

Палеонтологический журнал, 2021, № 2, стр. 42-51
Новые данные по семейству Siricidae (Hymenoptera, Symphyta) в балтийском янтаре
А. Р. Манукян a, *, А. В. Смирнова a, **
a Калининградский музей янтаря
Калининград, Россия
* E-mail: manukyan@list.ru
** E-mail: smirnit@gmail.com
Поступила в редакцию 24.04.2020
После доработки 14.05.2020
Принята к публикации 14.05.2020
Аннотация
Из эоценового балтийского янтаря описан вид Xeris dorbnikensis Manukyan, sp. nov. (Hymenoptera, Siricidae); ранее отсюда семейство Siricidae было известно лишь видом Eoxeris klebsi (Brues, 1926). Приведены также сведения о другой находке рогохвоста рода Xeris из балтийского янтаря без уточнения видового статуса. Выявлены фаунистические связи между сообществами эоценового балтийского янтаря и современными гималайскими биологическими комплексами. Дан обзор палеонтологической истории семейства Siricidae.
Сидячебрюхие перепончатокрылые (Hymenoptera, Symphyta) насекомые в целом, и семейство Siricidae в частности, имеют важное значение для понимания эволюционной истории отряда Hymenoptera (Расницын, 1980; Klopfstein et al., 2013 и др.). Cреди современных Siricidae Р. Бэнсон (Benson, 1943) выделял подсемейства Siricinae (роды Sirex L., Xeris Costa, Siricosoma Forsius, Urocerus Geoffroy и Xoanon Semenov) и Tremecinae (роды Teredon Norton, Tremex Jurine, Eriotremex Benson и Afrotremex Pasteels). Несколько измененную родовую комбинацию приводят Я. Голд и Л. Моунд (Gauld, Mound, 1982) – род Siricosoma ими был перенесен в состав Tremecinae и противопоставлялся как сестринский таксон к подсемейству Siricinae (роды Xeris, Sirex, Xoanon и Urocerus). В составе Siricinae был описан род Neoxeris из горных районов Индии (Saini, Singh, 1987), позже сведенный в синонимы (Schiff et al., 2012) к роду Xeris. Для рецентных сирицид в мировой фауне в настоящее время известно 10 родов и около 122 видов (Schiff et al., 2012).
Надсемейство Siricoidea, помимо рецентного Siricidae и условно рецентного Anaxyelidae (в современной фауне только один вид Syntexis libocedrii Rohwer, 1915), включает также ископаемое семейство Pseudosiricidae (=Myrmiciidae) (Krogmann, Nel, 2012; Wedmann et al., 2014). Сюда же помещалось сем. Daohugoidae, которое позже было перенесено в состав Xyelidae в ранге подсемейства (Wang et al., 2019). Ранее к надсемейству Siricoidea также относили верхнеюрско-нижнемеловое семейство Praesiricidae, которое позже (Taeger et al., 2010; Wang et al., 2016) было перенесено в состав Pamphilioidea.
Монофилетический характер рецентного подсемейства Siricinae был показан по морфологическим данным (Königsmann, 1977; Rasnitsyn, 1988; Schulmeister, 2003) и позже был подтвержден генетико-молекулярными исследованиями (Heraty et al., 2011; Sharkey et al., 2012; Ronquist et al., 2012; Klopfstein et al., 2013; Malm, Nyman, 2015). Относительно недавно (Schiff et al., 2012) была проведена ревизия рецентных Siricidae и предложена новая группировка родов в подсемействах Siricinae и Tremecinae, основанная на признаках жилкования переднего крыла: роды Sirex, Sirotremex, Urocerus и Xoanon были включены в состав Siricinae, а роды Afrotremex, Eriotremex, Siricosoma, Teredon, Tremex и Xeris – в подсемейство Tremecinae.
Деление на основе жилкования крыла следует считать ненадежным из-за нестабильности признаков – существенные различия проявляются даже в крыльях у одних и тех же особей (Wedmann et al., 2014). Различия в биологии подсемейств также незначительны (Archibald, Rasnitsyn, 2016), поэтому деление сем. Siricidae s. s. на подсемейства следует считать необоснованным.
Современные сирициды – сравнительно крупные перепончатокрылые; тело цилиндрическое, преимущественно 15–45 мм длиной. Яйцеклад сверловидный, массивный, длиной до 45 мм, используется для просверливания коры и древесины при откладке яиц в ослабленные деревья. Самки характеризуются нитевидными, на вершине слабо уплощенными 14–25-члениковыми усиками. Преимущественно бореально-неморальная группа, связанная с хвойными и лиственными лесами Северного полушария. Небольшое число видов, эндемиков и завезенных, известно в Мексике, Кубе и Доминиканской Республике. Сведения о находках в Центральной Америке, Папуа – Новой Гвинее и в Африке к югу от Сахары базируются или на недостоверных сообщениях, или являются следствием завоза с древесным материалом за пределы естественного ареала (Smith, 1988; Schiff et al., 2012).
Рогохвосты заселяют ослабленные или усыхающие деревья, когда стволы заселяются жуками-усачами (Coleoptera, Cerambycidae) (церамбицидная стадия; Мамаев, 1960). Яйца откладываются на глубину 10–20 мм, при этом самки вместе с каждым яйцом вносят в древесину также споры дереворазрушающих эндосимбионтов – базидиомикотных (Basidiomycota) грибов Amylostereum areolatum Boidin, 1958 и A. chailletii (Persoon, 1822), а также секрет, который снижает устойчивость дерева к грибу. Базидиомикотные грибы вырабатывают фермент, который разрушает целлюлозу, трансформируя ее в доступные для питания личинки вещества. В результате совместного действия гриба и секрета гибель дерева ускоряется (Мамаев, Кравченко, 1973; Spradbery, 1973). Рогохвосты более обычны в старых хвойных лесах. В Европе встречаются c июня до ранней осени. Самцы большую часть жизни проводят в кроне деревьев, где и происходит спаривание. Самок часто можно обнаружить близ свежесрубленных деревьев. Живут несколько дней, не питаются, затраты энергии обеспечивают за счет накопленных запасов жира. Личинка в благоприятные годы развивается 2–4 года, при неблагоприятных условиях (например, в сухой древесине) развитие может продлиться еще на 2–4 года. Ходы проникают в глубину ствола на 10–15 см. Окукливание происходит в колыбельках чуть ниже камбия в 1.2–4.0 см от поверхности ствола.
Представители ископаемых сирицид и родственных групп (надсем. Siricoidea) известны с ранней юры, продвинутые Siricidae в характерном облике появились не позже конца поздней юры (Расницын, 1980). Cудя по строению яйцеклада, уже тогда они были связаны с мертвой древесиной (Rasnitsyn, 2002).
Т. Остен (Osten, 2007, с. 351, цит. по: Wedmann et al., 2014) приводит рисунки хорошо сохранившегося экземпляра неописанного вида (подсем. Siricinae) из нижнемелового (апт) местонахождения Крато (Бразилия). Систематическое положение этого вида как представителя семейства Pseudosiricidae было определено С. Арчибальдом и А.П. Расницыным (Archibald, Rasnitsyn, 2016).
Из формации Исянь (Yixian; нижний мел) Китая был описан вид Sinosirex gigantea Hong, 1975 (местонахождение Weichang, Хэбэй), из той же формации на северо-востоке (местонахождение Dawangzhangzi, Ляонин) М. Ван с соавт. (Wang et al., 2018) описали вид Cretosirex xiaoi. Два экземпляра не названных сирицид, сходных с Xeris и Eoxeris Maa, 1949, и новый род и вид сирикоида неясного положения были отмечены из меловых отложений бассейна р. Хетаны (Россия, Западное Приохотье) (Расницын, 1993).
Современные роды сирицид впервые достоверно отмечаются в палеогене: Urocerus ligniticus (Piton, 1940) из палеоцена Menat (Франция) и U. patagonicus Fidalgo et Smith, 1987 из нижнего эоцена бассейна р. Чубут (юг Аргентины). Вид U. ligniticus известен по 4 экз., что необычно для кайнозойских рогохвостов. Вымерший род и вид Ypresiosirex orthosemos Archibald et Rasnitsyn, 2016 описан из нижнего эоцена Западной Канады.
Из среднего эоцена Месселя (Германия) С. Ведманн с соавт. (Wedmann et al., 2014) по фрагменту переднего крыла описали Xoanon? eocenicus Wedmann et al., 2014, условно отнесенный к восточноазиатско-ориентальному (Сундуков, Лелей, 2012, с. 112) роду Xoanon.
Вид Eoxeris klebsi (Brues, 1926) был описан из балтийского янтаря по единственному экземпляру самца в составе рода Urocerus, позже вид был перенесен в ископаемый род Eoxeris Maa, 1949. В описании Ч. Брюса (Brues, 1926) приводится лишь рисунок переднего крыла; тип, хранившийся в коллекции Кёнигсбергского ун-та, ныне утрачен, поэтому истинный статус вида трудно определим. Новые находки сирицид в балтийском янтаре обсуждаются в настоящей статье.
В окаменевшей древесине хвойного дерева (предположительно Cupressaceae) из олигоценовых менилитовых слоев (Польша, Восточные Карпаты) обнаружены ихнофоссилии личинок сирицид – ходы и лётные отверстия (Rajchel, Uchman, 1998).
Ведманн описала из верхнего олигоцена Эншпель (Enspel, Германия) рогохвоста неясного систематического положения, близкого к родам Eriotremeх и Afrotremex, а также Tremex sp. из плиоцена Виллерсхаузена (Германия) (Wedmann, 1998). Из миоцена Франции (Montagne d’Andance, Saint-Bauzile) описан вид Proximoxeris tripoliensis Nel, 1991 (=P. tripoliensis Riou, 1999, объективный синоним).
Виды Formicium brodiei Westwood, 1854 и Megapterites mirabilis Cockerell, 1920 были описаны как рогохвосты по крыльям из эоцена Англии (Cockerell, 1920, 1921), но оказались гигантскими муравьями (Lutz, 1986, 1990).
МАТЕРИАЛ И МЕТОДИКА
В статье принята терминология по: Тобиас, 1978; Goulet, Huber, 1993; Goulet et al., 2015; жилкование крыльев – по: Расницын, 1969, 1980; Archibald, Rasnitsyn, 2016. Использованные в работе обозначения жилок показаны на рис. 1, 2 . Все размеры даны в мм. Исследованные образцы балтийского янтаря происходят из калининградского (Приморского) месторождения. Материал субаэрального происхождения не подвергался предварительной химической и физической обработке11. Весь исследованный материал хранится в коллекции Калининградского музея янтаря (КМЯ). Возраст балтийского янтаря нами принимается как приабонский (верхний эоцен), который был определен по остаткам фораминифер и палинологическим комплексам во вмещающих отложениях (Александрова, Запорожец, 2008) и подтвержден сравнительно-энтомологическими исследованиями (Perkovsky et al., 2007). Фотографии подготовлены с помощью фотокамеры Canon EOS 6D, соединенной со стереомикроскопами Leiсa, МС-2ZOOM, для увеличения глубины резкости использовалось программа Helicon Focus 6.7.1.

СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
НАДСЕМЕЙСТВО SIRICOIDEA BILLBERG, 1820
СЕМЕЙСТВО SIRICIDAE BILLBERG, 1820
Род Xeris Costa, 1894
Xeris dorbnikensis Manukyan sp. nov.
Табл. IV , фиг. 1–6
Название вида – от названия пос. Дорбникен, входящего в состав Пальмникенского округа (в настоящее время район пос. Янтарный), где находятся значительные пока не освоенные запасы балтийского янтаря.
Объяснение к таблице IV
Фиг. 1–6 . Хeris dorbnikensis Manukyan, sp. nov., голотип, КМЯ, № 5318: 1 – расположение в камне; 2 – голова сбоку; 3 – лапка средней ноги; 4 – вершина брюшка, яйцеклад и роговидный отросток; 5 – общий вид слева; 6 – переднее крыло.

Голотип – КМЯ, № 5318, самка; балтийский янтарь, верхний эоцен. Камень размером 45 × 15 × 5 мм. Целое насекомое, отсутствуют (повреждены при механической обработке) вершина бедра правой средней ноги, основание правой средней голени и лапки задних ног. Брюшко полностью, грудь слева частично покрыты белой светонепроницаемой пленкой. Правое заднее крыло плотно прилегает к брюшку, закрывая его сверху. Сининклюзы: Arachnida, Acari; Collembola, Arthropleona; волоски дуба более или менее равномерно распределены по всему объему камня.
Описание (рис. 1). Длина тела около 15.0 мм (табл. IV , фиг. 1). Наличник с глубокой вырезкой посередине, двулопастный. Верхняя губа массивная, отчетливо выступает из-под наличника. Ширина ротовой выемки приблизительно в 3 раза больше диаметра глазка. Мандибулы, судя по строению, не смыкаются, короткие, их длина лишь в 1.3 раза больше ширины в основании; двузубые, нижний зубец длиннее верхнего более чем в 2 раза. Внутренние орбиты глаз широкие, выступают над лицом. Лицо в густой пунктировке, пунктирные ямки глубокие, в густом и длинном опушении, волоски направлены вперед.
Минимальное расстояние между глазами (измеренное на вершине глаз) в 1.2–1.3 раза больше высоты глаза. Расстояние между внутренними краями усиковых ямок приблизительно вдвое больше, чем кратчайшее расстояние от наружного края усиковых ямок до глаза. Глаза едва овальной формы, соотношение высота/ширина 5 : 4; выпуклые, значительно выступают над поверхностью головы.
Кратчайшее расстояние от латерального глазка до глаза (OOL) в два раза больше, чем расстояние от среднего до заднего глазка. Расстояние от заднего глазка до глаза чуть больше расстояния между латеральными глазками. Расстояние от заднего глазка до заднего края головы (OCL) более чем в 4 раза больше, чем расстояние от заднего глазка до глаза (OOL). Вдоль заднего края глаз имеется четко разграниченная вдавлением узкая площадка в резкой, глубокой и густой пунктировке (табл. IV , фиг. 2 ). Темя посередине с бороздой; поверхность, также теменная площадка и лобное поле в густой, грубой сотообразно-ячеистой скульптуре. Виски с едва намеченным вертикальным валиком (гребнем), в редких, но глубоких вдавлениях (pits), поверхность между вдавлениями гладкая, блестящая. Затылочный валик слабый. Щеки с пучком длинных, торчащих волосков; длина щеки (кратчайшее расстояние от нижнего края глаз до основания мандибулы) составляет около половины высоты глаза. Скапус едва изогнутый и слабо утолщенный посередине (табл. IV , фиг. 2 ). Жгутик усика 17-члениковый, плавно нитевидно утончается в верхней половине, членики цилиндрические.
Пронотум соединен со среднеспинкой неподвижно, верхне-боковые края вытянуты вперед в виде лопастей, передний край между ними выемчатый; воротничок отчетливо утолщенный, по краю окаймлен валиком; заднебоковые углы достигают тегул. Простернум разделен по средней линии отчетливым швом.
Среднеспинка короткая, ее длина около 2.0 мм; цельная (нотаулы отсутствуют), умеренно выпуклая, кзади слабо сужена. Поверхность матовая, в мелкобугристой скульптуре; в густом опушении, волоски тонкие. Мезоплевры слабо выпуклые, в густой пунктировке. Щитик в мелкой пунктировке, в густом, торчащем опушении. Заднещитик узкий, его ширина чуть больше, чем ширина щитика в основании.
Передние крылья (рис. 1) дымчато-затемненные (табл. IV , фиг. 6 ); длина от вершины до тегул 12.1 мм; стройные – длина в 3.6–3.7 раза больше максимальной ширины. Дистальная половина крыла продольно-гофрированная; в базальной половине ячейки 1cua и 1rm поперечно-гофрированные. Птеростигма слабо склеротизованная. RS+M явственная, 1r-m отсутствует. Жилка 2r-m вертикальная, 2m-cu инкливальная; длина отрезка Rs за поперечной жилкой 3r-m около 0.4 мм. Ячейка 1rm выпуклая, выступает над поверхностью крыла, гофр более тонкий, чем на ячейке 1cua. Жилка 3A отсутствует. Задние крылья прозрачные, длина около 7.5 мм, передний край с 20–21 зацепками, крупные зацепки чередуются с более мелкими; анальная ячейка открытая.
Брюшко полностью покрыто светонепроницаемой белой пленкой, контуры сегментов хорошо просматриваются (табл. IV , фиг. 1, 4, 5 ). Длина ножен яйцеклада 10.6 мм. Роговидный отросток 2.6 мм, копьевидный, плавно сужается к вершине (табл. IV , фиг. 4 ), без срединного сужения; ширина в основании 1.5, соотношение ширина в основании/длина – 1.7; дорсальная поверхность апикальной трети с небольшим зубцом, вентральная – с двумя рядами резких зубцов.
Бедра передних и средних ног цилиндрической формы, слабо утолщенные в базальной половине; бедра задних ног сжаты с боков, короткие, длина – 1.2 мм, максимальная ширина лишь в 2 раза меньше длины. Все голени цилиндрические (не сплюснутые с боков) с одной шпорой, передние и средние голени слабоизогнутые, задняя прямая; вершина голеней напротив шпор с глубокой вырезкой. Первые членики лапок передних и средних ног удлиненные, их длина равна длине голени: длина первого членика передней лапки 1.8 мм, средней – 2.0 мм. Четвертый членик лапки короткий (длина чуть больше 0.3 мм), на вершине с вентральной стороны лопастевидно выступающий. Коготки передних и средних ног острые, серповидные, с базальным зубцом (табл. IV , фиг. 3 ).
Тело черное, ноги за тазиками красные (окраска поврежденных участков ног неизвестна); окраска брюшка неопределима.
Самец неизвестен.
Сравнение. Вид отнесен к роду Xeris в первую очередь по следующим признакам: 1) наличие валика (гребня) на висках; 2) жилка 1r-m в переднем крыле отсутствует; 3) членики жгутика цилиндрические, не уплощенные на дистальной половине; 4) голени средних ног с одной шпорой; 5) задние крылья с открытой анальной ячейкой; 6) глаза овальные (соотношение длина/ширина 5 : 4); 7) видимые части тела черной окраски; 8) соотношение ширина в основании/длина роговидного отростка равно 1.7.
По признакам окраски (черные жгутики и тазики), наличию резкой и глубокой пунктировки по заднему краю глаз сближается с видом X. himalayensis Bradley, 1934 (=X. melanocephalus Saini et Singh, 1987; =X. indianus Vesu et Saini, 1999), отличия показаны ниже:
Тело около 15 мм. Жгутик 17-члениковый. Задний край глаз с четко разграниченной вдавлением площадкой, где пунктировка густая и глубокая. Наличник глубоко вырезан посередине, двулопастный. Мандибулы не смыкаются. Длина переднего крыла в 3.6–3.7 раза больше максимальной ширины. Cрединное сужение роговидного отростка отсутствует…..………….……… .........................................X. dorbnikensis sp. nov.
– Тело 20.2–20.8 мм. Жгутик 23-члениковый. Задний край глаз с площадкой в густой и глубокой пунктировке, не отграниченной вдавлением. Наличник с неглубокой вырезкой посредине, не двулопастный. Мандибулы смыкаются. Длина переднего крыла в 3.4 раза больше максимальной ширины. Cрединное сужение роговидного отростка имеется. ……………….…… .....................................................X. himalayensis
Замечания. Отличается от описанного из балтийского янтаря Eoxeris klebsi наличием только одной шпоры средней и задней голеней. Жгутик усика Х. dorbnikensis sp. nov. отличается меньшим количеством члеников (17), у единственного известного экземпляра самца E. klebsi усики 21‑члениковые. Возможно, что меньшее количество члеников обусловлено половым диморфизмом, который проявляется также цилиндрической (не сплюснутой) формой голеней задних ног. Обнаружены следующие различия в жилковании крыльев: жилка 2r-m вертикальная, у E. klebsi инкливальная, посередине изогнутая; жилка 2m-cu инкливальная, у E. klebsi вертикальная; отрезок Rs за поперечной жилкой 3r-m у X. dorbnikensis почти в два раза длиннее, чем тот же отрезок у E. klebsi. Возможно, что эти виды отличаются также строением крыловой пластины: судя по рисунку к описанию вида E. klebsi, дистальная половина переднего крыла отчетливо продольно-гофрирована, поверхность ячеек 1rm и 1cua гладкая, в отличие от Х. dorbnikensis, у которого последние ячейки поперечно-морщинистые (гофрированные) (рис. 1). Сходное с Х. dorbnikensis строение ячеек 1rm и 1cua характерно для ископаемого вида Ypresiosirex orthosemos.
Примечательны сравнительно мелкие размеры тела X. dorbnikensis sp. nov. – около 15 мм, примерно такие же размеры у Eoxeris klebsi. Для других ископаемых сирикоид характерны крупные размеры тела: Sinosirex gigantea – около 40, Ypresiosirex orthosemos – 59.3 мм. При этом новый вид укладывается в рамки средних размерных вариаций для рецентных представителей рода Xeris, которые при крайних выражениях имеют размеры от 9 до 45 мм.
* * *
Xeris sp., экз. КМЯ, № 5656, самка (табл. V , фиг. 1–3 ), сохранилась в слоистом куске янтаря размерами 72 × 25 × 8 мм; сининклюз: фрагмент древесного детрита. Экземпляр плохой сохранности; просматриваются лишь отдельные детали тела. Верхняя половина передних крыльев была сточена при механической обработке янтаря. Тело покрыто белой пленкой и слоями янтаря. Правое переднее и заднее крылья, передние и средние ноги и бедра задних ног плотно прилегают к брюшку, покрыты белым налетом. Ниже приводится описание лишь тех частей тела, которые доступны исследованию.
Объяснение к таблице V
Фиг. 1–3 . Xeris sp., экз. КМЯ, № 5656: 1 – расположение в камне; 2 – общий вид справа; 3 – вершина брюшка и яйцеклад.
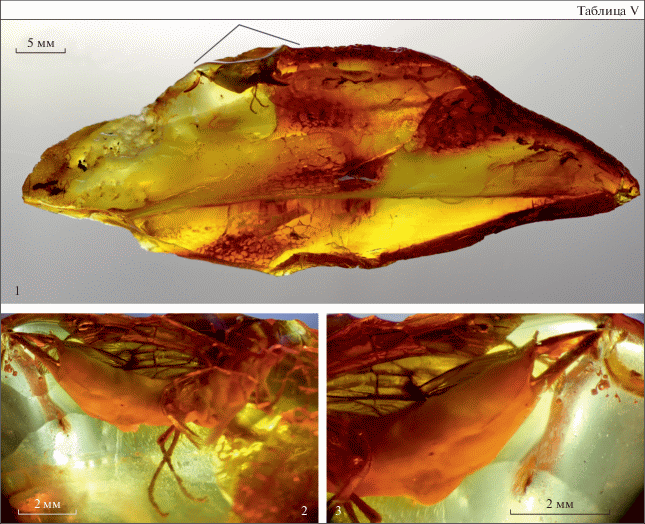
Длина тела около 11.5 мм (табл. V , фиг. 1, 2 ). Жгутик усика 11-члениковый, плавно нитевидно утончается в верхней половине, членики цилиндрические, последний (11-й) членик маленький, конусовидный. Голени передних и задних ног с одной шпорой; голени cредних ног не просматриваются.
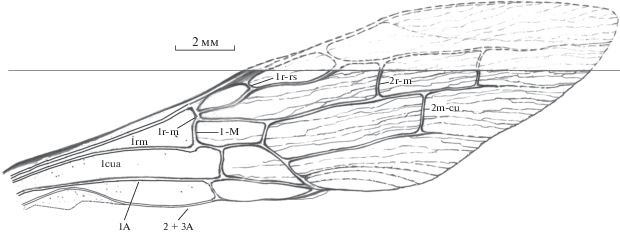
Передние крылья прозрачные; длина от вершины до тегул около 7.5 мм. Крыловая пластина гофрированная только в дистальной половине, ячейки 1cua и 1rm негофрированные. Жилка 1r-m имеется (рис. 2 ).
Брюшко полностью покрыто светонепроницаемой белой пленкой, контуры сегментов едва просматриваются (табл. V , фиг. 2 ). Длина ножен яйцеклада около 7.0 мм; роговидный отросток 1.0 мм, копьевидный, плавно сужается к вершине, без срединного сужения (табл. V , фиг. 3 ); ширина в основании 0.7 мм, соотношение ширина в основании/длина – 1.4 мм; дорсальная поверхность в единичных щетинках, вентро-латеральная – не менее чем с двумя рядами резких зубцов.
Задние ноги. Бедра сжаты с боков, их длина – 1.2 мм, максимальная ширина лишь в 2 раза меньше длины. Голень прямая, вершина напротив шпоры с глубокой вырезкой. Длина 1-го членика лапки 1.5 мм; 4-й членик лапки короткий, длина чуть более 3 мм, лопастевидно выступающий на вершине с вентральной стороны; коготки острые, загнутые под прямым углом.
На основании одной шпоры на задней голени экземпляр может быть лишь условно отнесен к роду Xeris, хотя главный диагностический признак рода – наличие вертикального валика на висках – не просматривается из-за плохой сохранности. По мелким размерам тела и меньшему количеству члеников жгутика усика может быть сближен к X. dorbnikensis; отличается гладкими (негофрированными) ячейками 1cua и 1rm переднего крыла и наличием жилки 1r-m. От E. klebsi отличается наличием только одной шпоры на голенях задних ног, мелкими размерами и меньшим количеством члеников жгутика.
ОБСУЖДЕНИЕ
В мировой фауне известно 16 видов рода Xeris, все из Голарктики. В Неарктике и Палеарктике обитает равное число видов – по восемь (Goulet et al., 2015). Из них только пять видов выходят за пределы южных границ Голарктики, при этом все отмечались в хвойно-широколиственном поясе горных районов: X. himalayensis встречается вдоль Гималаев от Пакистана до Непала на высоте от 1700 до 3000 м над уровнем моря; X. malaisei Maa, 1949, встречается в Приморском крае России, на северо-востоке Китая (Гирин), в Японии (Хоккайдо, Хонсю) и высокогорьях Южной Кореи (Goulet et al., 2015); X. umbra Goulet, 2015 и X. xanthoceros Goulet, 2015, Китай (Юньнань), встречается на высоте до 4000 м; X. xylocola Goulet, 2015, Лаос (Хуапхан), на высоте 1350–1900 м; X. tropicalis Goulet, 2012 известен по единичным находкам из Мексики (Чьяпас).
Весьма примечательны результаты молекулярно-генетических исследований (Goulet et al., 2015) – кладограмма, построенная на основе анализа митохондриальной ДНК, показывает изолированное положение вида X. himalayensis, при этом все остальные восемь изученных видов рода Xeris противопоставляются как сестринская группа к виду X. himalayensis.
Биология рода Xeris известна по трем видам – X. caudatus, X. malaisei и X. spectrum. Наиболее примечательной особенностью биологии этих видов является отсутствие у имаго самок симбиотических грибов, необходимых для развития личинки (Fukuda, Hijii, 1997; Schiff et al., 2012). Тем не менее, эти грибы всегда присутствуют в ходах личинок X. malaisei и X. caudatus. Наблюдения также показали, что виды рода Xeris всегда откладывают яйца в деревья, уже зараженные другими рогохвостами (в частности, из родов Sirex и Urocerus), летные отверстия также всегда находятся в непосредственной близости от отверстий других рогохвостов (Fukuda, Hijii, 1997). Предполагается, что самки рода Xeris в поиске подходящих для заражения деревьев ориентируются по запаху грибов Amylostereum spp.
Все виды рода развиваются исключительно на хвойных деревьях (Cupressaceae и Pinaceae), для вида X. himalayensis отмечались гималайские виды Abies pindrow (пихта гималайская), Cedrus deodara (кедр гималайский), Picea smithiana (ель гималайская) и Pinus roxburghii (индийская или гималайская сосна).
Новые сведения о фауне сирицид балтийского янтаря свидетельствуют о фаунистических связях эоценовой балтийской биоты с гималайской. Совершенно определенные сведения ранее были известны по эоценовым мхам (Frahm, 1999) и архаичным представителям жесткокрылых семейства Nitidulidae (Coleoptera) (Kirejtshuk, 2005; Кирейчук, 2017). К этой же группе следует отнести архаичных термитов рода Archotermopsis Desneux, 1904 (Isoptera, Archotermopsidae), которые известны по трем видам; из балтийского янтаря описан A. tornquisti Rosen, 1913, близкие современные виды распространены в Гималаях [A. wroughtoni (Desneux, 1904)] и во Вьетнаме (A. kuznetsovi Beljaeva, 2004) (Engel et al., 2009; Krishna et al., 2013).
Из приведенных данных следует, что гипотеза А.Г. Кирейчука (2017) о том, что в гималайском регионе сохранились не только отдельные таксономические элементы “янтарных” биот, но и целые сообщества в слабо трансформированном виде, получает дополнительный аргумент.
Таким образом, замечания по биологии и тафономии сирицид в балтийском янтаре в обобщенном виде следующие:
1. Малая встречаемость сирицид в балтийском янтаре обусловлена в первую очередь крупными размерами, а также короткой продолжительностью жизни имаго.
2. Поскольку виды рода Xeris не обладают собственными эндосимбионтами, для их выживания необходимо было присутствие и других сирицид, что говорит о потенциальном богатстве фауны сем. Siricidae в «янтарном» сообществе.
3. Данные по сирицидам балтийского янтаря являются дополнительным свидетельством того, что в гималайском регионе сохранились не только позднепалеогеновые таксономические компоненты, но и, возможно, малоизмененные сообщества этих биот.
* * *
Авторы выражают признательность А.П. Расницыну (Палеонтологический ин-т им. А.А. Борисяка РАН) за неоценимую помощь в подготовке работы, а также коллекционерам С.И. Шишову (г. Калининград) и В.А. Гусакову (г. Москва) за передачу в фонды Калининградского музея янтаря образцов с включениями рогохвостов. Работа поддержана Российским фондом фундаментальных исследований (грант № 19-05-00207).
Список литературы
Александрова Г.Н., Запорожец Н.И. Палинологическая характеристика верхнемеловых и палеогеновых отложений Самбийского полуострова (Калининградская область). Статья 1 // Стратигр. Геол. корреляция. 2008. Т. 16. № 3. С. 75–96.
Кирейчук А.Г. Гималаи: регион реликтов и центр современных диверсификаций биоты на примере жуков-блестянок (Coleoptera, Nitidulidae) // Российские гималайские исследования: вчера, сегодня, завтра. Сборн. научн. статей / Ред. Боркин Л.Я. СПб.: Изд-во “Европейский дом”, 2017. С. 168–172.
Мамаев Б.М. Зоологическая оценка стадий естественного разрушения древесины // Изв. АН СССР. 1960. № 4. С. 610 –617.
Мамаев Б.М., Кравченко М.А. Личинки рогохвостов // Насекомые–разрушители древесины в лесных биоценозах Южного Приморья. М.: Наука, 1973. С. 74–85.
Расницын А.П. Происхождение и эволюция низших перепончатокрылых. М.: Изд-во АН СССР, 1969. 196 с. (Тр. Палеонтол. ин-та АН СССР. Т. 123).
Расницын А.П. Происхождение и эволюция перепончатокрылых насекомых. М.: Изд-во АН СССР, 1980. 192 с. (Тр. Палеонтол. ин-та AH CCCP. Т. 174).
Расницын А.П. Отряд перепончатокрылые (Hymenoptera) // Меловые энтомофауны бассейна р. Ульи (Западное Приохотье). Мезозойские насекомые и остракоды Азии. М.: Наука, 1993. С. 50–57.
Сундуков Ю.Н., Лелей А.С. Надсем. Siricoidea 10. Сем. Siricidae – рогохвосты // Аннотированный каталог насекомых Дальнего Востока России. Т. I. Перепончатокрылые. Владивосток: Дальнаука, 2012. С. 112–113.
Тобиас В.И. Отряд Hymenoptera – перепончатокрылые. Введение // Определитель Насекомых Европейской Части СССР. Т. III. Перепончатокрылые. I часть. Л.: Наука, 1978. С. 7–42 (Определители по фауне СССР. Вып. 119).
Archibald S.B., Rasnitsyn A.P. New early Eocene Siricomorpha (Hymenoptera: Symphyta: Pamphiliidae, Siricidae, Cephidae) from the Okanagan Highlands, western North America // Can. Entomol. 2016. V. 148. P. 209–228.
Benson R.B. Studies in Siricidae, especially of Europe and Southern Asia (Hymenoptera, Symphyta) // Bull. Entomol. Res. 1943. V. 34. P. 27–51.
Brues C.T. A species of Urocerus from Baltic Amber // Psyche. 1926. V. 33. P. 168–169.
Cockerell T.D.A. Fossil arthropods in the British Museum // Ann. Mag. Natur. Hist. 1920. V. 5. P. 273–279.
Cockerell T.D.A. Some British fossil insects // Can. Entomol. 1921. V. 53. P. 22–23.
Engel M.S., Grimaldi D., Krishna K. Termites (Isoptera): their phylogeny, classification, and rise to ecological dominance // Amer. Mus. Novit. 2009. № 3650. P. 1–27.
Frahm P. Die Laubmoosflora des Baltischen und Bitterfelder Bernsteins // Mitt. Geol.-Paläontol. Staatsinst. 1999. Bd 83. S. 219–238.
Fukuda H., Hijii N. Reproductive strategy of a woodwasp with no fungal symbionts, Xeris spectrum (Hymenoptera: Siricidae) // Oecologia. 1997. V. 112. P. 551–556.
Gauld H., Mound L.A. Homoplasy and the delineation of holophyletic genera in some insect groups // Syst. Entomol. 1982. V. 7. P. 73–86.
Goulet H., Boudreault C., Schiff N.M. Revision of the World species of Xeris Costa (Hymenoptera: Siricidae) // Can. J. Arthropod Identification. 2015. № 28. P. 1–101.
Goulet H., Huber J.T. Hymenoptera of the World: an Identification Guide to Families. V. 4. Ottawa: Agriculture Canada, Res. branch, 1993. 668 p.
Heraty J., Ronquist F., Carpenter J.M. et al. Evolution of the hymenopteran megaradiation // Mol. Phylog. Evol. 2011. V. 60. P. 73–88.
Kirejtshuk A.G. On the similarities between the Nitidulid faunas from Baltic amber and the Himalayas and mountains of South China (Coleoptera, Nitidulidae) // 2nd Intern. Symp. on Biodiversität und Naturausstattagung im Himalaya: Abstracts. Erfurt, 2005.
Klopfstein S., Vilhelmsen L., Heraty J.M. et al. The Hymenopteran tree of life: Evidence from protein-coding genes and objectively aligned ribosomal data // PLOS. 2013. V. 8. e69344.
Königsmann E. Das phylogenetische system der Hymenoptera. Teil 2 “Symphyta” // Dtsch. Entomol. Z. 1977. H. 24. P. 1–40.
Krishna K., Grimaldi D., Krishna V. et al. Treatise on the Isoptera of the world. 2. Basal families // Bull. Amer. Mus. Natur. Hist. 2013. V. 377. P. 252–258.
Krogmann L., Nel A. On the edge of parasitoidism, a new Lower Cretaceous woodwasp forming the putative sister group of Xiphydriidae+ Euhymenoptera // Syst. Entomol. 2012. V. 37. P. 215–222.
Lutz H. Eine neue Unterfamilie der Formicidae (Insecta: Hymenoptera) aus dem mittel-eozänen Ölschiefer der “Grube Messel” bei Darmstadt (Deutschland, S-Hessen) // Senckenb. lethaea. 1986. V. 67. S. 177–218.
Lutz H. Systematische und palökologische Untersuchungen an Insekten aus dem Mittel-Eozän der Grube Messel bei Darmstadt // Cour. Forsch.-Inst. Senckenb. 1990. V. 124. S. 1–165.
Malm T., Nyman T. Phylogeny of the symphytan grade of Hymenoptera, new pieces into the old jigsaw(fly) puzzle // Cladistics. 2015. V. 31. P. 1–17.
Perkovsky E.E., Rasnitsyn A.P., Vlaskin A.P. et al. A comparative analysis of the Baltic and Rovno amber arthropod faunas: representative samples // Afr. Invertebr. 2007. V. 48. № 1. P. 229–245.
Rajchel J., Uchman A. Insect borings in Oligocene wood, Kliwa sandstones, Outer Carpathians, Poland // Ann. Soc. Geol. Pol. 1998. V. 68. P. 219–224.
Rasnitsyn A.P. An outline of evolution of the hymenopterous insects (order Vespida) // Oriental Insects. 1988. V. 22. P. 115–145.
Rasnitsyn A.P. Superorder Vespidea Laicharting, 1781. Order Hymenoptera Linné, 1758 // History of Insects / Eds. Rasnitsyn A.P., Quicke D.L.J. Dordrecht: Kluwer Acad. Publ., 2002. P. 242–254.
Ronquist F., Klopfstein S., Vilhelmsen L. et al. A total evidence approach to dating with fossils, applied to the early radiation of the Hymenoptera // Syst. Biol. 2012. V. 61. P. 973–999.
Saini M.S., Singh D. A new genus and a new species of Siricinae (Insecta, Hymenoptera, Siricidae) from India with a revised key to its world genera // Zool. Scripta. 1987. V. 16. № 2. P. 177–180.
Schiff N.M., Goulet H., Smith D.R. et al. Siricidae (Hymenoptera: Symphyta: Siricoidea) of the Western Hemisphere // Can. J. Arthropod Identification. 2012. V. 21. 305 p.
Schulmeister S. Review of morphological evidence on the phylogeny of basal Hymenoptera (Insecta), with a discussion of the ordering characters // Biol. J. Linn. Soc. 2003. V. 79. P. 209–243.
Sharkey M.J., Carpenter J.M., Vilhelmsen L. et al. Phylogenetic relationships among superfamilies of Hymenoptera // Cladistics. 2012. V. 28. P. 80–112.
Smith D.R. A synopsis of the sawflies (Hymenoptera: Symphyta) of America south of the United States: introduction, Xyelidae, Pamphiliidae, Cimbicidae, Diprionidae, Xiphydriidae, Siricidae, Orussidae, Cephidae // Syst. Entomol. 1988. V. 13. P. 205–261.
Spradbery J.P. A comparative study of the phytotoxic effects of siricid woodwasps on conifers // Ann. Appl. Biol. 1973. V. 75. № 3. P. 309–320.
Taeger A., Blank S.M., Liston A.D. World Catalog of Symphyta (Hymenoptera) // Zootaxa. 2010. V. 2580. 1064 p.
Wang M., Rasnitsyn A.P., Han G. et al. A new genus and species of basal horntail (Hymenoptera, Siricidae) from the Lower Cretaceous of China // Cret. Res. 2018. V. 91. P. 195–201.
Wang M., Rasnitsyn A.P., Shih Ch. et al. New fossils from China elucidating the phylogeny of Praesiricidae (Insecta: Hymenoptera) // Syst. Entomol. 2016. V. 41. P. 41–55.
Wang M., Rasnitsyn A.P., Zhang H. et al. Revising the systematic position of the extinct family Daohugoidae (basal Hymenoptera) // J. Syst. Palaeontol. 2019. V. 17. № 14. P. 1245–1255.
Wedmann S. First records of fossil tremecine hymenopterans // Palaeontology. 1998. V. 41. P. 929–938.
Wedmann S., Pouillon J.-M., Nel A. New Palaeogene horntail wasps (Hymenoptera, Siricidae) and a discussion of their fossil record // Zootaxa. 2014. V. 3869. № 1. P. 33–43.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал