

Палеонтологический журнал, 2021, № 3, стр. 70-82
Возрастная изменчивость в гистологическом строении посткраниального скелета Platyoposaurus stuckenbergi (Temnospondyli, Archegosauridae) из средней перми Восточной Европы
А. В. Ульяхин a, b, *, П. П. Скучас c, **, П. Г. Сабуров c, ***
a Московский государственный университет имени М.В. Ломоносова
Москва, Россия
b Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
c Санкт-Петербургский государственный университет
Санкт-Петербург, Россия
* E-mail: ulyakhin@paleo.ru
** E-mail: p.skutschas@spbu.ru
*** E-mail: p.saburov@spbu.ru
Поступила в редакцию 21.09.2020
После доработки 16.10.2020
Принята к публикации 20.10.2020
Аннотация
Описывается микроанатомическое и гистологическое строение разноразмерных бедренных костей и гипоцентров архегозавроидной амфибии Platyoposaurus stuckenbergi из средней перми Восточной Европы (уржумский горизонт, зона Estemmenosuchus uralensis). Дается сравнительная характеристика возрастной изменчивости, рассматриваются особенности экологии Platyoposaurus. Бедренные кости P. stuckenbergi характеризуются полным отсутствием кальцинированного хряща и наличием линий остановки роста (“LAG”) числом до 15; гипоцентры – остеопорозным строением, отсутствием кальцинированного хряща и линий остановок роста. Микроанатомические и морфологические особенности указывают на преимущественно водный образ жизни Platyoposaurus, как активного хищника речных и озерных мест обитания.
ВВЕДЕНИЕ
Архегозавриды (Archegosauridae) – группа темноспондильных амфибий, известных из нижне-среднепермских отложений Центральной и Восточной Европы, а также Бразилии и Индии (Schoch, Milner, 2000). Представители семейства предположительно были облигатно-водными рыбоядными хищниками (Губин, 1991; Ивахненко, 2001) с размером тела до 5–6 м (Price, 1948). Отличительными особенностями морфологии архегозаврид являются удлиненная и узкая предорбитальная часть черепа у взрослых особей, сближенная с лобной слезная кость, слабо редуцированная основная затылочная кость, умеренно глубокие ушные вырезки, глубокий затылок, отсутствие межхоанального зубного ряда, ровная crista arcuata нижней челюсти (Губин, 1991; Gubin, 1997).
Архегозавридные темноспондилы рода Platyoposaurus были крупными (общая реконструируемая длина черепа до 0.7 м; длина тела около 3.5 м) гавиалоподобными амфибиями, широко распространенными в средней перми на территории современной Восточной Европы (Ивахненко, 2001). Род Platyoposaurus представлен четырьмя описанными видами (Ивахненко и др., 1997), остатки которых достоверно известны на сегодняшний день из 18 местонахождений. Однако есть основания выделять только три вида (Schoch, Milner, 2000), поскольку P. vjuschkovi Gubin, 1989 описан по бедренной кости (Губин, 1989), которая не несет диагностических признаков, поэтому ее стоит рассматривать как Platyoposaurus sp. Платиопозавры впервые появляются в ископаемой летописи в казанском веке и исчезают в конце уржумского (возможно, в начале северодвинского) века по региональной геохронологической шкале Восточно-Европейской платформы, характеризуя очерский и ишеевский комплексы тетрапод (Ивахненко и др., 1997). Древнейшая из известных находок представителей рода Platyoposaurus представлена единственной заднетеменной костью (экз. ПИН, № 4276/37) из нижнеказанского местонахождения Голюшерма республики Удмуртия, реперного для голюшерминского фаунистического субкомплекса (Голубев, 1996; Ивахненко и др., 1997). Наиболее ранний из описанных видов P. stuckenbergi Trautschold, 1884 известен по относительно полному скелету (экз. ПИН, № 164/1) из низов верхней казани местонахождения Белебей республики Башкортостан (голюшерминский фаунистический субкомплекс; Новожилов, 1955; Ивахненко и др., 1997). Более высокое стратиграфическое положение занимает Platyoposaurus watsoni Efremov, известный по массовым, преимущественно черепным остаткам (колл. ПИН, №№ 161, 2250, 5021) из верхов верхней казани местонахождения Шихово-Чирки Кировской обл. (голюшерминский фаунистический субкомплекс; Efremov, 1932; Ивахненко и др., 1997). Наиболее молодой представитель рода Platyoposaurus (Platyoposaurus vjuschkovi по: Губин, 1991; голотип – бедренная кость ПИН, № 272/57) происходит из верхнеуржумского (?) местонахождения Малая Кинель Оренбургской обл. (ишеевский фаунистический комплекс; Вьюшков, 1955; Ивахненко и др., 1997). Стратиграфическое положение утраченного фрагмента черепа Platyoposaurus rickardi Twelvetrees, 1880 (Twelvetrees, 1880; Ивахненко и др., 1997), известного из местонахождения Рождественский Рудник из группы Каргалинских рудников, остается неясным.
В настоящее время морфологические особенности Platyoposaurus хорошо охарактеризованы; выявлены черты сходства и различия между видами рода и прочими представителями семейства Archegosauridae (Конжукова, 1955; Губин, 1991). При этом вопросы, касающиеся особенностей онтогенеза, предполагаемого индивидуального возраста (Конжукова, 1964), палеоэкологии и типов адаптации (Конжукова, 1955; Губин, 1991; Witzmann, Brainerd, 2017) архегозаврид в целом и Platyoposaurus, в частности, остаются недостаточно изученными и спорными.
Палеогистологические исследования способны решать ряд важных вопросов, в том числе, связанных с биологическими особенностями ископаемых позвоночных, а также с реконструкцией среды их обитания (напр., климатические особенности) и условиями гибели. В настоящее время известно, что существует прямая связь между микроанатомическим строением костей и образом жизни животных (de Ricqlès, de Buffrénil, 2001; Laurin et al., 2004). Устанавливаемые по палеогистологических данным особенности индивидуального развития (характер и темпы роста) и, соответственно, возрастная изменчивость, в свою очередь, позволяют выявить условия, при которых организм развивался. Это касается также стратегий питания и трофических взаимодействий (Sanchez et al., 2010). Помимо прямой связи морфологических параметров костей с особенностями локомоции (Wall, 1983; Botha, Chinsamy, 2004; Ray, Chinsamy, 2004; Ray et al., 2005; Kriloff et al., 2008), “биомеханически” информативными оказываются также микроанатомические и гистологические параметры (Sanchez et al., 2010). Остановка и замедление роста костей, которые проявляются в наличии линий остановки (LAG’s) и задержки роста (“annuli”), помимо характера индивидуального развития, могут отражать общие особенности среды обитания животных (Steyer et al., 2004). Кроме того, подсчет линий остановки роста используется в скелетохронологии (определение индивидуального возраста) и установлении связи возраста с размерами животных (Steyer et al., 2004; Gee et al., 2020).
Цель настоящей работы – на основании гистологических данных уточнить характер индивидуального развития и выявить особенности экологии Platyoposaurus, а также возможную смену местообитания в онтогенезе на основании сравнения с гистологическими данными по другим темноспондилам, в том числе облигатно-водного и неотенического Dvinosaurus campbelli (Ульяхин и др., 2020).
Гистологическое строение костей Archegosauridae в настоящее время практически не изучено. Первый опыт подобных исследований для представителей рассматриваемого семейства принадлежит Г.А. Траутшольду, который описал микроанатомическое строение Platyoposaurus по пришлифовкам покровных элементов черепа, нижней челюсти и зубам P. stuckenbergi (Trautschold, 1884). Позднее А.П. Быстров охарактеризовал микроанатомическое строение покровных элементов черепа на примере P. watsoni, сравнив его с Benthosuchus sushkini (Efremov, 1929) и Dvinosaurus (Bystrow, 1938; Быстров, 1957) и отметив схожесть больших каналов базальной части костей и сети анастомозирующих канальцев у первых двух видов. Исследования по палеогистологии Archegosauridae, начатые Траутшольдом и Быстровым, впоследствии никем не были продолжены. В данной работе представлены результаты анализа костной гистологии наиболее информативных посткраниальных элементов Platyoposaurus stuckenbergi, в том числе позвонков, не часто используемых при палеогистологических исследованиях (Mukherjee et al., 2010), несмотря на то, что элементы позвоночника могут встречаться в ископаемом состоянии чаще прочих частей скелета.
Авторы благодарны В.К. Голубеву (ПИН РАН) за оказанную помощь и консультации при подготовке работы, В.В. Силантьеву и Р.Д. Петровой (КФУ) за предоставленный для исследования материал, а также А.В. Барановой (ВНИГНИ) за съемку гистологических шлифов. Работа выполнена при финансовой поддержке РФФИ в рамках научных проектов №№ 19-04-00060, 20-04-00545 и 20-54-12013.
МАТЕРИАЛ И МЕТОДИКА
Изучен материал из коллекций №№ 49, 164, 1955, 2250, 2255, 3968 Палеонтологического ин-та им. А.А. Борисяка РАН (ПИН), а также коллекции № 1044/П 637 Казанского (Приволжского) федерального университета (КФУ).
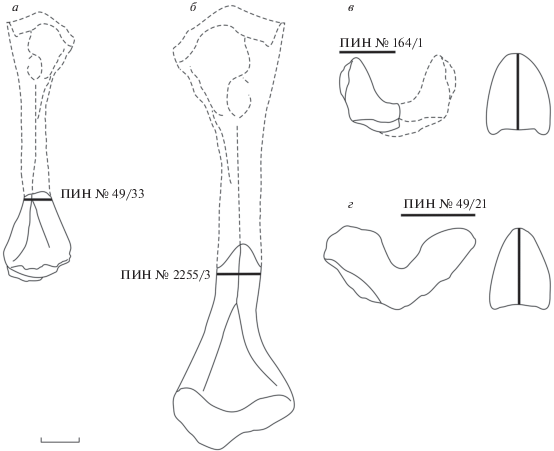
Для изучения гистологического строения посткраниальных элементов Platyoposaurus stuckenbergi были изготовлены стандартные тонкие петрографические срезы следующих образцов, происходящих из различных местонахождений одного стратиграфического уровня (уржумский горизонт, зона Estemmenosuchus uralensis): дистальный фрагмент мелкой (реконструируемая длина около 80 мм) левой бедренной кости (экз. ПИН, № 49/33) (рис. 1, а) и крупный (ширина 35 мм) гипоцентр (экз. ПИН, № 49/21) (рис. 1, г) из местонахождения Акбатыровский Рудник Малмышского р-на Кировской обл.; дистальный фрагмент крупной (реконструируемая длина около 120 мм) правой бедренной кости (экз. ПИН, № 2255/3) из местонахождения Большой Китяк-1 Малмышского р-на Кировской обл. (рис. 1, б); фрагмент мелкого (реконструируемая ширина около 25 мм) гипоцентра (экз. ПИН, № 164/1) из местонахождения Белебей Белебеевского р-на Башкортостана (рис. 1, в). Отобранный материал происходит из разных местонахождений схожего генезиса (справедливо для бедренных костей), поскольку относимые к P. stuckenbergi экземпляры из отдельно взятого местонахождения не позволяют рассматривать возрастную изменчивость по гистологическим данным.
Рис. 1.
Platyoposaurus stuckenbergi Trautschold, 1884, контуры мелкой (а) и крупной (б) бедренных костей; мелкого (в) и крупного (г) гипоцентров с указанием уровня взятия проб для микроанатомического и гистологического анализа.
Размерные показатели по бедренным костям и гипоцентрам P. stuckenbergi (рис. 2) оценивались в процентах относительно максимально известной реконструируемой длины черепа для рассматриваемого вида в 0.68 м (экз. ПИН б/н, отвалы медных рудников Кировской обл.), принятого за 100%. Максимальная длина черепа была определена с учетом имеющихся размерных параметров единственного и относительно полного скелета из местонахождения Белебей (экз. ПИН, № 164/1). Величины сопоставлялись с учетом одинаковой пропорции тела для разноразмерных особей. По такому же принципу были определены длины черепов для исследуемого в работе посткраниального материала.
Рис. 2.
Platyoposaurus stuckenbergi Trautschold, 1884, общие длины черепов, реконструированные по размерным параметрам посткраниальных элементов. Красным цветом отмечено положение исследуемых в работе экземпляров.

Фрагмент мелкой бедренной кости (экз. ПИН, № 49/33) имеет реконструируемую длину приблизительно 80 мм, приблизительную ширину диафиза 7 мм и ширину дистального эпифиза 20 мм, что соответствует длине черепа 0.22 м (32.3% от максимального размера). Фрагмент крупной бедренной кости (экз. ПИН, № 2255/3) имеет реконструируемую длину приблизительно 120 мм, ширину диафиза 12 мм и ширину дистального эпифиза 35 мм – соответствует длине черепа 0.40 м (58.8% от максимального размера). Размеры исследуемых фрагментов бедренных костей P. stuckenbergi сопоставлялись с размерами пяти целых экземпляров данного вида. Всего было изготовлено два поперечных гистологических среза бедренных костей: несколько дистальнее середины диафиза (экз. ПИН, № 2255/3) и в основании дистального эпифиза (экз. ПИН, № 49/33).
Исследуемые гипоцентры рахитомного плана строения, типичного для пермских Temnospondyli, массивные, имеющие характерную серповидную форму. Для гистологического анализа были отобраны два гипоцентра предкрестцового отдела наименьшего и наибольшего размеров с реконструируемой шириной 25 и 35 мм. Стоит отметить, что на примере скелета P. stuckenbergi (экз. ПИН, № 164/1) с хорошо сохранившимся предкрестцовым отделом позвоночного столба, представленного 22 сочлененными элементами, можно говорить об отсутствии четкой дифференциации осевого скелета у Platyoposaurus (Конжукова, 1955), поскольку размеры позвонковых элементов (гипоцентров) колеблются в незначительных пределах: длина от 15 до 20 мм, а ширина – от 22 до 28 мм. Размеры исследуемых гипоцентров соответствуют черепам с длиной 0.33 м (48.5% от максимального размера) и 0.46 м (67.6% от максимального размера), соответственно. Всего было изготовлено по одному поперечному гистологическому срезу на уровне середины гипоцентров. Срезы наиболее мелкого (экз. ПИН, № 164/1) и наиболее крупного (экз. ПИН, № 49/21) экземпляров различаются по размеру в 1.4 раза.
Для каждого поперечного среза бедренной кости вычислялось значение кортико-диафизарного индекса (CDI), отражающего компактность кости и выраженного соотношением толщины кортекса к радиусу среза (Skutschas, Stein, 2015).
В качестве математического параметра, характеризующего особенности экологических адаптаций, применялся индекс RBT (процентное отношение среднего значения толщины кортекса к среднему значению диаметра поперечного сечения длинной кости), отражающий связь толщины кортекса с адаптацией к тому или иному образу жизни (Wall, 1983).
Для определения типа адаптаций P. stuckenbergi в дополнение к полученным гистологических данным морфология костей конечностей сравнивалась с аналогичной по Dvinosaurus на примере D. campbelli (Губин, 2004; Ульяхин и др., 2020). Выбор Dvinosaurus обусловлен тем, что для представителей этого рода надежно установлены особенности палеоэкологии с учетом явной неотеничности и облигатно-водного образа жизни.
Гистологические срезы изучались в ПИН РАН с использованием модульного стереомикроскопа Leica M165 C, а также во ВНИГНИ с использованием оптического микроскопа Axio Imager.A2m. Детальные изображения костного матрикса были получены с помощью камер Leica DFC425 и AxioCamHRc. Используемая гистологическая терминология приведена по работам: Francillon-Vieillot et al. (1990) и Danto et al. (2017).
МИКРОАНАТОМИЧЕСКОЕ И ГИСТОЛОГИЧЕСКОЕ ОПИСАНИЕ ПОСТКРАНИАЛЬНОГО СКЕЛЕТА PLATYOPOSAURUS STUCKENBERGI
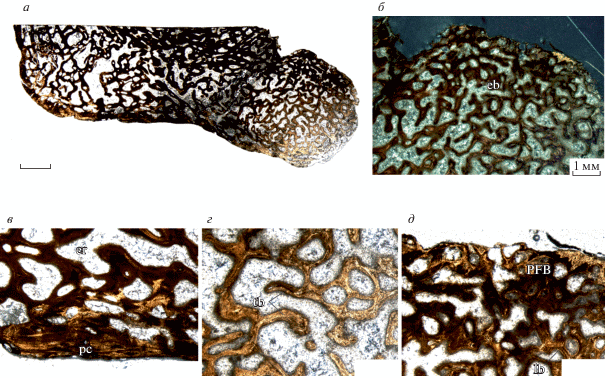
Наиболее мелкая бедренная кость в материале (экз. ПИН, № 49/33; 32.3% от максимального размера) характеризуется губчатым строением, т.е., наличием хорошо выраженной спонгиозы (рис. 3, а, б). Первичный кортекс содержит линии остановки роста (LAG’s) – около 3 (рис. 3, б, г). Внешняя часть представлена сравнительно тонким периостелиальным кортексом, сформированным первичной рыхло-волокнистой костью и содержащим многочисленные хаотично расположенные продольно-ориентированные васкулярные каналы и первичные остеоны (рис. 3, в, г). Более внутренняя часть кортекса демонстрирует значительную степень вторичных преобразований (ремоделинга), выраженную наличием крупных полостей эрозии, наличие немногочисленных вторичных остеонов (рис. 3, в, г). Медуллярная область содержит трабекулы, образованные вторичной ламеллярной костью, вторичными остеонами и первичной костной тканью во внутренней части (рис. 3, д). Лакуны кортикальных остеоцитов многочисленные, относительно небольшие, округлой или реже овальной формы. Кальцинированный хрящ отсутствует. Индекс CDI равен 0.59, индекс RBT – 26.4%.
Рис. 3.
Platyoposaurus stuckenbergi Trautschold, 1884, поперечный гистологический срез мелкой бедренной кости (экз. ПИН, № 49/33; 32.3% от максимального размера): а – общий микроанатомический вид среза; б – срез с прорисовкой ростовых меток и обозначением первичной и вторичной кости; в – кортикальная часть (в поляризованном свете); г – кортикальная часть с ростовыми метками; д – медуллярная часть. Обозначения: cx – кортекс; er – эрозионные полости; LAG’s – линии остановки роста; lb – ламеллярная кость; mc – медуллярная полость; ol – лакуны остеоцитов; pb – первичная кость; pс – периостелиальный кортекс; WFB – рыхло-волокнистая кость; sb – вторичная кость; so – вторичные остеоны; tb – трабекулы. Длина масштабной линейки 1 мм.

Бедренная кость большего размера (экз. ПИН, № 2255/3; 58.8% от максимального размера) характеризуется наличием относительно толстого кортекса, плотного внешнего (периостелиального) кортекса и медуллярной полостью, заполненной сравнительно тонкими трабекулами (рис. 4, а, б). Периостелиальный кортекс сформирован параллельно-волокнистой тканью и содержит значительное число ростовых меток: среди них можно выделить 15 линий остановки роста (LAG’s), которые располагаются ближе друг к другу во внешних частях кортекса (рис. 4, в, г). Периостелиальный кортекс средне васкуляризирован и содержит продольно и косо ориентированные васкулярные каналы, а также единичные вторичные остеоны. Васкулярные каналы сосредоточены преимущественно в области гребня аддуктора crista aspera femoris. В кортексе наблюдается усложнение васкуляризации по сравнению с меньшей по размеру бедренной костью: помимо продольных, появляются поперечные косо ориентированные сосуды в районе гребня аддуктора, а также формируется ретикулярный тип васкуляризации с неравномерным анастомозированием. Перемедуллярная область демонстрирует сильные вторичные преобразования, выраженные наличием крупных полостей эрозии и вторичных остеонов. Медуллярная полость относительно крупная, содержит сравнительно тонкие трабекулы, сформированные плотной ламеллярной костью (рис. 4, д). Лакуны кортикальных остеоцитов чаще овальной (до сильно вытянутой) и реже округлой формы. Кальцинированный хрящ также не обнаружен. Индекс CDI равен 0.46, индекс RBT – 21.9%.
Рис. 4.
Platyoposaurus stuckenbergi Trautschold, 1884, поперечный гистологический срез крупной бедренной кости (экз. ПИН, № 2255/3; 58.8% от максимального размера): а – общий микроанатомический вид среза; б – срез с обозначением первичной и вторичной кости; в – кортикальная часть (в поляризованном свете); г – кортикальная часть с многочисленными ростовыми метками; д – медуллярная часть. Обозначения: PFB – параллельно-волокнистая кость; sv – васкулярные каналы; остальные как на рис. 3. Длина масштабной линейки 2 мм.

Наиболее мелкий гипоцентр (экз. ПИН, № 164/1; 48.5% от максимального размера) имеет губчатое (“остеопорозное”) строение и характеризуется наличием сравнительно тонкого периостелиального кортекса (рис. 5, а). Периостелиальный кортекс плотный, аваскулярный, сформирован параллельно-волокнистой костью (рис. 5, б, в). Кортекс содержит многочисленные косо ориентированные шарпеевы волокна (рис. 5, б). Внутренняя часть кортекса частично резорбирована и содержит крупные полости эрозии (рис. 5, в, д). Крупные трабекулы во внутренней части гипоцентра сформированы плотной ламеллярной тканью снаружи и менее компактной параллельно-волокнистой первичной тканью внутри трабекул (рис. 5, д). Лакуны остеоцитов многочисленны, от округлой и неправильной до сильно вытянутой формы. Ростовые метки отсутствуют. Кальцинированный хрящ полностью резорбирован.
Рис. 5.
Platyoposaurus stuckenbergi Trautschold, 1884, поперечный гистологический срез мелкого гипоцентра (экз. ПИН, № 164/1; 48.5% от максимального размера): а – общий микроанатомический вид среза; б – нижняя часть с шарпеевыми волокнами; в – детали строения нижней части (в поляризованном свете); г – детали строения средней части (в поляризованном свете); д – детали строения верхней части (в поляризованном свете). Обозначения: eb – эндохондральная кость; ShF – шарпеевы волокна; tb – трабекулы губчатой кости; остальные как на рис. 3 и 4. Длина масштабной линейки 2 мм.

Гипоцентр большего размера (экз. ПИН, № 49/21; 67.6% от максимального размера) характеризуется наличием очень тонкого периостелиального кортекса, губчатым строением (рис. 6, а, б). Периостелиальный кортекс с единичными продольно ориентированными первичными сосудами сформирован параллельно-волокнистой костью (рис. 6, в). Внутренняя часть кортекса характеризуется наличием первичной спонгиозы – значительно резорбирована и содержит крупные полости эрозии (рис. 6, г, д). В кортексе шарпеевы волокна не выявлены. Крупные трабекулы во внутренней части гипоцентра сформированы плотной вторичной ламеллярной тканью в наружной части и остатками первичной параллельно-волокнистой кости во внутренней части (рис. 6, д). Лакуны остеоцитов многочисленны, чаще сильно вытянутой формы и реже округлой и неправильной формы. Ростовые метки и кальцинированный хрящ отсутствуют.
Рис. 6.
Platyoposaurus stuckenbergi Trautschold, 1884, поперечный гистологический срез крупного гипоцентра (экз. ПИН, № 49/21; 67.6% от максимального размера): а – общий микроанатомический вид среза; б – часть (в поляризованном свете); в – детали строения нижней части; г – детали строения средней части; д – детали строения верхней части. Обозначения: как на рис. 3–5. Длина масштабной линейки 2 мм.
СРАВНЕНИЕ И ОБСУЖДЕНИЕ
На основании полученных палеогистологических данных по разноразмерным посткраниальным элементам скелета P. stuckenbergi стало возможным установление возрастных различий, проявляющихся на микроанатомическом и гистологическом уровнях. Стоит отметить, что значительная разница в гистологической организации между мелкими и крупными особями не всегда может быть четко установленной у Temnospondyli, когда биомеханические и экологические особенности менялись на протяжении всей жизни, что отражалось в строении костного матрикса, как, например, у Dutuitosaurus ouazzoui и Gerrothorax sp. (Sanchez et al., 2010).
Микроанатомический и гистологический анализы поперечных срезов посткраниальных элементов Platyoposaurus stuckenbergi позволили выявить следующие общие для разноразмерных костных элементов особенности:
1) Как мелкая (32.3% от максимального размера), так и более крупная бедренная кость (58.8% от максимального размера) демонстрируют значительную степень вторичных преобразований в примедуллярной области кортекса.
2) Оба гипоцентра (48.5 и 67.6% от максимального размера соответственно) имеют остеопорозное (облегченное) строение, при этом значительных отличий в ходе онтогенеза, за исключением наличия выраженных шарпеевых волокон и меньшей площади эрозионных пустот у мелкого гипоцентра, не выявлено. Несмотря на отсутствие четкой дифференциации осевого скелета у P. stuckenbergi, нельзя исключать, что заметная разница в размере гипоцентров может быть результатом не только возрастных различий, но и индивидуальной изменчивости (Konietzko-Meier et al., 2013).
3) Важной характерной чертой всех скелетных элементов, микроанатомическое и гистологическое строение которых описывается в данном исследовании, является полное отсутствие кальцинированного хряща как в мелких, так и в более крупных костях. Данная особенность может быть объяснена резорбцией хряща на очень ранних стадиях онтогенеза, указывающая на отсутствие педоморфных черт в гистологическом строении P. stuckenbergi.
Кроме этого, были выявлены следующие характерные изменения в ходе онтогенеза:
1) периостелиальный кортекс мелкой бедренной кости (32.3% от максимального размера) содержит первичную рыхло-волокнистую костную ткань, что свидетельствует о быстром росте на ранних стадиях онтогенеза. Преобладание в кортексе крупной бедренной кости (58.8% от максимального размера) более плотной и упорядоченной параллельно-волокнистой ткани свидетельствует о замедлении темпов роста.
2) Периостелиальный кортекс мелкой бедренной кости P. stuckenbergi содержит продольно-ориентированные васкулярные каналы, в то время как бедро более крупной особи характеризуется наличием как продольных каналов, так и появлением поперечных радиальных сосудов в районе гребня аддуктора crista aspera femoris и формированием васкулярной сети ретикулярного типа в кортексе – следствие смены и усложнения ориентации сосудов в костном матриксе.
3) Ростовые метки (LAG’s) обнаружены как в бедренной кости мелкого размера, так и в более крупной, что свидетельствует о цикличности роста Platyoposaurus и смене благоприятных условий среды на неблагоприятные (сезонность климата) в течение жизни животных. При этом бедренная кость мелкой особи характеризуются наличием меньшего количества меток и, соответственно, меньшей толщиной кортекса. Сближение ростовых меток во внешней части периостелиального кортекса у более крупной бедренной кости свидетельствует о замедлении темпов роста.
Для молодых особей представителей группы Temnospondyli характерно большое количество кальцинированного хряща в костях. По мере роста происходило формирование эндохондральной кости, которая увеличивалась с одновременным уменьшением количества хряща (Konietzko-Meier et al., 2013). В отличие от Dvinosaurus и Metoposaurus, у которых хрящ присутствует как у мелких, так и у крупных особей, что может указывать на неотенический характер развития (Konietzko-Meier et al., 2013), у Platyoposaurus хрящ у исследованных экземпляров полностью отсутствует. Анализ морфологии Temnospondyli предполагает неотению для водных таксонов, таких как Dvinosaurus и Stereospondyli (Pawley, Warren, 2004), но не для Platyoposaurus, что полностью согласуется с результатами данного исследования. Необнаруженный в бедренных костях Platyoposaurus кальцинированный хрящ у мелко- и крупноразмерной особей косвенно указывает на отсутствие выраженных педоморфных черт в гистологическом строении. Выводы об отсутствии значительного числа педоморфных черт также подкрепляются данными по морфологии скелета и его отдельных элементов при сопоставлении микро- и макроанатомических данных строения костей у Dvinosaurus, неотеничность которого доказана (Шишкин, 1973). Окостенение элементов скелета Platyoposaurus было практически полным, за исключением слабо окостеневающих эпифизных частей плечевой и бедренной костей, коракоидной части скапулокоракоида, intermedium и неокостеневающих pubis и ischium (Губин, 1991). У Dvinosaurus число неокостеневающих или частично окостеневающих элементов скелета заметно больше. Отсутствие гиобранхиальных окостенений для неличиночных особей Platyoposaurus указывает на полностью легочное или частично жаберное дыхание с использованием легких. Предполагается, что у наиболее близкого к Platyoposaurus рода Archegosaurus наружные ветвистые жабры присутствовали на начальных этапах онтогенеза, после чего в процессе метаморфоза трансформировались во внутренние примитивные легкие, как у полиптеридных рыб (Witzmann, Brainerd, 2017). Данный тип дыхания нельзя исключать и для Platyoposaurus. Dvinosaurus, не проходящий полный метаморфоз вследствие неотении, сохранял жабры в течение всей жизни, что подтверждается хорошо окостеневающими элементами гиобранхиального аппарата у крупноразмерных особей (Шишкин, 1973).
Микроанатомическое и гистологическое строение рассмотренных бедренных костей P. stuckenbergi схоже с таковыми у метопозаврида Metoposaurus diagnosticus krasiejowensis (Konietzko-Meier, Klein, 2013) и Dutuitosaurus ouazzouii (Steyer et al., 2004) наличием рыхло-волокнистой ткани у молодых особей, линий остановки роста, а также формированием трабекул в медуллярной области. При этом бедренные кости P. stuckenbergi демонстрируют более губчатое строение и меньшую компактность костного матрикса, что может быть связано с другим типом экологической адаптации по сравнению с Metoposaurus и Dutuitosaurus.
Представители группы Temnospondyli занимали различные экологические ниши, включая водные, полуводные и наземные (Lindemann, 1991; Schoch, 2014). При сравнении P. stuckenbergi с неотеническим облигатно-водным Dvinosaurus campbelli был выявлен ряд морфологических отличий в строении костей конечностей. У более вытянутых бедренных костей P. stuckenbergi диафиз в сечении треугольный за счет хорошо выраженного гребня аддуктора (crista aspera femoris), который отсутствует у D. campbelli с округлым сечением диафиза. Плоскость проксимального конца относительно дистального конца у P. stuckenbergi развернута сильнее (34°–37°) (Губин, 1991), чем у D. campbelli (23°–27°). У P. stuckenbergi проксимальный эпифиз слабо окостеневший, большая часть межтрохантерной впадины оставалась хрящевой, четвертый трохантер развит слабо, представлен коротким и утолщенным гребнем (Губин, 1991). Для бедренной кости D. campbelli характерны слабо окостеневшие как проксимальный, так и дистальный эпифизы, четвертый трохантер выражен хуже, чем у P. stuckenbergi и представлен коротким, узким и более низким гребнем (Губин, 2004). Длина передней конечности P. stuckenbergi, реконструируемая по скелету из местонахождения Белебей, составляла 48% от длины позвоночного столба до крестцового отдела, длина задней конечности – 51%. Для D. campbelli по уместной аналогии с наиболее близким видом D. primus, известным по относительно полному сохранившемуся скелету (экз. ПИН, № 2005/41), данные показатели будут 20 и 22% соответственно. Исходя из приведенного морфологического сравнения бедренных костей, а также относительной длины реконструируемых конечностей, у P. stuckenbergi последние были длиннее относительно длины туловища и лучше развиты, чем у D. campbelli. Это может указывать на их опорно-локомоторную функцию в условиях спорадического пребывания в сухопутной обстановке, как это предполагается для Archegosaurus (Witzmann, Brainerd, 2017).
Сохранение плотного и сильно минерализованного хряща в эндохондральной части костей у водных Temnospondyli могло играть роль дополнительного балласта для контроля плавучести при придонном образе жизни (de Buffrénil, Mazin, 1990; Danto et al., 2017). Отсутствие в бедренных костях как у мелко- так и у крупноразмерных особей P. stuckenbergi кальцинированного хряща и остеопорозное строение позвонков не свидетельствует в пользу упомянутого типа адаптации. Однако длинные кости вполне придонных представителей семейства Plagiosauridae и рода Plagiosuchus в частности несут явные признаки остеопороза (Konietzko-Meier, Schmitt, 2013). Схожесть Platyoposarus с Plagiosuchus проявляется в том, что на брюхе они имели сплошной покров (Конжукова, 1955; Губин, 1991; Witzmann, Soler-Gijón, 2010), состоящий из костных кожных чешуй (брюшной панцирь), добавляющих вес к общей костной массе, что могло быть дополнительным приспособлением к придонному образу жизни (Sanchez, Schoch, 2013). Подобная адаптация сходна с таковой у современных черепах или вымерших плакодонтов (Witzmann, Soler-Gijón, 2010; Witzmann, 2011).
Отдельно рассматриваемые микро- или макроанатомические параметры костей скелета не всегда могут надежно указывать на тот или иной тип адаптации. Так, для Eryops на основании изучения костей конечности адаптация была определена как полуназемная (Pawley, Warren, 2006). Однако губчатое строение его гипоцентров указывает на преимущественно водный образ жизни (Sanchez et al., 2010).
Исследования толщины кортекса бедренных костей у нескольких современных крокодилов, ящериц и млекопитающих, а также ископаемых терапсид показали, что для большинства исследованных животных с полуводным типом адаптации индекс RBT близок к 30% или больше (Magwene, 1993). Значение RBT в 30% считается пороговым для определения особенностей экологии. Для животных, у которых индекс RBT для костей конечностей превышает 30%, предполагается водный образ жизни (Mukherjee et al., 2010). У P. stuckenbergi индекс RBT бедренной кости (составляющий 22–26%) очень близок по значению к таковому у раннетриасовых Trematosauridae (Mukherjee et al., 2010). Для сравнения, у облигатно-водных D. campbelli индекс RBT бедренной кости составляет 27.2%, что сопоставимо с таковым для мелкой бедренной кости P. stuckenbergi (RBT = 26.4%). Стоит отметить, что индекс RBT мелкой бедренной кости P. stuckenbergi на 4.5% больше, чем для крупной бедренной кости, что может указывать на изменение типа адаптации с возрастом от облигатно-водного образа жизни в сторону полуназемного.
Компактность (CDI) бедренной кости P. stuckenbergi с возрастом закономерно уменьшалась за счет резорбции внутренних частей кортекса, причем для мелкого экземпляра показатель CDI равен 0.59, что близко по значению к таковому для крупной бедренной кости D. campbelli (CDI = 0.54). Высокая пористость и низкая компактность костного матрикса для крупной бедренной кости P. stuckenbergi указывает на низкое биомеханическое сопротивление (Teschner et al., 2018) и меньшую устойчивость к скручивающей нагрузке (Sanchez et al., 2010). Все перечисленные параметры могут указывать скорее на водную адаптацию P. stuckenbergi как активно плавающего, нежели придонного хищника. Конечности при передвижении в воде, по всей видимости, мало использовались. У близкого родственника P. stuckenbergi, Archegosaurus decheni, был хорошо развитый хвост (длиннее остальной части тела), активно используемый при плавании (Witzmann, Schoch, 2006). Не полностью окостеневающие кости конечностей P. stuckenbergi, как педоморфная черта, оставались слаборазвитыми как у молодых, так и у взрослых особей даже по сравнению с вполне водным стереоспондиломорфом Australerpeton, у которого окостенение было более полным (Dias, Schultz, 2003).
В качестве дополнительной информации в разрешении вопроса об особенностях экологической адаптации P. stuckenbergi были привлечены данные по литологии костеносных пород тех местонахождений, откуда известны все достоверные находки Platyoposaurus. Остатки Platyoposaurus примерно в равной степени приурочены к песчано-гравийным литофациям русел и глинисто-карбонатным литофациям озерных водоемов. В шести местонахождениях остатки приурочены к русловым песчаникам; в одном – к гравелитам и конгломератам временных и быстрых потоков; в восьми – к мергелям, алевролитам и глинам непроточных опресненным водоемов; в одном – к известнякам непроточных слабосоленых (?) водоемов (табл. 1). Это может указывать на большую экологическую пластичность по сравнению с архегозавроидами семейства Melosauridae, остатки которых чаще всего встречаются в русловых отложениях. Для сравнения, остатки других архегозаврид найдены в следующих типах отложений: Archegosaurus – в озерных и речных (Geinitz, Deichmüller, 1882; Witzmann, Brainerd, 2017); Bashkirosaurus – в речных (Новожилов, 1955; Губин, 1981), Collidosuchus – в речных (Ефремов, Вьюшков, 1955; Губин, 1986); Prionosuchus – в озерных (Iannuzzi et al., 2018). Остатки как мелко-, так и крупноразмерных Platyoposaurus одинаково часто встречаются и в речных, и в озерных отложениях, т.е., между разноразмерными (разновозрастными) особями нет четкой зависимости от типа водного объекта (река, озеро).
Таблица 1.
Приуроченность представителей Platyoposaurus к различным типам отложений с указанием количества местонахождений для тех или иных литофаций
| Тип литофаций | Число местонахождений с достоверно известными находками Platyoposaurus |
|---|---|
| Русловые песчаники | 6 |
| Гравелиты и конгломераты временных и быстрых потоков | 1 |
| Мергели, алевролиты и глины непроточных опресненных водоемов | 8 |
| Известняки непроточных слабосоленых водоемов | 1 |
Сезонные климатические колебания находят отражение у экзотермических животных в характере циклического роста костей, который устанавливается по видимым границам, представленным чередующимися слоями медленно и быстро растущей костной ткани, а также линиями остановок роста (LAG’s), маркирующими прекращение остеогенеза (Konietzko-Meier, Klein, 2013). Для зависимых от водной среды обитания животных, таких как Platyoposaurus, неблагоприятным временем года являлся сухой сезон, характеризующийся повышенной температурой и снижением количества осадков, что приводило к сокращению площадей обитания и кормления. Непостоянная толщина промежутков между LAG’s в глубоких слоях кортекса у P. stuckenbergi, скорее всего, указывает на неодинаковую продолжительность благоприятных сезонов. На протяжении жизни Platyoposaurus неблагоприятные периоды, в течение которых животное испытывало физиологический стресс, наблюдались очень часто, что подтверждается многочисленными ростовыми метками. Физиологический стресс мог быть вызван, в частности, конкуренцией за пищевые ресурсы.
Конкурентами для Platyoposaurus в водном блоке хищных среднепермских тетрапод Восточной Европы были Melosauridae. Так, остатки Platyoposaurus достоверно известны в 15 ориктокомплексах, а Melosauridae – в 18, причем только в шести из них установлено их совместное нахождение. При этом наблюдается зависимость – если из местонахождения известно много остатков Platyoposaurus, то мало Melosauridae, и наоборот, – что может указывать на высокую конкуренцию этих архегозавроидов внутри водных биотопов.
Скелетохронологические данные по бедренным костям P. stuckenbergi ставят под сомнение заключение Е.Д. Конжуковой (1964) о том, что архегозавроиды могли доживать до 300 лет. Это заключение было сделано по результатам изучения черепа крупноразмерной особи Melosaurus uralensis из местонахождения Мамадыш Кировской обл. (экз. ПИН, № 683/1), который сопоставим со среднеразмерным черепом P. stuckenbergi. В отношении рассматриваемой группы среднепермских Temnospondyli скорее уместно говорить о десятках лет, чему не противоречат данные по скелетохронологии P. stuckenbergi с подсчитанным количеством LAG’s в количестве не менее 15 для крупной бедренной кости изученного материала.
ЗАКЛЮЧЕНИЕ
Микроанатомическое и гистологическое исследования разноразмерных посткраниальных элементов скелета (бедренные кости, гипоцентры) P. stuckenbergi позволили установить ряд возрастных различий, позволяющих оценить степень онтогенетической зрелости мелких и крупных экземпляров. При этом учитывался тот факт, что разница в гистологической организации между мелкими и крупными особями не всегда бывает явной по причине биомеханических и экологических изменений на протяжении всей жизни. Характерные для многих представителей группы Temnospondyli педоморфные черты, например, наличие кальцинированного хряща, у P. stuckenbergi по гистологических данным выявлены не были, при том, что некоторые части костей и отдельные элементы скелета не подвергались окостенению. Это может объясняться резорбцией хряща на ранних стадиях онтогенеза, когда происходил быстрый рост, замедляющийся уже при достижении размеров, близких к 60% от максимальных значений. Комплекс полученных данных позволяет говорить о P. stuckenbergi как о достаточно подвижном водном хищнике, который при передвижении в большей степени использовал хвост, нежели конечности. P. stuckenbergi с возрастом, вероятно, мог изменять тип адаптации от облигатно-водного образа жизни в сторону полуназемного, что разительно отличает его от неотенического Dvinosaurus – засадного хищника, не способного покидать водную среду. В качестве биотопов для Platyoposaurus примерно в равной степени выступали реки и озера, что очень схоже с Archegosaurus, и может указывать на значительную экологическую пластичность представителей Archegosauridae в целом. Скелетохронологические данные о количестве подсчитанных линий остановки роста (N = 15) у P. stuckenbergi дают совершенно новое представление о возрасте и опровергают ранее высказанное предположение о нескольких сотнях лет жизни архегозавроидов.
Список литературы
Быстров А.П. Стегоцефалы как показатели климата // Вопросы палеобиогеографии и биостратиграфии. М.: Госгеолтехиздат, 1957. С. 147–153.
Вьюшков Б.П. О фауне верхнепермских наземных позвоночных с реки Малая Кинель. М.: Изд-во АН СССР, 1955. 177 с.
Голубев В.К. Наземные позвоночные // Стратотипы и опорные разрезы верхней перми Поволжья и Прикамья. Казань: Экоцентр, 1996. С. 381–389.
Губин Ю.М. Новый платиопозаврид из Башкирии // Палеонтол. журн. 1981. № 2. С. 141–143.
Губин Ю.М. Новые данные об архегозавроидах Восточно-Европейской платформы // Палеонтол. журн. 1986. № 2. С. 75–80.
Губин Ю.М. О систематическом положении лабиринтодонтов из местонахождения Малая Кинель // Палеонтол. журн. 1989. № 4. С. 116–120.
Губин Ю.М. Пермские архегозавроидные амфибии СССР. М.: Наука, 1991. 140 с. (Тр. Палеонтол. ин-та АН СССР. Т. 249).
Губин Ю.М. Новый двинозавр (Amphibia: Temnospondyli) из верхнетатарских отложений Среднего Поволжья // Палеонтол. журн. 2004. № 2. С. 72–80.
Ефремов И.А., Вьюшков Б.П. Каталог местонахождений пермских и триасовых наземных позвоночных на территории СССР. М.; Л.: Изд-во АН СССР, 1955. 186 с. (Тр. Палеонтол. ин-та АН СССР. Т. 46).
Ивахненко М.Ф. Тетраподы Восточно-Европейского плакката – позднепалеозойского территориально-природного комплекса. Пермь: Пермский обл. краевед. муз., 2001. 200 с. (Тр. Палеонтол. ин-та РАН. Т. 283).
Ивахненко М.Ф., Голубев В.К., Губин Ю.М. и др. Пермские и триасовые тетраподы Восточной Европы. М.: Наука, 1997. 216 с. (Тр. Палеонтол. ин-та РАН. Т. 268).
Конжукова Е.Д. Platyops stuсkеnbergi Trautsch. – архегозавроидный лабиринтодонт нижних зон верхней перми Приуралья // Материалы по пермским и триасовым наземным позвоночным СССР. М.: Изд-во АН СССР, 1955. С. 89–127.
Конжукова Е.Д. Старческая особь мелозавра из верхней перми р. Вятка // Палеонтол. журн. 1964. № 4. С. 122–126.
Новожилов Н.И. Белебеевское местонахождение пермских Tetrapoda // Материалы по пермским и триасовым наземным позвоночным СССР. М.: Изд-во АН СССР, 1955. С. 190–200.
Ульяхин А.В., Скучас П.П., Сабуров П.Г. Гистология Dvinosaurus campbelli (Temnospondyli, Dvinosauria) из позднепермского местонахождения Гороховец, Владимирская область // Палеонтол. журн. 2020. № 6. С. 74–82.
Шишкин М.А. Морфология древних земноводных и проблемы эволюции низших тетрапод. М.: Наука, 1973. 260 с. (Тр. Палеонтол. ин-та АН СССР. Т. 137).
Botha J., Chinsamy A. Growth and life habits of the Triassic cynodont Trirachodon, inferred from bone histology // Acta Palaeontol. Pol. 2004. V. 49. P. 619–627.
Bystrow A.P. Zahnstruktur der Labyrinthodonten // Acta Zool. 1938. V. 19. P. 387–425.
Danto M., Witzmann F., Pierce S.E., Fröbisch N.B. Intercentrum versus pleurocentrum growth in early tetrapods: A paleohistological approach // J. Morphol. 2017. V. 278. P. 1–22.
de Buffrénil V., Mazin J.M. Bone histology of the Ichthyosaurs: comparative data and functional interpretation // Paleobiology. 1990. V. 16. P. 435–447.
de Ricqlès A., de Buffrénil V. Bone histology, heterochronies and the return of tetrapods to life in water: where are we // Secondary adaptation of tetrapods to life in water / Eds. Mazin J., de Buffrénil V. München: Verlag Dr. F. Pfeil, 2001. P. 289–310.
Dias E.V., Schultz C.L. The first Paleozoic temnospondyl postcranial skeleton from South America // Rev. Brasil. Paleontol. 2003. V. 6. P. 29–42.
Efremov I.A. Peгmischen Labyrinthodonten des fruheren Gouvernements Wjatka // Uber die Labyrinthodonten der UdSSR. M.: Изд-вo AH CCCP, 1932. C. 117–164.
Francillon-Vieillot H., de Buffrénil V., Castanet J. et al. Microstructure and mineralization of vertebrate skeletal tissues // Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends / Ed. Carter J. N.Y.: Van Nostrand Reinhold, 1990. P. 471–530.
Gee B.M., Haridy Y., Reisz R.R. Histological skeletochronology indicates developmental plasticity in the early Permian stem lissamphibian Doleserpeton annectens // Ecol. Evol. 2020. V. 10. P. 1–17.
Geinitz H.B., Deichmüller J.V. Die Saurier der unteren Dyas von Sachsen // Palaeontogr. 1882. Bd 9. S. 1–46.
Gubin Y.M. Skull morphology of Archegosaurus decheni Goldfuss (Amphibia, Temnospondyli) from the Early Permian of Germany // Alcheringa. 1997. V. 21. P. 103–121.
Iannuzzi R., Neregato R., Cisneros J.C. et al. Re-evaluation of the Permian macrofossils from the Parnaíba Basin: biostratigraphic, palaeoenvironmental and palaeogeographical implications // Geol. Soc. L. Spec. Publ. 2018. V. 472. P. 223–249.
Konietzko-Meier D., Bodzioch A., Sander P.M. Histological characteristics of the vertebral intercentra of Metoposaurus diagnosticus (Temnospondyli) from the Upper Triassic of Krasiejów (Upper Silesia, Poland) // Trans. R. Soc. Edinburgh. Earth Envir. Sci. 2013. V. 103. P. 1–14.
Konietzko-Meier D., Klein N. Unique growth pattern of Metoposaurus diagnosticus krasiejowensis (Amphibia, Temnospondyli) from the Upper Triassic of Krasiejów, Poland // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2013. V. 370. P. 145–157.
Konietzko-Meier D., Schmitt A. A histological study of a femur of Plagiosuchus, a Middle Triassic temnospondyl amphibian from southern Germany, using thin sections and micro-CT scanning // Neth. J. Geosci. 2013. V. 92. P. 65–71.
Kriloff A., Germain D., Canoville A. et al. Evolution of bone microanatomy of the tetrapod tibia and its use in palaeobiological inference // J. Evol. Biol. 2008. V. 21. P. 807–826.
Laurin M., Girondot M., Loth M.-M. The evolution of long bone microanatomy and lifestyle in lissamphibians // Paleobiology. 2004. V. 30. P. 589–613.
Lindemann F.J. Temnospondyls and the Lower Triassic paleogeography of Spitsbergen // Fifth Symp. on Mesozoic Terrestrial Ecosystems and Biota / Eds. Kielan-Jaworowska Z., Heintz N., Naktrem H.A. Oslo, 1991. P. 39–40 (Contrib. Paleontol. Mus. Univ. Oslo. V. 364).
Magwene P.M. What’s bred in the bone: histology and cross-sectional geometry of mammal-like reptile long bones–evidence of changing physiological and biomechanical demands. MSc thesis. Harvard Univ., 1993.
Mukherjee D., Ray S., Sengupta D.P. Preliminary observations on the bone microstructure, growth patterns, and life habits of some Triassic temnospondyls from India // J. Vertebr. Paleontol. 2010. V. 30. P. 78–93.
Pawley K., Warren A.A. Immaturity vs. paedomorphism: a rhinesuchid stereospondyl postcranium from the Upper Permian of South Africa // Palaeontol. Afr. 2004. V. 40. P. 1–10.
Pawley K., Warren A.A. The appendicular skeleton of Eryops megacephalus (Temnospondyli: Eryopoidea) from the Lower Permian of North America // J. Paleontol. 2006. V. 80. P. 561–580.
Price L.I. Um anfibio labirintodonte da formacao Pedra de Fogo, estado do Maranhao // Bol. Div. Geol. Mineral. 1948. V. 124. P. 1–32.
Ray S., Chinsamy A. Diictodon feliceps (Therapsida, Dicynodontia): bone histology, growth and biomechanics // J. Vertebr. Paleontol. 2004. V. 24. P. 180–194.
Ray S., Chinsamy A., Bandyopadhyay S. Lystrosaurus murrayi (Therapsida Dicynodontia): bone histology, growth and lifestyle adaptations // Palaeontology. 2005. V. 48. P. 1169–1185.
Sanchez S., Germain D., de Ricqlès A. et al. Limb-bone histology of temnospondyls: implications for understanding the diversification of palaeoecologies and patterns of locomotion of Permo-Triassic tetrapods // J. Evol. Biol. 2010. V. 23. P. 2076–2090.
Sanchez S., Schoch R.R. Bone histology reveals a high environmental and metabolic plasticity as a successful evolutionary strategy in a long-lived homeostatic temnospondyl // Evol. Biol. 2013. V. 40. P. 627–647.
Schoch R.R. Amphibian evolution: the life of early land vertebrates. Chichester: Wiley Blackwell, 2014. 276 p.
Schoch R.R., Milner A.R. Stereospondyli // Encyclopedia of Paleoherpetology. Pt 3B. München: Pfeil, 2000. 203 p.
Skutschas P.P., Stein K. Long bone histology of the stem salamander Kokartus honorarius (Amphibia: Caudata) from the Middle Jurassic of Kyrgyzstan // J. Anat. 2015. V. 226. P. 334–347.
Steyer J.S., Laurin M., Castanet J., de Ricqlès A. First histological and skeleto-chronological data on temnospondyl growth: palaeoecological and palaeoclimatological implications // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2004. V. 206. P. 193–201.
Teschner E., Sander P.M., Konietzko-Meier D. Variability of growth pattern observed in Metoposaurus krasiejowensis humeri and its biological meaning // J. Iber. Geol. 2018. V. 44. P. 99–111.
Trautschold H. Die Reste Permischer Reptilien des Paläontologischen Kabinets der Universität Kasan. Moscou: Impr. de l’Univ. Impériale, 1884. 38 p.
Twelvetrees W.H. On a labyrinthodont skull (Platyops rickardi, Twelvetr.) from the Upper Permian cupriferous strata of Kargala, near Orenburg // Bull. Soc. Natur. M. 1880. V. 55. P. 117–122.
Wall W.P. The correlation between high limb–bone density and aquatic habits in recent mammals // J. Paleontol. 1983. V. 57. P. 197–207.
Witzmann F. Morphological and histological changes of dermal scales during the fish-to-tetrapod transition // Acta Zool. 2011. V. 92. P. 281–302.
Witzmann F., Brainerd E. Modeling the physiology of the aquatic temnospondyl Archegosaurus decheni from the early Permian of Germany // Foss. Rec. 2017. V. 20. P. 105–127.
Witzmann F., Schoch R.R. The postcranium of Archegosaurus decheni, and a phylogenetic analysis of temnospondyl postcrania // Palaeontology. 2006. V. 49. P. 1211–1235.
Witzmann F., Soler-Gijón R. The bone histology of osteoderms in temnospondyl amphibians and in the chroniosuchian Bystrowiella // Acta Zool. 2010. V. 91. P. 96–114.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал