

Палеонтологический журнал, 2021, № 3, стр. 61-69
Новая фебодонтидная акула из девона Урала и распространение видов рода Phoebodus
a Санкт-Петербургский государственный университет
Санкт-Петербург, Россия
b Казанский (Приволжский) федеральный университет
Казань, Россия
* E-mail: IvanovA-Paleo@yandex.ru
Поступила в редакцию 10.07.2020
После доработки 22.09.2020
Принята к публикации 24.09.2020
Аннотация
Из отложений среднего и верхнего девона Среднего и Южного Урала описывается новый вид фебодонтных хрящевых рыб Phoebodus curvatus. Зубы нового вида отличаются от зубов других видов этого рода очень длинными, лингвально изогнутыми боковыми вершинами, узким лабиобазальным отростком, выступающим за лабиальный край. Проведен анализ распространения всех известных видов Phoebodus. Изменен объем пяти фебодонтидных зон. Две радиации видов отмечаются в позднем фране и в среднем–позднем фамене.
ВВЕДЕНИЕ
Фебодонтидные хрящевые рыбы (отряд Phoebodontiformes) – одна из самых широко распространенных групп хрящевых рыб в позднем девоне. В данный отряд включено три рода: Diademodus Harris, 1951, Phoebodus St. John et Worthen, 1875 и Thrinacodus St. John et Worthen, 1875 (Ginter et al., 2010). Остатки фебодонтид в основном представлены изолированными зубами, но почти полный скелет Thrinacodus обнаружен в нижнекаменноугольных отложениях известного местонахожения Бер-Галч в Монтане, США (Grogan, Lund, 2008). Недавно скелет нового вида Phoebodus saidselachus найден в среднем фамене Марокко (Frey et al., 2019). Зубы акул, относящихся к 13 видам рода Phoebodus, встречены в морских отложениях живет–фаменского интервала многих регионов мира. Широкое распространение этих видов было использовано в предложенной фебодонтидной зональной шкале (Ginter, Ivanov, 1995, 2000).
Род Phoebodus, как и первый описанный его вид Ph. sophiae St. John et Worthen, были установлены по материалу из живетских отложений Айовы, США (St. John, Worthen, 1875). На протяжении достаточно продолжительного периода к этому роду относили зубы с тремя вершинами, почти равными по высоте, но, как выяснилось позже, некоторые из них к фебодонтидам не относятся (Ginter et al., 2010). Первая ревизия фебодонтид (Ginter, Ivanov, 1992) позволила пересмотреть видовой состав рода, установить три новых вида и три неназванных таксона: Phoebodus sp. A, Ph. sp. B и Ph. sp. C. Два первых были позже описаны, как Ph. latus Ginter et Ivanov и Ph. typicus Ginter et Ivanov (Ginter, Ivanov, 1995). Ph. sp. C так и остался неописанным в качестве нового вида, несмотря на то, что его определяли в Австралии и Кузнецком бассейне (Trinajstic, George, 2009; Иванов, Родина, 2010), помимо тех местонахождений на Южном Урале и в Свентокшиских горах, где он был впервые отмечен. В предлагаемой работе описывается этот таксон, как новый вид, на основе ранее известных и новых экземпляров из живетских и франских отложений Среднего и Южного Урала.
МАТЕРИАЛ И МЕТОДЫ
Зубы нового вида рода Phoebodus обнаружены в нескольких местонахождениях на Среднем и Южном Урале. На Среднем Урале они найдены в двух местонахождениях – Покровское (рис. 1, б) и Кривой рог (рис. 1, а). Первое местонахождение находится в окрестностях с. Покровское на р. Бобровке (левый приток р. Ирбит), в 10 км к западу от г. Артемовский, Свердловская обл. Обнажения живетских и франских отложений расположены по обоим берегам р. Бобровки на протяжении более 2 км (Наседкина, Бороздина, 1999; Бикбаев и др., 2014). Отложения высотинского горизонта среднего и верхнего живета представлены песчанистыми известняками с прослоями глин, а породы бродовского горизонта нижнего франа (возможно, и верхов живета) включают в себя неслоистые и массивные рифогенные известняки (Бикбаев и др., 2013). Кроме остатков разнообразных беспозвоночных, в местонахождении встречены микроостатки рыб: зубы Phoebodus fastigatus Ginter et Ivanov, Ph. latus Ginter et Ivanov и нового вида, разнообразные чешуи хрящевых, чешуи акантод, фрагменты пластинок птиктодонтидных плакодерм, зубы струнииформных саркоптеригий, зубы и чешуи лучеперых. Данные о присутствии зубов Omalodus sp. в этом местонахождении ошибочны (Бикбаев и др., 2014): омалодонтиды никогда не были найдены на Урале. Новый вид установлен в двух обнажениях в верхней части высотинского горизонта и нижней части бродовского горизонта, в интервале конодонтовых зон disparilis – falsiovalis (Бикбаев и др., 2014).
Рис. 1.
Расположение изученных районов (а) и местонахождений нового вида в Свердловской области (б) и Башкортостане (в).

Местонахождение Кривой рог расположено на левом берегу р. Вильвы, в 2 км к юго-востоку от г. Гремячинска, Пермский край (рис. 1, а). Здесь в карбонатных отложениях мендымского горизонта верхнего франа, соответствующего конодонтовой зоне Lower rhenana, найдены зубы нового вида, а также Phoebodus bifurcatus Ginter et Ivanov, Ph. latus Ginter et Ivanov, фрагменты пластинок птиктодонтид и артродир, зубы струнииформов.
На Южном Урале новый вид встречен в двух местонахождениях Башкортостана – Лемезинский и Ряузяк (рис. 1, в). Лемезинский находится на левом берегу р. Инзер, у бывшего пос. Лемезинский, в 4 км к север-северо-западу от д. Габдюково. Отложения мендымского горизонта верхнего франа (соответствующие интервалу конодонтовой зоны Lower rhenana), где встречен новый таксон, представлены толщей переслаивания тонкоплитчатых битуминозных и толстоплитчатых пелитоморфных известняков с прослоями углисто-глинистых сланцев (Маслов и др., 2019). Помимо нового вида, комплекс ихтиофауны из этого местонахождения включает в себя хрящевых Phoebodus bifurcatus Ginter et Ivanov, Protacrodontidae indet., акантод и лучеперых.
Местонахождение Ряузяк расположено на правом берегу р. Ряузяк, в 5 км выше д. Саргаево. Зубы описываемого вида обнаружены здесь в верхнефранских отложениях аскынского горизонта, в интервале, соответствующем конодонтовой зоне Upper rhenana, и представленном органогенными и массивными пелитоморфными известняками с обильной фауной беспозвоночных (Artyushkova et al., 2011). Ихтиофауна этого интервала включает в себя зубы хрящевых Phoebodus bifurcatus Ginter et Ivanov, Ph. latus Ginter et Ivanov, Ph. aff. fastigatus Ginter et Ivanov, Cladodoides sp., Protacrodus sp., чешуи хрящевых, акантод и палеонисков.
Зубы нового вида различной сохранности, часто с обломанными вершинами, без повреждений наружной поверхности, но несколько экземпляров из местонахождения Покровское несут следы биоэрозии на поверхности коронки (табл. XII , фиг. 3 ; см. вклейку).
Фотографии выполнены на сканирующих электронных микроскопах JEOL JSM-T300, Cambridge CamScan-4 и TescanVEGA-II XMU в ресурсном центре “Геомодель” Научного парка Санкт-Петербургского государственного ун-та (СПбГУ) и в Палеонтологическом ин-те им. А.А. Борисяка (ПИН) РАН. Для изучения внутреннего строения зубов нового вида использовался рентгеновский микротомограф Skyscan 1172 ресурсного центра “Рентгенодифракционные методы исследования” Научного парка СПбГУ. Сканирование производилось с использованием следующих параметров: фильтр Al, напряжение 80–100 кВ, сила тока 100–124 мкA, шаг вращения 0.4°. Полученные микротомографические данные были обработаны с помощью компьютерных программ NRecon, DataViewer, CTVox, CTAn. Томографирование трех зубов показало, что васкулярные каналы частично заполнены веществом, близким по химическому составу к фосфату зубов, и из-за низкой контрастности на виртуальной реконструкции прослеживается только часть каналов.
Описанные экземпляры хранятся в Палеонтологическом музее СПбГУ (колл. ПМ СПбГУ №№ 7 и 74).
СИСТЕМАТИЧЕСКАЯ ПАЛЕОНТОЛОГИЯ
КЛАСС CHONDRICHTHYES
ПОДКЛАСС ELASMOBRANCHII
ОТРЯД PHOEBODONTIFORMES
СЕМЕЙСТВО PHOEBODONTIDAE WILLIAMS IN ZANGERL, 1981
Род Phoebodus St. John et Worthen, 1875
Phoebodus curvatus Ivanov, sp. nov.
Табл. XII , фиг. 1–6
Рис. 2.
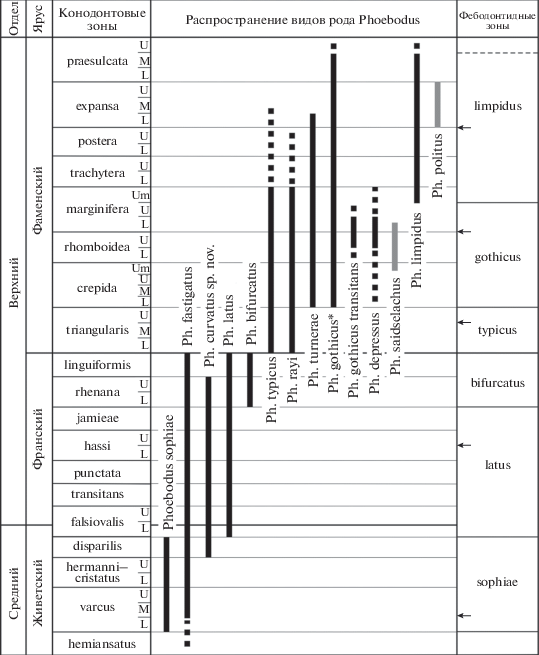
Распространение видов рода Phoebodus и зоны по фебодонтидам. Черная сплошная линия – точно датированный интервал по конодонтовым зонам; черная пунктирная – интервал таксона, определенного как “cf.” или “aff.”; серая сплошная – интервал, предположительно соответствующий конодонтовым зонам. Сокращения: L – Lower, M – Middle, U – Upper, Um – Uppermost. В колонке с фебодонтидными зонами стрелками показано положение границ зон в предыдущей варианте зональности (Ginter, Ivanov, 1995). * – интервал распространения Ph. gothicus, неразделенный на подвиды, или Ph. gothicus gothicus.
Phoebodus sp. C: Ginter, Ivanov, 1992, c. 71–72, фиг. 5 A–C ; Trinajstic, George, 2009, c. 651, табл. 1 , фиг. 10 ; Иванов, Родина, 2010, рис. 2, фиг. 3 .
Phoebodus fastigatus Ginter et Ivanov: Trinajstic, George, 2009, c. 648, 650 (part.), табл. 1 , фиг. 11, 14 .
Название вида curvatus лат. – изогнутый.
Голотип – ПМ СПбГУ 74-1, зуб; Южный Урал, Республика Башкортостан, лев. берег р. Инзер, Лемезинский; верхний девон, фран, мендымский горизонт.
Описание. Небольшие зубы с основанием шириной от 0.8 до 1.1 мм мезио-дистально. Фебодонтная коронка состоит из трех основных вершин: центральной и пары боковых, иногда с небольшими промежуточными вершинками. Вершины тонкие, заостренные, сигмоидально изогнутые, существенно наклоненные в лингвальном направлении, округлые в поперечном сечении, не учитывая бокового канта. Боковые вершины длинные: их высота более чем в 1.5 раза превышает высоту центральной, расходятся под углом в 30°–40° к центральной вершине. Хорошо выраженные режущие канты разделяют выпуклые лингвальную и лабиальную стороны вершин, у основания коронки канты соседних вершин сливаются. На внешней стороне боковых вершин кант не достигает основания. Боковые вершины расходятся от центральной в мезиальном и дистальном направлениях, образуя острый угол между ними. Вершины чаще гладкие, скульптура в виде тонких, редких ребрышек иногда заметна на лабиальной стороне. Основания центральной и боковых вершин располагаются по прямой линии вдоль лабиального края, промежуточные вершинки смещены в лабиальном направлении.
Основание овальной, реже округло-прямоугольной формы, направленное лингвально, вытянуто мезио-дистально, с узкой лабиальной частью, иногда с оттянутыми латеральными углами (табл. XII , фиг. 4–6 ). Пропорции основания варьируют: ширина превышает длину в 1.3–2 раза. Окклюзарная сторона выпуклая, несет апикальный сочленовный бугорок. Бугорок достаточно крупный и выпуклый, овальной формы, занимает большую часть поверхности основания и расположен близко к лингвальному краю. Базальная сторона вогнутая, с узким лабиобазальным отростком и утолщенным лингвальным краем. Отросток смещен за лабиальный край в окклюзарной проекции.
Вершины сложены ортодентином на большей части длины, и лишь в их основании присутствует остеодентин. Васкулярная система состоит из двух главных горизонтальных каналов, немногочисленных второстепенных горизонтальных и восходящих вертикальных, и пульпарных каналов (табл. XII , фиг. 1в). Крупные отверстия главных каналов расположены у лингвального края и в центре вогнутой базальной стороны основания. Мелкие отверстия второстепенных каналов окружают апикальный бугорок, а также образуют ряд над лабиобазальным отростком.
Сравнение. От всех известных видов рода Ph. curvatus sp. nov. отличается очень длинными, лингвально изогнутыми боковыми вершинами, узким лабиобазальным отростком, выступающим за лабиальный край. У зубов большинства видов Phoebodus боковые вершины сильнее расходятся от центральной в мезиальном и дистальном направлениях, образуя угол более 50°, в отличие от нового вида, у которого они направлены в лингвальную сторону под углом 30°–40° к центральной вершине. Зубы Ph. limpidus Ginter, Ph. fastigatus Ginter et Ivanov, Ph. typicus Ginter et Ivanov напоминают зубы нового вида. Однако зубы Ph. limpidus отличаются субтреугольным основанием, широкой лабиальной частью и лабиобазальным отростком; у Ph. fastigatus основание зуба трапециевидной формы с небольшим апикальным бугорком, а у Ph. typicus – прямоугольной формы (у нового вида основание овальной, реже округло-прямоугольной формы).
Замечания. Авторы (Ginter, Ivanov, 1992), впервые упомянувшие Phoebodus sp. C, не располагали достаточным материалом для описания нового вида. В настоящее время это стало возможным, благодаря новым находкам в среднем девоне Среднего Урала и экземплярам, обнаруженным в нижнефранских отложениях яйской свиты Кузнецкого бассейна (Иванов, Родина, 2010) и в отложениях верхнего франа бассейна Каннинг Западной Австралии (Trinajstic, George, 2009).
Распространение. Средний девон, верхи живетского яруса – верхний девон, франский ярус Среднего и Южного Урала, Кузнецкого бассейна, Польши и Австралии.
Материал. Кроме голотипа, шесть зубов из местонахождения Покровское, средний девон, живетский ярус, высотинский горизонт и верхний девон, франский ярус, бродовский горизонт (сборы А.З. Бикбаева и М.П. Снигиревой); три зуба из местонахождения Ряузяк, франский ярус, аскынский горизонт; два зуба из местонахождения Лемезинский (сборы А.Н. Абрамовой) и один зуб из местонахождения Кривой рог (сборы А.З. Бикбаева и М.П. Снигиревой), франский ярус, мендымский горизонт.
РАСПРОСТРАНЕНИЕ ВИДОВ PHOEBODUS В СРЕДНЕМ И ПОЗДНЕМ ДЕВОНЕ
В настоящее время род Phoebodus включает в себя 13 видов с учетом нового такона: Ph. bifurcatus Ginter et Ivanov; Ph. curvatus sp. nov.; Ph. depressus Ginter, Hairapetian et Klug; Ph. fastigatus Ginter et Ivanov; Ph. gothicus Ginter; Ph. latus Ginter et Ivanov; Ph. limpidus Ginter; Ph. politus Newberry; Ph. rayi Ginter et Turner; Ph. saidselachus Frey, Coates, Ginter, Hairapetian, Rücklin, Jerjen et Klug; Ph. sophiae St. John et Worthen; Ph. turnerae Ginter et Ivanov; Ph. typicus Ginter et Ivanov.
Два самых древних вида, Ph. sophiae и Ph. fastigatus, появились в живете среднего девона (рис. 2). Живетский вид Ph. sophiae встречен в интервале конодонтовых зон Middle varcus – disparilis в Северо-Восточной Австралии, Центральном Иране, Южной Мавритании, Свентокшиских горах Польши, Рейнских сланцевых горах Германии, Испании, Португалии, штатах Айова, Индиана и Нью-Йорк США (Ivanov, Derycke, 1999; Turner et al., 2000; Ginter et al., 2010), а в Кузнецком бассейне Западной Сибири (Россия) – в интервале зон Lower – Middle varcus (Ivanov, Rodina, 2001).
Ph. fastigatus известен из живет–франских отложений (интервал конодонтовых зон varcus – rhenana) Западной Австралии, Китая, Мавритании, Марокко, Кузнецкого бассейна, Среднего и Южного Урала России, Польши, Испании, штатов Айова, Индиана, Нью-Йорк, Юта США (Ginter, Ivanov, 1992; Ivanov, Derycke, 1999; Ivanov, 2008; Ginter et al., 2010; Roelofs et al., 2015). В нескольких работах упоминается возможное нахождение вида в более древних отложениях зоны hemiansatus (Liao et al., 2007; Derycke, 2017). В верхнем фране Среднего Урала этот вид найден и в конодонтовой зоне linguiformis (Ivanov, 2008). Таким образом, интервал стратиграфического распространения Ph. fastigatus отвечает почти всему живету и франу.
Новый вид Ph. curvatus установлен в верхнеживетских – нижнефранских отложениях (конодонтовые зоны disparilis – Middle falsiovalis) Среднего Урала, верхнефранских отложениях (конодонтовая зона rhenana) Среднего и Южного Урала, в верхнефранских отложениях Свентокшиских гор Польши (Ginter, Ivanov, 1992), а также в нижнефранских отложениях Кузнецкого бассейна (Иванов, Родина, 2010). В Западной Австралии данный вид обнаружен в отложениях, соответствующих интервалу hassi – Lower rhenana (Trinajstic, George, 2009). Эти авторы привели распространение вида (под названием Ph. sp. C) в других регионах мира, соответствующее интервалу конодонтовых зон punctata–rhenana, ссылаясь на ранее опубликованные данные (Ginter, Ivanov, 1995a), несмотря на то, что в упомянутой статье нет сведений об этом таксоне. Ph. curvatus появился в позднем живете, но стал широко распространенным видом в позднем фране.
Традиционно считают Ph. latus франским видом (Ginter et al., 2010). Действительно, в большинстве регионов мира он встречен в интервале конодонтовых зон hassi–linguiformis. Вид отмечают в среднем–верхнем фране Западной Австралии (Roelofs et al., 2015); Южного и Среднего Урала, Южного Тимана, Кузнецкого бассейна, Горного Алтая России (Ivanov, 1995, 1999, 2008; Ivanov, Rodina, 2001); Мавритании (Racheboeuf et al., 2001); Свентокшиских гор Польши (Ginter, Ivanov, 1992). Но Ph. latus установлен также и в нижнем фране (интервал Middle falsiovalis–hassi) Ирана (Hairapetian et al., 2008), а на Среднем Урале он определен в верхах живета, в зоне Lower falsiovalis (Ivanov, 2008).
Широко распространенный вид Ph. bifurcatus известен из достаточно узкого интервала конодонтовых зон rhenana–linguiformis верхнего франа Западной Австралии, Китая, Мавритании, Ирана, Польши, Чехии, Бельгии, штата Юта США (Ginter et al., 2010, 2017; Roelofs et al., 2015). В России этот вид найден в верхнем фране Южного Тимана, Центрального девонского поля, Среднего и Южного Урала, Кузнецкого бассейна, Горного Алтая (Ivanov, 1995, 1999; Ivanov, Rodina, 2001).
На границе франа и фамена наблюдается полная смена видового состава Phoebodus: живет–франские виды исчезают, а с фамена начинается новая радиация фебодонтид (рис. 2). Два вида – Ph. typicus и Ph. rayi – появляются в раннем фамене, начиная с зоны Lower triangularis. Ph. typicus встречается в большинстве регионов в нижнем и среднем фамене (Ginter et al., 2010). Данный вид обнаружен в интервале конодонтовых зон Upper triangularis–Lower marginifera Австралии, Марокко, Ирана, Южного Урала и Кузнецкого бассейна России (Ginter, Ivanov, 2000; Hairapetian, Ginter, 2009; Ginter et al., 2010). Но в Кузнецком бассейне он найден также в интервале Lower–Middle triangularis (Ivanov, Rodina, 2001). Ph. cf. typicus установлен в нижнем и среднем фамене Армении и Беларуси (Ginter et al., 2011; Ivanov, Plax, 2018), а в Кузнецком бассейне – в верхнем фамене (Ivanov, Rodina, 2004). Недавно Ph. typicus определен автором из отложений верхней части покаямской свиты на р. Волонга Северного Тимана (сборы П.А. Безносова).
Ph. rayi установлен из нижнего фамена (конодонтовая зона crepida) Австралии и Северной Канады (Ginter, Turner, 1999; Ginter et al., 2010). В Кузнецком бассейне вид найден в интервале зон Lower crepida–Uppermost marginifera (Иванов, Родина, 2002), а в Иране – в зоне Lower triangularis (Hairapetian, Ginter, 2010). Находки Phoebodus cf. rayi известены из интервала Middle crepida–postera Ирана (Yazdi, Turner, 2000; Ginter et al., 2002), и из зоны crepida Литвы (Valiukevičius, Ovnatanova, 2005).
Вид Ph. turnerae в основном распространен в интервале конодонтовых зон Lower crepida–Uppermost marginifera Ирана, Алжира, Армении, Беларуси, Польши, Бельгии, штата Аляска США (Derycke-Khatir, 2005; Ginter et al., 2010, 2011; Ivanov, Plax, 2018). На Южном Урале он установлен в широком интервале, с Lower crepida по Lower expansa (Ivanov, 1995), в Кузнецком бассейне – с rhomboidea по trachytera (Иванов, Родина, 2010). В Марокко распространение вида указывается в интервале конодонтовых зон с crepida по postera (Derycke, 2017). Кроме того, Ph. turnerae установлен автором в верхней части покаямской свиты на р. Волонга Северного Тимана (сборы П.А. Безносова). Ph. cf. turnerae отмечен в самом верхнем фамене Западной Австралии (Roelofs et al., 2015).
Вид Ph. gothicus был разделен на два подвида, Ph. gothicus gothicus Ginter и Ph. g. transitans Ginter, Hairapetian et Klug (Ginter et al., 2002). Судя по данным этих авторов, большинство находок, определенных до этого как Ph. gothicus, относится к первому подвиду. Этот подвид распространен преимущественно в интервале конодонтовых зон Lower marginifera–Middle praesulcata во многих регионах мира: в Южном Китае, Центральном Иране, Армении, Марокко, Южном Урале России, Польше, Германии, Франции, штатах Айова, Огайо, Юта США (Ginter, Ivanov, 1992; Hampe, 2000; Ginter et al., 2002, 2010, 2011; Hampe, Schindler, 2004). Но в Иране и Марокко вид известен с зоны Lower crepida (Ginter et al., 2002; Hairapetian, Ginter, 2009; Derycke, 2017). Ph. cf. gothicus определен из интервала конодонтовой зоны marginifera Вьетнама (Long, 1993). Ph. gothicus transitans описан только из нижнего или среднего фамена Алжира (Ginter et al., 2002), но позже показано его распространение в интервале конодонтовых зон Upper rhomboidea–Lower marginifera (Ginter et al., 2010). Ph. cf. gothicus transitans встречен в интервале rhomboidea–Upper marginifera в Иране (Hairapetian, Ginter, 2009).
Находка Ph. depressus отмечена в среднем фамене Алжира (интервал Upper rhomboidea–Lower marginifera по: Ginter et al., 2010), а Ph. cf. depressus найден в Иране в интервале crepida–Upper marginifera (Hairapetian, Ginter, 2009).
Интервал распространения Ph. saidselachus определен авторами таксона как нижняя часть среднего фамена, без указания точного интервала конодонтовых зон (Frey et al., 2019).
Вид Ph. limpidus в основном встречен в верхнем фамене, в интервале конодонтовых зон Lower expansa–Upper praesulcata в Южном Китае, на Южном Урале и Северном Кавказе России, в Марокко, Польше, Германии, Франции, штатах Невада, Вайоминг и Юта США (Ginter, Ivanov, 2000; Ginter, 2001; Ivanov, 2003; Ginter et al., 2010). Но в Карнийских Альпах Италии вид найден и в более древних отложениях, здесь он определен в интервале конодонтовых зон Uppermost marginifera–Lower praesulcata (Randon et al., 2007).
Позднефаменский вид Ph. politus описан из пачки сланцев Кливленд штата Огайо, США (Newberry, 1889; Ginter et al., 2010). Эти отложения Д. Блад и др. (Blood et al., 2019) сопоставляют с конодонтовыми зонами aculeatus и, возможно, costatus старой конодонтовой шкалы (Sandberg et al., 1972), или зоной expansa новой шкалы.
Виды-космополиты Phoebodus – Ph. bifurcatus, Ph. fastigatus, Ph. gothicus, Ph. latus, Ph. limpidus, Ph. sophiae, Ph. turnerae и Ph. typicus – встречены во многих регионах мира. Большинство из упомянутых таксонов найдены как на территории Лавруссии, так и Северной и Восточной Гондваны, Сибири, иногда и Южного Китая. В среднем и позднем фамене появились несколько видов-эндемиков, таких как Ph. depressus, Ph. politus, Ph. saidselachus. Следует отметить две радиации видов рода Phoebodus, приходящиеся на поздний фран и средний–поздний фамен. Во время второй радиации одновременно существовало шесть видов.
ФЕБОДОНТИДНЫЕ ЗОНЫ
На основе анализа распространения видов Phoebodus было предложено шесть фебодонтидных зон для живета, франа и фамена (Ginter, Ivanov, 1995, 2000). Фебодонтидная зона sophiae соответствовала интервалу конодонтовых зон Middle varcus–Lower hassi живета и франа, зона latus отвечала конодонтовым зонам Upper hassi–jamieae франа, зона bifurcatus – интервалу rhenana–linguiformis франа (Ginter, Ivanov, 1995). В фамене фебодонтидная зона typicus начиналась с Upper triangularis и продолжалась до Upper rhomboidea, зона gothicus соответствовала интервалу конодонтовых зон Lower marginifera–Upper postera, а зона limpidus отвечала интервалу Lower expansa–Middle praesulcata (Ginter, Ivanov, 1995). Таким образом, в интервале конодонтовых зон Lower – Middle triangularis нижнего фамена не было данных о распространении видов фебодонтид.
Несмотря на то, что Phoebodus gothicus был найден в Иране в интервале конодонтовых зон crepida–rhomboidea (Ginter et al., 2002), в последующих публикациях нижняя граница этой зоны осталась неизменной (Ginter et al., 2010). Как видно из анализа распространения видов Phoebodus, за последнее время появилось много новых данных о находках некоторых зональных видов за пределами ранее известных стратиграфических интервалов. Ph. latus появился с конодонтовой зоны Lower falsiovalis. Ph. typicus установлен в Lower и Middle triangularis, заполняя пробел в данных о фебодонтидах в этом интервале. Ph. gothicus теперь известен из интервала crepida–rhomboidea, а самая ранняя находка Ph. limpidus датирована конодонтовой зоной Uppermost marginifera. В результате модификации объемов зон и положения границ (рис. 2), неизменной осталась одна фебодонтидная зона, bifurcatus. Зона sophiae теперь соответствует интервалу конодонтовых зон Lower varcus–disparilis, зона latus расширена до интервала конодонтовых зон Lower falsiovalis–jamieae, зона typicus отвечает только интервалу Lower–Upper triangularis, зона gothicus изменена до интервала Lower crepida–Upper marginifera, а нижняя граница зоны limpidus смещена на нижнюю границу Uppermost marginifera.
* * *
Автор статьи благодарен А.Н. Абрамовой и О.В. Артюшковой (ИГ УФИЦ РАН), А.З. Бикбаеву и М.П. Снигиревой (ИГГ УРО РАН), П.А. Безносову (ИГ КНЦ УРО РАН) за предоставленные образцы для изучения и информацию по стратиграфии; С.П. Нилову (СПбГУ) за микротомографию зубов; Н.М. Власенко (СПбГУ) и Р.А. Ракитову (ПИН РАН) за помощь при работе на сканирующих электронных микроскопах, а также О.А. Лебедеву (ПИН РАН) и Е.В. Попову (СГУ) за рецензирование рукописи и ценные замечания. Работа выполнена за счет средств Программы стратегического академического лидерства Казанского (Приволжского) федерального университета и при частичной финансовой поддержке РФФИ, грант № 20-05-00445а. Исследования проведены с использованием оборудования ресурсных центров “Геомодель” и “Рентгенодифракционные методы исследования” Научного парка СПбГУ.
Список литературы
Бикбаев А.З., Снигирева М.П., Тупицына М.А. К вопросу о возрасте брахиоподового комплекса с Rhyssochonetes rugosus в живетских отложениях типового разреза “Покровское” (восточный склон Среднего Урала) // Тр. ИГГ УрО РАН. 2013. Вып. 160. С. 8–12.
Бикбаев А.З., Снигирева М.П., Тупицына М.А. Литологическая характеристика и конодонты верхнеживетских отложений типового разреза “Покровское” // Тр. ИГГ УрО РАН. 2014. Вып. 161. С. 7–14.
Маслов А.В., Артюшкова О.В., Тагариева Р.Ч. и др. Систематика редкоземельных элементов, Y, Th, U и Mn в верхнедевонских конодонтах Западно-Уральской внешней зоны складчатости (Южный Урал) // Литосфера. 2019. Т. 19. № 2. С. 250–268.
Наседкина В.А., Бороздина Г.Н. Конодонты из пограничных отложений среднего и верхнего девона на восточном склоне Среднего Урала // Проблемы стратиграфии и палеонтологии Урала / Ред. Золоев К.К., Анцыгин Н.Я. Екатеринбург: Минприроды РФ, ОАО УГСЭ, 1999. С. 45–51.
Иванов А.О., Родина О.А. Смена комплексов хрящевых рыб на границе франа и фамена в Кузнецком бассейне // Геология девонской системы. Матер. Междунар. симпозиума. Сыктывкар, 2002. С. 84–87.
Иванов А.О., Родина О.А. Комплексы хрящевых рыб из верхнедевонских отложений бассейна р. Яя (Кузнецкий бассейн) // Природа и экономика Кузбасса и сопредельных территорий. Матер. Всеросс. науч. конф. / Ред. Удодов В.П. Новокузнецк, 2010. С. 47–52.
Artyushkova O.V., Maslov V.A., Pazukhin V.N. et al. Devonian and Lower Carboniferous type sections of the western South Urals. Pre-Conference Field Excursion Guidebook. Ufa: UIG RAN, 2011. 92 p.
Blood D.R., Baird G.C., Danielsen E.M. et al. Upper Devonian paleoenvironmental, diagenetic, and tectonic enigmas in the western Appalachian Basin: new discoveries and emerging questions associated with the Frasnian–Famennian boundary and end-Devonian disturbances in central Ohio. Field Trip Guidebook, Eastern Section Annual Meeting of the Amer. Assoc. of Petroleum Geologists. Columbus, Ohio: AAPG, 2019. 80 p.
Derycke C. Paléobiodiversité des gnathostomes (chondrichthyens, acanthodiens et actinoptérygiens) du Dévonien du Maroc (NW Gondwana) // Mém. Soc. Géol. Fr. 2017. № 180. P. 44–77.
Derycke-Khatir C. Microrestes de vertébrés du Paléozöique supérieur de la Manche au Rhin // Soc. Géol. du Nord Publ. 2005. № 33. P. 1–261.
Frey L., Coates M.I., Ginter M. et al. The early elasmobranch Phoebodus: phylogenetic relationships, ecomorphology and a new time-scale for shark evolution // Proc. Roy. Soc. Ser. B. 2019. V. 286. 20191336. P. 1–10.
Ginter M. Chondrichthyan biofacies in the late Famennian of Utah and Nevada // J. Vertebr. Paleontol. 2001. V. 21. P. 714–729.
Ginter M., Gouwy S., Goolaerts S. A classic Late Frasnian chondrichthyan assemblage from southern Belgium // Acta Geol. Pol. 2017. V. 67. № 3. P. 381–392.
Ginter M., Hairapetian V., Grigoryan A. Chondrichthyan microfossils from the Famennian and Tournaisian of Armenia // Acta Geol. Pol. 2011. V. 61. № 2. P. 153–173.
Ginter M., Hairapetian V., Klug C. Famennian chondrichthyans from the shelves of North Gondwana // Acta Geol. Pol. 2002. V. 52. № 2. P. 169–215.
Ginter M., Hampe O., Duffin C.J. Chondrichthyes Paleozoic Elasmobranchii: Teeth // Handbook of Palaeoichthyology, v. 3D / Ed. Schultze H.-P. München: Verl. Dr. F. Pfeil, 2010. P. 1–168.
Ginter M., Ivanov A. Devonian phoebodont shark teeth // Acta Palaeontol. Pol. 1992. V. 37. № 1. P. 55–75.
Ginter M., Ivanov A. Middle/Late Devonian phoebodont-based ichthyolith zonation // Géobios. 1995. Mém. Spec. № 19. P. 351–355.
Ginter M., Ivanov A. Stratigraphic distribution of chondrichthyans in the Devonian on the East European Platform margins // Cour. Forsch.-Inst. Senckenb. 2000. Bd 223. P. 325–339.
Ginter M., Turner S. The early Famennian recovery of phoebodont sharks // Acta Geol. Pol. 1999. V. 49. № 2. P. 105–117.
Grogan E.D., Lund R. A basal elasmobranch, Thrinacoselache gracia n. gen. & sp. (Thrinacodontidae, new family) from the Bear Gulch Limestone, Serpukhovian of Montana, USA // J. Vertebr. Paleontol. 2008. V. 28. № 4. P. 970–988.
Hairapetian V., Ginter M. Famennian chondrichthyan remains from the Chahriseh section, central Iran // Acta Geol. Pol. 2009. V. 59. № 2. P. 173–200.
Hairapetian V., Ginter M. Pelagic chondrichthyan microremains from the Upper Devonian of the Kale Sardar section, eastern Iran // Acta Geol. Pol. 2010. V. 60. № 3. P. 357–371.
Hairapetian V., Ginter M., Yazdi M. Early Frasnian sharks from central Iran // Acta Geol. Pol. 2008. V. 58. № 2. P. 173–179.
Hampe O. Occurrence of Phoebodus gothicus (Chondrichthyes: Elasmobranchii) in the middle Famennian of northwestern Iran (Province East Azerbaijan) // Acta Geol. Pol. 2000. V. 50. № 3. P. 355–367.
Hampe O., Schindler T. Late Famennian fish remains from the Refrath 1 Borehole (Bergisch Gladbach-Paffrath Syncline; Ardennes-Rhenish Massif, Germany) // Cour. Forsch.-Inst. Senckenb. 2004. Bd 251. P. 267–289.
Ivanov A. Late Devonian vertebrate fauna of the South Urals // Geobios. 1995. Mém. Spec. № 19. P. 357–359.
Ivanov A. Late Devonian – Early Permian chondrichthyans of the Russian Arctic // Acta Geol. Pol. 1999. V. 49. № 3. P. 267–285.
Ivanov A. Vertebrates from the Late Devonian–Carboniferous of North Caucasus (Russia) // Ichthyolith Iss., Spec. Publ. 2003. № 7. P. 27–28.
Ivanov A. Vertebrate assemblages from the lower and upper Frasnian boundaries of the Middle Urals // Abstr. of 20 Intern. Senckenberg-Conf. and Second Geinitz-Conf. Frankfurt am Main, 2008. P. 56.
Ivanov A., Derycke C. Distribution of the Givetian Omalodus shark assemblage // Ichthyolith Iss., Spec. Publ. 1999. № 5. P. 22–24.
Ivanov A.O., Plax D.P. Chondrichthyans from the Devonian–Early Carboniferous of Belarus // Est. J. Earth Sci. 2018. V. 67. № 1. P. 43–58.
Ivanov A., Rodina O. Middle and Late Devonian vertebrate biostratigraphy of SW Siberia, Russia // Abstr. of the 15th Int. Senckenberg Conference. Frankfurt am Main, 2001. P. 50.
Ivanov A., Rodina O. A new omalodontid-like shark from the Late Devonian (Famennian) of western Siberia, Russia // Fossils and Strata. 2004. № 50. P. 82–91.
Liao J.-C., Ginter M., Valenzuela-Rios J.I. Chondrichthyan microremains from the Givetian of the Aragonian Pyrenees (Spain) // Bull. Soc. Géol. Fr. 2007. T. 178. № 3. P. 171–178.
Long J.A. Palaeozoic vertebrate biostratigraphy of south-east Asia and Japan. Chapter 11 // Palaeozoic Vertebrate Biostratigraphy and Biogeography / Ed. Long J.A. L.: Belhaven Press, 1993. P. 277–289.
Newberry J.S. The Paleozoic fishes of North America // U.S. Geol. Surv. Monogr. 1889. V. 16. P. 1–340.
Racheboeuf P.R., Girard C., Lethiers F. et al. Evidence for Givetian stage in the Mauritanian Adrar (West Africa): biostratigraphical data and palaeogeographic implications // Newsl. Stratigr. 2001. V. 38. № 2–3. P. 141–162.
Randon C., Derycke C., Blieck A. et al. Late Devonian – Early Carboniferous vertebrate microremains from the Carnic Alps, northern Italy // Geobios. 2007. V. 40. № 6. P. 809–826.
Roelofs B., Playton T., Barham M. et al. Upper Devonian microvertebrates from the Canning Basin, Western Australia // Acta Geol. Pol. 2015. V. 65. № 1. P. 69–101.
Sandberg C.A., Streel M., Scott R.A. Comparison between conodont zonation and spore assemblages in the Devonian–Carboniferous boundary in the western and central United States and in Europe // Septième Congrès Internationale de Stratigraphie Géologie du Carbonifère, Krefeld, 23–28 August 1971, Compte Rendu 1. Krefeld, 1972. P. 179–203.
St. John O., Worthen A.H. Description of fossil fishes // Geol. Surv. Ill., Paleontol. 1875. V. 6. P. 245–488.
Trinajstic K., George A.D. Microvertebrate biostratigraphy of Upper Devonian (Frasnian) carbonate rocks in the Canning and Carnarvon basins of Western Australia // Palaeontology. 2009. V. 52. P. 641–659.
Turner S., Basden A., Burrow C.J. Devonian vertebrates of Queensland // Cour. Forsch.-Inst. Senckenb. 2000. Bd 223. P. 487–521.
Valiukevičius J., Ovnatanova N. The Early Famennian conodonts and fishes of Lithuania // Geologija. 2005. T. 49. P. 21–28.
Yazdi M., Turner S. Late Devonian and Carboniferous vertebrates from the Shishtu and Sardar Formations of the Shotori Range, Iran // Rec. West. Austral. Mus., Suppl. 2000. V. 58. P. 223–240.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал