

Палеонтологический журнал, 2021, № 3, стр. 10-23
Радиолярии из класса Stauraxonaria позднего карбона–средней перми: Nazarovispongus Kozur sensu Kozur et Mostler, restituo et emend. nov. и новый род Kozurispongus gen. nov.
М. С. Афанасьева *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: afanasieva@paleo.ru
Поступила в редакцию 02.10.2020
После доработки 20.10.2020
Принята к публикации 27.10.2020
Аннотация
Анализ морфологических особенностей скелетов радиолярий из класса Stauraxonaria позднего карбона и ранней перми Южного Урала России и Западных Мугоджар Казахстана, а также средней перми Западного Техаса США позволил восстановить валидность рода Nazarovispongus Kozur, 1980 и пересмотреть систематическое положение видов N. pavlovi и N. permicus, установленных Х. Коцуром (Kozur, 1980). Дополнен диагноз восстановленного рода Nazarovispongus Kozur, 1980 sensu Kozur et Mostler, 1989, restituo et emend. nov., а видовой состав расширен до шести видов: N. pavlovi Kozur, 1980 (типовой вид), N. aequilateralis (Nazarov in Isakova et Nazarov, 1986, comb. nov.), N. delicatus (Nazarov et Ormiston, 1985, part., comb. nov.), N. globosus Nestell et Nestell 2010, N. inflatus (Nazarov et Ormiston, 1985, comb. nov.) и N. subrotundus (Nazarov in Isakova et Nazarov, 1986, comb. nov.). Описан новый род Kozurispongus gen. nov., объединяющий шесть видов: K. aktastiensis (Nazarov et Ormiston, 1993, comb. nov.), K. circumfusus (Nazarov et Ormiston, 1985, comb. nov.), K. permicus (Kozur, 1980, comb. nov.), K. semilaminus (Nazarov in Isakova et Nazarov, 1986, part., comb. nov.) и два новых вида, K. concavilatus sp. nov. и K. rotundatus sp. nov. Предложена новая схема последовательных этапов формирования игл скелета.
ВВЕДЕНИЕ
Детальное изучение раннепермских радиолярий Южного Урала и Западных Мугоджар потребовало обратить особое внимание и на общие вопросы систематики и таксономии двух родов ставраксонных радиолярий.
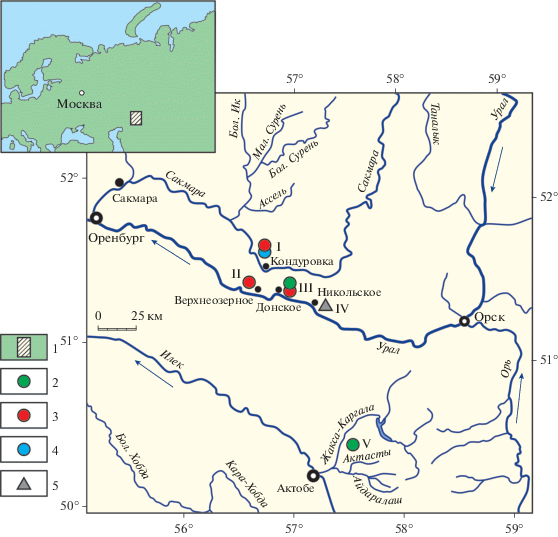
Морфологическое и таксономическое исследование радиолярий из отложений гжельского яруса верхнего карбона и ассельского, сакмарского и артинского ярусов нижней перми в опорных разрезах Кондуровский, Верхнеозерный, Никольский и Донское на Южном Урале России и Актасты в Западных Мугоджарах Казахстана (рис. 1) было осуществлено по литературным данным и на основании собственных наблюдений автора (Kozur, 1980; Nazarov, Ormiston, 1985, 1993; Исакова, Назаров, 1986; Афанасьева и др., 2014, 2015). Кроме этого, был привлечен литературный материал по радиоляриям из отложений роудского и кептенского ярусов средней перми Западного Техаса США (Nazarov, Ormiston, 1985; Nestell, Nestell, 2010).
Рис. 1.
Местонахождения радиолярий верхнего карбона и нижней перми на Южном Урале (I–IV) и Западных Мугоджарах (V): I – р. Сакмара, разрез Кондуровский (Назаров, 1984; Афанасьева и др., 2014); II–IV – р. Урал: II – разрез Верхнеозерный (Kozur, 1980; Назаров, 1984), III – разрез Донское (Назаров, 1984; Nazarov, Ormiston, 1985), IV – разрез Никольский (Назаров, 1984; Исакова, Назаров, 1986); V – Западные Мугоджары, разрез Актасты на р. Актасты (Назаров, 1984; Nazarov, Ormiston, 1993; Афанасьева и др., 2015). Обозначения: 1 – район исследований; 2–4 – нижняя пермь: 2 – артинский ярус, 3 – сакмарский ярус, 4 – ассельский ярус; 5 – верхний карбон, гжельский ярус.
Анализ морфологических особенностей скелетов ставраксонных радиолярий позволил восстановить валидность рода Nazarovispongus Kozur, 1980 sensu Kozur et Mostler, 1989, restituo et emend. nov., а его видовой состав расширить до шести видов (рис. 2).
Рис. 2.
Распространение видов из родов Nazarovispongus и Kozurispongus: 1–3 – Россия, Южный Урал: 1 – р. Урал, разрез Никольский (Назаров, 1984; Исакова, Назаров, 1986), 2 – р. Сакмара, разрез Кондуровский (Афанасьева и др., 2014), 3 – р. Урал, разрез Верхнеозерный (Kozur, 1980; Назаров, 1984), р. Урал, разрез Донское (Назаров, 1984; Nazarov, Ormiston, 1985), р. Сакмара, разрез Кондуровский (Назаров, 1984; Афанасьева и др., 2014); 4 – Казахстан, Западные Мугоджары, разрез Актасты на р. Актасты (Назаров, 1984; Nazarov, Ormiston, 1993; Афанасьева и др., 2015), Россия, Южный Урал, р. Урал, разрез Донское (Назаров, 1984); 5, 6 – США, Западный Техас: 5 – роудский ярус (Nazarov, Ormiston, 1985), 6 – кептенский ярус (Nazarov, Ormiston, 1985; Nestell, Nestell 2010). Обозначения ярусов: g – гжельский, as – ассельский, sk – сакмарский, ar – артинский, r –роудский, c – кептенский. Двойной линией отмечен перерыв во временнóм распространении видов.
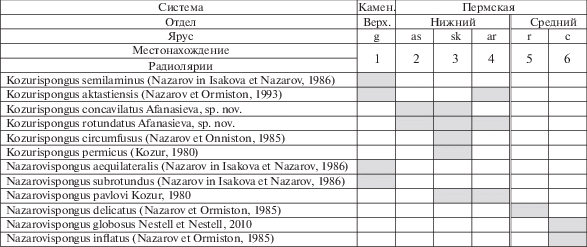
В результате исследований пересмотрено систематическое положение Nazarovispongus permicus, который в настоящее время рассматривается в составе нового рода Kozurispongus gen. nov. Новый род объединяет шесть видов, среди которых два являются новыми (рис. 2).
ТРЕУГОЛЬНЫЕ МОРФОТИПЫ СКЕЛЕТОВ РАДИОЛЯРИЙ
Исследование радиолярий проводилось под разными углами зрения: (1) анализ морфологических особенностей треугольных скелетов; (2) конструктивно-морфологические исследования.
Проведенное всестороннее изучение радиолярий позволило раскрыть основные особенности треугольных морфотипов скелетов и установить последовательные этапы формирования игл в скелете радиолярий.
Треугольные морфотипы скелетов
Анализ морфологических особенностей треугольных скелетов радиолярий позднего карбона и ранней перми Южного Урала России и Западных Мугоджар Казахстана, а также средней перми Западного Техаса США позволил установить четыре морфотипа, характеризующих четыре рода (рис. 3):
Рис. 3.
Схема строения скелетов радиолярий: а – Ruzhencevispongus Kozur, 1980 sensu Kozur et Mostler, 1989 (по Назаров, 1988, рис. 23 м); б – Ruzhencevispongus Kozur, 1980 sensu Nazarov et Ormiston, 1983 (по Nazarov, Ormiston, 1983, рис. 5); в – Nazarovispongus Kozur, 1980; г – Kozurispongus gen. nov.; д – Foremanhelena De Wever et Caridroit, 1984 (по Caridroit, De Wever, 1986, рис. 23 ).
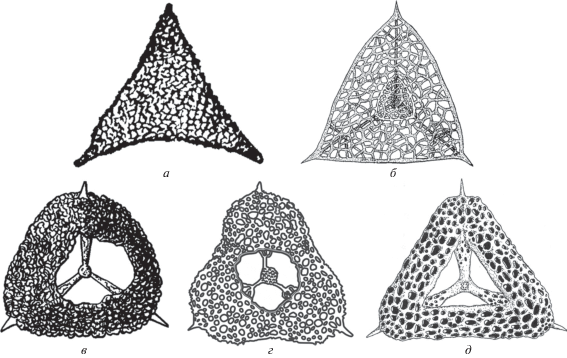
• Ruzhencevispongus Kozur, 1980 sensu Kozur et Mostler, 1989 (рис. 3, а; 4, а) [non Ruzhencevispongus Kozur, 1980 sensu Nazarov et Ormiston, 1983 (рис. 3, б)] отличается формированием треугольного скелета с прямыми или слегка вогнутыми боковыми и слегка выпуклыми верхней и нижней сторонами, а также закрытой центральной частью внешней оболочки;
• Nazarovispongus Kozur, 1980 (рис. 3, в; 4, б–п) характеризуется образованием скелета от сильно выпуклой субтреугольной до почти субсферической формы с выпуклыми боковыми сторонами и всегда закрытой центральной частью внешней оболочки скелета;
• Kozurispongus gen. nov. (рис. 3, г; 5) выделяется формированием скелета уплощенной субтреугольной формы с прямыми или вогнутыми боковыми сторонами и, как правило, со слегка вогнутой и открытой центральной частью внешней оболочки (рис. 5, а, б, г, д; табл. III , см. вклейку), которая, однако, может быть частично или полностью закрыта очень тонким покровным слоем (рис. 5, в, е–з; табл. IV , см. вклейку);
Рис. 4.
Ruzhencevispongus uralicus Kozur, 1980 (а) и виды рода Nazarovispongus Kozur, 1980 (б–п): а–е – Россия, Южный Урал: а – нижняя пермь, верхнекунгурский подъярус, кошелевская свита, Южный Урал, р. Ай, с. Алегазово (Kozur, 1980), б, в – нижняя пермь, сакмарский ярус, сарабильская свита, р. Урал, разрез Верхнеозерный (Kozur, 1980), г – нижняя пермь, артинский ярус, р. Урал, разрез Донское (Назаров, 1984), д, е – верхний карбон, гжельский ярус, р. Урал, разрез Никольский (Исакова, Назаров, 1986); ж–п – США, Западный Техас, средняя пермь: ж – роудский ярус (Nazarov, Ormiston, 1985), з–п – кептенский ярус (Nazarov, Ormiston, 1985; Nestell, Nestell, 2010). [Авторские права на фотографии радиолярий (а–в) из: Kozur, 1980 принадлежат “©Universität Innsbruck, Institut für Geologie” и используются с любезного разрешения проф. Д. Сандерса. Фотографии радиолярий (г–з) приведены из фотоархива Б.Б. Назарова. Авторские права на фотографии радиолярий (и–п) из: Nestell, Nestell, 2010 принадлежат “©Micropaleontology” и используются с любезного разрешения д-ра М. Камински]. а – Ruzhencevispongus uralicus Kozur, 1980, экз. № Ko-1979/III-1, штрих = 57 мкм (Kozur, 1980, табл. 1 , фиг. 1); б–г – Nazarovispongus pavlovi Kozur, 1980, экз. № Ko-1979/III-3: б – штрих = 84 мкм (Kozur, 1980, pl. 1, fig. 6), в – фрагмент, штрих = 22 мкм (Kozur, 1980, pl. 2, fig. 1), г – экз. № ГИН 4673/121, штрих = 136 мкм; д – N. aequilateralis (Nazarov in Isakova et Nazarov, 1986), экз. ГИН, № 4488/162, штрих = 125 мкм (Исакова, Назаров, 1986, табл. 26 , фиг. 9 ); е – N. subrotundus (Nazarov in Isakova et Nazarov, 1986), экз. ГИН, № 4488/179, штрих = = 115 мкм; ж – N. delicatus (Nazarov et Ormiston, 1985), экз. № USNM 257613, штрих = 107 мкм (Nazarov, Ormiston, 1985, табл. 5 , фиг. 4); з–л – N. inflatus (Nazarov et Ormiston, 1985): з – экз. № USNM 257614, штрих = 100 мкм (Nazarov, Ormiston, 1985, табл. 5 , фиг. 6), и – экз. № USNM 538706, штрих = 109 мкм (Nestell, Nestell, 2010, табл. 17 , фиг. 9 ), к – экз. № USNM 538710, фрагмент, штрих = 300 мкм (Nestell, Nestell, 2010, табл. 17 , фиг. 13a ), л – экз. № USNM 538709, штрих = 100 мкм (Nestell, Nestell, 2010, табл. 17 , фиг. 12a ); м–п – N. globosus Nestell et Nestell, 2010: м – экз. № USNM 538711, штрих = 110 мкм (Nestell, Nestell, 2010, табл. 17 , фиг. 14 ), н – экз. № USNM 538713, штрих = 120 мкм (Nestell, Nestell, 2010, табл. 17 , фиг. 16 ), о – экз. № USNM 538712, штрих = 120 мкм (Nestell, Nestell, 2010, табл. 17 , фиг. 15 ), п – экз. № USNM 538714, фрагмент, штрих = 300 мкм (Nestell, Nestell, 2010, табл. 17 , фиг. 17a ).
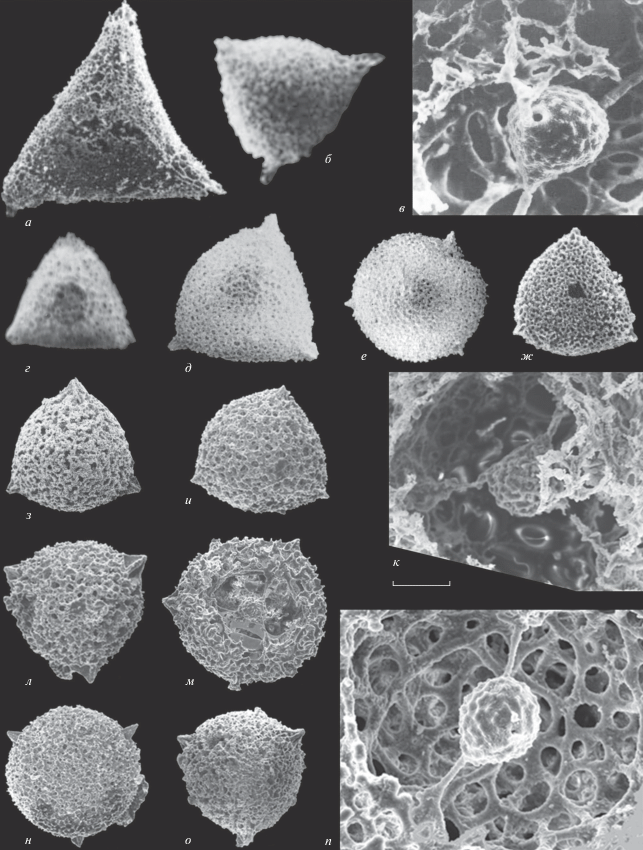
Рис. 5.
Виды рода Kozurispongus gen. nov.: а–ж – Россия, Южный Урал: а, б – нижняя пермь, сакмарский ярус, сарабильская свита, р. Урал, разрез Верхнеозерный (Kozur, 1980), в, е – верхний карбон, гжельский ярус, р. Урал, разрез Никольский (Исакова, Назаров, 1986), г, д – нижняя пермь, сакмарский ярус, малоикская свита, р. Урал, разрез Донское (Назаров, 1984; Nazarov, Ormiston, 1985), ж – нижняя пермь, артинский ярус, р. Урал, разрез Донское (Назаров, 1984); з – Казахстан, Западные Мугоджары, р. Актасты, разрез Актасты, нижняя пермь, артинский ярус (Назаров, 1984; Nazarov, Ormiston, 1993). [Авторские права на фотографии радиолярий (а, б) из работы Kozur, 1980 принадлежат “©Universität Innsbruck, Institut für Geologie” и используются с любезного разрешения проф. Д. Сандерса. Фотографии радиолярий (в–з) приведены из фотоархива Б.Б. Назарова]. а, б – K. permicus (Kozur, 1980): а – экз. № Ko-1979/III-5, штрих = 128 мкм (Kozur, 1980, табл. 1 , фиг. 5), б – экз. № Ko-1979/III-4, штрих = 150 мкм (Kozur, 1980, табл. 1 , фиг. 4); в – K. semilaminus (Nazarov in Isakova et Nazarov, 1986), экз. ГИН, № 4488/264, штрих = 134 мкм (Исакова, Назаров, 1986, табл. 21 , фиг. 8 ); г, д – K. circumfusus (Nazarov et Ormiston, 1985): г – экз. ГИН, № 4672/16, штрих = 158 мкм (Nazarov, Ormiston, 1985, табл. 5 , фиг. 7), д – экз. ГИН, № 4672/26, штрих = 156 мкм; е–з – K. aktastiensis (Nazarov et Ormiston, 1993): е – экз. ГИН, № 4488/285, штрих = 167 мкм (Исакова, Назаров, 1986, табл. 24 , фиг. 2), ж – экз. ГИН, № 4673/104, штрих = 156 мкм, з – экз. ГИН, № 4673/138, штрих = 150 мкм (Nazarov, Ormiston, 1993, табл. 6 , фиг. 1).

• Foremanhelena De Wever et Caridroit, 1984 (рис. 3, д) отличается развитием треугольного скелета со всегда открытой центральной частью внешней оболочки скелета, образованной грубой губчатой скелетной тканью.
Формирование игл радиолярий
Внешние иглы разнообразной формы очень характерны для радиолярий. Иглы закладываются на самом первом этапе становления скелетов у всех радиолярий. Процесс формирования игл подобен на начальном этапе заложения игл, но имеет существенные отличия на последующих этапах развития.
Исследования Sphaerellaria и Stauraxonaria палеозоя позволили построить две схемы последовательных этапов формирования игл (рис. 6).
Рис. 6.
Схемы формирования игл: а, б – Kozurispongus concavilatus sp. nov.: а – схема иглы, б – экз. ПИН, № 5481/16667, фрагмент, штрих = 40 мкм; Южный Урал, р. Сакмара, разрез Кондуровский; нижняя пермь, сакмарский ярус, карамурунская свита; в, г – Bientactinosphaera pinica Afanasieva, 2000: в – схема иглы, г – экз. ПИН, № 5311/08604, фрагмент, штрих = 50 мкм; Тимано-Печорский бассейн, скв. Шуда-Яг-1003, обр. 72 (инт. 71.9–72.4 м); верхний девон, среднефранский подъярус, доманиковая свита. Обозначения: 1, 2 – внутренний каркас: 1 – полая сфера, 2 – спикула; 3 – первичная внутренняя сфера, 4 – первичные шипы, 5 – начальный этап формирования игл, 6 – часть внешней сферы скелета, 7 – основная игла, 8 – структурный конус.

Сферические скелеты радиолярий. Строение внутреннего скелета Bientactinosphaera pinica Afanasieva, 2000 позволило предложить схему последовательных этапов формирования основных игл у сферических скелетов радиолярий (Afanasieva et al., 2005; Афанасьева, Амон, 2006; Афанасьева, 2007). В сферическом скелете сначала закладывается внутренний каркас в виде многолучевой спикулы (рис. 6, в, г). Затем образуется первичная внутренняя сфера. На поверхности первичной внутренней сферы и на продолжении лучей спикулы закладываются первичные шипы. Первичные шипы служат основой для начального этапа формирования основной иглы: она как бы “насаживается” на первичную иглу по принципу матрешки или cone-in-cone.
Далее наступает завершающий этап – формирование собственно основной иглы. Он несколько отличается отдельными деталями в разных классах Sphaerellaria и Spumellaria, но в целом прослеживается закономерное постепенное нарастание скелета иглы от основания к дистальной части.
Ставраксонные радиолярии. Анализ прижизненного дефекта в строении скелета Kozurispongus concavilatus sp. nov. (рис. 6, б; табл. III , фиг. 6) позволяет восстановить возможную последовательность образования игл на первичной сфере.
Процесс формирования игл начинается с образования внутреннего каркаса в форме неперфорированной полой сферы (рис. 6, а, б). На поверхности первичной внутренней сферы, так же как и у Sphaerellaria, закладываются первичные шипы. Они также служат основой для начального этапа формирования полых цилиндрических игл, которые как бы “насаживаются” на первичный шип по принципу матрешки или cone-in-cone.
Завершающий этап формирования цилиндрических игл Ruzhencevispongidae отличается от образования основных игл Sphaerellaria, поскольку связан со спецификой нарастания губчатой скелетной ткани внешней оболочки скелета.
Иглы расширяются по направлению к внешней оболочке, и на их окончании формируются высокие структурные конусы за счет обрастания игл губчатой скелетной тканью внешней оболочки скелета (рис. 6, а, б; табл. III , фиг. 1–9 ).
Рис. 7.
Измерение скелета: H – максимальная высота, W – ширина внутренней полости, R – валик, D – диаметр внутренней сферы, L – длина терминальных шипов, dp − диаметр пор внешней оболочки, dc − диаметр ячей покровной оболочки.
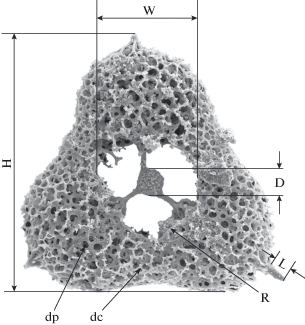
Кроме этого, среди исследованных радиолярий распространены формы с терминальными шипами, обрастающими скелетной тканью внешней оболочки скелета (табл. IV , фиг. 4, 5).
РОД NAZAROVISPONGUS
История изучения рода Nazarovispongus началась в 1980 г., когда Х. Коцур (Kozur, 1980) установил и описал новый род Nazarovispongus Kozur, 1980 с двумя видами N. pavlovi Kozur, 1980 (типовой вид) и N. permicus Kozur, 1980.
Род Nazarovispongus характеризуется наличием уплощенно-выпуклого треугольного губчатого скелета с выпуклыми сторонами, короткими терминальными трехлопастными шипами и первичной неперфорированной сферой. Пространство между внешней губчатой оболочкой и первичной сферой остается свободным.
Б.Б. Назаров и А.Р. Ормистон (Nazarov, Ormiston, 1983) без какого-либо пояснения отметили, что, вероятно, род Nazarovispongus, выделенный Коцуром (Kozur, 1980), принадлежит к семейству Tormentidae и может быть отнесен к категории родов-спутников. П. Думитрика также без объяснения причин указал Nazarovispongus Kozur, 1980 в качестве младшего синонима Ruzhencevispongus Kozur, 1980: “Ruzhencevispongus Kozur (?=Nazarovispongus Kozur)” (Думитрика, 1984, с. 100).
В 1985 г. Назаров и Ормистон (Nazarov, Ormiston, 1985, с. 11) вновь предложили рассматривать Nazarovispongus в качестве рода-спутника в семействе Tormentidae и подвергли род Nazarovispongus необъективной критической оценке:
• неясна структура внутренней сферы N. pavlovi и соединение ее лучей с наружными шипами и губчатым слоем;
• не указано строение центральной сферы N. permicus;
• неясно, является ли скелетная ткань внутренней сферы N. permicus губчатой, мелкопористой или пластинчатой, или внутренняя сфера вообще отсутствует.
Между тем в работе Коцура (Kozur, 1980) четко сказано:
– строение скелетной ткани внутренней сферы N. pavlovi и N. permicus соответствует ее описанию в диагнозе рода: оболочка внутренней сферы неперфорированная, с тремя цилиндрическими иглами, в основании которых находятся три большие поры11 (рис. 4, в) (Kozur, 1980, с. 237, табл. 2 , фиг. 1, 2);
– внутренняя сфера у N. permicus присутствует, а ее строение Коцуром изображено (рис. 5, а) (Kozur, 1980, с. 237, табл. 1 , фиг. 5).
В 1984–1993 гг. Назаров и Ормистон (Назаров, 1984, 1988; Назаров, Ормистон, 1984, 1990; Nazarov, Ormiston, 1985, 1993) больше не упоминают Nazarovispongus.
В 1989 г. Х. Коцур и Х. Мостлер (Kozur, Mostler, 1989) не согласились с выводами Назарова, Ормистона (Nazarov, Ormiston, 1983, 1985) и Думитрики (1984). Они справедливо заметили22, что Nazarovispongus имеет многослойный губчатый внешний скелет с тонкопористым, не всегда сохранившимся внешним покровным слоем и более крупнопористой внутренней скелетной тканью, что указано в диагнозе семейства и также хорошо видно на снимках SEM. Более того, Коцур и Мостлер (Kozur, Mostler, 1989) исправили диагноз рода Tormentum, оставив в его составе только виды, сходные по морфологии с типовым видом T. protei Nazarov et Ormiston, 1983, и показали, что виды, описанные как Tormentum Назаровым и Ормистоном (Nazarov, Ormiston, 1985), принадлежат к двум различным родам – Tormentum s.s. и Nazarovispongus. Виды с “вздутыми” скелетами, такими как у Nazarovispongus pavlovi и Tormentum inflatum, были включены в состав рода Nazarovispongus.
Кроме этого Коцур и Мостлер (Kozur, Mostler, 1989) обратили внимание на то, что голотип Tormentum delicatum Nazarov et Ormiston, 1985 по морфологическим признакам внешнего скелета может быть отнесен к роду Nazarovispongus. Однако внутренняя сфера T. delicatum, согласно данным Назарова и Ормистона (Nazarov, Ormiston, 1985), отличается формированием пористой оболочки, тогда как у Nazarovispongus первичная внутренняя сфера непористая.
Следует обратить внимание на то, что при первичном описании Tormentum delicatum Назаров и Ормистон указали на изменение конфигурации скелета от округло-треугольной до формы клеверного листа33 и привели изображения двух совершенно разных морфотипов (Nazarov, Ormiston, 1985, табл. 5 , фиг. 4 и 5). При этом у “округло-треугольного” голотипа внутренняя сфера не видна (рис. 4, ж), а у “клеверного” морфотипа внутренняя сфера присутствует, но структура ее пористой поверхности может быть установлена с большим трудом в силу плохого качества изображения. Кроме этого, следует отметить, что у представителей семейства Ruzhencevispongidae поверхность внутренней сферы имеет бугристую структуру (рис. 4, в, к, п). При плохом качестве изображения бугристость поверхности можно принять за пористость.
К сожалению, Назаров и Ормистон (Nazarov, Ormiston, 1983, 1985; Исакова, Назаров, 1986; Назаров, 1988) достаточно вольно обращались с определением характера пористости или не пористости внутренней сферы:
• в диагнозах семейства Tormentidae и рода Tormentum показано присутствие полой внутренней сферы, но без указания наличия или отсутствия пор;
• только в 1988 г. Назаров отметил наличие неперфорированной внутренней сферы при описании семейства Tormentidae;
• в диагнозах видов из рода Tormentum также не акцентируется внимание на наличии или отсутствии пор на внутренней сфере, отмечается только присутствие полой внутренней сферы;
• два вида из рода Tormentum являются исключением:
– типовой вид T. protei, в диагнозе которого указано присутствие неперфорированной внутренней сферы (Nazarov, Ormiston, 1983, 1985; Исакова, Назаров, 1986);
– T. delicatum, характеризующийся наличием пористой внутренней оболочки44 (Nazarov, Ormiston, 1985), которая с большим трудом наблюдается у морфотипа с клеверной формой скелета, но отсутствует у голотипа округло-треугольной формы.
Таким образом, можно допустить принадлежность “округло-треугольного” голотипа T. delicatum к роду Nazarovispongus: N. delicatus:
(1) исходя из не определенной однозначно структуры внутренней сферы у представителей семейства Tormentidae и рода Tormentum в работах Назарова и Ормистона (Nazarov, Ormiston, 1983, 1985; Исакова, Назаров, 1986);
(2) ориентируясь на указание наличия неперфорированной внутренней сферы при описании семейства Tormentidae (Назаров, 1988) и типового вида T. protei (Nazarov, Ormiston, 1983, 1985; Исакова, Назаров, 1986);
(3) учитывая отсутствие изображения пористой первичной сферы в округло-треугольном скелете голотипа T. delicatum в какой-либо публикации Назарова и Ормистона.
С другой стороны “клеверный” морфотип T. delicatum должен быть рассмотрен в составе иного рода. По мнению Коцура и Мостлера (Kozur, Mostler, 1989) этот вид, возможно, принадлежит к Scharfenbergia Won, 1983 (Won, 1983), скелеты которого отличаются наличием внутренней пористой сферы. Вместе с тем, название Scharfenbergia преоккупировано и оказалось младшим омонимом паукообразных Scharfenbergia Oudemans, 1936 (Oudemans, 1936). В связи с этим для радиолярий было предложено (Özdikmen, 2009) новое замещающее название Wonia Özdikmen, 2009. К сожалению, истинная родовая приуроченность “клеверного” морфотипа Tormentum delicatum до сих пор остается неопределенной.
Г. и М. Нестелл (Nestell, Nestell, 2010) на основании исследования радиолярий из среднепермских, кептенских отложений Западного Техаса рассматривают род Nazarovispongus в качестве самостоятельного валидного таксона, включающего три вида: N. pavlovi Kozur, 1980 (типовой вид), N. inflatus (Nazarov et Ormiston, 1985) и N. globosus Nestell et Nestell, 2010.
П. Нобл с коллегами (Noble et al., 2017, с. 470) включили Nazarovispongus в состав рода Ruzhencevispongus в качестве младшего синонима на основании необоснованного заключения Думитрики (1984, с. 100). Видовой состав рода объединяет три вида (Aitchison et al., 2017, с. 594): Nazarovispongus pavlovi, N. permicus и N. globosus. К сожалению, эти авторы (Aitchison et al., 2017; Noble et al., 2017) не обратили должного внимания на аргументированное восстановление самостоятельности и валидного статуса рода Nazarovispongus, а также на видовой состав рода, предложенные Нестелл и Нестелл (Nestell, Nestell, 2010).
В настоящей публикации автор позволяет себе усомниться в правильности решения об упразднении Nazarovispongus в качестве самостоятельной систематической единицы (Думитрика, 1984; Noble et al., 2017) и согласен с мнением Нестелл и Нестелл (Nestell, Nestell, 2010) о валидности рода Nazarovispongus.
Новый уточненный диагноз вновь восстановленного и валидного рода Nazarovispongus Kozur, 1980 sensu Kozur et Mostler, 1989, restituo et emend. nov. принимается в редакции Коцура и Мостлера (Kozur, Mostler, 1989) с дополнениями на основе собственного анализа морфологии скелета и с учетом диагноза, сделанного ранее Нестелл и Нестелл (Nestell, Nestell, 2010, с. 56). Видовой состав рода расширен до шести видов: N. pavlovi Kozur, 1980 (типовой вид), N. aequilateralis (Nazarov in Isakova et Nazarov, 1986, comb. nov.), N. delicatus (Nazarov et Ormiston, 1985, part., comb. nov.), N. globosus Nestell et Nestell 2010, N. inflatus (Nazarov et Ormiston, 1985, comb. nov.) и N. subrotundus (Nazarov in Isakova et Nazarov, 1986, comb. nov.) (рис. 2; 4, б–п).
РОД KOZURISPONGUS
При описании рода Nazarovispongus Коцур (Kozur, 1980) указал в его составе два вида – N. pavlovi (типовой вид) и N. permicus. Однако Коцур и Мостлер (Kozur, Mostler, 1989) предложили перевести N. permicus в состав рода Foremanhelena De Wever et Caridroit, 1984 и рассматривать его в качестве переходной формой между двумя родами: Nazarovispongus и Foremanhelena. Вместе с тем, морфотип Nazarovispongus permicus отличается от морфотипов рода Foremanhelena (рис. 3, д) формированием более тонкой губчатой скелетной ткани внешней оболочки скелета и менее открытой центральной частью скелета, которая, в случае очень хорошо сохранившихся форм, может быть частично или полностью закрыта очень тонким покровным слоем (рис. 5, е–з; табл. IV ).
Нестелл и Нестелл (Nestell, Nestell, 2010) не упоминают N. permicus в составе рода Nazarovispongus, но и не относят его к какому-либо другому роду. С другой стороны, Дж. Ачисон и др. (Aitchison et al., 2017, с. 594) рассматривают N. permicus в составе рода Nazarovispongus.
Новое изучение треугольных морфотипов из авторской коллекции раннепермских радиолярий, происходящих из ассельских и сакмарских отложений опорного разреза Кондуровский на Южном Урале, а также привлечение литературных данных по радиоляриям: (1) из артинских отложений разреза Актасты (рис. 1, V) (Назаров, 1988; Nazarov, Ormiston, 1993), (2) из сакмарских отложений разрезов Верхнеозерный (рис. 1, II) (Kozur, 1980) и Донское (рис. 1, III) (Nazarov, Ormiston, 1985), (3) из гжельских отложений разреза Никольский (рис. 1, IV) (Исакова, Назаров, 1986), позволяет установить самостоятельный новый род Kozurispongus gen. nov. Видовой состав нового рода представлен шестью видами: K. aktastiensis (Nazarov et Ormiston, 1993, comb. nov.), K. circumfusus (Nazarov et Ormiston, 1985, comb. nov.), K. permicus (Kozur, 1980, comb. nov.), K. semilaminus (Nazarov in Isakova et Nazarov, 1986, part., comb. nov.) и два новых вида, K. concavilatus sp. nov. и K. rotundatus sp. nov. (рис. 2; 5; табл. III ; IV).
ОПИСАНИЕ ТАКСОНОВ
ТИП RADIOLARIA
НАДКЛАСС POLYCYSTINA
КЛАСС STAURAXONARIA
ОТРЯД PYRAMIDATA
СЕМЕЙСТВО RUZHENCEVISPONGIDAE KOZUR, 1980 SENSU KOZUR ET MOSTLER, 1989
Род Nazarovispongus Kozur, 1980 sensu Kozur et Mostler, 1989, restituo et emend. nov.
Nazarovispongus: Kozur, 1980, с. 237; Kozur, Mostler, 1989, с. 205; Nestell, Nestell, 2010, с. 56; Noble et al., 2017, с. 470.
Типовой вид – Nazarovispongus pavlovi Kozur, 1980; Россия, Южный Урал; нижняя пермь, сакмарский ярус, сарабильская свита (Kozur, 1980, с. 238, табл. 1 , фиг. 6; табл. 2 , фиг. 1, 2).
Диагноз55. Скелет от субтреугольной до субсферической формы с выпуклыми сторонами (верхней, нижней и боковыми) и с острыми или закругленными углами (рис. 3, в; 4, б–п). Внешняя оболочка скелета губчатая, в бóльшей или меньшей степени заполняет внутреннюю часть скелета, при этом внутреннее пространство скелета вокруг внутренней сферы всегда свободно. Внутренний каркас представлен непористой полой сферой, поверхность которой имеет бугристую структуру. От поверхности внутренней сферы в плоскости гипотетического треугольника под углом 120° друг к другу отходят три полых цилиндрических иглы, при этом центр гипотетического пересечения игл смещен относительно центра внутренней сферы (рис. 4, к). На базе игл формируются высокие структурные конусы за счет обрастания игл губчатой скелетной тканью внешней оболочки скелета. В основании одной из игл образуется базальная пора (рис. 4, в). На внешней поверхности скелета, в вершинах гипотетического треугольника расположены три коротких трехгранных или конических шипа.
Видовой состав. Шесть видов из верхнего карбона – средней перми (рис. 2): типовой вид N. pavlovi Kozur, 1980, N. aequilateralis (Nazarov in Isakova et Nazarov, 1986, comb. nov.), N. delicatus (Nazarov et Ormiston, 1985, part., comb. nov.), N. globosus Nestell et Nestell 2010, N. inflatus (Nazarov et Ormiston, 1985, comb. nov.), N. subrotundus (Nazarov in Isakova et Nazarov, 1986, comb. nov.).
Сравнение. От Ruzhencevispongus Kozur, 1980 sensu Kozur et Mostler, 1989 (рис. 3, а; 4, а) [non Ruzhencevispongus Kozur, 1980 sensu Nazarov et Ormiston, 1983 (рис. 3, б)], отличающегося формированием треугольного скелета с прямыми или слегка вогнутыми боковыми и слегка выпуклыми верхней и нижней сторонами, Nazarovispongus отличается формированием вздутого скелета от субтреугольной до субсферической формы.
Род Kozurispongus Afanasieva, gen. nov.
Название рода в честь палеонтолога и стратиграфа Х. Коцура; м. р.
Типовой вид – Kozurispongus concavilatus sp. nov.
Диагноз. Скелет уплощенной субтреугольной формы с прямыми или вогнутыми боковыми сторонами, со слегка вогнутой центральной частью и с острыми, закругленными или плоскими углами гипотетического равностороннего треугольника (рис. 3, г; 5). Внешняя оболочка скелета губчатая. Центральная часть внешней оболочки, как правило, открыта (рис. 5, а, б, г, д; табл. III ), но может быть частично или полностью закрыта очень тонким покровным слоем (рис. 5, в, е–з; табл. IV ). Внутренний каркас представлен непористой полой сферой, поверхность которой имеет бугристую структуру. Внутреннее пространство скелета вокруг внутренней сферы всегда свободно. От поверхности внутренней сферы в плоскости гипотетического треугольника под углом 120° друг к другу отходят три полых цилиндрических иглы, при этом центр гипотетического пересечения игл смещен относительно центра внутренней сферы (рис. 5, а, г–е; табл. III ; табл. IV , фиг. 3а). Иглы расширяются по направлению к внешней оболочке скелета, и на их окончании формируются высокие структурные конусы за счет обрастания игл губчатой скелетной тканью внешней оболочки (рис. 6, а, б). В основании одной из игл образуется базальная пора (табл. III , фиг. 9 ). На внешней поверхности скелета, в вершинах гипотетического треугольника расположены три коротких конических шипа.
Видовой состав. Шесть видов из верхнего карбона – нижней перми (рис. 2): типовой вид K. concavilatus sp. nov., K. aktastiensis (Nazarov et Ormiston, 1993, comb. nov.), K. circumfusus (Nazarov et Ormiston, 1985, comb. nov.), K. permicus (Kozur, 1980, comb. nov.), K. rotundatus sp. nov., K. semilaminus (Nazarov in Isakova et Nazarov, 1986, part., comb. nov.).
Сравнение. От Ruzhencevispongus Kozur, 1980 sensu Kozur et Mostler, 1989 (рис. 3, а; 4, а) [non Ruzhencevispongus Kozur, 1980 sensu Nazarov et Ormiston, 1983 (рис. 3, б)], отличающегося формированием треугольного скелета со слегка выпуклыми верхней и нижней сторонами, а также закрытой центральной частью внешней оболочки, новый род отличается формированием уплощенного субтреугольного скелета со слегка вогнутой и, как правило, открытой центральной частью внешней оболочки. От Nazarovispongus Kozur, 1980 (рис. 3, в; 4, б–п), характеризующегося образованием скелета от сильно выпуклой субтреугольной до почти субсферической формы с выпуклыми боковыми сторонами и всегда закрытой центральной частью внешней оболочки скелета, новый род отличается формированием уплощенного субтреугольного скелета со слегка вогнутой и, как правило, открытой центральной частью внешней оболочки. От Foremanhelena De Wever et Caridroit, 1984 (рис. 3, д), отличающегося развитием грубой губчатой скелетной ткани и всегда открытой центральной частью внешней оболочки треугольного скелета, новый род отличается наличием тонкой губчатой скелетной ткани и менее открытой центральной частью скелета, которая, может быть частично или полностью закрыта очень тонким покровным слоем.
Kozurispongus concavilatus Afanasieva, sp. nov.
Табл. III , фиг. 1–8
Название вида от concavus лат. – вогнутый, и latus лат. – сторона, бок.
Голотип − ПИН, № 5481/16679; Южный Урал, р. Сакмара, разрез Кондуровский, слой 17; нижняя пермь, сакмарский ярус, тастубский горизонт, карамурунская свита, экозона радиолярий Arcoclathrata alekseevi–Apophysiacus sakmaraensis, конодонтовая зона Sweetognathus merrilli.
Описание (рис. 6, б). Скелет большой (H = = 327–369 мкм), уплощенный, с округлыми, вогнутыми боковыми сторонами и с закругленными углами гипотетического равностороннего треугольника. Внешняя оболочка скелета губчатая и состоит из переплетения относительно толстых прутьев (wp = 4 мкм), образующих крупные полигональные поры (dp = 8–12 мкм) (рис. 7, dp). Центральная часть внешней оболочки открыта, имеет лепестковидную или округлую форму и окаймлена валиком (рис. 7, R; табл. III , фиг. 6). Она составляет около 1/3 от высоты скелета (W = = 107–122 мкм) (рис. 7, W). Центральная часть скелета может быть частично или полностью закрыта очень тонким покровным решетчатым слоем. Покровный слой развивается непосредственно на каркасе внешней оболочки и потенциально закрывает всю поверхность скелета (табл. III , фиг. 4). Он образован переплетением тонких прутьев (wc = 2–3 мкм), образующих крупные полигональные ячейки (dc = 5–9 мкм) (рис. 7, dc). В некоторых случаях прутья покровного слоя могут частично разрастаться, формируя тонкий слой почти пластинчатой скелетной ткани с очень маленькими порами (1–1.5 мкм) (табл. III , фиг. 3, 7а, 8б). Внутренний каркас представлен очень маленькой (D = 32–35 мкм) непористой полой сферой, поверхность которой имеет бугристую структуру (рис. 7, D; табл. III ). Диаметр сферы составляет 9.4–10.1% от максимальной высоты скелета. На внешней поверхности скелета, в вершинах гипотетического треугольника расположены три коротких конических шипа (L = 20–29 мкм) (рис. 7, L).
| Экз. № | H | W | D | L | wL | dp | wp | dc | wc |
|---|---|---|---|---|---|---|---|---|---|
| 5481/16679 (голотип) | 327 | 108 | 33 | 29 | 13 | 9 | 4 | 9 | 2 |
| 5481/16667 | 369 | 122 | 35 | 27 | 16 | 12 | 4 | 9 | 3 |
| 5481/16668 | 361 | 107 | 34 | 20 | 14 | 8 | 4 | 5 | 2 |
| 5481/16606 | 334 | 110 | 32 | 22 | 13 | 10 | 4 | 5 | 2 |
Сравнение. От K. permicus Kozur, 1980 (рис. 5, а, б), отличающегося слегка припухлой формой треугольного скелета и плотной губчатой тканью внешней оболочки, новый вид отличается более уплощенной формой и слегка вогнутыми сторонами скелета, а также развитием более рыхлой губчатой ткани внешней оболочки скелета и развитием покровного слоя. От K. semilaminus (Nazarov in Isakova et Nazarov, 1986, part., comb. nov.) (рис. 5, в), отличающегося образованием равностороннего или равнобедренного треугольного скелета с пластинчатой скелетной тканью в центре внешней оболочки и губчатой на сетчатом основании по периферии, новый вид отличается развитием округлых вершин треугольного скелета и в той или иной степени вогнутыми боковыми сторонами скелета. От K. circumfusus (Nazarov et Ormiston, 1985, comb. nov.) (рис. 5, г, д), характеризующегося уплощенной формой скелета с прямыми сторонами, рыхлой губчатой тканью внешней оболочки и правильной треугольной формой открытой центральной части внешней оболочки, новый вид отличается слегка вогнутыми боковыми сторонами скелета, лепестковидной формой открытой центральной части внешней оболочки, а также развитием тонкого покровного слоя. От K. aktastiensis (Nazarov et Ormiston, 1993, comb. nov.) (рис. 5, е–з), выделяющегося развитием прямых, как бы усеченных вершин треугольного скелета и, как правило, закрытой покровным слоем центральной частью внешней оболочки, новый вид отличается формированием округлых вершин треугольного скелета, более вогнутыми боковыми сторонами и, как правило, открытой центральной частью внешней оболочки. От K. rotundatus sp. nov., для которого характерно образование округло-треугольного скелета, иногда практически округлого, поверхность которого полностью закрыта тонкой покровной тканью, новый вид отличается развитием скелета с открытой центральной частью и с вогнутыми боковыми сторонами.
Материал. 19 экз. из типового местонахождения: десять экз. из ассельских и девять экз. из сакмарских отложений.
Kozurispongus rotundatus Afanasieva, sp. nov.
Табл. IV , фиг. 1–9
Название вида rotundatus лат. – округлый.
Голотип − ПИН, № 5481/16246; Южный Урал, р. Сакмара, разрез Кондуровский, слой 17; нижняя пермь, сакмарский ярус, тастубский горизонт, карамурунская свита, экозона радиолярий Arcoclathrata alekseevi–Apophysiacus sakmaraensis, конодонтовая зона Sweetognathus merrilli.
Описание. Скелет большой (H = 316–353 мкм), уплощенный, как правило, округло-треугольный, но иногда формируется почти круглый скелет (табл. IV , фиг. 2). Внешняя оболочка скелета губчатая и состоит из переплетения относительно толстых прутьев (wp = 4–6 мкм), образующих крупные полигональные поры (dp = 7–8 мкм) (рис. 7, dp). Покровный слой развивается непосредственно на каркасе внешней оболочки и закрывает всю поверхность скелета. Он образован переплетением тонких прутьев (wc = 2 мкм), образующих крупные полигональные ячейки (dc = 4–5 мкм) (рис. 7, dc). В центральной части покровного слоя прутья разрастаются, формируя тонкий слой почти пластинчатой скелетной ткани с очень маленькими порами (1–1.5 мкм). Внутренний каркас представлен очень маленькой (D = 31–35 мкм) непористой полой сферой, поверхность которой имеет бугристую структуру (рис. 7, D; табл. IV , фиг. 3а). Диаметр сферы составляет 9.6–10.2% от максимальной высоты скелета. На внешней поверхности скелета, в вершинах гипотетического треугольника расположены три коротких конических шипа (L = 18–24 мкм) (рис. 7, L). Нижняя половина шипа обрастает скелетной тканью внешней оболочки скелета, что создает иллюзию формирования укрепляющих распорок (табл. IV , фиг. 5а).
Размеры в мкм (рис. 7):
| Экз. № | H | D | L | wL | dp | wp | dc | wc |
|---|---|---|---|---|---|---|---|---|
| 5481/16246 (голотип) | 353 | – | 24 | 11 | 7 | 4 | 4 | 2 |
| 5481/16238 | 316 | 32 | 18 | 15 | 7 | 4 | 4 | 2 |
| 5481/16240 | 342 | 35 | 24 | 18 | 8 | 6 | 5 | 2 |
| 5481/16247 | 322 | 31 | 21 | 16 | 8 | 6 | 4 | 2 |
Сравнение. От всех других видов рода Kozurispongus (рис. 5; табл. III ) новый вид отличается образованием округло-треугольного скелета, поверхность которого полностью закрыта тонкой покровной тканью, вплоть до образования очень тонкопористой, практически пластинчатой скелетной ткани в центральной части скелета.
Материал. 11 экз.: 10 экз. из типового местонахождения (один экз. из ассельских и девять экз. из сакмарских отложений); один экз. из фотоархива Б.Б. Назарова: артинские отложения разреза Актасты на р. Актасты, Западные Мугоджары, Казахстан.
ЗАКЛЮЧЕНИЕ
Анализ морфологических особенностей треугольных скелетов радиолярий позднего карбона и ранней перми Южного Урала России и Западных Мугоджар Казахстана, а также средней перми Западного Техаса США позволил: (1) установить четыре морфотипа, характеризующих четыре рода: Ruzhencevispongus, Nazarovispongus, Kozurispongus, Foremanhelena; (2) предложить новую схему последовательных этапов формирования игл скелета; (3) восстановить валидность и дополнить диагноз рода Nazarovispongus Kozur, 1980 sensu Kozur et Mostler, 1989, restituo et emend. nov.; (4) расширить видовой состав рода Nazarovispongus до шести видов (рис. 2): N. pavlovi Kozur, 1980 (типовой вид), N. aequilateralis (Nazarov in Isakova et Nazarov, 1986, comb. nov.), N. delicatus (Nazarov et Ormiston, 1985, part., comb. nov.), N. globosus Nestell et Nestell 2010, N. inflatus (Nazarov et Ormiston, 1985, comb. nov.) и N. subrotundus (Nazarov in Isakova et Nazarov, 1986, comb. nov.); (5) установить новый род Kozurispongus gen. nov., объединяющий шесть видов (рис. 2): K. aktastiensis (Nazarov et Ormiston, 1993, comb. nov.), K. circumfusus (Nazarov et Ormiston, 1985, comb. nov.), K. permicus (Kozur, 1980, comb. nov.), K. semilaminus (Nazarov in Isakova et Nazarov, 1986, part., comb. nov.) и два новых вида K. concavilatus sp. nov. и K. rotundatus sp. nov.; (6) пересмотреть систематическое положение Nazarovispongus permicus, который рассматривается в составе нового рода Kozurispongus permicus (Kozur, 1980, nov. comb.).
* * *
Автор выражают свою признательность А.С. Алексееву, В.С. Вишневской и Г.П. Нестелл за ценные советы и конструктивные замечания; А.Ф. Банникову, Я.М. Кузьминой и М.А. Кнорре за помощь при подготовке рукописи к печати.
Авторские права на фотографии радиолярий из работ Kozur, 1980 и Nestell, Nestell, 2010 принадлежат “©Universität Innsbruck, Institut für Geologie” и “©Micropaleontology” и используются с любезного разрешения проф. Д. Сандерса (Diethard Sanders) и д-ра М. Камински (Mike Kaminski).
Систематика радиолярий приведена по работам автора (Афанасьева, 2000; Afanasieva et al., 2005; Афанасьева, Амон, 2006). Исследование морфологии радиолярий проводилось на сканирующем электронном микроскопе CamScan на базе Палеонтологического ин-та им. А.А. Борисяка (ПИН) РАН. Коллекция радиолярий нижней перми № 5481 Южного Урала и коллекция фотографий радиолярий из архива Б.Б. Назарова хранятся в лаборатории протистологии ПИН РАН.
Работа выполнена в рамках бюджетной программы (государственного задания) ПИН РАН и при поддержке программы Президиума РАН “Эволюция органического мира. Роль и влияние планетарных процессов”.
Список литературы
Афанасьева М.С. Атлас радиолярий палеозоя Русской платформы. М.: Научн. мир, 2000. 480 с.
Афанасьева М.С. Скелет радиолярий: морфология игл, внутреннего каркаса и первичной внутренней сферы // Палеонтол. журн. 2007. № 1. С. 3–14.
Афанасьева М.С., Амон Э.О. Радиолярии. М.: ПИН РАН, 2006. 320 с.
Афанасьева М.С., Амон Э.О., Черных В.В. Радиолярии пограничных отложений ассельского и сакмарского ярусов (нижняя пермь) разреза Кондуровский (западный склон Южного Урала // Бюлл. Моск. об-ва испыт. прир. Отд. геол. 2014. Т. 89. Вып. 4. С. 64–82.
Афанасьева М.С., Амон Э.О., Черных В.В. Артинские (нижняя пермь) радиолярии из опорного разреза Актасты, Северные Мугоджары // Бюлл. Моск. об-ва испыт. прир. Отд. геол. 2015. Т. 90. Вып. 5. С. 9–27.
Думитрика П. Систематика радиолярий Sphaerellaria // Морфология, экология и эволюция радиолярий: Матер. IV симп. европейских радиоляристов EURORAD IV / Ред. Петрушевская М.Г., Степаньянц С.Д. Л.: Наука, 1984. С. 91–102.
Исакова Т.Н., Назаров Б.Б. Стратиграфия и микрофауна позднего карбона – ранней перми Южного Урала. М.: Наука, 1986. 184 с. (Тр. ГИН АН СССР. Вып. 402).
Международный кодекс зоологической номенклатуры. Изд. четвертое. М.: Т-во научн. изданий КМК, 2004. 223 с.
Назаров Б.Б. Радиолярии палеозоя. Дис. … докт. геол.-мин. наук. М.: ГИН АН СССР, 1984. 642 с.
Назаров Б.Б. Радиолярии палеозоя. Практическое руководство по микрофауне СССР. Справочник для палеонтологов и геологов. Т. 2. Л.: Недра, 1988. 231 с.
Назаров Б.Б., Ормистон А.Р. Возможная система радиолярий палеозоя // Морфология, экология и эволюция радиолярий: Матер. IV симп. европейских радиоляристов EURORAD IV / Ред. Петрушевская М.Г., Степаньянц С.Д. Л.: Наука, 1984. С. 64–87.
Назаров Б.Б., Ормистон А.Р. Биостратиграфический потенциал радиолярий палеозоя // Радиолярии в биостратиграфии. Свердловск: УрО АН СССР, 1990. С. 3–25.
Aitchison J.C., Suzuki N., O’Dogherty L. Inventory of Paleozoic radiolarian species (1880–2016) // Catalogue of Paleozoic radiolarian genera / Eds. Danelian T., Caridroit M., Noble P., Aitchison J. C. (Geodiversitas. 2017. V. 39. № 3. P. 533–637).
Afanasieva M.S., Amon E.O., Agarkov Yu.V., Boltovskoy D.S. Radiolarians in the geological record // Paleontol. J. 2005. V. 39. Suppl. 3. P. 135–392.
Caridroit M., De Wever P. Some Late Permian radiolarians from Pelitic Rocks of the Tatsuno Formation (Hyogo Prefecture), Southwest Japan // Marine Micropaleontol. 1986. V. 11. P. 55–90.
Kozur H. Ruzhencevispongidae, eine neue Spumellaria Familie aus dem oberen Kungurian (Leonardian) und Sakmarian des Vorurals // Geol.-Paläontol. Mitt. Innsbruck. 1980. Bd 10. H. 6. S. 235–242.
Kozur H., Mostler H. Radiolarien und Schwammskleren aus dem Unterperm des Vorurals // Geol.-Paläontol Mitt. Innsbruck. 1989. Sonderband 2. S. 147–275.
Nazarov B.B., Ormiston A.R. A new superfamily of stauraxon polycystine Radiolaria from the Late Paleozoic of the Soviet Union and North America // Senckenb. Lethaea. 1983. Bd 64. № 2/4. P. 363–379.
Nazarov B.B., Ormiston A.R. Radiolaria from Late Paleozoic of the Southern Urals, USSR, and West Texas, USA // Micropaleontology. 1985. V. 31. № 1. P. 1–54.
Nazarov B.B., Ormiston A.R. New biostratigraphically important Paleozoic Radiolaria of Eurasia and North America // Radiolaria of giant and subgiant fields in Asia. Micropaleontology. 1993. Spec. Publ. № 6. P. 22–60.
Nestell G.P., Nestell M.K. Late Capitanian (latest Guadalupian, Middle Permian) radiolarians from the Apache Mountains, West Texas // Micropaleontology. 2010. V. 56. № 1–2. P. 7–68.
Noble P., Aitchison J.C., Danelian T. et al. Taxonomy of Paleozoic radiolarian genera // Catalogue of Paleozoic radiolarian genera / Eds. Danelian T., Caridroit M., Noble P., Aitchison J. C. (Geodiversitas. 2017. V. 39. № 3. P. 419–502).
Oudemans A.C. Neues über Anystidae (Acari) // Archiv für Naturges. N.S. 1936. Bd 5. S. 364–446.
Özdikmen, H. Substitute names for some unicellular animal taxa (Protozoa) // Munis Entomol. and Zool. 2009. V. 4. № 1. P. 233–256.
Won M.-Z. Radiolarien aus dem Unter-Karbon des Rheinischen Schiefergebirges (Deutschland) // Palaeontogr. Abt. A. 1983. Bd 182. № 4–6. S. 116–175.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал