

Палеонтологический журнал, 2021, № 3, стр. 24-34
Пермские двустворчатые моллюски кряжа Прончищева (Север Сибири): новые данные по таксономическому составу, биостратиграфии и биогеографическим связям
А. С. Бяков a, b, *, М. К. Данукалова c, А. Б. Кузьмичев c, Е. С. Соболев d
a Северо-Восточный комплексный научно-исследовавтельский институт им. Н.А. Шило ДВО РАН
Магадан, Россия
b Казанский (Приволжский) федеральный университет
Казань, Россия
c Геологический институт РАН
Москва, Россия
d Институт нефтегазовой геологии и геофизики СО РАН им. А.А. Трофимука
Новосибирск, Россия
* E-mail: abiakov@mail.ru
Поступила в редакцию 13.07.2020
После доработки 17.07.2020
Принята к публикации 04.08.2020
Аннотация
Впервые систематически изучены пермские морские двустворчатые моллюски кряжа Прончищева (север Сибири). Фаунистически доказано присутствие среднего и верхнего отделов перми, где установлены пять биостратиграфических уровней. В средней перми это бивальвиевые зоны Kolymia inoceramiformis – K. plicata (нерасчлененные) и Kolymia multiformis, а в верхней – зоны Maitaia bella (слои с Phestia ex gr. ovata), Maitaia belliformis и Intomodesma costatum. Комплексы двустворок средней и большей нижней части верхней перми (до зоны Intomodesma costatum) по своему таксономическому составу близки к комплексам Западного и Северного Верхоянья. Ископаемые сообщества верхов перми очень своеобразны и представлены почти исключительно монотаксонными скоплениями крупных пектинид Streblopteria rotunda (Lutkevich et Lobanova) и довольно редких представителей рода Intomodesma. Почти на протяжении всей перми морской бассейн, существовавший на территории кряжа Прончищева, имел устойчивые связи с Верхоянским. Этот бассейн был весьма мелководным и, возможно, испытывал некоторое опреснение. В конце перми палеогеографическая ситуация изменилась, и резко усилились связи морского бассейна кряжа Прончищева с акваторией Восточного Таймыра. Приведены изображения наиболее характерных видов двустворчатых моллюсков, описан новый вид Kolymia rutskovi Biakov, sp. nov.
ВВЕДЕНИЕ
В последние годы в связи с развертыванием работ по изучению арктических регионов России активизировалось стратиграфическое изучение этих обширных слабо исследованных территорий, весьма перспективных на горючие полезные ископаемые. Не стал исключением север Сибирской платформы, в частности, кряж Прончищева. Он представляет собой узкую протяженную квестообразную возвышенность высотой 150–250 м, вытянутую в запад-северо-западном направлении более чем на 150 км между Анабарской губой и Оленекским заливом (рис. 1). Бóльшая часть кряжа сложена породами мезозоя, отложения перми фрагментарно обнажены лишь в центральном сегменте возвышенности – в бассейнах рек Синнигес, Песчаной, Лыгый-Юряге и Урасалах. Пермские породы слагают переходную часть от северного подножья первой гряды кряжа к приморской низменности. До последнего времени пермские отложения кряжа были очень слабо изучены, а их возраст ненадежно обоснован из-за плохой обнаженности и крайне редких находок ископаемой фауны.
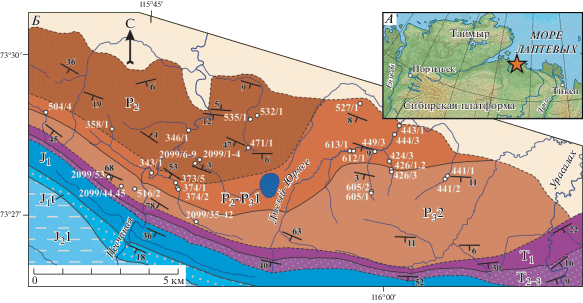
Рис. 1.
Расположение изученных местонахождений пермской фауны на кряже Прончищева: А – расположение района исследований на Севере Сибири. Звездочкой отмечен изученный участок пермских отложений в пределах кряжа Прончищева. Б – схематичная геологическая карта исследованного участка (составлена А.Б. Кузьмичевым и М.К. Данукаловой с добавлениями Е.С. Соболева). Цветом и индексами показаны: P2 – средняя пермь; P2–${\text{P}}_{{\text{3}}}^{{\text{1}}}$ – верхняя часть средней и низы верхней перми; T1 – нижний триас; T2–3 – средний–верхний триас, J1 – нижняя юра (геттанг–плинсбах); J1t – нижняя юра (тоар); ${\text{J}}_{{\text{2}}}^{{\text{1}}}$ – средняя юра (аален–бат). Белыми точками обозначены номера местонахождений пермской фауны.
Настоящая работа посвящена результатам изучения представительной коллекции морских двустворчатых моллюсков, систематически собранной авторами из пермских толщ кряжа Прончищева. Это впервые позволило провести обоснование их возраста и установить биогеографические связи с соседними пермскими бореальными акваториями.
КРАТКАЯ ИСТОРИЯ ИЗУЧЕНИЯ ПЕРМСКИХ ОТЛОЖЕНИЙ И ФАУНЫ КРЯЖА ПРОНЧИЩЕВА
Пермские отложения в пределах кряжа Прончищева впервые были обнаружены и изучены В.А. Руцковым, проводившим в этом районе геологическую съемку масштаба 1 : 200 000 (Руцков, 1955). Они были отнесены целиком к верхней перми (в старом, двучленном понимании объема системы)11 на основании находок в нижней видимой части разреза комплексов мелких фораминифер и палинокомплексов, а в средней части – одной раковины Kolymia sp. indet.
Позже, на единственной изданной на сегодня геологической карте масштаба 1 : 200 000 (Бобров, Горшкова, 1992), верхнепермские отложения кряжа Прончищева были расчленены на три безымянные толщи общей мощностью чуть более 1000 м. Толщи были охарактеризованы редкими находками морских двустворок (фестий, мионий и единичных колымиид), брахиопод-ринхопор и фораминифер (псевдонодозарий, нодозарий, фрондикулярий и ректогландулин), приуроченных, в основном, к верхней половине средней – нижней половине верхней толщ и датирующих позднюю пермь (в старом понимании).
В записке к мелкомасштабной карте третьего поколения (Проскурнин и др., 2017) стратиграфия пермских отложений кряжа Прончищева была пересмотрена (при сохранении литологической и фаунистической характеристик разреза). Авторы записки попытались выделить на кряже аналоги пермских свит Нордвикского р-на, расположенного к западу от рассматриваемой территории, где пермские отложения были изучены по скважинам (Бобров и др., 2000 и ссылки в этой работе). Так, нижняя толща была отнесена к нижнекожевниковской свите (кунгурский – уфимский ярусы нижней перми в современном понимании), а средняя толща и нижняя половина верхней толщи – к верхнекожевниковской свите (казанский – северодвинский ярусы средней–верхней перми). Вышележащие отложения были отнесены к мисайлапской свите (по-видимому, отвечающей вятскому ярусу). Однако, следует заметить, что в своей стратотипической местности последняя свита охарактеризована исключительно пресноводными комплексами микрофауны (остракод и конхострак) и листовой флорой (Бобров и др., 2000). Это делает невозможным ее выделение на кряже Прончищева, где весь видимый разрез перми сложен мелководно-морскими отложениями с соответствующей фауной, преимущественно двустворчатых моллюсков.
Таким образом, до начала наших исследований отсутствовали сколько-нибудь детальные сборы ископаемой фауны из пермских толщ кряжа Прончищева, в связи с чем оставался открытым вопрос обоснования их возраста. Кроме присутствия здесь аналогов казанского яруса, установленных по остаткам фораминифер (Бобров, Горшкова, 1992 и более ранние неопубликованные работы), другие биостратиграфические уровни не были фаунистически подтверждены. Также было неясным, насколько обосновано здесь картирование стратонов, установленных в Нордвикском р-не по скважинам. Это делало предположительным и мало обоснованным построение даже мелкомасштабных палеогеографических карт на рассматриваемую территорию (Budnikov et al., 2020).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Летом 2012 г. трем соавторам настоящей статьи (А.Б. Кузьмичеву и М.К. Данукаловой в составе экспедиционного отряда Геологического ин-та РАН, Москва и Е.С. Соболеву в составе отряда Ин-та нефтегазовой геологии и геофизики СО РАН, Новосибирск) впервые удалось осуществить целенаправленное изучение пермских отложений кряжа Прончищева. В результате были изучены все три толщи перми и собрана достаточно представительная коллекция ископаемой фауны, включающая в себя, главным образом, двустворчатых моллюсков, а также более редких гастропод и брахиопод.
Стратиграфия. Весь видимый разрез перми общей мощностью около 1400 м (рис. 2) сложен чередованием полимиктовых, в том числе известковистых, песчаников, алевролитов и аргиллитов в разных сочетаниях (рис. 3). Песчаники нередко биотурбированы, содержат растительный детрит, демонстрируют волновую рябь, горизонты уплощенной гальки эродированных местных пород. Авторами, как и предшественниками, выделено три толщи. Для нижней из них (плохо обнаженной) характерно обилие песчаников, встречены единичные развалы полимиктовых конгломератов и в одной точке – высыпки пластинчатого угля. В верхней части толщи заметную роль играют тонкообломочные породы. Для второй толщи характерно чередование глинистых или тонкоритмичных пачек (переслаивание аргиллитов и алевролитов, иногда маломощных песчаников) с менее мощными песчаниковыми. В кровле толщи залегает 50-метровая пачка аргиллитов с тонкими конкрециевидными прослоями сидерита и известняка. Завершает разрез перми толща, сложенная преимущественно песчаниками с крупномасштабной косой слоистостью, вероятно, барового типа. В основании этой толщи задокументированы многочисленные глендониты (рис. 4). В видимой кровле пермского разреза присутствуют известковистые песчаники с карбонатными конкрециями, пачки аргиллитов. Выше необнаженного интервала мощностью около 50–60 м залегают пестроцветные глины инда (?), перекрытые фаунистически охарактеризованными отложениями оленекского яруса (Бобров, Горшкова, 1992).
Рис. 2.
Сводная стратиграфическая колонка пермских отложений кряжа Прончищева. Справа от колонки показаны уровни отбора и номера образцов с фауной. Серым цветом обозначены образцы, собранные вне основного разреза. Звездочками показаны находки глендонитов. МСШ – Международная стратиграфическая шкала, ОСШ – Общая (российская) стратиграфической шкала. Сокращения в гранулометрической шкале (под колонкой): а – аргиллиты и алевролиты, ар – аргиллиты и глины, п – песчаники, к – конгломераты.

Рис. 3.
Характер обнаженности пермских пород нижней части толщи 3 по правому притоку р. Лыгый-Юряге (в районе места отбора образца 426/1, см. рис. 1, Б). Высота уступа в русле 0.7 м.


На основании определений двустворчатых моллюсков (табл. 1) и, в меньшей степени, по литологии нижнюю и среднюю толщи перми можно соотнести с верхнекожевниковской свитой Нордвикского р-на (Бобров и др., 2000). Таким образом, присутствие на кряже возрастных аналогов нижнекожевниковской свиты (Проскурнин и др., 2017) не подтверждается. Верхняя толща примерно соответствует мисайлапской свите. Однако иной литологический состав отложений, иные условия их образования и характер палеонтологических остатков указывают на необходимость отказа от использования свиты в пределах кряжа Прончищева. В задачи нашей статьи не входит разбор этого вопроса, поэтому на стратиграфической колонке название свиты дано условно.
Таблица 1.
Фауна пермских отложений кряжа Прончищева
| Бивальвиевые зоны (слои) | Местонахождения | Номера образцов с фауной | Фауна |
|---|---|---|---|
| Kolymia inoceramiformis– K. plicata | р. Песчаная | 346/1 | Kolymia rutskovi Biakov, sp. nov. (табл. V , фиг. 21–23 ) (многочисленные), Kolymia ex gr. yurii Astafieva (табл. V , фиг. 19 ), Kolymia sp. (K. ex gr. inoceramiformis Licharew ?) |
| северные выходы на правобережье р. Песчаной | 532/1 | Kolymia ex gr. yurii Astafieva | |
| 535/1 | Kolymia sp. (возможно, K. ex gr. inoceramiformis Licharew) (табл. V , фиг. 20 ) | ||
| Kolymia multiformis | 471/1 | Kolymia multiformis Biakov, K. ex gr. nikolaewi (Voronez), Maitaia vel, Evenia? sp. indet. | |
| правые притоки р. Песчаной | 2099/1-4 | отпечаток неопределимой колымииды, Myonia sp., Nuculopsis sp. indet. (табл. V , фиг. 1), Mourlonia aff. yabeshigerui (Kobayashi) | |
| 2099/6-9 | Kolymia sp. indet., Phestia sp. indet. (табл. V , фиг. 8 ) | ||
| 373/5 | Rhynchopora lobjaensis Tolmatchew (скопление) | ||
| 374/1, 2099/10-12 |
Kolymia sp. indet., Kolymia multiformis Biakov (табл. V , фиг. 10 ), Evenia lenaensis (Voronez) | ||
| 374/2, 2099/18-27 |
Kolymia cf. nikolaewi (Voronez), K. multiformis Biakov, Streblopteria ex gr. levis (Lutkevich et Lobanova), Pachymyonia sp. indet., Mourlonia aff. yabeshigerui (Kobayashi) | ||
| 374/3 | Myonia sp. | ||
| р. Песчаная (излучина) | 343/1, 2099/28-33 |
Kolymia aff. nikolaewi (Voronez), K. multiformis Biakov (табл. V , фиг. 11 ), K. ex gr. multiformis Biakov | |
| левые притоки р. Песчаной | 358/1 | Kolymia nikolaewi (Voronez), K. ex gr. nikolaewi (Voronez) (табл. V , фиг. 17 ), Evenia lenaensis (Voronez) (табл. V , фиг. 14 ). | |
| северные выходы в бассейне р. Лыгый-Юряге | 527/1 | Australomya longa (Lutkevich et Lobanova) (табл. VI , фиг. 1), Kolymia ex gr. multiformis Biakov, K. cf. pergamenti Muromzeva | |
| р. Лыгый-Юряге | 443/1, 2099/54-60 |
Kolymia nikolaewi (Voronez) (табл. V , фиг. 13, 15, 16 ) (многочисленные), K. multiformis Biakov (табл. V , фиг. 5–7, 9, 12 ), K. ex gr. multiformis Biakov (табл. V , фиг. 18 ) | |
| Maitaia bella (слои с Phestia ex gr. ovata) | 444/3, 2099/64 | Rhynchopora lobjaensis Tolmatchew (скопления), Rhynchopora sp. indet. |
|
| 449/3, 2099/65-67 |
Phestia ex gr. ovata (Laseron) (табл. V , фиг. 3), Maitaia sp. (табл. V , фиг. 24 ), Mourlonia aff. yabeshigerui (Kobayashi), Beecheria sp. | ||
| 612/1, 613/1 | Phestia sp. indet., Streblopteria cf. englehardti (Etheridge et Dun), Myonia sp. indet., Pyramus ? sp. indet., гастроподы Mourlonia aff. yabeshigerui (Kobayashi) | ||
| правый приток р. Лыгый-Юряге | 424/3 | Glyptoleda sp. indet. (табл. V , фиг. 2), Streblopteria levis (Lutkevich et Lobanova) (табл. VI , фиг. 9 ), Myonia aff. gibbosa (Maslennikow), Mourlonia aff. yabeshigerui (Kobayashi) (многочисленные) | |
| 426/1, 426/2 | Pachymyonia elata Popow (табл. VI , фиг. 3) | ||
| 426/3, 2099/68 | Streblopteria levis (Lutkevich et Lobanova) (табл. VI , фиг. 5 ), S. cf. levis (Lutkevich et Lobanova) (многочисленные), Myonia aff. gibbosa (Maslennikow) (табл. VI , фиг. 2, 7 ), Pachymyonia cf. elata (Popow) | ||
| правые притоки р. Синнигес-Песчаной | 504/4 | Streblopteria levis (Lutkevich et Lobanova) (табл. VI , фиг. 6 ) (многочисленные), Myonia aff. gibbosa (Maslennikow) | |
| левые притоки р. Синнигес-Песчаной | 302/1 | Rhynchopora lobjaensis Tolmatchew (скопление) | |
| Maitaia belliformis | между р. Лыгый-Юряге и р. Урасалах | 441/1 | Maitaia sp. indet. (возможно, M. ex gr. bella Biakov) |
| правый приток р. Лыгый-Юряге | 605/2 | Streblopteria cf. levis (Lutkevich et Lobanova) | |
| 605/1 | Maitaia bella Biakov (табл. V , фиг. 25–27 ) | ||
| Intomodesma costatum | левые притоки р. Песчаной | 516/2, 2099/43-52 | Streblopteria rotunda (Lutkevich et Lobanova) (табл. VI , фиг. 10–12 ) (многочисленные), Phestia sp. indet. (табл. V , фиг. 4) |
| правые притоки р. Песчаной | 2099/35-42 | Intomodesma ex gr. costatum Popow (табл. VI , фиг. 4), Intomodesma sp. (табл. VI , фиг. 8 ), Maitaia sp. | |
| левые притоки р. Песчаной | 2099/44, 45 | Intomodesma ? sp. indet. | |
| 2099/53 | Intomodesma ex gr. costatum Popow |
Биостратиграфия и таксономический состав комплексов. Определение коллекции двустворчатых моллюсков позволяет установить присутствие пяти биостратиграфических уровней.
В средней перми, согласно зональной бивальвиевой схеме северо-востока Азии (Бяков, 2000, 2012), выделяются:
1) зоны Kolymia inoceramiformis – K. plicata (нерасчлененные) на основании находок Kolymia ex gr. yurii Astafieva;
2) зона Кolymia multiformis, охарактеризованная Kolymia multiformis Biakov, K. ex gr. multiformis Biakov, K. nikolaewi (Voronez), Evenia lenaensis (Voronez).
В верхней перми могут быть выделены:
3) зона Maitaia bella (слои с Phestia ex gr. ovata) на основании находок Glyptoleda sp., Phestia ex gr. ovata (Laseron), Pachymyonia elata Popow, Myonia aff. gibbosa (Maslennikow), Streblopteria levis (Lutkevich et Lobanova);
4) зона Maitaia belliformis с многочисленными Maitaia bella Biakov;
5) зона Intomodesma costatum, где определены скопления крупных Streblopteria rotunda (Lutkevich et Lobanova), Intomodesma ex gr. costatum Popow и Intomodesma sp.
Первый биостратиграфический уровень (зоны Kolymia inoceramiformis – K. рlicata) охарактеризован Kolymia ex gr. yurii Astafieva (табл. V , фиг. 19 ; см. вклейку), K. rutskovi Biakov, sp. nov. (табл. V , фиг. 21–23 ) и Kolymia sp. (возможно, K. ex gr. inoceramiformis Licharew) (табл. V , фиг. 20 ). Первый из этих видов встречается на северо-востоке Азии (Верхоянье, Охотский массив и его обрамление) в интервале бивальвиевых зон Aphanaia dilatata – K. plicata (Бяков, 2012). Вид Kolymia inoceramiformis характерен преимущественно для одноименной бивальвиевой зоны, но в некоторых разрезах Верхоянья сходные формы встречены и в зоне Кolymia multiformis.
В целом на рассматриваемом стратиграфическом уровне находок фауны, представленной исключительно иноцерамоподобными двустворками, мало, и они имеют неважную сохранность.
Второй биостратиграфический уровень (зона Кolymia multiformis) является одним из наиболее хорошо охарактеризованных. Здесь установлены многочисленные Kolymia multiformis Biakov (табл. V , фиг. 5–7, 9–12 ), K. cf. multiformis, K. ex gr. multiformis (табл. V , фиг. 18 ), K. nikolaewi (Voronez) (табл. V , фиг. 13, 15, 16 ), K. aff. nikolaewi, K. ex gr. nikolaewi (табл. V , фиг. 17 ), Streblopteria ex gr. levis (Lutkevich et Lobanova) (редкие), Evenia lenaensis (Voronez) (табл.V, фиг. 14 ), Australomya longa (Astafieva-Urbajtis) (табл. VI , фиг. 1; см. вклейку), Myonia sp., Pachymyonia sp. indet., Nuculopsis sp. (табл. V , фиг. 1), Phestia sp. indet. (табл. V , фиг. 8 ). Иногда встречаются скопления брахиопод Rhynchopora lobjaensis Tolmatchew и гастропод Mourlonia aff. yabeshigerui (Kobayashi).
Перечисленные иноцерамоподобные двустворки, за исключением Evenia lenaensis, очень характерны для зоны Multiformis и встречаются практически повсеместно на северо-востоке Азии (Бяков, 2012). Вид E. lenaensis встречен в разрезах Северного Верхоянья, а также на cевере Восточно-Европейской платформы (где он описан как Atomodesma menneri Kulikov), п-ове Канин (Куликов, 1978; Канев, 2006) и в Восточной Гренландии, где он описан Н. Нюэлом (Newell, 1955) под названием Posidonia permica Newell (Бяков, 2013). Вид Australomya longa (Astafieva-Urbajtis) впервые описан с Центрального Таймыра (Люткевич, Лобанова, 1960), часто встречается также в Северном Верхоянье (Астафьева-Урбайтис и др., 1976). В целом таксономический состав описываемого комплекса очень близок к северо-верхоянскому (Biakov, Kutygin, 2018).
Streblopteria levis впервые описан с Восточного Таймыра (район мыса Цветкова) без точно установленной стратиграфической привязки из верхнепермских отложений (в старом понимании) (Люткевич, Лобанова, 1960). На северо-востоке Азии этот вид распространен в диапазоне зон Belliformis–Costatum в Верхоянье и на Омолонском массиве (Бяков, 2012). Данные находки расширяют (на две зоны) нижний диапазон стратиграфического распространения вида.
Третий биостратиграфический уровень (зона Maitaia bella, слои с Phestia ex gr. ovata) установлен на основании находок Phestia ex gr. ovata (Laseron) (табл. V , фиг. 3), многочисленных Pachymyonia elata Popow (табл. VI , фиг. 3) и Myonia aff. gibbosa (Maslennikow) (табл. VI , фиг. 2, 7 ). Хотя на кряже Прончищева в зоне Maitaia bella отсутствуют находки вида–индекса зоны, но присутствуют вышеуказанные виды зонального комплекса (Бяков, 2012), что позволяет достаточно уверенно выделять эту зону.
Также здесь встречены многочисленные Streblopteria levis (Lutkevich et Lobanova) (табл. VI , фиг. 5, 6, 9 ) и единичные Glyptoleda sp. indet. (табл. V , фиг. 2), Maitaia sp. (табл. V , фиг. 24 ), Streblopteria cf. englehardti (Etheridge et Dun), Myonia sp. indet., Pyramus? sp. indet., а также многочисленные брахиоподы Rhynchopora lobjaensis Tolmatchew (в низах), единичные Beecheria sp. и частые гастроподы Mourlonia aff. yabeshigerui (Kobayashi).
Phestia ex gr. ovata известна из разрезов Западного (Кутыгин и др., 2003) и Южного Верхоянья (определения первого автора из колл. Р.В. Кутыгина) из низов дулгалахского горизонта. Pachymyonia elata Popow встречается во многих разрезах Верхоянья, Охотского массива и его обрамления, на Омолонском массиве и на Центральном и Восточном Таймыре в узком стратиграфическом диапазоне – зоне Multiformis и низах зоны Bella. Myonia aff. gibbosa (Maslennikow), определенная нами, несколько отличается от типичных экземпляров, описанных Д.Ф. Масленниковым с Омолонского массива (Каширцев, 1959) из более молодых (хивачских = хальпирских) отложений. Аналогичные нашим экземпляры нередко встречаются в разрезах Западного Верхоянья на рассматриваемом биостратиграфическом уровне (Кутыгин и др., 2003).
Четвертый биостратиграфический уровень (зона Maitaia belliformis) выделен на основании многочисленных находок Maitaia bella Biakov (табл. V , фиг. 25–27 ). Хотя сам вид–индекс зоны belliformis здесь не обнаружен, именно для этой зоны характерны многочисленные находки (эпиболь) вида M. bella, в частности, в стратотипических разрезах зоны на Омолонском массиве (Бяков, 2012). Такая же картина наблюдается в большинстве разрезов северо-востока Азии, правда, там, как правило, присутствуют находки и самого вида–индекса. Поэтому мы сочли возможным выделить и на кряже Прончищева эту зону. Этот вид очень широко распространен не только на северо-востоке Азии, но и на Новой Земле (Бяков, 2013), Восточном Таймыре (авторские определения из колл. М.К. Данукаловой и А.Б. Кузьмичева), в Восточном Забайкалье, Приморье (Бяков, 2013) и, вероятно, на Шпицбергене (Bond et al., 2015). В последнем регионе “Retroceramus sp.” (там же, рис. 6F ), скорее всего, является M. bella. Также на рассматриваемом уровне встречены многочисленные Streblopteria cf. levis.
Пятый биостратиграфический уровень (зона Intomodesma costatum) установлен на основании находок местами многочисленных Intomodesma ex gr. costatum Popow (табл. VI , фиг. 4), Intomodesma sp. (табл. VI , фиг. 8 ), а также крупных Streblopteria rotunda (Lutkevich et Lobanova) (табл. VI , фиг. 10–12 ), образующих массовые скопления. Такие ракушечники характерны для разрезов Восточного Таймыра (Люткевич, Лобанова, 1960). Также здесь встречены единственная Phestia sp. indet. (табл. V , фиг. 4) и более многочисленные Maitaia sp.
Intomodesma costatum, вид–индекс одноименной бивальвиевой зоны, широко распространен на северо-востоке Азии (Бяков, 2012, 2013) и встречен на Новой Земле (Муромцева, 1981). Streblopteria rotunda впервые был описан с Восточного Таймыра без точно установленной стратиграфической привязки из “верхнепермских (в старом понимании) отложений” (Люткевич, Лобанова, 1960). На северо-востоке Азии изредка встречается в низах зоны Costatum (подзона Maitaia hurenensis) Северного и Западного Верхоянья (Кутыгин и др., 2003) и восточного обрамления Охотского массива (Бяков, 2007).
Определения пермской макрофауны с кряжа Прончищева сведены в табл. 1.
СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
В описании приняты сокращения: В – высота раковины, Вп – выпуклость створки, ГД – длина раковины по главной диагонали, Д – длина раковины, ДУ – длина переднего ушка, ДЗК – длина замочного края раковины, МУ – макушечный угол, УС – угол скоса створки.
Изученный материал хранится в музее Северо-Восточного комплексного научно-исследовательского ин-та им. Н.А. Шило ДВО РАН (СВКНИИ), г. Магадан, колл. № 02-06.106; в Центре коллективного пользования (ЦКП) “Геохрон” Ин-та нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН, г. Новосибирск, колл. № 2099.
СЕМЕЙСТВО KOLYMIIDAE KUSNEZOV, 1973
Род Kolymia Licharew, 1941
Kolymia rutskovi Biakov, sp. nov.
Табл. V , фиг. 21–23
Название вида в память геолога В.А. Руцкова, впервые установившего пермские отложения на кряже Прончищева.
Голотип – СВКНИИ, № 12/02-06.106, ядро правой створки; север Якутии, кряж Прончищева, р. Песчаная; средняя пермь, средняя часть деленджинского горизонта, бивальвиевые зоны Kolymia inoceramiformis–K. plicata.
Описание. Раковина от небольшого до среднего размера, до 60 мм по ГД, треугольно-овальная, от более развитой в длину до чуть вытянутой в высоту (Д : В = 0.94–1.11), умеренно скошенная (УС = 50°–60°). У большинства изученных экземпляров на задне-нижнем поле раковины имеется слабо выраженная депрессия, подчеркиваемая слабым изгибом концентрических морщин-складок. Замочный край длинный, значительно больше половины длины створок (ДЗК : Д = 0.60–0.72). Передний край почти прямой или слабо вогнутый, длинный, по крутой дуге переходит в выпуклый длинный нижний край, который постепенно переходит в почти прямой или в различной степени выпуклый, умеренно длинный задний край. Последний косо соединяется с замочным краем под углом 125°–135°. Выпуклость створок умеренная (Вп : В = 0.22–0.27), точка наибольшей выпуклости расположена примерно посередине створки или чуть сдвинута вверх и несколько приближена к переднему краю.
Макушки умеренно высокие, широкие, нависающие над замочным краем, притупленные. Скульптура створок представлена сглаженными, относительно неправильными концентрическими морщинами-складками, хорошо выраженными на ядрах раковин. На заднее-нижней части раковины скульптура более неправильная. Переднее ушко обычно хорошо выражено, от маленького до умеренно развитого (ДУ : Д = 0.08–0.16), в виде неправильного, вытянутого в высоту треугольника. Заднее крыло, как правило, хорошо выражено, умеренной величины, обычно хорошо обособлено от остальной части раковины. Призматический слой, реликты которого сохранились кое-где на ядрах раковин (в частности, у голотипа), был очень тонким, доли мм в толщину.
Размеры в мм и отношения:
| Экз. № | Д | В | Вп | ГД | ДЗК | ДУ | Д : В | Вп : В | ДЗК : Д | ДУ : Д | УС | ДУ |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 12/02-06.106 голотип | 37.7 | 34 | 8 | 41 | 26.5 | 6 | 1.11 | 0.24 | 0.70 | 0.16 | 55° | 60° |
| 11/02-06.106 | 34.6 | 34 | 8 | 39.2 | 20.5 | 3 | 1.02 | 0.22 | 0.60 | 0.09 | 50° | 60° |
| 14/02-06.106 | 50 | 45.2 | 11 | 58.2 | 31.3 | 4 | 1.11 | 0.24 | 0.63 | 0.08 | 50° | 60° |
| 13/02-06.106 | 29.8 | 31.6 | 8.5 | 34.8 | 21.5 | 4 | 0.94 | 0.27 | 0.72 | 0.13 | 60° | 65° |
Изменчивость. Может варьировать отношение Д : В (от 0.94 до 1.11); у более зрелых экземпляров оно более единицы. Изменяется также степень выраженности переднего ушка – от слабо заметного до хорошо сформированного, а также степень выраженности депрессии на задне-нижнем поле раковины.
Сравнение. По форме раковины некоторые экземпляры имеют сходство с K. nikolaewi (Voronez), впервые описанным Н.С. Воронец (1936) из верхов деленжинского горизонта Северного Верхоянья, Булкурская антиклиналь, отличаясь более треугольной, расширяющейся в нижне-заднем направлении раковиной, как правило, имеющей слабо выраженную депрессию, неправильной скульптурой и обычно более крупными размерами. От K. yurii Astafieva (Астафьева, 1993), с которым новый вид также имеет некоторое сходство по очертаниям раковины, отличается гораздо меньшей (почти в два раза) выпуклостью створок, присутствием депрессии на задне-нижнем поле, более неправильной скульптурой и, как правило, меньшими размерами.
Материал. 7 ядер створок из типового местонахождения; обр. 346/1.
ЗАКЛЮЧЕНИЕ
Изучение коллекции двустворчатых моллюсков позволяет сделать следующие выводы. Доказано присутствие на кряже Прончищева верхнего и среднего отделов перми, где установлено пять биостратиграфических уровней.
В средней перми выделяются зоны Kolymia inoceramiformis–K. plicata (нерасчлененные) и Кolymia multiformis. В верхней перми могут быть выделены зоны Maitaia bella (слои с Phestia ex gr. ovata), Maitaia belliformis и Intomodesma costatum. Кроме двустворчатых моллюсков, в зоне Кolymia multiformis определены также брахиоподы Rhynchopora lobjaensis Tolmatchew и гастроподы Mourlonia aff. yabeshigerui (Kobayashi), а в зоне Maitaia bella их дополняют единичные брахиоподы Beecheria sp.
На основании приведенных определений можно сделать некоторые палеобиогеографические и палеогеографические выводы. Комплексы двустворок средней и большей нижней части верхней перми (до зоны Intomodesma costatum) по своему таксономическому составу близки к комплексам Западного и Северного Верхоянья (Муромцева, Гуськов, 1984; Кутыгин и др., 2003; Бяков, 2013; Biakov, Kutygin, 2018). В то же время, в нижней–средней части средней перми присутствует специфичный новый вид Kolymia rutskovi Biakov, sp. nov., а в верхней части средней перми нет характерных североверхоянских эндемиков – K. verchoyanica Lutkevich et Lobanova и K. peregoedovi Biakov.
В низах верхней перми много пектинид Streblopteria levis (Lutkevich et Lobanova), присутствуют своеобразные Myonia aff. gibbosa (Maslennikow), Pachymyonia elata Popow, характерные как для Северного Верхоянья, так и для Восточного Таймыра. Особенно следует отметить скопления стреблоптерий, доминирующих в восточно-таймырских мелководных сообществах. Здесь же встречены многочисленные глендониты, фиксирующие эпизод похолодания климата.
Ископаемые комплексы верхов перми очень своеобразны и представлены почти исключительно монотаксонными скоплениями крупных (до 10 см) пектинид Streblopteria rotunda (Lutkevich et Lobanova) и отдельных представителей родов Intomodesma и Maitaia. Такой состав позднепермских сообществ известен только на Восточном Таймыре (Люткевич, Лобанова, 1960) и, очевидно, характеризует мелководные прибрежные фации.
Таким образом, можно сделать вывод, что на протяжении почти всей перми морской бассейн, существовавший на территории кряжа Прончищева, имел устойчивые связи с Верхоянским. Этот бассейн был весьма мелководным и, возможно, испытывал некоторое опреснение, о чем свидетельствует резко обедненный состав сообществ брахиопод, полное отсутствие среди них продуктид, и достаточно бедный в систематическом отношении состав комплексов двустворок. В конце перми палеогеографическая ситуация изменилась, и резко усилились связи бассейнов кряжа Прончищева и Восточного Таймыра. На протяжении перми эти связи постоянно усиливались и достигли максимума в конце пермского периода, о чем свидетельствует постепенное увеличение доли восточно-таймырских элементов в биоте первого.
Работа выполнена при поддержке РФФИ, проекты 18-05-00191, 19-05-00926 и 20-05-00604, а также за счет средств субсидии, выделенной Казанскому федеральному университету для выполнения государственного задания № 671-2020-0049 в сфере научной деятельности и в соответствии с госзаданиями СВКНИИ ДВО РАН, ГИН РАН и ИНГГ СО РАН (проект ФНИ № 0331-2019-0004). Часть исследований (палеогеографические выводы) выполнена по проекту РНФ № 19-17-00178.
Авторы выражают признательность А.В. Ядрёнкину, А.Ю. Попову и Л.Б. Хазину (ИНГГ СО РАН) за помощь в сборах ископаемой фауны, а также Т.В. Филимоновой (ГИН РАН) за консультации по пермским фораминиферам, И.Л. Ведерникову (СВКНИИ ДВО РАН) за подготовку изображений двустворчатых моллюсков, С.В. Попову и А.В. Гужову (ПИН РАН), замечания и предложения которых сделали текст статьи более полным и логично построенным.
Список литературы
Астафьева М.М. Пермские иноцерамоподобные двустворчатые моллюски России. М.: Наука, 1993. 129 с.
Астафьева-Урбайтис К.А., Лобанова О.В., Муромцева В.А. Род Myonia (Bivalvia) в перми Северо-Востока СССР // Палеонтол. журн. 1976. № 1. С. 27–40.
Бобров В.Н., Бардеева М.А., Клейкова Н.И. и др. Легенда Оленекской серии листов Государственной геологической карты Российской Федерации масштаба 1 : 200 000 (издание второе). Объяснительная записка. М.: ФГУ НПП “Аэрогеология”, 2000. 172 с.
Бобров В.Н., Горшкова Е.Р. Государственная геологическая карта СССР. Масштаб 1 : 200 000. Оленекская серия. Листы S-50-XIX, XX. Л.: Картфабрика ВСЕГЕИ, 1992.
Бяков А.С. Зональное расчленение перми Северо-Востока Азии по двустворчатым моллюскам // Стратигр. Геол. корреляция. 2000. Т. 8. № 1. С. 35–54.
Бяков А.С. Биостратиграфия пермских отложений Северного Приохотья (Северо-Восток Азии) // Стратигр. Геол. корреляция. 2007. Т. 15. № 2. С. 47–71.
Бяков А.С. Новая зональная схема пермских отложений Северо-Востока Азии по двустворчатым моллюскам. Статья 1. Зональное расчленение // Тихоокеанск. геол. 2012. Т. 31. № 5. С. 13–40.
Бяков А.С. Новая зональная схема пермских отложений Северо-Востока Азии по двустворчатым моллюскам. Статья 2. Вопросы корреляции // Тихоокеанск. геол. 2013. Т. 32. № 1. С. 3–17.
Воронец Н.С. Мезозойская фауна хребта Хараулахского // Тр. Арктич. ин-та. 1936. Т. 37. С. 7–36.
Канев Г.П. Класс Bivalvia // Верхняя пермь полуострова Канин. М.: Наука, 2006. С. 116–119.
Каширцев А.С. Полевой атлас фауны пермских отложений Северо-Востока СССР. М.: Изд-во АН СССР, 1959. 85 с.
Куликов М.В. О некоторых редких представителях моллюсков из нижнеказанских отложений Русской платформы // Докл. АН СССР. 1978. Т. 241. № 1. С. 177–180.
Кутыгин Р.В., Будников И.В., Бяков А.С. и др. Опорный разрез дулгалахского и хальпирского горизонтов Западного Верхоянья // Тихоокеанск. геол. 2003. Т. 22. № 6. С. 82–97.
Люткевич Е.М., Лобанова О.В. Пелециподы перми советского сектора Арктики. Л.: Гостоптехиздат, 1960. 294 с.
Муромцева В.А. Двустворчатые моллюски // Пермские отложения Новой Земли. Л.: Наука, 1981. С. 26–51.
Муромцева В.А., Гуськов В.А. Пермские морские отложения и двустворчатые моллюски Советской Арктики. Л.: Недра, 1984. 208 с.
Проскурнин В.Ф., Шкарубо С.И., Заварзина Г.А. и др. Государственная геологическая карта Российской Федерации. Масштаб 1 : 1 000 000 (третье поколение). Серия Лаптево-Сибироморская. Лист S-50. Объяснительная записка. СПб.: Картфабрика ВСЕГЕИ, 2017. 264 с.
Руцков В.А. Геологическое строение кряжа Прончищева. Отчет о комплексной геологической съемке масштаба 1 : 200 000, проведенной летом 1953 года в северной части междуречья Анабар-Оленек партией № 1 третьей Оленекской экспедиции. Якутск, 1955. 276 с.
Biakov A.S., Kutygin R.V. Bivalves from the Delenzhian-Dulgalakhian boundary beds of the Middle Permian of the lower reaches of the Lena River (Northern Verkhoyansk region, Northern Siberia) // Paleontol. J. 2018. V. 52. № 7. P. 761–767.
Bond D.P.G., Wignall P.B., Joachimski M.M. et al. An abrupt extinction in the Middle Permian (Capitanian) of the Boreal Realm (Spitsbergen) and its link to anoxia and acidification // GSA Bull. 2015. https://doi.org/10.1130/B31216.1
Budnikov I.V., Kutygin R.V., Shi G.R. et al. Permian stratigraphy and paleogeography of Central Siberia (Angaraland) − A review // J. Asian Earth Sci. 2020. V. 196. https://doi.org/10.1016/j.jseaes.2020.104365
Newell N.D. Permian pelecipods of East Greenland // Medd. Grønland undgivne af kommissionen for videnskabelige undersogelser i Grønland. 1955. Bd 110. № 4. 48 p.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал