

Палеонтологический журнал, 2021, № 3, стр. 104-116
Железистые кварциты (джеспилиты) – взгляд палеонтолога
М. М. Астафьева
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
Поступила в редакцию 10.09.2020
После доработки 07.10.2020
Принята к публикации 07.10.2020
Аннотация
Результаты изучения ископаемых бактерий показывают, что минералообразование под влиянием бактерий или с их участием – процесс, который происходил на Земле всегда и везде (после появления бактерий). Однотипность пород и породообразования докембрия и девона говорят о схожести условий, существовавших на Земле в те времена – в архее, протерозое и девоне – во время образования изученных пород.
ВВЕДЕНИЕ
Велика роль бактерий в образовании различных минералов железа. Но, пожалуй, наибольший интерес вызывает проблема появления полосчатых железистых кварцитов в связи с масштабами накопления железа в палеобассейнах раннего докембрия и несомненной связи формирования полосчатых железистых кварцитов с определенным этапом геологической истории Земли. Преобладающая часть железистых кварцитов отлагалась ранее рубежа 2.0 млрд лет.
Полосчатые железистые кварциты представляют собой метаморфизованные осадочные или вулканогенно-осадочные кварцево-железистые горные породы, широко распространенные в докембрийских образованиях. Характерной их чертой является чередование железистых (магнетит, гематит, сидерит) и кремнистых/карбонатных (кварц, яшма, доломит и анкерит) прослоев, образующих слои мощностью от нескольких микрон до нескольких метров (Posth et al., 2011). Тонко-полосчатые, яшмовидно-тонкозернистые разновидности железистых кварцитов называют джеспилитами. Содержание железа часто превышает 15%, и эти породы заключают в себе основную массу мировых запасов железа. В настоящее время большинство исследователей подразделяют все железистые кварциты на две главные группы: месторождения типа Алгома – небольшие месторождения, связанные с вулканитами, и месторождения типа оз. Верхнего – крупных размеров, представленные ассоциациями, образовавшимися чисто осадочным путем в мелководных обстановках седиментационных бассейнов (Холодов, 1993; Posth et al., 2011).
Полосчатые железистые кварциты известны на всех материках, появляясь в архее. Главные месторождения железистых кварцитов типа Алгома приурочены к архею, но известны и в протерозое. В Гренландии железистые кварциты района Исуа – одни из самых древних осадочных горных пород Земли (3.76 млрд лет) (Schopf, 1983; Posth et al., 2011). Наибольшего развития железистые кварциты достигают в протерозое, они являются составной частью метаморфических комплексов всех древних щитов. Максимум образования железистых кварцитов типа оз. Верхнее (Superior) на всех континентах приходится более или менее на одно и то же время в начале протерозоя (2.0–2.5 млрд лет тому назад). Полосчатые железистые кварциты этого типа простираются на сотни километров, достигая мощности сотен метров.
Своеобразие развития во времени, тонкая слоистость, ритмическая повторяемость прослоев вызывают большой интерес к выяснению генезиса этой формации.
Обычно считается, что все минералы железистых кварцитов не являются первичными, а несут следы как диагенетических, так и метаморфических изменений. Иногда предполагается, что первичными минералами железа железистых кварцитов были гидроокись железа, гриналит и сидерит (Klein, 2005; Posth et al., 2011).
Мысль о возможном биогенном происхождении раннедокембрийских джеспилитов возникла в начале XX в. Пионером был Ч. Лейс, одним из первых предположивший участие бактерий в образовании джеспилитов с возрастом 1.6–2.8 млрд лет. Эту идею подхватили Л. Кайе, Е. Муром, П. Клауд и др. (Cloud, 1968; Антошкина, 2011 и литература там). В 60–70-е гг. ХХ в. гипотезу биогенного происхождения раннедокембрийских полосчатых железистых кварцитов рассматривал Г. Лаберж (LaBerge, 1967, 1973) на основе результатов микропалеонтологических исследований десятков месторождений из различных регионов.
Потенциально все минералы железистых прослоев могут иметь биогенную природу, поскольку и для окисленных (гематит), и для восстановленных (магнетит и сидерит) форм микробиологами установлен прямой механизм бактериальной продукции. При этом не следует забывать, что участие микроорганизмов в образовании железных руд проявляется в их способности концентрировать элементы из среды и преодолевать кинетические барьеры, являясь, таким образом, катализатором химического процесса. Целый ряд вопросов, связанных с происхождением железистых кварцитов, таких как источник железа, пути его транспортировки, условия осаждения и т.д., остаются открытыми.
Вероятное участие диссимиляционных железоредукторов в формировании докембрийских месторождений железистых кварцитов исследовали Д.Г. Заварзина (2004а, б) и А.И. Слободкин (2008). Из-за деятельности железобактерий процесс окисления происходит очень быстро, вследствие этого возможно появление только водного окисла железа с наименее совершенной структурой – ферригидрита. Ферригидрит способен самопроизвольно переходить в гематит, что приводит к быстрому уплотнению осадков, при этом характерные бактериальные структуры утрачиваются (Заварзина, 2004а).
Мало того, выдвигалась гипотеза о том, что диссимиляционное восстановление Fe(III) могло быть первым возникшим типом метаболизма. Высказывались осторожные предположения, что нельзя полностью исключить возможность биогенного происхождения даже самых древних – эоархейских железистых кварцитов с возрастом >3.8 млрд лет (Dodd, Papineau, 2015). А.Г. Вологдин (1976) писал, что в рудах Кривого Рога и в аналогичных “кремнистых рудах” (т.е. в полосчатых железистых кварцитах) Кольского п-ова наблюдались “настоящие бактериальные структуры, лучше различимые именно в рудах, бедных железом, на фоне светлого кремнезема”. Тот же автор упоминал о присутствии остаточного органического, по-видимому, бактериального вещества (т.е., углерода) в этих рудах, что соответствует данным наших анализов. Однако изображения микрофоссилий, обнаруженных в нижнем протерозое криворожской серии, названных Вологдиным (1976) кораллитами, приведены без указания масштаба, и качество изображения оставляет желать лучшего. Еще одно изображение – фотография, сделанная С.И. Жмуром в “Бактериальной палеонтологии” (2002, с. 108, рис. 1, а).
Рис. 1.
Мелкие округло-овальные формы (<1 мкм) из железистых прослоев железистых кварцитов и химический анализ. Изображение на электронном микроскопе Zeiss № 3195 от 19.11.2012. Архей (>2.7 млрд лет), Оленегорская структура, Мурманская обл., Кольский п-ов.

Зеленокаменные пояса11 архея содержат набор осадочных и вулканических пород, который, как пишут О. Розен с соавт. (2006): “не обнаруживает заметных систематических отличий от соответствующих фанерозойских аналогов”. В этом наборе пород присутствуют все их типы, включая карбонаты, эвапориты и др. Таким образом, с точки зрения сравнения породных ассоциаций, нет оснований говорить о существенной разнице в условиях атмосферы и гидросферы неоархея и более поздних времен.
Отличительной чертой древних толщ является достаточно широкое присутствие железистых кварцитов – джеспилитов. Однако следует помнить об их присутствии не только в верхнем рифее Канады, Бразилии и Намибии (Windley, 1999), но и в венде–нижнем кембрии Восточного Саяна (Кизир-Казырская складчатая зона) и даже в девоне Казахстана – джеспилоиды Жайремского месторождения.
Нами были изучены джеспилиты и джеспилоиды как архея и протерозоя, так и девона, и во всех случаях подтверждалась бактериальная природа этих пород. Значит, присутствуют общие характеристики бассейнов осадконакопления. К числу таких принципиальных характеристик относится и аэрированность бассейнов, т.е., содержание свободного кислорода, и температура палеобассейнов. Таким образом, изучение железистых кварцитов может пролить свет и на условия жизни на ранней земле.
МАТЕРИАЛ
К числу изученных и изучаемых нами железорудных бассейнов относятся: железорудное проявление Полмос-Порос и Оленегорское месторождение Кольского п-ова, Костомукшское и Межозерное месторождения Карелии – тип Алгома; Читрадурга Южной Индии (AR); Старооскольское месторождение Курской магнитной аномалии (PZ1); Жайремское месторождение Казахстана (D3fm) – тип оз. Верхнее.
МЕТОДИКА ИССЛЕДОВАНИЙ
Поскольку проблема происхождения и условий образования железистых кварцитов представляет значительный интерес, для ее решения были привлечены микропалеонтологические, в частности, бактериально-палеонтологические методы исследования. Все исследования проводились на электронном сканирующем микроскопе CamScan-4 с микроанализатором Link-860, на сканирующем электронном микроскопе системы Zeiss EVO 50 с рентгеновским микроанализатором Inca Oxford (350); TESCAN VEGA II ZMU фирмы “TESCAN” с системой рентгеновского энергодисперсионного микроанализа INCA ENERGY 450 и TESCAN VEGA 3 фирмы “TESCAN”. Исследовались только свежие сколы образцов, напыление – золотом.
БАКТЕРИАЛЬНАЯ ПАЛЕОНТОЛОГИЯ ПОЗДНЕАРХЕЙСКИХ ЖЕЛЕЗИСТЫХ КВАРЦИТОВ (ТИП АЛГОМА)
Во всех изученных образцах позднеархейских железистых кварцитов обнаружены инситные микрофоссилии, как в железистых, так и в кремнистых прослоях. В железистых прослоях преобладают коккоидные, нитевидные, гантелевидные формы (рис. 1) часто очень малых размеров (<1 мкм) (рис. 2).
Рис. 2.
Мелкие округло-овальные формы (<1 мкм) из железистых прослоев железистых кварцитов и химический анализ. Изображение на электронном микроскопе Zeiss № 3200 от 19.11.2012. Архей (>2.7 млрд лет), Оленегорская структура, Мурманская обл., Кольский п-ов.

Интересно, что некоторые кокки несут следы деления – вполне вероятно, нам удалось наблюдать следы деления клеток (рис. 3). Подчеркиваем, что часто мы имеем дело с предполагаемыми организмами наноразмеров – нанобактериями, которые мало изучены.
Рис. 3.
Мелкие округло-овальные формы (<1 мкм) из кремнистых прослоев железистых кварцитов (на некоторых кокках, погруженных в породу, видны следы деления клеток), и химический анализ. Изображение на электронном микроскопе Zeiss № 4274 от 24.06.2014. Архей (2.7 млрд лет), Костомукшинская структура, Зап. Карелия.

Не следует думать, что микрофоссилии архейских железистых кварцитов ограничиваются наноформами. Встречены и обычные микрофоссилии микронных размеров. Это кокки, редкие нити (рис. 4). Изредка встречаются удлиненно-овальные или палочковидные формы (рис. 5). Длина их 2–3 мкм, ширина 1.0–1.5 мкм. Они часто группируются, образуя некое подобие нитевидных структур. Широко распространены образования, которые рассматриваются нами, как биопленки (рис. 6).
Рис. 4.
Нити со строением, напоминающим соединение клеток; они утоплены в породу, что дает возможность предположить их прижизненное захоронение. Изображение на электронном микроскопе Zeiss № 3183 от 19.11.2012. Архей (>2.7 млрд лет), Оленегорская структура, Мурманская обл., Кольский п-ов.

Рис. 5.
Удлиненно-овальные или палочковидные формы. Длина их 2–3 мкм, ширина 1.0–1.5 мкм; они часто группируются, образуя некое подобие нитевидных структур. Архей (>2.7 млрд лет), Оленегорская структура, Мурманская обл., Кольский п-ов.

Рис. 6.
Биопленка и ее химический анализ. Видны содержащиеся в биопленке коккоидные формы. Изображение на электронном микроскопе Zeiss № 4223 от 27.05.2014. Архей (2.7 млрд лет), Костомукшинская структура, Зап. Карелия.

Более крупные нити (которые могли бы оказаться цианобактериями) почти всегда лежат на поверхности породы или же заполняют трещины, поэтому мы склонны рассматривать их как более позднюю контаминацию.
Безусловно, часть обнаруженных микрофоссилий может являться засорением; прежде всего, это относится к крупным коккам и некоторым нитевидным формам. Но захоронение мелких коккоидов одновременно с осадконакоплением весьма вероятно. Это доказывают углубления в породе от выпавших кокков.
Представленные результаты свидетельствуют о разнообразии микробиальных форм в железистых кварцитах позднего архея – в крупнейших месторождениях Сев. Карелии и Кольского п-ова. Часть обнаруженных микрофоссилий может являться остатками магнетотактических и железоредуцирующих бактерий. Поскольку большинство подобных бактерий являются микроаэрофилами и развиваются на содержащем Fe3+ субстрате, правомерно предположить наличие свободного кислорода в зоне гипергенеза, как минимум, начиная с 2.7 млрд лет назад, причем содержание кислорода было достаточным для окисления железа. Транспортировка железа в бассейны формирования железистых кварцитов происходила в форме Fe3+, часть которого могла восстанавливаться посредством бактериальной редукции в среде с высоким содержанием органического вещества с образованием магнетита до стадий позднего диагенеза и метаморфизма. Возможно, некоторое количество магнетита образовалось на стадии транспортировки железа в континентальных обстановках в результате бактериальной редукции. Таким образом, участие бактериальной компоненты в образовании железистых кварцитов архея очень вероятно. Скорее всего, архейские моря были тепловодными, относительно обогащенными кислородом бассейнами. Другими словами, можно предположить, что архейские железистые кварциты являются одними из самых древних биогенных пород.
БАКТЕРИАЛЬНАЯ ПАЛЕОНТОЛОГИЯ РАННЕПРОТЕРОЗОЙСКИХ ЖЕЛЕЗИСТЫХ КВАРЦИТОВ (ТИП ОЗ. ВЕРХНЕЕ)
Месторождения Курской магнитной аномалии (КМА) относятся ко второй группе осадочных железистых кварцитов – железистых кварцитов типа оз. Верхнего (Superior). Мы изучали образцы массивных полосчатых железистых кварцитов Старо-Оскольского железорудного района КМА. Образцы получены в Центральном научно-исследовательском геолого-разведочном музее им. акад. Ф.Н. Чернышева, С.-Петербург (ЦНИГР музей).
Микрофоссилии позднепротерозойских железистых кварцитов отличаются от таковых архея. Характерной особенностью исследуемых пород явилось обилие фоссилизированного гликокаликса – внеклеточной полисахаридной слизи, выделяемой бактериями и цианобактериями, однородной или с включениями полисахаридных фибрилл. Гликокаликс образован переплетением полисахаридных волокон (декстраны и леваны). Основная его функция защитная, а также адгезия к различным субстратам; гликокаликс также является важным фактором при образовании колоний микроорганизмов и транспортировке питательных веществ. Мы также обнаружили остатки фоссилизированных бактерий или цианобактерий (Розанов и др., 2016). Судя по взаимоотношению с вмещающими породами, все микрофоссилии захоронены in situ. В исследуемом материале эти остатки были представлены в виде длинных (шириной до 2 мкм) переплетающихся расплющенных нитей (рис. 7), слагающих бактериальные и цианобактериальные маты, и в виде палочковидных форм размером 2–3 × 6–7 мкм.
Рис. 7.
Ископаемый бактериальный мат, состоящий из длинных (>20 мкм), взаимопроникающих, расплющенных нитей, шириной до 2 мкм. Центральная часть главной залежи Коробковского месторождения.

Но наибольший интерес представляла находка многочисленных трихомов, объединенных общим чехлом (рис. 8), напоминающая представителей современного рода цианобактерий Microcoleus (Schopf, 1983).
Рис. 8.
Многочисленные фоссилизированные, плавно изгибающиеся, длинные трихомы (диаметр около 2 мкм). В левой части снимка несколько трихомов (обозначены стрелками) объединены одним чехлом (белая полужирная стрелка). В нижней части снимка к этому пучку примыкает практически разрушенный пучок – видна оболочка (белая полужирная стрелка) и малочисленные трихомы (белая тонкая стрелка), заключенные в нее. Правее – следы (отверстия) от разрушенных трихомов (белая тонкая стрелка). Виден общий чехол (диаметром до 20 мкм), объединяющий все группы трихомов (черная стрелка). Подобная структура морфологически близка современным представителям цианобактерий рода Microcoleus. Лебединское месторождение, глуб. ~100 м.

В нашем случае несколько хорошо сохранившихся, плавно изгибающихся трихомов диаметром 1–2 мкм объединены в общий чехол. Диаметр этого пучка нитей был равен примерно 6 мкм. Несколько ниже (в левом нижнем углу снимка, рис. 7) находятся структуры, также напоминающие трихомы, объединенные общим чехлом. Возможно, что несколько таких пучков, в свою очередь, также заключены в общий чехол. Диаметр этого чехла приближается к 18–20 мкм. Также обращает на себя внимание наличие отверстий, похожих на следы от клеток, что характерно для цианобактерий.
Результаты исследования микрофоссилий свидетельствуют о том, что в образовании осадочных раннепротерозойских железистых кварцитов значительную роль играл биогенный фактор. Это было подтверждено нахождением в образцах ископаемых бактерий и предположительно цианобактерий, и обильного гликокаликса. Из этого следует, что осаждение железистых кварцитов осадочного типа происходило в условиях фотической зоны, т.е., на мелководье, и содержание О2 в атмосфере было довольно значительным.
В палеопротерозойских железистых кварцитах присутствуют и другие предположительно биогенные образования. В целом биогенная составляющая этих железистых кварцитов гораздо более представительна, чем архейская. Мало того, нам удалось сделать интересные наблюдения, иллюстрирующие преобразование некоторых минералов железистых кварцитов (Astafieva, Zaitseva, 2020). Приведены примеры, показывающие процесс преобразования железоокисляющими бактериями сидерита в окислы/гидроокислы железа (рис. 9).

БАКТЕРИАЛЬНАЯ ПАЛЕОНТОЛОГИЯ ПАЛЕОЗОЙСКИХ (ДЕВОНСКИХ) ДЖЕСПИЛОИДОВ
Исследуемый материал был получен из Жайремского месторождения полиметаллических руд в Карагандинской обл. Казахстана. Жайремский рудный узел находится в Центральном Казахстане вблизи пос. Жайрем. Здесь в пределах единой геологической структуры и на небольшом расстоянии друг от друга локализованы стратиформные залежи свинцово-цинковых, баритовых и железо-марганцевых руд среди осадочных отложений узкого временного интервала (D3fm). В этом отношении Жайремский узел представляется уникальным, а составляющий его комплекс месторождений уже давно выделяют в виде особого атасуйского типа.
Рудоносными являются отложения фаменского возраста. В распространении разных типов руд проявлен литолого-фациальный контроль. Пласты и линзы комплексных свинцово-цинковых и баритовых руд локализованы среди углеродистых глинисто-кремнисто-карбонатных отложений, галенит-баритовых – среди рифогенных известняков, а железо-марганцевых – преимущественно в кремнистых известняках. Во всех случаях железо-марганцевая минерализация располагается стратиграфически выше полиметаллической. Железные руды представлены мелкозернистыми тонкослоистыми породами с чередованием слоев, обогащенных кварцем или гематитом. Второстепенными минералами являются магнетит и кальцит (Брусницын и др., 2017).
Железные руды очень просты и однообразны по составу и строению. На всех месторождениях это мелкозернистые породы кирпично-красного цвета с хорошо выраженной слоистостью. Текстуры пород создаются чередованием слоев, обогащенных либо кварцем, либо гематитом. Участками, за счет процессов собирательной перекристаллизации, гематит образует относительно крупные (первые мм по удлинению) пластинчатые зерна, и тогда порода приобретает стально-серую до черной окраску. Второстепенными минералами являются магнетит и кальцит. По текстурам, структурам и минеральному составу железные руды Казахстана практически аналогичны классическим железистым кварцитам типа BIF (Banded Iron Formations) (Брусницын и др., 2018). Поэтому в отличие от классических джеспилитов железные руды Жайрема называют джеспилоидами.
Дискуссии о генезисе месторождений Жайремского рудного узла к началу 1980-х гг. были практически завершены. Исследователи сошлись во мнении о гидротермально-осадочном происхождении рудоносных залежей (Митряева, 1979; Скрипченко, 1980, 1989; Рожнов, 1982; Варенцов и др., 1993). Предполагалось, что рудоносные растворы, зарождавшиеся в вулканогенных породах фундамента рифтогенной впадины, выносили на поверхность морского дна Pb, Zn, Ba, Fe, Mn, Si и другие элементы. Здесь в различных фациальных и физико-химических обстановках происходила их дифференциация и накопление разных по составу металлоносных отложений. В стагнированных обстановках впадин образовывались Pb–Zn (±Ba) осадки, а на поднятиях в окислительных условиях – Pb–Ba и Fe–Mn (±Ba) (Брусницын и др., 2018).
Поскольку докембрийские джеспилиты и девонские джеспилоиды близко родственны, мы решили провести бактериально-палеонтологические исследования образцов, любезно предоставленных А.В. Ткачевым.
В образцах обнаружены ископаемые остатки не только прокариотных, но, скорее всего, и эвкариотных форм. Как ни странно, разнообразие микрофоссилий не увеличилось по сравнению с докембрийскими породами. Резко превалировали остатки чехлов биологических форм, скорее всего водорослевых или цианобактериальных (рис. 10), также присутствовали биопленки и единичные включения кокков и нитей (рис. 11, 12), а четко выраженные прокариоты и эвкариоты отсутствовали. Обилие биопленок при практическом отсутствии выраженных микрофоссилий не должно смущать. По современным представлениям, 95–99% микроорганизмов в природных местах обитания существует в биопленках (Николаев, Плакунов, 2007). Сама же идея того, что микроорганизмы в большинстве своем существуют в природе, главным образом, в виде структурированных сообществ, а не индивидуальных, свободно плавающих (“планктонных”) клеток, была сформулирована Дж. Костертоном еще в 1978 г. (Costerton et al., 1978). В ископаемое состояние они переходят в виде донных осадков или береговых штормовых выбросов, где особенно благоприятны условия захоронения. Но эти тафоценозы не отражают пространственной структуры сообщества в естественном состоянии (Заварзин, 1993).
Рис. 10.
Чехол цианобактерий или водорослей в джеспилоидах Жайремского месторождения. Изображение на электронном микроскопе Tescan Vega3 № Д-21-9 от 10.08.2020 г.

Рис. 11.
Нитевидные формы и чехлы цианобактерий или водорослей в джеспилоидах Жайремского месторождения. Изображение на электронном микроскопе Zeiss № 9972 от 28 ноября 2019 г. Тонкими стрелками указаны чехлы. Нитевидная изгибающаяся форма указана стрелкой средней толщины и жирной стрелкой – предполагаемый чехол бактериальной нитевидной формы. Нельзя исключить, что в нижней части нити в районе указателя стрелки наблюдаются следы деления нити на клетки.

Рис. 12.
Нитевидные формы в биопленках джеспилоидов Жайремского месторождения. Химические анализы. Изображение на электронном микроскопе Zeiss № 9974 от 28.11.2019 г. Стрелками указаны биопленки и бактериальные нити.

В настоящее время биологические микрообъекты воспринимаются как целостные, совместно эволюционирующие системы. Узкий взгляд на микроорганизмы как на просто одноклеточные формы жизни сменился осознанием их способности функционировать только в составе сообществ (Заварзин, 1993, 1995). И нет оснований предполагать, что в прошлом ситуация в корне отличалась от современной.
Поэтому присутствие чехлов биологических объектов и биопленок в жайремских джеспилоидах говорит о вероятном участии биологической компоненты в образовании этих пород. Ничего удивительного в этом нет, так как для джеспилитов и слагающих их минералов влияние биогенного фактора на их происхождение давно установлено (Астафьева и др., 2017 и литература там).
РЕЗУЛЬТАТЫ
Однотипность пород и породообразования говорит о схожести условий, существовавших на Земле в те времена – в архее, протерозое и девоне – во время образования изученных пород (рис. 13). Насыщенность девонской атмосферы кислородом сомнений не вызывает – соответственно, нелогично отрицать оксигенированность архейской атмосферы; то же можно сказать и о приповерхностной температуре Земли – архейская не может сильно отличаться от девонской, хотя долгое время, впрочем, как и сейчас, превалировало мнение о восстановительном характере архейско-раннепротерозойской атмосферы Земли и о невероятно высоких температурах ее поверхности в это время.
Рис. 13.
Обобщенная картина некоторых гео-биологических параметров в истории Земли (по: Розанов, 2009).
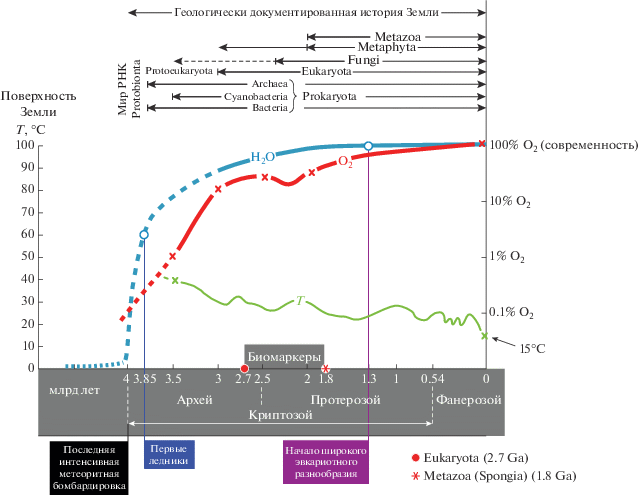
Сделанные нами выводы подтверждаются и данными по уровню организации ископаемых микроорганизмов, обнаруженных в архейско-протерозойских породах. Так, нахождение в архее и даже в раннем архее предположительных эвкариот (Белова, Ахмедов, 2006; Розанов, Астафьева, 2008, 2020; Buik, 2010; Ископаемые бактерии…, 2011) говорит о достаточно комфортных условиях на планете в те времена.
Бесспорные эвкариотические организмы были описаны из отложений возрастом 2–2.2 Ga (Белова, Ахмедов, 2006). Это своеобразные органические остатки, часть из которых может оказаться кандидами грибов или протистами; также присутствуют Leiosphaeridia со складками смятия. Эвкариоты известны и в фосфоритах возрастом 2 млрд лет печенгской серии района Печенги. Последняя находка достаточно уверенно определяется как празинофиты, и названа нами Pechengia (Розанов, Астафьева, 2008).
Изучая архейские предверхнелопийские коры выветривания по вулканитам (гранитам и плагиогранитам) охтинской серии Лехтинской структуры Карелии (оз. Воронье, устье р. Нижней Охты) с возрастом 2.8 млрд лет, мы обнаружили комплекс ископаемых микроорганизмов, который оказался интересным и довольно разнообразным. Среди прочих в этом комплексе была встречена форма, интерпретация которой вызвала значительные затруднения. Это полуразрушенная удлиненно-овальная микрофоссилия, но учитывая ее архейский возраст, можно сказать, что сохранилась она довольно хорошо. Эта форма имеет довольно сложное внутреннее строение. Вероятно, она была покрыта оболочкой толщиной 2–3 мкм. Было сделано предположение, что подобные формы напоминают раковинных амеб, причем данных, опровергающих это, мы не имеем (Розанов, Астафьева, 2020).
В том же местонахождении были встречены и другие удлиненно-овальные расплющенные бутылковидные формы. Относительно этих микрофоссилий было сделано предположение, что они могут являться подобием хитинозой (Ископаемые бактерии…, 2011). О некотором сходстве с хитинозоями говорилось и в описании еще одной расплющенной изогнутой формы (Ископаемые бактерии…, 2011).
* * *
Автор признательна А.Ю. Розанову, Е.А. Жегалло, Л.В. Зайцевой, Г.Т. Ушатинской, всем, кто помогал в проведении этой работы и участвовал в обсуждении ее результатов. Особая благодарность – А.В. Ткачеву, С.Б. Фелицыну, Н.А. Алфимовой и сотрудникам ЦНИГР музея, передавшим образцы жайремских джеспилоидов и архейско-протерозойских железистых кварцитов для изучения.
Список литературы
Антошкина А.И. Бактериальное породообразование – реальность современных методов исследований // Учен. зап. Казан. ун-та. Сер. естеств. науки. 2011. Т. 153. Кн. 4. С. 114–126.
Астафьева М.М., Фелицын С.Б., Алфимова Н.А. Бактериальная палеонтология неоархейских полосчатых железистых кварцитов Карелии и Кольского полуострова // Палеонтол. журн. 2017. № 4. С. 93–102.
Бактериальная палеонтология. М.: ПИН РАН, 2002. 188 с.
Белова М.Ю., Ахмедов А.М. Petsamomyces – новый род органостенных микрофоссилий в углеродсодержащих отложениях раннего протерозоя Кольского полуострова // Палеонтол. журн. 2006. № 5. С. 3–12.
Брусницын А.И., Перова Е.Н., Верещагин О.С. и др. Стратиформные свинцово-цинковые, баритовые и железо-марганцевые руды Жайремского рудного узла (Центральный Казахстан): условия залегания, состав, генезис // Металлогения древних и современных океанов. Миасс: Ин-т минералогии УрО РАН, 2017. С. 90–94.
Брусницын А.И., Перова Е.Н., Верещагин О.С. и др. Барит-свинцово-цинковые и железо-марганцевые месторождения Жайремского рудного узла: геологическая экскурсия в Центральный Казахстан // Минералогия. 2018. Т. 4. № 3. С. 82–92.
Варенцов И.М., Веймарн А.Б., Рожнов А.А. и др. Геохимическая модель формирования марганцевых руд фаменского рифтогенного бассейна Казахстана (главные компоненты, редкие земли, рассеянные элементы) // Литол. и полезн. ископ. 1993. № 3. С. 56–79.
Вологдин А.Г. Земля и жизнь. М.: Недра, 1976. 238 с.
Заварзин Г.А. Развитие микробных сообществ в истории Земли // Проблемы доантропогенной эволюции биосферы. М.: Наука, 1993. С. 212–222.
Заварзин Г.А. Смена парадигмы в биологии // Вестн. РАН. 1995. Т. 6. № l. С. 8–23.
Заварзина Д.Г. Роль диссимиляторных железоредуцирующих бактерий в преобразовании минералов железа // Палеонтол. журн. 2004а. № 3. С. 3–10.
Заварзина Д.Г. Образование магнетита и сидерита термофильными железоредуцирующими бактериями // Палеонтол. журн. 2004б. № 6. С. 3–8.
Ископаемые бактерии и другие микроорганизмы в земных породах и астроматериалах / Ред. Розанов А.Ю., Ушатинская Г.Т. М.: ПИН РАН, 2011. 172 с.
Митряева Н.М. Минералогия барито-цинково-свинцовых руд месторождений Атасуйского района. Алма-Ата: Наука, 1979. 219 с.
Николаев Ю.А., Плакунов В.К. Биопленка – “город микробов” или аналог многоклеточного организма? // Микробиология. 2007. Т. 76. № 2. С. 149–163.
Рожнов А.А. Сравнительная характеристика марганцевых месторождений Атасуйского и Никопольско-чиатурского типов // Геология и геохимия марганца. М.: Наука, 1982. С. 116–121.
Розанов А.Ю. Условия жизни на ранней Земле после 4.0 млрд лет назад // Проблемы происхождения жизни. М.: ПИН РАН, 2009. С. 185–201.
Розанов А.Ю., Астафьева М.М. Празинофиты (зеленые водоросли) из нижнего протерозоя Кольского полуострова // Палеонтол. журн. 2008. № 4. С. 90–93.
Розанов А. Ю., Астафьева М.М. Архейские эвкариоты: новый взгляд // Палеонтол. журн. 2020. № 1. С. 3–5.
Розанов А.Ю., Астафьева М.М., Зайцева Л.В. и др. Цианобактерии (?) в железистых кварцитах Курской магнитной аномалии // Докл. Акад. наук. 2016. Т. 470. № 3. С. 1–3.
Розен О.М., Аббясов А.А., Аксаментова Н.В. и др. Седиментация в раннем докембрии: типы осадков, метаморфизованные осадочные бассейны, эволюция терригенных отложений. М.: Научн. мир, 2006. 398 с. (Тр. Геол. ин-та РАН. Т. 569).
Скрипченко Н.С. Гидротермально-осадочные полиметаллические руды известково-сланцевых формаций. М.: Недра, 1980. 215 с.
Скрипченко Н.С. Прогнозирование месторождений цветных металлов в осадочных породах. М.: Недра, 1989. 207 с.
Слободкин А.И. Термофильные железовосстанавливающие прокариоты. Автореф. дис. … докт. биол. наук. М., 2008. 50 с.
Холодов В.Н. К проблеме эволюции осадочного процесса в истории Земли // Проблемы доантропогенной эволюции биосферы. М.: Наука, 1993. С. 123–167.
Astafieva M.M., Zaitseva L.V. Bacterial-paleontological study of siderite transformation in the Early Proterozoic jaspilites of the Kursk Magnetic Anomaly // Paleontol. J. 2020. V. 54. № 8. P. 922–929.
Buik R. Ancient acritarchs // Nature. 2010. V. 463. P. 885–886.
Cloud P. Atmospheric and hydrospheric evolution on the primitive Earth // Science. 1968. V. 160. P. 729–736.
Costerton J.W., Geesey G.G., Cheng K.J. How bacteria stick // Sci. Amer. 1978. V. 238. P. 86–95.
Dodd M., Papineau D. Biosignatures of early life in >3.8 Ga Banded Iron Formations? // Geophysical Research Abstracts. 2015. V. 17. EGU2015-12987-1.
Klein C. Some Precambrian banded iron formations (BIFs) from around the world: their age, geologic setting, mineralogy, metamorphism, geochemistry, and origin // Amer. Mineralogist. 2005. V. 90. P. 1473–1499.
LaBerge G.L. Microfossils and Precambrian iron formations // Bull. Geol. Soc. Amer. 1967. V. 61. P. 572–581.
LaBerge G.L. Possible biological origin of Precambrian iron-formations // Econ. Geol. 1973. V. 68. P. 1098–1109.
Posth N.R., Konhauser K.O., Kappler A. Banded iron formations // Encyclopedia of Geobiology / Eds. Reitner J., Thiel V. Springer, 2011. 927 p.
Schopf J.W. (ed.). Earth’s Biosphere, its Origin and Evolution. Princeton: Princeton Univ. Press, 1983. 543 p.
Windley B.F. The evolving continents. Chichester: J. Wiley and Sons, 1999. 526 p.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал