

Палеонтологический журнал, 2021, № 3, стр. 92-103
Мелкие позвоночные из плейстоценовых отложений восточной галереи Денисовой пещеры, Северо-Западный Алтай
А. К. Агаджанян a, *, М. В. Шуньков b, **, М. Б. Козликин b, ***
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Институт археологии и этнографии СО РАН
Новосибирск, Россия
* E-mail: aagadj@paleo.ru
** E-mail: shunkov77@gmail.com
*** E-mail: kmb777@yandex.ru
Поступила в редакцию 07.08.2020
После доработки 17.09.2020
Принята к публикации 29.09.2020
Аннотация
Мелкие позвоночные из плейстоценовых отложений восточной галереи Денисовой пещеры свидетельствуют о том, что на протяжении всей истории осадконакопления в ее окрестностях сохранялась высокая мозаичность биотопов. Лесные массивы чередовались с участками луговой и степной растительности. Показано, что изменение тафоценозов различных слоев коррелируется с динамикой климата, которую отражают стадии Морской изотопной шкалы.
ВВЕДЕНИЕ
Денисова пещера – один из важнейших палеолитических памятников Северо-Западного Алтая (Деревянко, Молодин, 1994; Деревянко и др., 2003, 2014). Его изучение дало много новых материалов, которые проливают свет на ранние этапы становления человека (Деревянко, 2015). В палеолитических слоях были найдены останки древнего человека, из которых были выделены митохондриальная и ядерная ДНК (Prüfer et al., 2014; Brown et al., 2015). Получена ДНК и непосредственно из осадков полости пещеры (Slon et al., 2017a, b). Вариабельность нуклеотидных замен мтДНК разных образцов указывает на длительное, более 200 тыс. лет, существование денисовского человека в долине Ануя (Sawyer et al., 2015). Установлено, что пещеру наряду с денисовцами заселяли неандертальцы – денисовцы (Viola et al., 2012; Деревянко, 2015). Доказаны случаи метисации между представителями этих популяций (Slon et al., 2018). Эти уникальные результаты показали более сложную схему ранней истории рода Homo, чем считалось ранее. При создании новых моделей эволюции человека необходимо учитывать динамику природной среды на протяжении каждого этапа ее преобразований. Это позволяет подойти к пониманию связи и взаимозависимости эволюции человека и развития природных комплексов, в которых эта эволюция происходила.
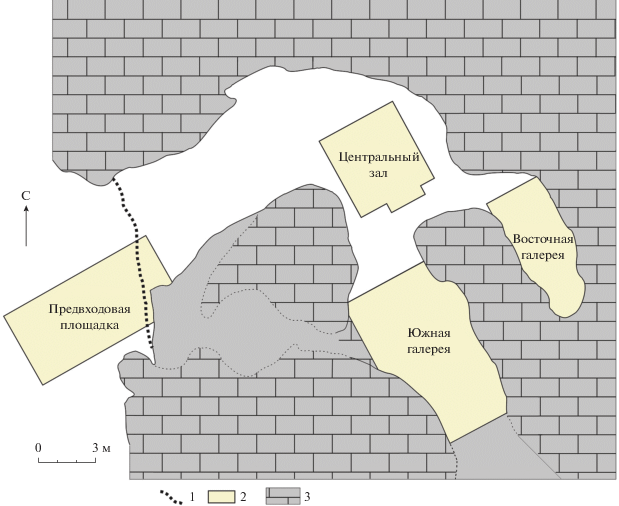
Денисова пещера расположена в центральной части Северо-Западного Алтая на юге Алтайского края на правом берегу р. Ануй, ее координаты: 51°23′51′′ N, 84°40′36′′ Е (рис. 1). Пещера состоит из центрального зала и двух галерей.
Рис. 1.
Географическое положение Денисовой пещеры и других близлежащих палеолитических памятников Северо-Западного и Центрального Алтая: 1 – Денисова пещера; 2 – стоянка Усть-Каракол; 3 – Каминная пещера; 4 – пещера Окладникова; 5 – Усть-Канская пещера.

Настоящая работа посвящена результатам изучения остатков мелких позвоночных, преимущественно млекопитающих, из восточной галереи (секторы 5 и 6) Денисовой пещеры (рис. 2). При раскопках порода каждого слоя промывалась на ситах с ячеей 1 × 1 мм. Измерения и определение материала проводились под микроскопом МБС‑10 при увеличении не менее ×10. Для уточнения диагностики привлекались коллекции Палеонтологического ин-та им. А.А. Борисяка РАН и Зоологического музея МГУ.
РЕЗУЛЬТАТЫ
Из плейстоценовых отложений восточной галереи (секторы 5 и 6) Денисовой пещеры для определения было изучено 17559 костей мелких позвоночных. Далее приведен их таксономический состава от нижних слоев к верхним.
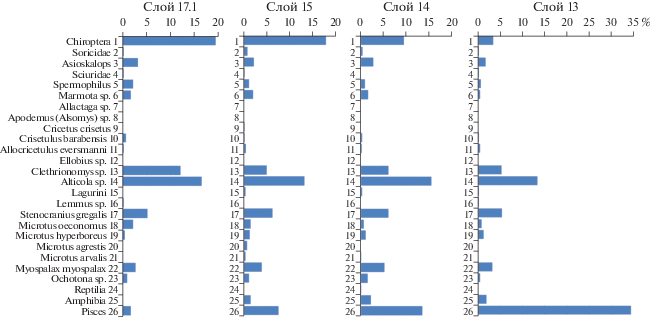
Из слоя 17.1 нижней части разреза получено 2202 костных остатка. Средняя мощность этого слоя 30–35 см, местами до 1 м. Кости светло-коричневого, почти желтого цвета. Они имеют матовую “припудренную” поверхность, что обусловлено мельчайшими частичками сцементированных карбонатов. В составе тафоценоза преобладают летучие мыши – 19.46% от общего количества мелких позвоночных (рис. 3 ). Вторая группа по численности – скальные полевки Alticola (16.5%) и рыжие полевки Clethrionomys (12.1%). Узкочерепная полевка составляет 5.2% тафоценоза, полевка-экономка Microtus oeconomus – 2.2%, северосибирская полевка Microtus hyperboreus – 0.5%. В небольшом количестве присутствуют лемминги рода Lemmus – 0.2%. Установлено наличие длиннохвостого суслика Spermophilus – 2.22%, сурка Marmota sp. – 1.7%, хомяков: барабинского – 0.74% и Эверсманна 0,25%. Среди насекомоядных численность землероек составляет 0.25%, а крота Asioscalops – 3.2%. Роющие грызуны представлены цокором Myospalax myospalax – 2.7%. В составе тафоценоза отсутствуют амфибии и рептилии, что не случайно при общем значительном объеме исследованного материала. Carnivora представлены родом Mustela – 0.74%. Относительно много молочных зубов медведей – 2.45%. Такое высокое их содержание свидетельствует о регулярном использовании пещеры медведями для зимней спячки, что предполагает очень редкое или полное отсутствие визитов древнего человека в Денисову пещеру в период формирования слоя 17.1. Это подтверждает отсутствие в осадках орудий палеолитического человека и высокая численность летучих мышей. Перечисленные факты хорошо согласуются друг с другом.
Рис. 3.
Количественное соотношение основных экологических групп мелких позвоночных из слоев 17.1, 15, 14 и 13 восточной галереи Денисовой пещеры в %.
Состав мелких позвоночных из слоя 17.1 свидетельствует о том, что в период формирования этой части разреза лесные массивы чередовались с биотопами скальной и луговой растительности. Отсутствие рептилий и лягушек указывает на низкие летние температуры. Распределение костей мелких позвоночных по уровням слоя 17.1 позволяет сделать дополнительные заключения.
Наибольшее количество костей крота, рыжих полевок, скальных полевок, цокора приходится на верхнюю половину слоя 17.1 (уровни 4–2). Максимум рыжих полевок приходится на средние уровни, что указывает на смягчение природных условий в период накопления осадков указанных уровней. Лишь в самом верхнем уровне 1 эти показатели снижаются. В заключительную фазу накопления слоя 17.1 произошло довольно сильное, но кратковременное похолодание, о чем свидетельствует пылеватый налет карбонатов на костях из этого слоя. На прохладный климат в целом указывает также наличие лемминга, полное отсутствие костей лягушек и данные палинологического анализа (Болиховская и др., 2016). Однако при этом длительного промерзания почвы в долине Ануя не происходило, о чем говорит постоянное присутствие крота Asioscalops и цокора Myospalax. Примечательно, что лемминг обнаружен в осадках уровня 10, где рыжие полевки Clethrionomys отсутствуют полностью. Осадки этого уровня накапливались в эпоху наибольшего похолодания.
В целом, время накопления данного слоя можно сопоставить с самаровской ледниковой эпохой или со стадией 8 Морской изотопной шкалы (МИС) – MIS. По данным изучения изотопного состава δ18О донных осадков Мирового океана (Lisiecki, Raymo, 2005), похолодание МИС 8 было менее сильным, чем предшествующее похолодание МИС 10 и последующее похолодание МИС 6. Кроме того, похолодание МИС 8 в начале и в конце осложнялось короткими эпизодами потепления, которые, по-видимому, и отражает состав фауны мелких позвоночных из слоя 17.1.
Слой 15 сравнительно маломощный: 15–20 см, из него было получено 835 костей и зубов мелких позвоночных (рис. 3 ). Набор таксонов в этом тафоценозе напоминает таковой слоя 17.1, но их количественное соотношение изменяется. На долю рукокрылых приходится 17.99%. Важную роль играют полевки: скальные Alticola – 13.3%, и рыжие Clethrionomys – 4.92%. Немного увеличивается количество серых полевок: узкочерепная полевка Stenocranius gregalis составляет 6.25%, полевка-экономка Microtus oeconomus – 1.4%, северосибирская полевка M. hyperboreus – 1.33%. Отмечена пашенная полевка M. agrestis, но лишь в малом количестве – 0.66%. Найдено здесь и несколько зубов степной пеструшки Lagurus lagurus – 0.3%. В слое 15 найдено четыре фрагмента моляров водяной полевки Arvicola, что составляет 0.39%. Они имеют морфологические признаки, отличные от моляров позднеплейстоценовой Arvicola cf. sapidus и от современной A. terrestris. Древнейшие представители этого рода — A. mosbachensis лихвинского времени имели структуру эмали моляров, аналогичную древним полевкам Mimomys. На протяжении второй половины среднего плейстоцена происходила ее перестройка. A. sapidus начала позднего плейстоцена имели эмаль промежуточного типа (Heinrich, 1982, 1987, 1990; Koenigswald, 1982; Agadzhanyan, 1983, 2009; Агаджанян, 2003).
Участие хомяков в составе тафоценоза слоя 15 довольно низкое – 0.76%, но среди них, в отличие от слоя 17.1, появляется хомяк обыкновенный Cricetus criсetus – 0.19%. Изменяется соотношение наземных беличьих: количество суслика Spermophilus снижается – 1.14%, а сурка Marmota – увеличивается до 1.99% (рис. 3 ). Среди насекомоядных численность землероек нарастает до 0.76%, а доля крота Asioscalops снижается до 2.2%. Роющие грызуны представлены цокором Myospalax myospalax, численность которого – 2.7%. В составе тафоценоза появляются лягушки – 1.4% и существенно увеличивается количество костей рыб – до 7.6%. Хищники представлены куньими: Mustela – 0.19% и Martes – 0.19%, и молочными зубами медведей – 0.19%.
Приведенные цифры указывают на то, что природные условия в начальный период накопления слоя 15 были примерно такие же, как во время накопления слоя 17.1. В тафоценозе лишь немного сокращается количество летучих мышей и увеличивается число костей рыб. Снижается численность рыжих и скальных полевок. Это предполагает сокращение площадей, занятых лесами, на раннем этапе седиментогенеза более чем вдвое, и уменьшение площади скальных биотопов приблизительно на четверть. Одновременно происходило расширение лугов с густым высокотравием. От нижних уровней к верхним нарастает численность рыжих лесных полевок. На уровнях 14–12 Clethrionomys составляют 2.8%, на уровнях 11 и 10 – 3.9%, на уровнях от 9 до 4 – 7.3%. К верхним уровням слоя 15 приурочены находки бурундука и лягушек, что свидетельствует о расширении площади лесных биотопов и о наиболее благоприятных климатических условиях в период накопления средней и, особенно, верхней части слоя 15. Такое изменение состава фауны позвоночных отражает постепенное потепление и увлажнение климата. Очевидно, в этот период происходила сложная перестройка природных комплексов от самаровской холодной эпохи к сообществам ширтинского межледниковья, или времени МИС 7.
Из слоя 14 в определении было использовано 5629 образцов мелких позвоночных. Костный материал светло-коричневого цвета, хорошей сохранности, не имеет следов омарганцевания и осаждения карбонатов. Количественное соотношение позвоночных напоминает предшествующий слой. В этом слое происходит дальнейшее снижение численности летучих мышей до 9.5% и одновременно нарастание количества костей рыб до 13.5% (рис. 3 ). Среди останков грызунов зарегистрирована белка Sciurus vulgaris – 0.03%. Наземные беличьи представлены сусликом Spermophilus – 0.96% и сурком Marmota – 1.65%. Найдена мышовка Sicista sp. – 0.07% и лесная мышь – Apodemus (Alsomys) – 0.05%. Как и в слое 15, доминируют скальные (Alticola) – 15.5% и рыжие (Clethrionomys) – 6.14% полевки. Немного увеличивается количество Microptini. Становится ощутимым участие в тафоценозе степной пеструшки Lagurus – 0.38%, хотя, конечно, их роль в сообществах была в целом незначительной. Интересна находка лемминга рода Lemmus, что составляет 0.02%. Это указывает на существование растительных ассоциаций с участием зеленых мхов Bryales, которые, правда, занимали незначительные площади. Важной является находка водяной полевки – 0.1%. По своей морфологии она тяготеет к Arvicola cf. sapidus микулинского времени.
Численность цокора Myospalax myospalax в слое 14 увеличивается до 5.2%, лягушек – до 2.3% (132 экз.). Впервые в палеонтологической летописи восточной галереи появляются ящерицы, 12 экз. – 0.21% (рис. 3 ). Количество землероек Soricidae уменьшается до 0.4%, а крота Asioscalops увеличивается до 2.8%. На долю пищух приходится 1.6%. Среди них отмечена степная пищуха Ochotona pusilla, правда, в очень малом количестве – 0.02%. Из хищников присутствуют куньи родов Mustela – 0.2% и Martes – 0.1%, довольно многочисленны молочные зубы медведей – 0.4%.
Формирование слоя 14, судя по таксономическому составу тафоценоза, происходило в условиях некоторого расширения площади лесных массивов и одновременного увеличения доли ксерофитных скальных и открытых биотопов. Время формирования уровней 10–8 характеризуется значительным потеплением и некоторым увлажнением климата. Именно на эти уровни приходится наибольшее количество крота, рыжих полевок, цокора и лягушек; установлено присутствие белки и ящериц. В целом климат этого времени был значительно теплее, чем во время накопления слоя 15. Верхняя часть слоя 14 характеризуется ростом доли скальных полевок, сусликов, сурка, степных пеструшек и уменьшением численности землероек, что свидетельствует о снижении влагообеспеченности. Время накопления слоя 14 можно сопоставить с ширтинским межледниковьем, т.е., со второй половиной МИС 7. Ухудшение природной обстановки на заключительных этапах седиментогенеза отражает преддверие последующей ледниковой эпохи.
Слой 13 содержит костный материал светло-коричневого цвета, хорошей сохранности, он не имеет следов осаждения марганца и карбонатов. В определении использовано 2738 образцов. Численность летучих мышей в этом слое резко уменьшается до 3.3% (рис. 3 ). Количество костей рыб увеличивается до 34.3%. Снижается участие рыжих и скальных полевок. На долю Clethrionomys приходится 5.2%, на долю Alticola – 13.27%. Среди Microtini самая многочисленная Stenocranius gregalis – 5.3% состава тафоценоза. В слое 13 снижается численность крота до 1.7%, сусликов до 0.6%, сурков до 0.4%, барабинского хомячка Crisetulus barabensis – до 0.14%. Но доля хомячка Эверсманна увеличивается почти в два раза, до 0.4%. Впервые в палеонтологической летописи восточной галереи появляется слепушонка Ellobius sp., составляя 0.1%. В небольшом количестве присутствует лемминг Lemmus sp. – 0.14%. Доля цокора Myospalax myospalax снижается до 3.2%. Падает численность лягушек до 1.8%. Более чем вдвое сокращается численность землероек Soricidae. На общем фоне падения численности наземных позвоночных происходит увеличение количества древесных беличьих: формально в 5 раз по сравнению со слоем 14. На белку Sciurus vulgaris приходится 0.1%, на бурундука Eutamias sibiricus – 0.04%. В небольшом количестве в слое 13 найдены хищники: Mustela – 0.04% и Martes – 0.04%, молочные зубы медведей – 0.04%.
Приведенные цифры свидетельствуют о сложных преобразованиях природы в долине Ануя при накоплении слоя 13 (рис. 3 ). В целом, в это время происходило падение численности большинства наземных млекопитающих, в том числе землероев (крота и цокора), появилась слепушонка Ellobius – роющий вид грызунов. Возросла численность хомячка Allocricetulus eversmanni, обитателя злаково-полынных степей. Все это говорит о значительном похолодании и аридизации климата, которые происходили в период формирования уровней 6–4. Анализ микроструктуры осадков слоя 13 также выявляет периодическое, возможно сезонное, промерзание средней части толщи (Morley et al., 2019). В то же время, осадки уровней 9–8 и 3–2 характеризуются наличием древесных беличьих, повышением численности рыжих полевок и цокора, что отражает увеличение площади лесных массивов в условиях более теплого климата. По-видимому, формирование слоя 13 происходило в условиях чередования холодных и теплых климатических фаз. Данные пыльцевого анализа также свидетельствуют о сложной трехэтапной истории природных условий этого периода (Болиховская и др., 2017).
Можно предполагать, что накопление слоя 13 пришлось на первую половину тазовской холодной эпохи в начале МИС 6, когда общее похолодание нарушалось периодическими потеплениями климата (Lisiecki, Raymo, 2005).
Из слоя 12.3 было получено 7061 определимых образцов. Они светло-коричневого цвета, хорошей сохранности, не имеют следов осаждения марганца и карбонатов. Для тафоценоза этого слоя характерно дальнейшее снижение численности летучих мышей до 2.86%, при одновременном снижении количества костей рыб до 22.2%. Численность рыжих и скальных полевок, напротив, увеличивается: Clethrionomys до 7.04%, Alticola до 18.1% (рис. 4 ). Таксономическое разнообразие серых полевок сокращается. Численность обычных для пещеры видов Microtini остается на прежнем уровне. Найдена водяная полевка Arvicola – 0.13%, которая по морфологическим признакам близка Arvicola cf. sapidus позднего плейстоцена. Немного увеличивается количество степной пеструшки Lagurus – 0.6%. Изменяется количество наземных беличьих: численность суслика увеличилась до 0.8%, сурка – уменьшилась до 0.3%. Найдена лесная мышь Apodemus (Alsomys) sp., на долю которой приходится 0.014%. Хомячки представлены двумя видами: Crisetulus barabensis – 0.6% и Allocricetulus eversmanni – 0.3%. Присутствует, но очень редок, лемминг Lemmus – 0.01%. Численность пищух равна 1.02%. Установлено присутствие степной пищухи Ochotona pusilla в количестве 0.04%. Появляются ящерицы (Lacertilia) – 0.06%, и довольно много лягушек – 2.2%. Видовое разнообразие и численность землероек Soricidae немного увеличивается и достигает 0.2%. Поднялась численность крота до 2.02%, а численность цокора, напротив, снизилась до 2.7%. Среди хищников определены куньи: Mustela 0.13%, Martes – 0.06%, и молочные зубы крупных хищников – 0.4%.
Рис. 4.
Количественное соотношение основных экологических групп мелких позвоночных из слоев 12.3, 12.2, 12.1, 11.4 и 11.3 восточной галереи Денисовой пещеры в %.

Количественное соотношение различных групп мелких позвоночных (рис. 4 ) указывает на сложную историю развития природных комплексов в период формирования слоя 12.3. Падение численности рыжих полевок в осадках уровней 7, 4 и 1 отражает значительную редукцию лесов, что соответствует похолоданию и некоторой аридизации климата.
Вместе с тем, в осадках уровня 5 рыжие полевки составляют 8.7%, найдена лесная мышь Apodemus (Alsomys), происходит уменьшение численности сурка (рис. 4 ), что отражает некоторое увеличение площади лесов.
Кажущееся противоречие в составе тафоценоза объяснимо сложной динамикой природных условий в пределах возрастного интервала слоя 12.3. Анализ распределения остатков позвоночных по горизонтам указывает на то, что на фоне общей криоксеротической обстановки в эпоху накопления слоя 12.3 происходили временные потепления и увлажнения климата, что приводило к описанному выше изменению состава фауны мелких позвоночных. Начало одного из них приходится на уровень 6, максимум фиксируется на уровне 5, спад отмечен в осадках уровня 4. Еще одну флуктуацию потепления фиксируют осадки уровня 3, где обилие Clethrionomys достигает 9.05%.
Такая ритмика условий могла соответствовать тазовской ледниковой эпохе, т.е., МИС 6, которая, впрочем, не приводила к формированию покровных ледников в пределах Горного Алтая. Кроме того, в середине данного интервала (уровни 6–5 и 3) ясно фиксируется временное потепление и увлажнение климата. Эти выводы согласуются с результатами пыльцевого анализа, который указывает на чередование перигляциальной лесотундры, тундро-лесостепи, перигляциальной тундры (Болиховская и др., 2017). По данным изотопного анализа δ18О морских осадков (Lisiecki, Raymo, 2005), переход от теплой МИС 7 к холодной МИС 6 был достаточно долгим и постепенным. Он многократно осложнялся кратковременными потеплениями, что и фиксирует состав фауны мелких позвоночных различных уровней слоя 12.3.
Из слоя 12.2 получено 649 определимых образцов. Кости светло-коричневого цвета, хорошей сохранности, не имеют следов омарганцевания и осаждения карбонатов. Здесь возрастает количество летучих мышей Chiroptera до 4% и костей рыб до 24.6% (рис. 4 ). Немного увеличилась доля суслика Spermophilus – 0.92%, а численность сурка снижается до 0.1%. Происходит снижение численности Clethrionomys до 5.1%, скальных полевок Alticola до 15.1% и серых полевок рода Microtini до 21.9%. Возрастает количество степных пеструшек Lagurus, достигая 0.8%. Численность крота поднимается до 2.5%, а хомячка Эверсманна – до 0.92%, т.е., увеличивается более чем в два раза. Опять появляется слепушонка Ellobius sp. – 0.15%. Соотношение видов внутри группы Microtini меняется: Stenocranius gregalis составляет 6.01%, Microtus oeconomus – 0.8%, M. hyperboreus – 1.8%. Немного снижается численность цокора: до 2%. Пищухи составляют 0.8% тафоценоза. Численность лягушек снижается до 1.5%. Хищники представлены зубами мелкого куньего Mustela sp. – 0.1% и молочными зубами медвежат – 0.5%.
Приведенные цифры указывают на сокращение площади лесных биотопов и, следовательно, на холодные и достаточно сухие природные условия в период накопления осадков слоя 12.2. Вместе с тем, для этого слоя характерно увеличение количества крота и, почти в два раза – количества землероек (рис. 4 ). Численность цокора снижается незначительно, что указывает на отсутствие длительного и глубокого промерзания почвы. Эти показатели предполагают одновременное сокращение площади лесных массивов и ксерофильных степных биотопов. Расширились площади лугового разнотравья, что в целом отражает прохладную климатическую обстановку. Все это свидетельствует о чередовании холодных и теплых климатических фаз в период накопления слоя 12.2. На сложное и многоступенчатое преобразование природной обстановки для верхней части слоя 12.3 и нижней части слоя 12.2 указывают и данные спорово-пыльцевого анализа (Болиховская и др., 2017). Признаки дальнейшего похолодания и аридизации климата в этом слое хорошо выражены, что позволяет относить его ко второй половине тазовской холодной эпохи, к концу МИС 6.
Из слоя 12.1 определено 780 костей и зубов мелких позвоночных. Они светло-коричневого цвета, хорошей сохранности, не имеют следов осаждения марганца и карбонатов. Для этого тафоценоза характерно увеличение количества летучих мышей до 9.5% и снижение количества костей рыб до 14% (рис. 4 ). Численность рыжих полевок Clethrionomys на прежнем уровне – 5%, а количество Alticola увеличивается, достигая 19.35%. Возрастает таксономическое разнообразие Microtini. В слое 12.1 становится больше землероек Soricidae – 1.1%, установлено присутствие Neomys – 0.1%. Меньше становится пищух Ochotona – 0.4%. Снижается численность крота Asioscalops до 1.1% и цокора до 1.3%. В тафоценозе этого слоя снижается численность суслика до 0.5%, сурок вообще не обнаружен. В небольшом количестве зарегистрирован лемминг Lemmus – 0.1%. Найдена слепушонка Ellobius – 0.1%, присутствуют кости ящериц – 0.1% и лягушек – 1.5%. Изредка встречаются остатки мелких куньих рода Mustela – 0.26% и молочные зубы медвежат – 0.26%.
Приведенные цифры указывают на сокращение степных участков и увеличение доли скальных биотопов, поросших кустарничками, расширение площади лугового разнотравья. Это отражает прохладную климатическую обстановку, которая нарушалась потеплением в эпоху формирования осадков уровней 10 и 9, о чем свидетельствует повышенное содержание рукокрылых, повышенная численность рыжих полевок и лягушек. Начало формирования слоя 12.1 характеризуется потеплением и увлажнением климата, а затем произошло небольшое похолодание и увеличение сухости. Однако существенного изменения температурного режима не происходило. Вероятно, слой 12.1 соответствует переходному этапу от тазовского холодного времени к казанцевскому межледниковью.
Из слоя 11.4 получено 757 определимых костных остатков мелких позвоночных. Они светло-коричневого цвета, хорошей сохранности, не имеют следов омарганцевания и осаждения карбонатов. Доля летучих мышей в составе тафоценоза снижается и равна 5.8%, доля костей рыб уменьшается до 8.6%. Участие рыжих полевок Clethrionomys увеличивается до 7.3%, численность скальных полевок Alticola снижается до 14.4% (рис. 4 ). При этом количество Clethrionomys в нижних горизонтах слоях 11.4 приблизительно вдвое больше, чем в верхних горизонтах. Серые полевки Microtini достигают здесь максимального количества и таксономического разнообразия. В слое 11.4 появляются степные пеструшки Lagurus – 0.1%, зарегистрирован лемминг Lemmus sp. – 0.5%. (причем четыре моляра этого лемминга найдены в верхней части слоя 11.4). Присутствует водяная полевка Arvicola cf. sapidus – 0.1%. Примечательны некоторое увеличение численности землероек до 1.3% и сокращение в два раза численности крота – 0.5%. Среди землероек присутствуют Sorex minutus – 0.1%, Sorex araneus – 0.1%, водная землеройка Neomys – 0.1%. Большая часть землероек, восемь экз., найдена в нижней части слоя, а из верхних уровней получен только один образец. Наземные беличьи представлены лишь сусликом – 0.9%. Среди хомячков на долю барабинского приходится 0.1%, хомячка Эверсманна – 0.3%. Очень важной является находка моляра летяги Pteromys volans в нижней части слоя, что составляет 0.1%. Немного увеличивается количество цокора до 1.4%. Численность лягушек возрастает до 0.4%. Среди хищников присутствуют Mustela – 0.3% и Martes – 0.1%. Найдены два молочных зуба медведей.
Cостав тафоценоза позволяет сделать вполне определенные выводы. Нижние горизонты слоя 11.4 (рис. 4 ) накапливались в условиях лесных сообществ. Об этом свидетельствуют высокая численность рыжих полевок и присутствие летяги – типичного обитателя древесных ассоциаций. Даже отдельные находки летяг, как, например, в отложениях уровня 6, есть свидетельство их постоянного присутствия в исчезнувших биоценозах и, таким образом, существования высокоствольных лесных ассоциаций. Значительная концентрация землероек Soricidae именно в нижних горизонтах слоя 11.4 (уровни 8–6) указывает на обилие почвенных беспозвоночных летом и семян хвойных деревьев на поверхности почвы зимой. Все вместе это позволяет предполагать климатический режим, характерный для межледниковья.
Фауна верхних уровней (5–3) слоя 11.4 свидетельствует об изменении биотопической обстановки, о сокращении площади лесов и расширении открытых биотопов. Она маркирует начало очередного холодного этапа развития природной среды, что подтверждают данные спорово-пыльцевого анализа (Болиховская и др., 2016). В целом уровни 8–6 соответствуют теплой климатической фазе казанцевского межледниковья МИС 5e. Верхние уровни 5–3 отражают постепенное похолодание в преддверии ермаковского оледенения.
Остеологическая коллекция из слоя 11.3 насчитывает 704 экз. Кости светло-коричневого цвета, хорошей сохранности. Участие летучих мышей Chiroptera в тафоценозе равно 7.95%, что несколько выше, чем в слое 11.4. Количество костей рыб увеличивается вдвое, составляя 17.7% (рис. 4 ). В слое нарастает численность землероек, достигая 1.99%. Более чем в два раза увеличивается численность крота – 1.28%. Доля полевок Clethrionomys сокращается до 2.3%, а полевок Alticola уменьшается до 13.6%. Количество степных пеструшек Lagurus увеличивается до 0.28%. Возрастает численность суслика до 1.56%. Общее количество серых полевок рода Microtus остается на прежнем уровне – 34.1%. Почти в два раза увеличивается количество пищух Ochotona, до 0.57% и цокора Myospalax myospalax, до 2.8%. Заметно увеличивается численность хомячка Эверсмана – до 0.57%, а количество слепушонки Ellobius, напротив, снижается до 0.14%. На долю костей птиц приходится 7.39%, лягушек – 0.43%.
Состав тафоценоза слоя 11.3 (рис. 4 ) говорит о некотором повышении зимних температур и, возможно, об увеличении количества зимних осадков. Происходит сокращение лесных массивов и сохранение прежних размеров открытых каменистых склонов. Анализ распределения костных остатков по уровням свидетельствует о том, что в нижней части слоя (уровни 5–6) на квадрат раскопа приходится 11 моляров Clethrionomys, а в верхней части (уровни 1–4) – только 5 моляров, т.е., вдвое меньше. Количество моляров Alticolа на квадрат раскопа в нижней части слоя (уровни 6–5) приходится 57 моляров, а в верхней части слоя (уровни 4–1) – 39 моляров, т.е., приблизительно на одну треть меньше. Такие изменения в количественном соотношении этих групп указывают на похолодание климата в процессе накопления слоя 11.3. Эту тенденцию подтверждает увеличение количества сусликов от нижней части осадков к верхней. Вместе с тем, от нижних уровней слоя 11.3 к верхним увеличивается количество крота, полевки экономки и костей рыб. Это свидетельствует об увеличении влагообеспеченности почвенного покрова, о возросшей мощности водотоков в окрестностях пещеры. Такое заключение совпадает с данными пыльцевого анализа, который регистрирует появление в спектрах пыльцы прибрежных и водных растений (Болиховская и др., 2016).
Тафоценоз слоя 11.3 свидетельствует об изменениях природной среды, которые отмечены уже в кровле слоя 11.4. Осадки слоя 11.3, по-видимому, отвечают разным этапам ермаковского оледенения в период МИС 4.
Из слоя 11.2 определено 1350 экз. мелких позвоночных. Кости светло-коричневого цвета, хорошей сохранности, не имеют следов осаждения марганца и карбонатов. Для этого тафоценоза характерно значительное уменьшение количества летучих мышей до 0.3% и увеличение количества костей рыб до 42.9% (рис. 5 ). Такое количество рыб максимально для всего разреза отложений восточной галереи Денисовой пещеры. Насекомоядные Insectivora встречаются спорадически. Численность землероек снижается до 0.3%. Численность крота Asioscalops возрастает до 2.15%. Снижается численность полевок Clethrionomys, до 0.07%. Численность скальных полевок Alticola, напротив, увеличивается до 19.3% (рис. 5 ). Количество степных пеструшек Lagurus небольшое – 0.07%. Снижается общее количество серых полевок Microtini до 18.9. Однако видовое разнообразие этой группы остается высоким. На прежнем уровне остается численность суслика – 1.41%, зарегистрирован сурок – 0.07%. Возрастает количество хомячка Эверсманна до 2.9% и слепушонки Ellobius до 0.3%. Численность цокора снижается до 1.77%, а пищух равна 0.44%. Среди них присутствует степная пищуха Ochotona pusilla – 0.07%. В слое 11.2 участие костей лягушек равно 0.15%, мелких хищников рода Mustela – 0.07%.
Рис. 5.
Количественное соотношение основных экологических групп мелких позвоночных из слоев 11.2, 11.1 и 9 восточной галереи Денисовой пещеры в %.

Приведенный состав тафоценоза (рис. 5 ) свидетельствует о распространении открытых ландшафтов, занятых степными и луговыми ассоциациями, включая участки с разреженным травостоем. Судя по всему, лесные массивы в это время уменьшились по сравнению с эпохой накопления слоя 11.3. Вероятно, они сохранялись лишь в поймах рек. Важное место в ландшафтах долины Ануя занимали луговые ассоциации. Распределение костных остатков по уровням в пределах этого слоя показывает, что в нижней его части (уровни 3 и 2) количество остатков крота на квадрат раскопа – 25 экз., а в верхней части (уровень 1) – 4 экз. Количество полевки-экономки на квадрат раскопа в нижней части слоя (уровни 3 и 2) – 3 экз., а в верхней (уровень 1) они не найдены. Водяная полевка обнаружена только в нижней части слоя – 5 экз. Цокор в нижней части слоя (уровни 3 и 2) представлен 20 экз., в верхней части слоя – 4 экз. Степная пеструшка и степная пищуха найдены только в верхней части слоя (уровень 1). Все эти данные отражают аридизацию природной обстановки на протяжении накопления слоя 11.2.
Слой 11.2, вероятно, отражает переходный этап от ермаковского к каргинскому времени, который характеризуется холодными флюктуациями, что хорошо выражено на кривой изменения изотопного состава кислорода в океанических отложениях (Lisiecki, Raymo, 2005). График в указанной работе (ibid., рис. 4) показывает, что потепление МИС 3 было почти в три раза слабее, чем потепление в период МИС 5 казанцевского межледниковья и современного, т.е. голоценового (МИС 1), потепления. Этот вывод согласуется с данными по растительности и по составу мелких позвоночных из слоя 11.2. Они свидетельствуют о том, что лесные массивы занимали небольшие площади (Болиховская и др., 2017).
Слой 11.1 охарактеризован 821 экз. костей мелких позвоночных. В тафоценозе этого слоя количество летучих мышей Chiroptera увеличивается на одну треть и достигает 0.49%. Количество костей рыб, напротив, снижается до 26.9%. Увеличивается по сравнению с предыдущим слоем численность полевок Clethrionomys до 1.1%. Количество Alticola снижается до 17.9% (рис. 5 ). Существенно нарастает численность Lagurus – 3.17%. Найден один моляр лемминга Lemmus, что составляет 0.1% состава тафоценоза. Заметно увеличивается численность серых полевок, до 27.2%. Среди них Stenocranius gregalis составляет 1.1%, Microtus oeconomus – 0.2%, M. hyperboreus – 4.9%, M. arvalis – 0.24%. Arvicola составляют 0.4%. Численность Myospalax myospalax поднимается до 0.5%. Увеличивается видовое разнообразие хомяков. В их составе: Cricetus criсetus – 0.24%, Cricetulus barabensis – 0.7%, Allocricetulus eversmanni – 2.6%. Присутствует Ellobius – 0.24%. Численность Ochotona остается на прежнем уровне – 1.1, количество лягушек – 0.5%.
Для слоя 11.1 характерно нарастание видового разнообразия мелких позвоночных, увеличение численности землероек и крота, рыжих полевок и цокора, что можно интерпретировать как развитие каргинского межледниковья в период МИС 3. Фактически, каргинский горизонт стратиграфической схемы является лишь небольшим интерстадиалом, который нарушил общий процесс похолодания климата от казанцевской межледниковой эпохи в период МИС 5 до максимума сартанской ледниковой эпохи периода МИС 2, что и отражают данные по пыльце и мелким позвоночным из отложений слоя 11.1.
Определимые костные остатки из слоя 9 насчитывают 14 экз. (рис. 5 ). Количество полученного материала недостаточно для достоверных выводов. На основе спорово-пыльцевых спектров для времени формирования толщи слоя 9 реконструированы три фазы развития растительности и климата сартанского позднеледнековья – от суровых криоаридных условий до распространения перигляциальных межстадиальных лесостепей (Болиховская и др., 2017).
ЗАКЛЮЧЕНИЕ
Состав мелких позвоночных из отложений восточной галереи Денисовой пещеры свидетельствует о том, что на протяжении всей истории осадконакопления в окрестностях пещеры сохранялась высокая мозаичность биотопов. Лесные массивы чередовались с участками луговой и степной растительности. Небольшими пятнами были вкраплены ассоциации сухих степей и, возможно, тундр. Формирование каждого слоя протекало в условиях сложной динамики природных комплексов, обусловленной колебаниями климата. Полученные результаты позволяют сопоставить эту динамику с изотопной кривой кислорода δ18O океанических осадков (Lisiecki, Raymo, 2005), которая отражает общепланетарные климатические изменения (рис. 6). Они хорошо согласуются между собой.
Рис. 6.
Сопоставление времени накопления отложений в восточной галерее Денисовой пещеры с колебаниями изотопной кривой δ18 O океанических осадков. Датировки слоев восточной галереи Денисовой пещеры даны по работам К. Доука и др. (Douka et al., 2019) и З. Якобс и др. (Jacobs et al., 2019). Кривая колебаний изотопной кривой δ18O океанических осадков из работы Л. Лисиецки и М. Раймо (Lisiecki, Raymo, 2005).

Увеличение численности летучих мышей в слое 9, резкое сокращение костей рыб и уменьшение количества артефактов указывают на ослабление антропогенного воздействия в процессе формирования пещерного тафоценоза в этот период.
Такое заключение подтверждается изучением фосфатной минерализация отложений восточной галереи Денисовой пещеры. Показано, что в верхней части слоя 9 и в слоях 8–6 присутствуют аутигенные фосфаты, образование которых связано с процессами биодеградации фосфорсодержащих органических веществ, главным образом, гуано летучих мышей (Шуньков и др., 2018). Более того, в черных горизонтах слоев 6–8 и, отчасти, 9 сохранились многочисленные фрагменты хитина насекомых, принесенные летучими мышами. Все это свидетельствует о минимальной активности древних обитателей Денисовой пещеры и о временном перерыве обитания здесь человека в конце плейстоцена и в начале голоцена.
Работа выполнена при поддержке грантов РФФИ 18-09-40100, РНФ № 21-18-00089 и Программы фундаментальных исследований Президиума РАН “Эволюция органического мира. Роль и влияние планетарных процессов”.
Список литературы
Агаджанян A.K. Мелкие млекопитающие среднего плиоцена Русской равнины. Стратигр. Геол. корреляция. 2003. Т. 11. № 6. С. 98–114.
Болиховская Н.С., Козликин М.Б., Шуньков М.В. Климатостратиграфия верхней части плейстоценовой толщи в восточной галерее Денисовой пещеры и смены древних природных обстановок на Северо-Западе Алтая (по данным палинологического анализа) // Пути эволюционной географии: Матер. Всеросс. научн. конф., посвященной памяти проф. А.А. Величко (Москва, 23–25 ноября 2016 г.). М.: Ин-т географии РАН, 2016. С. 631–635.
Болиховская Н.С., Козликин М.Б., Шуньков М.В. и др. Новые данные в палинологии уникального памятника палеолита Денисова пещера на Северо-Западе Алтая // Бюл. Моск. об-ва испыт. прир. Отд. биол. 2017. Т. 122. Вып. 4. С. 46–60.
Деревянко А.П., Молодин В.И. Денисова пещера. Ч. 1. Новосибирск: Наука, 1994. 262 с.
Деревянко А.П., Шуньков М.В., Агаджанян А.К. и др. Природная среда и человек в палеолите Горного Алтая. Новосибирск: Изд-во Ин-та археол. и этногр. СО РАН, 2003. 445 с.
Деревянко А.П., Шуньков М.В., Маркин С.В. Динамика палеолитических индустрий в Африке и Евразии в позднем плейстоцене и проблема формирования Homo sapiens. Новосибирск: Изд-во Ин-та археол. и этногр. СО РАН, 2014. 226 с.
Шуньков М.В., Кулик Н.А., Козликин М.Б. и др. Фосфатная минерализация плейстоцен-голоценовых отложений восточной галереи Денисовой пещеры // Докл. Акад. наук. Геол. 2018. Т. 478. № 3. С. 318–322.
Agadzhanyan A.K. Eine altertümliche Arvicola (Rodentia, Mammalia) aus dem Mittelpleistozän der Russischen Ebene // Schr. Geol. Wiss. 1983. H. 19(20). S. 9–29.
Brown S., Buckley M., Derevianko A. et al. Identification of faunal remains from Denisova Cave through the Application of ZooMS // 5th Annual Meeting London, UK, 10–12 September, 2015. P. 60 (Proc. European Soc. for the study of Human Evolution 4).
Douka K., Slon V., Jacobs Z. et al. Age estimates for hominin fossils and the onset of the Upper Palaeolithic at Denisova Cave // Nature. 2019. V. 565. P. 640–644.
Heinrich W.-D. Zur Evolution und Biostratigraphie von Arvicola (Rodentia, Mammalia) im Pleistozän Europas // Z. Geol. Wiss. 1982. Bd 10. S. 693–735.
Heinrich W.-D. Neue Ergebnisse zur Evolution und Biostratigraphie von Arvicola (Rodentia, Mammalia) im Quartär Europas // Z. Geol. Wiss. 1987. Bd 15. S. 389–406.
Heinrich W.-D. Some aspects of evolution and biostratigraphy of Arvicola (Mammalia, Rodentia) in the Central European Pleistocene // Intern. Symp. Evol. Phylog. Biostr. Arvicolids. Praha: Geol. Survey, 1990. P. 165–182.
Jacobs Z., Li B., Shunkov M.V. et al. Timing of archaic hominin occupation of Denisova Cave in southern Siberia // Nature. 2019. V. 565. P. 594–599.
Koenigswald W.V. Arvicolidae – Wühlmause. Stammesgeschichte und Schmelzmuster // Handbuch der Säugetiere Europas. 1982. Bd 2. Pt 1. S. 60–69.
Lisiecki L.E., Raymo M.E. A Plio-Pleistocene stack of 57 globally distributed benthic δ18O records // Paleoceanogr. and Paleoclimatol. 2005. V. 20. P. 1–16. https://doi.org/10.1029/2004PA001071
Morley M.W., Goldberg P., Uliyanov V.A. et al. Hominin and animal activities in the microstratigraphic record from Denisova Cave (Altai Mountains, Russia) // Sci. Reports. 2019. V. 9. Article number: 13785. P. 1–12. https://doi.org/10.1038/s41598-019-49930-3
Prüfer K., Racimo F., Patterson N. et al. The complete genome sequence of a Neanderthal from the Altai Mountains // Nature. 2014. V. 505. P. 43–54.
Sawyera S., Renauda G., Viola B. et al. Nuclear and mitochondrial DNA sequences from two Denisovan individuals // PNAS. 2015. V. 112. № 51. P. 15697–15700. www.pnas.org/cgi/doi/10.1073/pnas.1519905112.
Slon V., Hopfe C., Weiß C.L. et al. Neandertal and Denisovan DNA from Pleistocene sediments // Science. 2017a. V. 356. P. 605–608. doi 10.1126/science.aam9695 [PubMed: 28450384]
Slon V., Viola B., Renaud G. et al. A fourth Denisovan individual // Science Advances. 2017b. V. 3. P. 1–8. e1700186. http://advances.sciencemag.org/.
Slon V., Mafessoni F., Vernot B. et al. The genome of the offspring of a Neandertal mother and a Denisovan father // Nature. 2018 Sept.; V. 561(7721). P. 1–18. https://doi.org/10.1038/s41586-018-0455-x
Viola B.T., Markin S.V., Buzhilova A.P. et al. New Neanderthal remains from Chagyrskaya Cave (Altai Mountains, Russian Federation) // Amer. J. Phys. Anthropol. 2012. V. 147. Suppl. № 54. P. 293–294.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал