

Палеонтологический журнал, 2021, № 2, стр. 77-87
Striatolamia tchelkarnurensis Glickman (Elasmobranchii: Lamniformes), самый молодой валидный вид рода Striatolamia
Т. П. Малышкина *
Институт геологии и геохимии им. А.Н. Заварицкого УрО РАН
Екатеринбург, Россия
* E-mail: prionace@yandex.ru
Поступила в редакцию 22.11.2019
После доработки 18.02.2020
Принята к публикации 14.05.2020
Аннотация
Изучение в коллекции Государственного Дарвиновского музея голотипа считавшегося невалидным вида Striatolamia tchelkarnurensis Glickman, 1964 (Lamniformes: Carchariidae), описанного из чеганской свиты Южного Тургая и содержащего более 30 изолированных зубов от одной особи, делает необходимым признать вид валидным. Морфологические характеристики зубов S. tchelkarnurensis как терминального таксона рода, вымершего в позднем эоцене, соответствуют эволюционным тенденциям рода – увеличению размеров зубов и усилению их режущей функции за счет изменения формы коронки и корня. В статье приводится переописание вида в соответствии с требованиями “Международного кодекса зоологической номенклатуры”.
ВВЕДЕНИЕ
Зубы акул рода Striatolamia часто встречаются в отложениях мелководных и прибрежных фаций палеоцена и эоцена по всему миру; они отмечены в Северной Африке, Северной Америке, Западной Европе, Украине, Причерноморье, Поволжье, Зауралье, Казахстане, Средней Азии, Китае, Антарктиде (Case et al., 1996; Малышкина, 2004; Kriwet, 2005; Удовиченко, 2009; Cappetta, 2012; Малышкина и др., 2013; Malyshkina, Ward, 2016; Trif et al., 2019 и др.). Наиболее многочисленны остатки стриатоламий в комплексах нижнего и низов среднего эоцена; в верхах среднего эоцена встречаемость Striatolamia падает; в низах верхнего эоцена род отмечается крайне редко, после чего он окончательно исчезает из палеонтологической летописи. Значительная эволюционная изменчивость и широкое географическое распространение рода делают его удобным объектом для решения задач биостратиграфии, но систематика его разработана недостаточно и требует ревизии.
Род Striatolamia был основан Л.С. Гликманом (1964, с. 102) на типовом виде Otodus macrotus Agassiz, 1843. В составе рода описано до девяти номинальных таксонов: палеоцен – S. cederstroemi Siverson, 1995, S. whitei (Arambourg, 1952), S. striata (Winkler, 1874); эоцен – S. rossica (Jaekel, 1895), S. elegans (Agassiz, 1843), S. macrota (Agassiz, 1843), S. sibirica Zhelezko in Zhelezko et Kozlov, 1999, S. tchelkarnurensis Glickman, 1964. По устоявшейся научной практике в настоящее время все эоценовые находки относят к виду S. macrota (Cappetta, 2012).
Наряду с выделением диагоноза рода, Гликман (1964, с. 124) описал новый вид, S. tchelkarnurensis, из “позднего эоцена” Тургайского прогиба, по найденным геологами Л.В. Мироновой и В.А. Броневым на плато Челкар-Нура (Шалкар-Нура) в отложениях чеганской свиты остаткам акулы. Голотип, хранящийся в Государственном Дарвиновском музее (ГДМ), содержит разрозненные позвонки, фрагменты кальцинированного хряща и более 30 отдельных зубов от одной особи. Формального описания вида Гликманом сделано не было, сопроводительные иллюстрации были выполнены на недостаточном уровне, вследствие чего вид в мировой научной практике был практически забыт, и лишь дважды упомянут в синонимике S. macrota (Железко, Козлов, 1999; Cappetta, 2006). Недавнее переизучение голотипа S. tchelkarnurensis выявило его четкие отличия от других видов в общем эволюционном тренде рода. Цель данной работы – переописание вида S. tchelkarnurensis Glickman, 1964 в соответствии с требованиями “Международного кодекса зоологической номенклатуры” (2004).
ГЕОГРАФИЯ, ГЕОЛОГИЯ И ВОЗРАСТ НАХОДКИ
Типовое местонахождение, Челкар-Нура, расположено в центральной части Тургайского прогиба, в южных отрогах Тургайского плато, называемых “горы Челкарнура” (или Шалкарнура на некоторых географических картах, что является вариациями в произношении местного географического названия), в Актюбинской области Казахстана. Горы Челкарнура протяженностью около 150 км вытянуты полукругом с севера на юго-восток. Максимальные высоты, достигающие 190–230 м, находятся в южной части. Южные склоны Челкарнуры круто спускаются к бессточному солончаку Шаркалтениз с абсолютной высотой 50 м. Приблизительные координаты находки 48°23′ с.ш., 63°05′ в.д. (рис. 1, а).
Рис. 1.
а – Географическое положение находки Striatolamia tchelkarnurensis Glickman, б – сводный разрез эоценовых отложений в районе находки (составлен по данным: Беньямовский и др., 1993; Васильева, 2014а, б; Геологии СССР…, 1971; Гликман, 1964). Обозначения: 1 – пески и алевриты, 2 – песчаники, 3 – глина алевритистая, 4 – перерыв, несогласное залегание пород, 5 – расположение скважины № 85, 6 – расположение типового местонахождения Челкар-Нура, 7 – предполагаемое место находки голотипа S. tchelkarnurensis; df – зоны по динофлагеллятам.
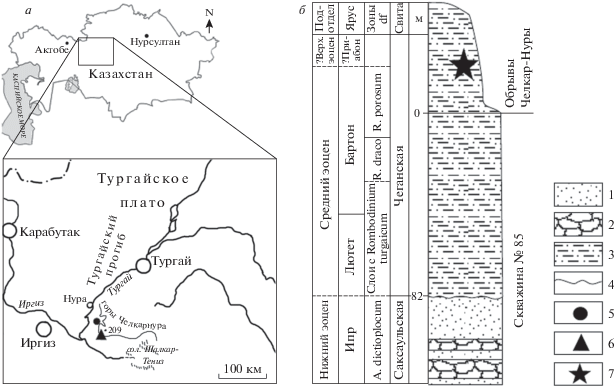
Палеоген в центральной и южной части Тургайского прогиба представлен морскими отложениями; таупская, тасаранская, саксаульская и чеганская свиты последовательно накапливались в течение эоценового периода (Геология СССР…, 1971; Беньямовский и др., 1993). Отложения чеганской свиты, формировавшейся в нормальных морских условиях в течение лютетского, бартонского и приабонского веков, представлены глинами некарбонатными, голубовато-серыми, пепельно-серыми, зеленовато-серыми, алевритистыми, переходящими в алевриты глинистые, особенно в верхней части (Беньямовский и др., 1993). Недавние исследования палеогеновых отложений по скважине № 85, пробуренной в 80-х гг. XX в. в 55 км к северо-востоку от устья р. Иргиз у подножия обрывов гор Челкарнура, выявили в верхней части разреза скважины, в чеганской свите, диноцист двух смежных зон, зоны Rombodinium draco и зоны R. porosum, что соответствует нанопланктонной зоне NP17 (бартон) (Васильева, 2014а, б; рис. 1, б). Выше по разрезу глины чеганской свиты, вскрытые скважиной № 85, выходят на дневную поверхность и слагают чинки Челкарнуры (Геология СССР…, 1971). Гликман (1964, с. 124) указывает, что все зубы Striatolamia tchelkarnurensis “… происходят из чеганских отложений Челкар-Нуры”, то есть из обрывов, что находится гипсометрически и стратиграфически выше верхнего уровня скважины и указывает на возраст находки не древнее зоны NP17 (верхняя часть бартона или приабон). Сам Гликман (1964) считал, что возраст находки соответствует верхнему тонгру, или “зоне турителл” по Н.B. Овечкину (1962), т.е., приабону.
МАТЕРИАЛ И МЕТОДЫ ИЗУЧЕНИЯ
Образцы были собраны сотрудниками Тургайской геологической партии Л.В. Мироновой и В.А. Броневым в 50-х гг. XX века в обрывах гор Челкарнура, в южной части Тургайского прогиба (Гликман, 1964), и хранятся в ГДМ с коллекционными номерами КП ОФ 8057/92–8057/211. Все образцы принадлежат одной особи – это более 30 разрозненных зубов, отдельные позвонки и фрагменты кальцифицированного хряща (табл. 1, рис. 2, а, б). Зубы были сфотографированы без напыления фотокамерой Canon EOS с макрообъективом Canon Macro Lens EF-S 60 mm 1:2.8 USM.
Таблица 1.
Список зубов Striatolamia tchelkarnurensis в коллекции КП ОФ 8057, ГДМ (голотип)
| Номер в коллекции | Положение в челюсти | Примечание |
|---|---|---|
| Верхняя челюсть | ||
| 8057/108 | 1-й передний | поврежден |
| 8057/97 8057/100 |
2-й передний | поврежден |
| 8057/95 | 3-й передний | |
| 8057/94 8057/109 8057/119 8057/125 |
боковой | поврежден поврежден |
| 8057/99 8057/111 8057/116 8057/119 8057/124 8057/127 8057/128 |
боковой дистальной серии | поврежден коронка коронка поврежден поврежден поврежден поврежден |
| 8057/122 8057/? |
боковой самых дистальных серий | поврежден поврежден |
| Нижняя челюсть | ||
| 8057/93 8057/101 8057/102 |
1-й передний | поврежден поврежден |
| 8057/92 | 2-й передний | поврежден |
| 8057/107 8057/135 |
3-й передний | коронка поврежден |
| 8057/105 8057/106 8057/114 8057/115 8057/136 |
передний неопределимой позиции | коронка коронка коронка коронка поврежден |
| 8057/96 | боковой | |
| 8057/104 8057/110 |
боковой неопределимой позиции | поврежден коронка |
Рис. 2.
Striatolamia tchelkarnurensis Glickman из колл. ГДМ КП ОФ 8057: а – фрагмент хряща; б – позвонок; в – реконструкция зубных рядов верхней и нижней челюстей на основе имеющихся зубов: S – симфизный зуб, A1–A3 – зубы передних серий, L1–L6 – зубы боковых серий, P – зубы задних серий. Отсутствующие позиции взяты в скобки.

Определение позиции зубов в челюсти проводилось с использованием реконструкции зубных рядов Striatolamia macrota, выполненной C. Каннингемом (Cunningham, 2000).
Межвидовой сравнительный анализ зубов акул рода Striatolamia проводился путем визуального сравнения, а также методом наложения контуров зубов одинакового местоположения в челюсти, приведенных к одной высоте.
При описании зубов использовалась стандартная терминология (Applegate, 1965; Cappetta, 2012). Традиционная система группы (Cappetta, 2012) пересматривается, что обсуждается ниже.
Благодарю А.И. Клюкину и Е.М. Байкину (Москва) за возможность изучения коллекции ГДМ; Т.В. Шевченко и В.Л. Стефанского (Киев) за консультации по современной стратиграфии Украины; А.В. Лаврова (Москва) за предоставление фотоаппаратуры; Ф.A. Триколиди (С.-Петербург), М.П. Покровского, В.В. Черных (Екатеринбург) и Д. Ворда (Лондон) за помощь в работе над статьей, А.Ф. Банникова (Москва) и Е.В. Попова (Саратов) за ценные замечания в ходе рецензирования, позволившие улучшить рукопись.
СИСТЕМАТИЧЕСКАЯ ПАЛЕОНТОЛОГИЯ
ОТРЯД LAMNIFORMES
СЕМЕЙСТВО CARCHARIIDAE JORDAN ET GILBERT, 1883
Род Striatolamia Glickman, 1964
Типовой вид – Otodus macrotus Agassiz, 1843, cредний эоцен Франции.
Замечания. Зубы, принадлежащие палеоценовым представителям рода, описаны М. Леришем (Leriche, 1902). Гликман классифицировал род Striatolamia в составе семейства Odontaspididae Müller et Henle, 1839, что долгое время было неоспоримым (Cappetta, 1987). Задолго до этого Э. Уайт (White, 1931), изучавший детали строения зубных систем ряда акул отряда Lamniformes, причислил S. striata к роду Synodontaspis (Carcharias), принадлежащему семейству Odontaspididae. Реконструкция зубных рядов S. macrota на основании сравнения их с озублением современного вида Carcharias taurus показывает очевидное морфологическое и функциональное сходство зубных систем родов Striatolamia и Carcharias (Cunningham, 2000).
М. Сиверсон (Siverson, 1995) предположил происхождение рода Striatolamia в позднем мелу от общего предка с родом Anomotodon (семейство Mitsukurinidae). A. Каппетта и Д. Нольф (Cappetta, Nolf, 2005) поддержали принадлежность Striatolamia к семейству Mitsukurinidae, и в настоящее время эта точка зрения получила довольно широкое распространение (Cappetta, 2012). Однако мы не придерживаемся данного взгляда, что основано в том числе и на генетических данных. Молекулярно-генетические исследования (Naylor et al., 1997, 2012; Martin et al., 2002; Human et al., 2006; Vélez-Zuazo, Agnarsson, 2011) свидетельствуют о том, что современные виды рода Odontaspis [O. ferox (Risso, 1810) и O. noronhai (Maul, 1955)] имеют отдаленное родство с современным представителем рода Carcharias (C. taurus Rafinesque, 1810), а значит, семейство Odontaspididae полифилетично. Ряд исследователей (Adolfssen, Ward, 2013; Stone, Shimada, 2019) предлагают разделить семейство Odontaspididae, восстановив семейство Carchariidae Jordan et Gilbert, 1883; в данной работе мы, вслед за Я. Адолфссеном и Д. Вордом (Adolfssen, Ward, 2013) помещаем род Striatolamia в семейство Carchariidae, отделяя его от Odontaspididae sensu stricto.
Striatolamia tchelkarnurensis Glickman, 1964
Табл. IX, X (см. вклейку)
Striatolamia tchelkarnurensis: Гликман, 1964, c. 124, табл. XI , фиг. 1–8 , 14 ; табл. XII , фиг. 1, 2.
Голотип – ГДМ колл. № КП ОФ 8057, разрозненные челюстные зубы от одной особи; Казахстан, Актюбинская обл., горы Челкарнура; эоцен, поздний бартон или приабон, верхняя часть чеганской свиты (табл. IX, X).
Описание (рис. 2). Гликман (1964, с. 124) писал: “Зубы характеризуются предельной для Striatolamia величиной, максимально расширенными коронками, внутренняя сторона которых несет уже лишь рудиментарную ребристость, столь характерную для зубов представителей этого рода”.
На основании изучения зубов голотипа описание Гликмана может быть расширено. Ламнообразные акулы с крупными зубами (передние зубы до 7 см в высоту). Коронки зубов робустные. Режущий край прямой, не заходит на лабиальную сторону коронки и достигает ее основания. Коронки передних зубов верхней челюсти в профиль не имеют S-образного изгиба, коронки передних зубов нижней челюсти имеют очень слабый S-образный изгиб. Зубы боковых серий имеют мезио-дистально широкие, лабио-лингвально уплощенные коронки и тесно примыкающие к ним крупные добавочные вершины, часто треугольной формы и с дополнительными зубчиками в основании, как у наружного края, так и между главной и добавочной вершинами. Корень у боковых зубов верхней челюсти расширен мезио-дистально.
Описание отдельных зубов. Верхнечелюстные зубы передних серий (табл. IX , фиг. 1–4 ) крупные, в высоту до 7 см. Коронки мощные. Режущие края всегда достигают основания коронки и не заходят на лабиальную поверхность коронки. Центральный бугорок развит в средней степени, питающая борозда неглубокая, начинается на верхушке бугорка с центрального питающего отверстия. Лабиальная поверхность коронки слегка выпуклая (табл. IX , фиг. 1а, 3а), при взгляде в профиль прямая (табл. IX , фиг. 1б, 2б, 3б, 4б). Лингвальная поверхность выпуклая. Энамелоид гладкий на лабиальной стороне и слегка орнаментирован легкой вертикальной штриховкой в нижней половине на лингвальной стороне.
Первый верхний зуб передней серии 8057/108 (табл. IX , фиг. 1) имеет крепкую, почти вертикальную коронку и корень с короткими, узко поставленными ветвями (дистальная ветвь сломана). Режущие края хорошо развиты, у нижнего края коронки тонкие, достигают основания коронки. Мезиальная добавочная вершина маленькая, треугольная, не соединена с главной вершиной. Дистальная сломана.
Второй верхний зуб передней серии 8057/97 и 8057/100 (табл. IX , фиг. 2, 4 ) имеет наклоненную дистально в верхней половине коронку, которая более плоская и сильнее расширена в нижней части, чем у первого переднего зуба. Ветви корня широко расставлены, дистальная длиннее и направлена более горизонтально; проксимальная короче и почти вертикальная, так что мезиальные края коронки и корня образуют почти прямую линию. Режущие края хорошо развиты по всей длине и достигают нижнего края коронки. Добавочные вершины треугольные и расположены у нижнего края коронки у 8057/100; у 8057/97 сломаны.
Третий верхний зуб передней серии 8057/95 (табл. IX , фиг. 3 ) имеет наклоненную дистально коронку с вертикально направленной верхней частью, и широко расставленные ветви корня, среди которых мезиальная значительно уже и длиннее. Коронка довольно плоская; лабиальная поверхность коронки слегка выпуклая, прямая в профиль, лингвальная поверхность выпуклая, режущие края достигают основания коронки. Добавочные вершины небольшие, с закругленными верхушками, расположены близко к главной вершине, но отделены от нее промежутком. Энамелоид на лингвальной стороне коронки имеет легкую вертикальную штриховку.
Зубы нижней челюсти передних серий (табл. IX , фиг. 5–9 ) имеют легкий лабио-лингвальный S-образный изгиб. Энамелоид на лингвальной стороне коронки в нижней трети покрыт очень легкой вертикальной штриховкой. Центральный бугорок развит в средней степени; питающая борозда неглубокая, начинается на верхушке бугорка, в верхней ее части расположено центральное питающее отверстие.
Первый нижний зуб передней серии 8057/93, 8057/101 и 8057/102 (табл. IX , фиг. 5, 7, 9 ) имеет наибольший S-образный изгиб коронки и выступающий центральный бугорок при взгляде сбоку. Режущие края немного не достигают нижнего края коронки. Ветви корня короткие, с закругленными концами, расставлены не широко.
Второй нижний зуб передней серии 8057/92 (табл. IX , фиг. 6) имеет прямую коронку, немного наклоненную к углу челюсти, слегка расширяющуюся у основания при лабиальном взгляде. В профиль S-образный изгиб очень слабо выражен. Лабиальная поверхность коронки немного выпуклая, лингвальная выпуклая. Лингвальная орнаментация энамелоида присутствует в нижней трети коронки, слабо выражена. Режущие края достигают основания коронки, не выходят на лабиальную поверхность. Добавочные вершины маленькие, треугольные, отделены от главной вершины. Центральный бугорок выражен в средней степени. Ветви корня длинные, закругленные на концах, широко расставлены.
Третий нижний зуб передней серии 8057/135 (табл. IX , фиг. 8 ) очень своеобразный, по характеристикам отдельных признаков представляет переходную форму между передними и боковыми зубами: довольно широкая коронка и короткие ветви корня сближают данный зуб с боковыми зубами, но значительная лабио-лингвальная толщина коронки, а также небольшой размер и форма добавочной вершины говорят о том, что зуб занимал переднюю позицию. Коронка мощная, S‑образный изгиб и центральный бугорок наименее выражен среди всех передних зубов. Лабиальная поверхность коронки слегка выпуклая, лингвальная выпукла в меньшей степени, чем у зубов, расположенных проксимальнее в зубном ряду. Режущий край достигает основания коронки. Дистальная ветвь корня широкая и короткая, мезиальная сломана. Дистальная добавочная вершина маленькая, с закругленной верхушкой, лабио-лингвально уплощенная, имеет выраженный режущий край; примыкает к главной вершине, спускаясь по корню ниже ее основания; мезиальная добавочная вершина сломана.
Зубы боковых серий (табл. X ) заметно отличаются от передних широкими коронками и короткими ветвями корня.
Зубы боковых серий верхней челюсти (табл. X , фиг. 3–6, 8 ). Коронка имеет главную и добавочные вершины. Главная вершина наклонена и слегка изогнута в сторону угла пасти, лабио-лингвально уплощена, степень выпуклости лингвальной и лабиальной сторон может быть почти равной у некоторых зубов. Режущие края почти прямые, благодаря чему форма главной вершины практически треугольная. Орнаментация энамелоида на лингвальной поверхности коронки присутствует в виде слабой вертикальной штриховки в нижней половине высоты. Лабиальная поверхность гладкая, либо несет несколько неглубоких вертикальных борозд в нижней трети высоты (табл. X , фиг. 3, 5 ). Добавочные вершины близко посажены к главной вершине, по одной с каждой стороны от нее. Мезиальная добавочная вершина может быть мельче дистальной (табл. X , фиг. 6). Добавочные вершины имеют различную форму. Часто они треугольные (табл. X , фиг. 3, 5 ), округлые (табл. X , фиг. 4, 6 ), а также могут раздваиваться (табл. X , фиг. 8 ). У основания добавочных вершин могут присутствовать небольшие зубцы, расположенные как между главной и добавочной вершиной, так и дистально (табл. X , фиг. 3, 5, 6 ). Корень боковых зубов верхней челюсти невысокий, расширен мезио-дистально, выступает за наружные края добавочных вершин; лабио-лингвально уплощен. Ветви корня округлые, короткие, арка невысокая и широкая, по контуру промежуточная между U- и V-образной. Гребни корня и центральный бугорок слабо выражены, питающая борозда неглубокая и неширокая, центральное питающее отверстие небольшое. Каннингем (Cunningham, 2000) указывает, что у S. macrota зубы верхних боковых серий имеют более длинную мезиальную ветвь корня (на материале из лондонской глины ипрского возраста). Данный признак не может быть в полной мере прослежен у S. tchelkarnurensis, так как сохранность некоторых зубов неполная; у зуба 8057/94 (табл. X , фиг. 6 ) ветви разной формы, но одной длины.
Боковые зубы нижней челюсти (табл. X , фиг. 1, 2, 7 ) отличаются от верхнечелюстных меньшим размером, меньшим дистальным наклоном главной вершины, меньшей шириной коронки и корня, более выпуклым центральным бугорком и гребнями корня. Режущие края коронки сильнее изогнуты, вследствие чего очертания коронки более изящные, чем у боковых зубов верхней челюсти. S-образный изгиб почти отсутствует. Добавочные вершины также имеют изменчивую форму, от треугольной, как у зуба одной из первых боковых серий (табл. X , фиг. 7 ), до раздвоенной у боковых зубов более дистальных серий (табл. X , фиг. 1, 2). Небольшие зубцы в основании добавочных вершин также могут присутствовать (табл. X , фиг. 7 ). Корень в меньшей степени расширен мезио-дистально, по сравнению с боковыми зубами верхней челюсти, лабио-лингвально уплощен; ветви корня округлые.
Сравнение. Зубы S. tchelkarnurensis можно дифференцировать от других позднепалеоценовых и эоценовых видов Striatolamia по ряду признаков (для сравнения везде взяты зубы одинакового местоположения в челюсти, принадлежавшие взрослым особям). От всех видов своего рода S. tchelkarnurensis отличается третьим передним зубом нижней челюсти (8057/135, табл. IX , фиг. 8 ), отчасти напоминающим боковой зуб сравнительно широкой и толстой коронкой и широкими и короткими ветвями корня. У остальных видов данный зуб имеет все признаки зубов передних позиций.
От S. macrota, циркумглобально распространенного в ранне- и среднеэоценовых отложениях, S. tchelkarnurensis отличается следующими признаками. Передние зубы S. tchelkarnurensis более мощные, прямые, S-образный изгиб проявлен в меньшей степени; режущие края более прямые, поскольку коронка не имеет сужения в средней части; режущий край на передних зубах S. tchelkarnurensis всегда достигает основания коронки (табл. IX ), исключая 1-й нижний передний зуб (табл. IX , фиг. 5, 7, 9 ), тогда как у зубов S. macrota всех передних серий режущий край заметно не достигает основания коронки (Железко, Козлов, 1999; Trif et al., 2019): ср. 2-й передний зуб верхней челюсти (Trif et al., 2019, рис. 5.5 -7') с аналогичным, изображенным на табл. IX , фиг. 2 и 4 настоящей статьи. Орнаментация лингвальной поверхности коронки более тонкая и легкая. Боковые зубы S. tchelkarnurensis отличаются от зубов S. macrota наличием более крупных и острых добавочных вершин. Они часто имеют форму почти равносторонних треугольников, тогда как у зубов S. macrota добавочные вершины, даже если имеют треугольную форму (в основном у молодых особей), то заметно ниже по высоте. В основном добавочные вершины боковых зубов S. macrota имеют закругленную вершину. Дополнительные зубцы у основания добавочных вершин встречаются очень редко у зубов S. macrota, и они не так хорошо развиты, как у зубов S. tchelkarnurensis. Дигнатическая гетеродонтия у S. tchelkarnurensis менее выражена, чем у S. macrota. Для зубов во всех положениях отношение высоты коронки к ее ширине у S. macrota больше, чем у S. tchelkarnurensis. Корень у верхних боковых зубов S. tchelkarnurensis мезио-дистально значительно шире, чем у S. macrota.
Зубы S. striata из палеоценовых отложений Евразии и Америки намного мельче зубов S. tchelkarnurensis, более тонкие и имеют более резкую лингвальную орнаментацию энамелоида. У боковых зубов S. striata дигнатическая гетеродонтия выражена сильнее, чем у S. tchelkarnurensis. Коронки боковых зубов S. striata расширяются от вершины к основанию неравномерно, имея узкую верхнюю часть и расширенную нижнюю, тогда как у S. tchelkarnurensis коронки равномерно расширены. Добавочные вершины на боковых зубах S. striata расположены очень близко к главной вершине, у S. macrota они разделены промежутком, а у S. tchelkarnurensis вновь сближены.
Зубы взрослых особей S. sibirica из среднего эоцена Зауралья в отличие от таковых S. tchelkarnurensis более грацильные и характеризуются сильнее изогнутыми режущими краями, по сравнению с прямыми режущими краями S. tchelkarnurensis. Коронки боковых зубов у S. sibirica более тонкие при взгляде с лабиальной стороны, отношение высоты коронки к ее ширине у основания больше, добавочные вершины посажены тесно к главной вершине. Некоторое сходство этих видов в зубных характеристиках заключается в том, что боковые зубы S. sibirica имеют довольно высокий процент добавочных вершин в форме равностороннего треугольника. Но корень у верхних боковых зубов S. tchelkarnurensis мезио-дистально значительно шире, чем у S. sibirica.
Зубы “S. elegans” из раннего эоцена Европы и Средней Азии меньше по размеру, чем у S. tchelkarnurensis, значительно тоньше и грациознее, с меньшими, более низкими добавочными вершинами и более выраженной орнаментацией энамелоида на лингвальной стороне коронки. Режущий край на передних зубах “S. elegans” заканчивается значительно выше основания коронки, тогда как у S. tchelkarnurensis он достигает корня.
Зубы S. macrota var. rossica из бартонских отложений Украины, обладая широкими и крепкими коронками, на первый взгляд, наиболее схожи с S. tchelkarnurensis. Главное их различие состоит в том, что у верхних боковых зубов S. tchelkarnurensis корни под добавочными вершинами сильнее расширены мезио-дистально. Наряду с меньшей шириной корня, у верхних боковых зубов S. macrota var. rossica относительно меньший размер добавочных вершин. Таксон S. macrota var. rossica обсуждается ниже.
Материал. Более 30 отдельных зубов, позвонки и фрагменты хряща от одной особи (табл. 1; рис. 2, а, б) из местонахождения Челкар-Нура.
ОБСУЖДЕНИЕ
Род Striatolamia включает в себя несколько видов средне- и крупноразмерных акул, обитавших в зоне шельфа, начиная со второй половины палеоцена, и ставших доминантами в прибрежных сообществах хрящевых рыб в раннем и среднем эоцене. Чаще всего в литературе упоминается вид S. macrota. Вероятно, это связано со значительной продолжительностью лютетского века, около 8 млн лет (Ogg et al., 2016), а также потому, что к данному таксону относят довольно разнородные морфотипы, часть из которых может принадлежать другим видам. Верхняя граница существования рода дискуссионна, по разным мнениям, род исчез в бартоне (Cappetta, 1987) или в приабоне (Железко, Козлов, 1999; Удовиченко, 2008; Cappetta, 2012). Сообщения о находках зубов Striatolamia в олигоцене (Йекель, 1895), вероятно, связаны с устаревшими стратиграфическими данными.
Как было показано выше, вмещающие S. tchelkarnurensis отложения Челкарнуры не могут быть датированы древнее зоны NP17 по наннопланктону, что соответствует верхней части бартона или приабону. Наиболее близкой к S. tchelkarnurensis формой, как по признакам зубной системы, так и по времени существования, можно считать S. macrota var. rossica, поэтому следует остановиться на ней подробнее. Данную форму, отличавшуюся от вида S. macrota более крепкими коронками и более крупными добавочными вершинами, описал О. Йекель как Odontaspis macrota var. rossica, из “нижнеолигоценовых” глауконитовых песков окрестностей Днепропетровска (Йекель, 1895, табл. I , фиг. 8–17 ). Гликман (1964) возвел эту форму в ранг вида Striatolamia rossica (Jaekel), с тремя подвидами в стратиграфическом интервале бартон–приабон: S. r. prima (тип происходит из аманкизилитской свиты Мангышлака), S. r. usakensis (тип из “рыбной свиты”, или средней части шорымской свиты Мангышлака, ранний или средний бартон) и S. r. rossica из адаевской свиты Мангышлака, приабон. В.И. Железко (Железко, Козлов, 1999) перевел подвиды Гликмана в разряд “стадий”, согласно своей оригинальной биостратиграфической концепции. В качестве эталона для S. macrota стадия rossica им был выбран зуб, изображенный Йекелем (1895, табл. I , фиг. 17 ), из слоя глауконитовых песков, вскрытых в ложе р. Днепр у г. Днепропетровск (ранее Екатеринослав) при строительстве железнодорожного моста в 1892 г. Отложения были вскрыты единожды и очень локально, в месте погружения кессона № 10; больше подобные отложения нигде не встречались. В данном слое В.А. Домгером была собрана и Н.А. Соколовым изучена коллекция из 21 вида моллюсков (Соколов, 1894); тогда же было найдено несколько зубов акул, причисленных Йекелем (1895) к трем таксонам: Odontaspis macrota var. rossica Jkl., Carcharodon sokolowi Jkl., Hypotodus trigonalis Jkl. (роды Striatolamia, Otodus, Jaekelotodus соответственно, согласно современной систематике). Железко полагал (Железко, Козлов, 1999), что данный слой глауконитовых песков относится к позднеэоценовым мандриковским слоям, но ни Домгер, ни Соколов не отождествляли глауконитовые пески и мандриковские слои (Соколов, 1894). Соколов (1894), описывавший малакофауну, коррелировал глауконитовые пески с латдорфскими песками Германии (поздний эоцен). Недавнее переизучение коллекции Домгера (Удовиченко, 2009; Стефанский и др., 2011) свидетельствует о более древнем, чем у мандриковских слоев, возрасте песков кессона №10. Согласно этим исследованиям, возраст комплексов из мандриковских слоев соответствует зонам NP19–основание NP20 ОСШ по наннопланктону (приабон), а из глауконитовых песков – NP17–NP18 (поздний бартон–ранний приабон). Возраст бартон–ранний приабон подтверждается и присутствием в сборах Домгера зубов акулы Otodus sokolowi (Jaekel, 1895). Также высказывалось мнение о более молодом возрасте песков относительно мандриковских слоев (Березовский, 2013).
Важно заметить, что зубы, изображенные Йекелем (1895, табл. II ) как ?O. macrota var. rossica, происходят из окрестностей г. Градижска. Н.И. Удовиченко (2004, 2006) установил, что эти зубы были перемыты в четвертичные отложения из киевской свиты, датируемой Т.С. Рябоконь (2002) на основании анализа комплексов фораминифер лютетом или ранним бартоном. Данные зубы не несут характерных для O. macrota var. rossica черт.
Типовая серия O. macrota var. rossica была помещена в ЦНИГР музей (ВСЕГЕИ, С. Петербург) и была утеряна. Из всех описанных Йекелем (1895) экземпляров на сегодняшний день в коллекции сохранилось лишь два зуба ?O. macrota var. rossica (Йекель, 1895, табл. II , фиг. 8 и 10 ) из окрестностей Градижска, являющихся, как было показано выше, перемытыми в четвертичные отложения из киевской свиты и справедливо отнесенных самим Йекелем к O. macrota var. rossica под знаком вопроса. Поэтому, принимая во внимание факт недоступности типового нахождения (пески из кессона № 10) для переизучения, вид S. rossica должен быть признан nomen dubium.
S. sibirica Zhelezko (Железко, Козлов, 1999) был описан из чеганской свиты в Северном Тургае (местонахождение Качар, голотип и паратипы) и из тавдинской свиты Южного Зауралья (местонахождение Курган, паратип). Вмещающие отложения были датированы приабоном, что делало S. sibirica самым молодым видом рода (Объяснительная записка…, 1997; Железко, Козлов, 1999). Однако последующие исследования по диноцистам показали, что чеганская свита в разрезе Качар имеет более древний возраст, очевидно, лютет (Васильева, 2014а, б, 2015). Зубы с аналогичными S. sibirica признаками были описаны А. Роговичем (1860, табл. V , фиг. 1–23 ) из голубых мергелей в окрестностях Киева, которые ныне относятся к киевской свите и датируются лютетом–ранним бартоном (Рябоконь, 2002). Вопрос о валидности вида S. sibirica требует дополнительных исследований.
Реконструкция зубной системы. Восстановление зубных рядов верхней и нижней челюстей S. tchelkarnurensis проводилось с использованием реконструкции зубной системы S. macrota, выполненной Каннингемом (Cunningham, 2000) на основании сравнения зубов S. macrota с современным представителем семейства, Carcharias taurus. С. Апплегейт (Applegate, 1965), изучив 12 челюстей C. taurus, указывает на внутривидовое различие в количестве зубов боковых и задних местоположений и приводит следующую зубную формулу с количественными вариациями:
A3 I1 L6-8 P6-19
S1 A3 L5-6 P4-14
В коллекции ГДМ КП ОФ 8057 отсутствуют зубы симфизной, интермедиальной и всех задних позиций (см. табл. 1). Многие зубы представлены только коронками, что не позволило точно определить их положение в челюсти и восстановить зубные ряды максимально полно. Получившаяся реконструкция представлена на рис. 2, в.
ЗАКЛЮЧЕНИЕ
S. tchelkarnurensis является самым молодым видом рода. Эволюционные тенденции линии Striatolamia, начиная с эоцена, шли по пути усиления режущей функции зубов, что проявлялось в постепенном лабио-лингвальном уплощении зубов боковых серий, расширении коронок вместе с увеличением ширины и высоты добавочных вершин у зубов боковых серий, в сближении добавочных вершин с главной и формировании в результате почти непрерывного общего режущего края. Возможно, что обособление S. tchelkarnurensis было связано с изолированностью бассейна Тургайского пролива в позднем эоцене.
Список литературы
Беньямовский В.Н., Васильева О.Н., Левина А.П., Пронин В.Г. Палеоген Южного Зауралья. Статья I. Палеоген в южной части Тургайского прогиба // Изв. ВУЗов. Геол. и разведка. 1993. № 6. С. 22–37.
Березовский А.А. Возраст глауконитовых песков Днепропетровска // Збiрн. наук. праць Iнст. геол. наук НАН Украïни. 2013. Т. 6. Вип. 1. С. 128–130.
Васильева О.Н. Эоценовые последовательности диноцист в Южном Тургае // Ежегодник-2013. Екатеринбург: УрО РАН, 2014а. С. 15–26 (Тр. ИГГ УрО РАН. Т. 161).
Васильева О.Н. Зональная последовательность диноцист эоцена южной части Тургайского прогиба (Челкарская впадина, Казахстан) // Литосфера. 2014б. № 6. С. 132–140.
Васильева О.Н. О возрасте тавдинской свиты в Тургайском прогибе // Ежегодник-2014. Екатеринбург: УрО РАН, 2015. С. 8–20 (Тр. ИГГ УрО РАН. Т. 162).
Гликман Л.С. Акулы палеогена и их стратиграфическое значение. М.–Л., 1964. 229 с.
Железко В.И. О развитии зубного аппарата пелагических акул рода Hispidaspis Sokolov, 1978 (по материалам из отложений среднего и верхнего мела Средней Азии и Казахстана) // Материалы по стратиграфии и палеонтологии Урала. № 4. Екатеринбург: УрО РАН, 2000. С. 128–135.
Железко В.И., Козлов В.А. Эласмобранхии и биостратиграфия палеогена Зауралья и Средней Азии // Материалы по стратиграфии и палеонтологии Урала. № 3. Екатеринбург: УрО РАН, 1999. С. 1–324.
Йекель О. Нижнетретичные селяхии Южной России // Тр. Геол. ком-та. 1895. Т. 9. № 4. С. 1–25.
Малышкина Т.П., Гонсалес-Барба Р., Банников А.Ф. Находки зубов эласмобранхий в бартоне Северного Кавказа (Россия) и Крыма (Украина) // Палеонтол. журн. 2013. № 1. С. 76–80.
Международный кодекс зоологической номенклатуры. Изд. четвертое. М.: Тов-во научн. изданий КМК, 2004. 224 с.
Овечкин Н.В. Биостратиграфия палеогеновых отложений Тургайского прогиба и Северного Приаралья // Тр. ВСЕГЕИ. Нов. сер. 1962. Т. 77. 170 с.
Рогович А. Об ископаемых рыбах губерний Киевского учебного округа. Палеонтология, часть систематическая. Киев, 1860. 87 с.
Рябоконь Т.С. Биостратиграфия опорного разреза киевской свиты (средний эоцен) Днепровско-Донецкой впадины по данным изучения фораминифер // Геол.-мінерал. вісн. 2002. № 2(8). С. 39–50.
Соколов Н.А. Фауна нижнеолигоценовых отложений окрестностей Екатеринослава. I. Фауна глауконитовых песков Екатеринославского железнодорожного моста // Тр. Геол. ком-та. 1894. Т. 9. № 3. С. 1–136.
Стефанский В.Л., Удовиченко Н.И., Стефанский М.В., Братишко А.В. О литологическом составе и стратиграфическом объеме мандриковских слоев г. Днепропетровска // Проблеми стратиграфії і кореляції фанерозойських відкладів України. Матер. XXXIII сес. Палеонтол. тов-ва НАН України. Kиїв: ІГН НАНУ, 2011. С. 74–75.
Стратиграфические схемы Урала (мезозой, кайнозой). Матер. IV Уральского межведомственного стратиграфического совещания (Свердловск, 20 апреля 1990 г.). Екатеринбург: Межведомственный стратигр. комитет России; АООТ “Уральская геологосъемочная экспедиция”, 1997.
Удовиченко Н.И. О переотложенных зубах акул из района Градижска Полтавской области // Проблеми стратиграфії фанерозою України. Kиїв, 2004. С. 141–143.
Удовиченко Н.И. Зубы акул из отложений киевского региояруса района Градижска // Проблеми палеонтології та біостратиграфії протерозою і фанерозою України: Зб. наук. пр. ІГН НАН Украины. Kиїв, 2006. С. 201–208.
Удовиченко М.И. Еласмобранхiï мандрикiвських верств Днiпропетровська. 1. Squatiniformes, Heterodontiformes, Orectolobiformes, Lamniformes // Палеонтол. зб. № 41. Лвiв: ЛНУ, 2009. С. 27–36.
Adolfssen J.S., Ward D.J. Neoselachians from the Danian (Early Paleocene) of Denmark // Acta Palaeontol. Pol. 2013. V. 60. № 2. P. 313–338.
Agassiz L.J.R. Recherches sur les Poissons Fossiles. T. 3. Neuchâtel, 1833–1844. 390 p.
Applegate S.P. Tooth terminology and variation in sharks with special reference to the sand shark, Carcharias taurus Rafinesque// Contrib. in Sci. Los Angeles Cty Museum. 1965. № 86. 18 p.
Arambourg C. Les Vertebres Fossiles des Gisements de Phosphates (Maroc–Algerie–Tunesie) // Notes Mém. Serv. Mines Carte Géol. Maroc. 1952. V. 92. 372 p.
Baut J.-P., Genault B. Contribution a l’étude des Elasmobranches du Thanetien (Paléocène) du Bassin de Paris. 1. Découverte d’une faune d’Elasmobranches dans la partie supèrieure des Sables de Bracheux (Thanetien, Paléocène) des regions de Compiegne (Oise) et de Montdidier (Somme) // Belg. Geol. Surv. Prof. Pap. 1995. № 278. P. 185–260.
Casier E. La Faune Ichthyologique de l’Ypresien de la Belgique // Mém. Mus. Roy. Hist. Natur. Belg. 1946. № 104. 267 p.
Cappetta H. Chondrichthyes II, Mesozoic and Cenozoic Elasmobranchii. Stuttgart, 1987. 193 p. (Handbook of Paleoichthyology. V. 3B).
Cappetta H. Elasmobranchii Post-Triadici (Index specierum et generum). Leiden: Backhuys Publ., 2006. 142 p.
Cappetta H. Mesozoic and Cenozoic Elasmobranchii: Teeth. München, 2012. 512 p. (Handbook of Paleoichthyology. V. 3E. Chondrichthyes / Ed. Schultze H.P.).
Cappetta H., Nolf D. Révision de quelques Odontaspididae (Neoselachii: Lamniformes) du Paléocène et de l’Éocène du Bassin de la Mer du Nord // Bull. Inst. Roy. Sci. Natur. Belg. Sci. Terre. 2005. V. 75. P. 237–266.
Case G.R., Udovichenko N.I., Nessov L.A. et al. A Middle Eocene selachian fauna from the White Mountain Formation of the Kizylkum Desert, Uzbekistan, C.I.S. // Palaeontogr. Abt. A. 1996. Bd 242. Lfg. 4–6. P. 99–126.
Cunningham S.B. A comparison of isolated teeth of early Eocene Striatolamia macrota (Chondrichthyes, Lamniformes), with those of a Recent sand shark, Carcharias taurus // Tertiary Res. 2000. V. 20. № 1–4. P. 17–31.
Human B.A., Owen E.P., Compagno L.J.V., Harley E.H. Testing morphologically based phylogenetic theories within the cartilaginous fishes with molecular data, with special reference to the catshark family (Chondrichthyes; Scyliorhinidae) and the interrelationships within them // Mol. Phylog. Evol. 2006. V. 39. P. 384–391.
Kriwet J. Additions to the Eocene selachian fauna of Antarctic with comments on Antarctic selachian diversity // J. Vertebr. Paleontol. 2005. V. 25. № 1. P. 1–7.
Leriche M. Les poissons paléocènes de la Belgique // Mém. Mus. Roy. Hist. Natur. Belg. 1902. V. 22. P. 1–48.
Malyshkina T.P., Ward D.J. The Turanian Basin in the Eocene: the new data on the fossil sharks and rays from the Kyzylkum desert (Uzbekistan) // Proc. Zool. Inst. RAS. 2016. V. 320. № 1. P. 50–65.
Martin A.P., Pardini A.T., Noble L.R., Jones C.S. Conservation of a dinucleotide simple sequence repeat locus in sharks // Mol. Phylog. Evol. 2002. V. 23. P. 205–213.
Müller J., Henle J. Systematische beshreibung der plagiostomen. Berlin: Veit & Co., 1838–1841. P. 1–38 (1838), p. 39–102 (1840), p. 103–200 (1841).
Naylor G.J.P., Caira J.N., Ensen K.J. et al. A DNA sequence-based approach to the identification of shark and ray species and its implications for global elasmobranch diversity and parasitology // Bull. Amer. Mus. Natur. Hist. 2012. V. 367. P. 1–262.
Naylor G.J.P., Martin A.P., Mattison E., Brown W.M. The inter-relationships of lamniform sharks: Testing phylogenetic hypotheses with sequencedata // Molecular Systematics of Fishes / Eds. Kocher T.D., Stepien C. N.Y.: Acad. Press, 1997. P. 199–218.
Ogg J.C., Ogg G., Gradstein F. The Concise Geologic Time Scale. Amsterdam: Elsevier, 2016. 240 p.
Reinecke T., Englehard P. The selachian fauna from the Geschiebe of the Lower Selandian Basal Conglomerate in the Danish Subbasin // Erratica. Wankendorf. 1997. V. 2. P. 3–45.
Siverson M. Revision of the Danian cow sharks, sand tiger sharks, and goblin sharks (Hexanchidae, Odontaspididae, and Mitsukurinidae) from Southern Sweden // J. Vertebr. Paleontol. 1995. V. 15. № 1. P. 1–12.
Trif N., Codrea V., Arghius A. A fish fauna from the lowermost Bartonian of the Transylvanian Basin, Romania // Palaeontol. Electron. 2019. 22.3.56. P. 1–29. https://doi.org/10.26879/909
Vélez-Zuazo X., Agnarsson I. Shark tales: A molecular species-level phylogeny of sharks (Selachiomorpha, Condrichthyes) // Mol. Phylog. Evol. 2011. V. 58. P. 207–217.
Ward D.J. The distribution of cartilaginous fish, turtles, birds and mammals in the British Palaeogene // Tertiary Res. 1980. V. 3. № 1. P. 1–21.
Ward D.J., Wiest R. A checklist of Palaeocene and Eocene sharks and rays (Chondrichthyes) from the Pamunkey Group, Maryland and Virginia, USA // Tertiary Res. 1990. V. 12. № 2. P. 81–88.
Welton B.J., Zinsmeister W.J. Eocene neoselachians from the La Meseta Formation, Seymour Island, Antarctic Peninsula // Contrib. in Sci. Los Angeles Cty Mus. 1980. № 329. P. 1–10.
White E.I. From the Thanet Sands to the Basement Bed of the London Clay // The Vertebrate Faunas of the English Eocene. V. 1. L.: Brit. Mus. (Natur. Hist.), 1931. P. 1–123.
Winkler T.C. Memoire sur des dents de poissons du terrain bruxellien // Arch. Mus. Teyler. Haarlem. 1874. V. 3. P. 295–304.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал