hard tissues in fish evolution: history and current issues // Cybium. 2018. V. 42. № 1. P. 29–39.
Senay C., Boisclair D., Peres-Neto P.R. Habitat-based polymorphism is common in stream fishes // J. Animal Ecol. 2015. V. 84. P. 219–227.
Sire J.-Y., Géraudie J., Meunier F.J., Zylberberg L. On the origin of ganoine: histological and ultrastructural data on the experimental regeneration of the scales of Calamoichthys calabaricus (Osteichthyes, Brachiopterygii, Polypteridae) // Amer. J. Anat. 1987. V. 180. № 4. P. 391–402.
Trinajstic K. Scale morphology of the Late Devonian palaeoniscoid Moythomasia durgaringa Gardiner & Bartram, 1977 // Alcheringa. 1999a. V. 23. P. 9–19.
Trinajstic K. Scales of palaeoniscoid fishes (Osteichthyes: Actinopterygii) from the Late Devonian of Western Australia // Rec. Western Austral. Mus. 1999b. V. 57. P. 93–106.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал