

Палеонтологический журнал, 2021, № 4, стр. 18-23
Первые находки двустворчатых моллюсков рода Unionites Wissmann в бореальной перми Северо-Востока России
А. С. Бяков a, b, *, Р. В. Кутыгин c
a Северо-Восточный комплексный научно-исследовательский институт им. Н.А. Шило ДВО РАН
Магадан, Россия
b Казанский (Приволжский) федеральный университет
Казань, Россия
c Институт геологии алмаза и благородных металлов СО РАН
Якутск, Россия
* E-mail: abiakov@mail.ru
Поступила в редакцию 13.10.2020
После доработки 19.10.2020
Принята к публикации 19.10.2020
Аннотация
Впервые в верхнепермских отложениях Северо-Восточной России найдены представители двустворчатых моллюсков рода Unionites Wissmann совместно с типично высокобореальными иноцерамоподобными двустворками Intomodesma sp. Ранее унионитесы считались исключительно триасово-раннеюрскими и лишь недавно были обнаружены в самых верхах перми Италии. Находка представителей рода Unionites в бореальной верхней перми свидетельствует о том, что к концу пермского периода усилились связи высокобореальных бассейнов и надобласти Тетис, наметившиеся еще в среднем и позднем вучапине. Описан новый вид U. kobyumensis Biakov, sp. nov.
Двустворчатые моллюски рода Unionites – морских униоподобных представителей семейства Trigonodoidae Modell – долгое время считались исключительно триасовыми (Невесская и др., 2013). Они довольно часто встречаются в отложениях триаса, как бореальных (Spath, 1935; Дагис, Корчинская, 1987; Foster et al., 2017), так и тетических бассейнов (Кипарисова, Курбатов, 1952; Ciriacks, 1963; Hofmann et al., 2013, 2015; Chu et al., 2016; Feng et al., 2019; и др.), а также Панталассы (Hofmann et al., 2014). Лишь недавно унионитесы были отмечены в самых верхах пермского разреза (верхи чансинского яруса) Италии (Farabegoli et al., 2007; Posenato, 2009).
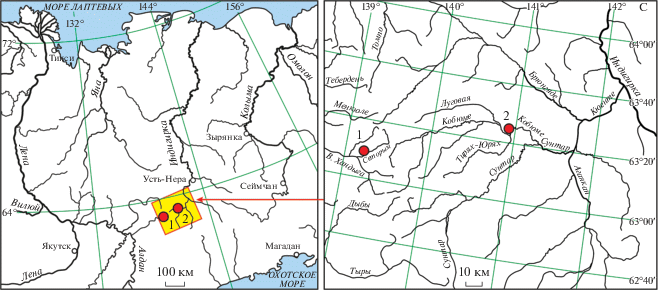
В России унионитесы ранее также были известны исключительно из триасовых отложений Северо-Востока России (Дагис и др., 1996) и Приморья (Кипарисова, 1938). В 2017 г. при изучении пограничных отложений перми и триаса Южного Верхоянья (разрез по руч. Правый Суол, правый приток р. Сеторым, бассейн р. Восточной Хандыги; рис. 1) нами впервые был найден представитель этого рода – Unionites cf. canalensis (Catullo) в нижней части отоцерасовых слоев (зона Otoceras concavum), ранее относимых к низам триаса (Дагис и др., 1996), а в настоящее время рассматриваемых как верхи чансинского яруса верхней перми (Бяков и др., 2018).
Рис. 1.
Местонахождения двустворчатых моллюсков рода Unionites на Северо-Востоке России: 1 – басс. р. Сеторым, верховья р. Восточной Хандыги, 2 – р. Кобюме.
Однако недавно У. Фостер с соавт., изучая нижнеиндских двустворок Шпицбергена, высказали мнение, что вид U. canalensis может быть отнесен к миоцен–cовременному роду нукуланоидных двустворок Austrotindaria (Foster et al., 2017). Это предположение основано на отсутствии данных о внутреннем строении раковины у типовых экземпляров Unionites canalensis и схожести, по Фостеру, этих форм с австротиндариями. В частности, по Фостеру, для этого вида характерно центральное (медианное) положение макушек и их ортогирность (Foster, 2017, c. 861).
В то же время, простое внешнее сравнение типичных Unionites canalensis с представителями рода Austrotindaria, в том числе, описанными самим Фостером, на наш взгляд, заставляют усомниться в правильности отнесения этого вида к австротиндариям. Во-первых, у многих ранее описанных представителей Unionites canalensis макушка не центральная и ортогирная, а явно сдвинута к переднему краю и прозогирная (см., напр., Ciriaks, 1963; Kumagae, Nakazawa, 2009; Song et al., 2019); во-вторых, она не такая миниатюрная, как у типичных австротиндарий, а довольно массивная. Поэтому мы оставляем данный вид в составе рода Unionites, в то же время признавая определенную условность этой принадлежности, поскольку, действительно, данных о внутреннем строении этого вида почти нет.
В 2016 г. при изучении другого разреза пермских и пограничных пермо–триасовых отложений Южного Верхоянья – по р. Кобюме, левому притоку р. Сунтар верховьев р. Индигирки (рис. 1), также были найдены представители рода Unionites. Но здесь они были встречены в более древних отложениях – в верхах зоны Intomodesma costatum (Бяков, 2012) в одном слое с типично высокобореальной гигантской иноцерамоподобной двустворкой Intomodesma sp. Согласно последним данным (Бяков и др., 2017), верхи зоны Intomodesma costatum примерно соответствуют верхнему чансину без самых его верхов.
Кобюминские унионитесы были встречены в верхах привольнинской свиты, которая относится к интервалу от кепитенского яруса (верхняя часть) до чансинского, включительно (Budnikov et al., 2020). Терминальный слой 33 привольнинской свиты (рис. 2) характеризуется отчетливым трансгрессивно-регрессивным строением (Кутыгин и др., 2019) и делится на три части, литологические переходы между которыми неотчетливые.
Рис. 2.
Геологическая схема участка и фрагмент разреза Тирях-Кобюме, включающий уровень находки двустворчатых моллюсков рода Unionites, обр. 3и/16-33(803): 1–5 – свиты: 1 – кобюминская, 2 – тиряхская, 3 – луговская, 4 – привольнинская, 5 – некучанская; 6 – оленекские отложения; 7 – анизийские отложения; 8 – четвертичные отложения; 9 – разрез Тирях-Кобюме; 10 – алевролиты мелкозернистые; 11 – алевролиты крупнозернистые; 12 – песчаники: a – слоистые, b – массивные; 13 – туфы (внемасштабно); 14 – скопления обломков призматического слоя двустворок (внемасштабно); 15 – диамиктиты; 16 – песчаные знаки ряби; 17 – следы деятельности илоедов; 18 – карбонатно-глинистые и карбонатно-кремнистые конкреции; 19 – двустворчатые моллюски; 20 – аммоноидеи; 3и/16-33(803): Unionites kobyumaensis Biakov, sp. nov., Palaeoneilo? vel Palaeonucula?? sp. indet.; 3и/16(19)-33(818): Intomodesma sp.; 19R3-34-1p: Otoceras aff. gracile Tozer, Otoceras sp., Palaeonucula? sp. indet.; 16RK20: Otoceras boreale Spath; 19R3-35-6p: Tompophiceras? sp.; 19R3-35-16p: Tompophiceras morpheos (Popow), Palaeonucula aldanenssis Kurushin, Nuculopsis setorymensis Kurushin; 19R3-35-19p: Tompophiceras sp., Maitaia ex gr. errabunda (Popow).
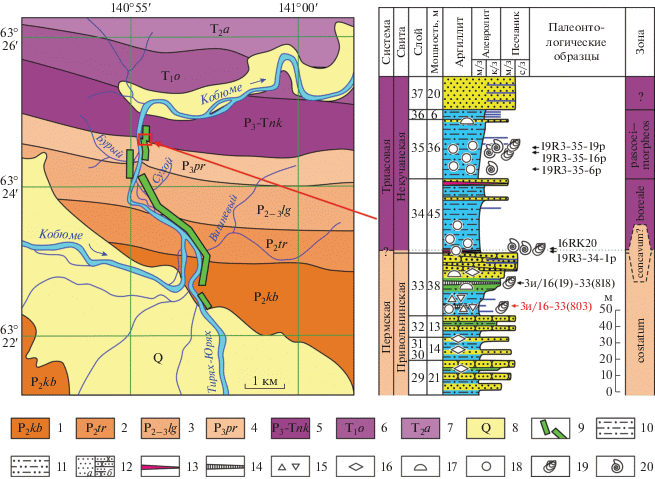
Нижняя, трансгрессивная часть (15 м) слоя 33 сложена неяснослоистыми, монотонными алевролитами, плавно огрубляющимися вверх по разрезу. В наиболее тонкозернистых нижних девяти–десяти метрах слоя наблюдаются рассеянные окатанные обломки кварцитов (размером до 1–2 см, реже – до 3 см) и многочисленные тонкие (первые сантиметры) линзочки, заполненные раздробленными остатками призматического слоя иноцерамоподобных двустворок. В 6 м выше подошвы слоя 33 обнаружены остатки трех раковин унионитесов, которые обладают морфологическим сходством с U. fassaensis (Wissmann) и описаны в настоящей статье как новый вид U. kobyumensis Biakov. На этом же уровне были обнаружены плохо сохранившиеся нукулиды Nuculopsis aff. wymensis (Keyserling), Palaeoneilo? vel Palaeonucula?? sp. indet. и очень мелкая неопределимая небеллерофонтидная гастропода.
Средняя часть (8 м) слоя 33 сложена горизонтально-слоистыми крупнозернистыми алевролитами с прослоями (5–10, реже до 20 см) глинистых песчаников и мелкозернистых алевролитов. Для этой части разреза характерны наиболее мощные (до 10–15 см) слойки с обломками призматического слоя иноцерамоподобных двустворок. В одном из таких слойков, на уровне в 20 м выше подошвы слоя 33 была обнаружена гигантская раковина Intomodesma sp.
Доминирующую роль в строении верхней, регрессивной части (15 м) слоя 33 приобретают разнозернистые песчаники. Разрез начинается с полого-волнистого переслаивания разнозернистых алевролитов, алевропесчаников и глинистых биотурбированных песчаников, а завершается относительно мощной толщей песчаников. Нижняя половина этой толщи имеет горизонтальную слоистость и знаки ряби на поверхностях напластования, а верхняя – является наиболее грубозернистой и характеризуется массивной, иногда крупной косослоистой текстурой.
Выше слоя 33 залегает мощная толща (слои 34 и 35) мелкозернистых алевролитов нижненекучанской подсвиты, в которой ранее по аммоноидеям была выявлена следующая последовательность зон (снизу вверх): Otoceras concavum?, O. boreale, Tompophiceras pascoei–T. morpheos (Кутыгин и др., 2019). Здесь, по аналогии с эталонным разрезом нижненекучанской подсвиты в бассейне р. Сеторым (Ядренкин и др., 2020), положение границы перми и триаса нами предполагается в основании зоны Otoceras boreale. Соответственно, слой 33, содержащий унионитесов и интомодесм, и основание слоя 34 с ранними отоцерасами относятся к терминальной части пермской системы.
Находка рода Unionites вместе с высокобореальными иноцерамоподобными двустворками свидетельствует о том, что к концу перми усилились связи бассейнов Северо-Восточной Азии и надобласти Тетис, наметившиеся еще в среднем и позднем вучапине, когда в сообществах двустворок Северо-Восточной Азии появились теплолюбивые Guizhoupecten, Cyrtorostra и Obliquipecten (Муромцева, Гуськов, 1984; Бяков, 2020) и произошла инвазия аммоноидей Paramexicoceras и двустворчатых моллюсков Atomodesma s.s. (Бяков, Кутыгин, 2020). Это проникновение может объясняться как некоторым потеплением климата, так и существенным ослаблением изолирующего значения Охотско-Тайгоносской вулканической дуги, игравшей роль крупного биогеографического барьера на протяжении почти всей пермской истории (Biakov, Shi, 2010).
Таким образом, в результате этой находки впервые получены данные о присутствии унионитесов в заведомо пермских отложениях северо-востока России. Ее описанию и посвящена настоящая статья.
В описании приняты сокращения: В – высота раковины, Вп – выпуклость створки, Д – длина раковины, ДЗК – длина замочного края раковины, ДПЧ – длина передней части створки.
Изученный материал хранится в музее Северо-Восточного комплексного научно-исследовательского ин-та им. Н.А. Шило ДВО РАН (СВКНИИ), г. Магадан, колл. № 02-06.113. Авторы признательны И.В. Будникову, В.И. Давыдову и А.Н. Килясову за помощь при проведении полевых работ.
Исследования выполнены по государственным заданиям СВКНИИ ДВО РАН и ИГАБМ СО РАН при поддержке грантов РФФИ №№ 18-05-00191, 20-05-00604, гранта РНФ № 19-17-00178 (палеогеографические и палеоклиматические выводы) и частично за счет средств субсидии, выделенной Казанскому федеральному университету для выполнения государственного задания № 671-2020-0049 в сфере научной деятельности.
Авторы признательны С.В. Попову и анонимному рецензенту, чьи замечания способствовали улучшению текста статьи.
ОТРЯД UNIONIDA
НАДСЕМЕЙСТВО TRIGONODOIDEA MODELL, 1942
СЕМЕЙСТВО TRIGONODOIDAE MODELL, 1942
Род Unionites Wissmann, 1841
Unionites kobyumensis Biakov, sp. nov.
Название вида – по р. Кобюме.
Голотип – СВКНИИ, № 1/02-06.113, ядро правой створки; Южное Верхоянье, правобережье р. Кобюме в нижнем течении; верхи привольнинской свиты; верхняя пермь, верхняя часть чансинского яруса, хальпирский горизонт, верхняя часть бивальвиевой зоны Intomodesma costatum; обн. 3и, нижняя часть слоя 33; сборы Р.В. Кутыгина, 2016 г.
Описание (рис. 3, б–д). Раковина небольших размеров для рода, чуть более 20 мм в длину, слабо удлиненная (Д : В = 1.47–1.67), треугольно-овальная, слабо неравносторонняя (ДПЧ : Д = = 0.40–0.43); раковинный слой был, по-видимому, относительно тонким, до 0.3 мм. Замочный край длинный (ДЗК : Д = 0.86–0.87), его передняя ветвь в значительной степени вогнута, несколько короче задней, соединяется с ней под тупым углом порядка 110°–120°. Задняя ветвь замочного края слабовыпуклая, длинная, плавно соединяется с коротким, сильно выпуклым задним краем, по крутой дуге переходящим в длинный, слабовыпуклый нижний край. Последний по крутой дуге постепенно переходит в короткий, сильновыпуклый передний край, плавно смыкающийся с передней ветвью замочного края. Выпуклость створок умеренная (Вп : В = 0.28), точка наибольшей выпуклости расположена в верхней части створок и несколько сдвинута к передней ветви замочного края. Макушки относительно небольшие, невысокие, притупленные, направленные вперед. Килевой перегиб не выражен.
Рис. 3.
Двустворчатые моллюски рода Unionites из верхнепермских отложений Северо-Востока России: а – U.? cf. canalensis (Catullo), экз. № 1/02-06.111, ядро левой створки; правобережье р. Сеторым, руч. Левый Суол; верхняя пермь, нижняя часть некучанской свиты, верхи чансинского яруса, обр. 9–2.8/АБ-17; б–д – U. kobyumensis Biakov, sp. nov.: б – голотип № 2/02-06.113, ядро правой створки; в, г – экз. № 1/02-06.113: в – ядро правой створки, г – отпечаток этой же створки; д – экз. № 3/02-06.113, ядро двустворчатого экземпляра со стороны левой створки; правобережье нижнего течения р. Кобюме; верхняя пермь, верхняя часть чансинского яруса, обр. 3и/16-33(803). Длина масштабной линейки 10 мм.

Замок, по-видимому, типичен для рода. Кардинальные зубы, по-видимому, редуцированы. На отпечатке экз. № 1/02-06.113 наблюдается слабозаметная вытянутая ямка от заднего бокового зуба (рис. 3, г), у голотипа сохранился один небольшой удлиненный передний боковой зуб (рис. 3, б).
Скульптура створок из частых, тонких, относительно правильных концентрических линий нарастания, сохраняющихся на ядрах раковин. Мускульные отпечатки относительно небольшие, удлиненно-овальные, передний расположен вблизи места смыкания переднего и замочного краев, задний по форме и размерам аналогичен переднему, тот и другой очень слабо заметны на ядрах раковин. Мантийная линия цельная. Остальные элементы строения раковины не наблюдались.
Размеры в мм и отношения:
| Экз. № | Д | В | Вп | ДПЧ | ДЗК | Д : В | Вп : В | ДПЧ : Д | ДЗК : Д |
|---|---|---|---|---|---|---|---|---|---|
| 1/02-06.113 | 18.5 | 11.8 | – | 8.0 | 16 | 1.57 | – | 0.43 | 0.86 |
| 2/02-06.113 голотип |
15.0 | 9.0 | 2.5 | 6.0 | 13 | 1.67 | 0.28 | 0.40 | 0.87 |
| 3/02-06.113 | ~22 | 15.0 | – | 9.0 | ~19 | 1.47 | – | ~0.41 | ~0.86 |
Сравнение. По очертаниям раковины наиболее близок к U. fassaensis (Wissmann), особенно сходен с экземплярами, описанными К. Сириаксом (Ciriacks, 1963) из нижнеиндской формации Динвуди Вайоминга, США, отличаясь менее крупными макушками и почти полным отсутствием килевого перегиба створок. От остальных видов рода заметно отличается формой раковины.
Материал. Два ядра правых створок, ядро двустворчатого экземпляра и отпечаток правой створки преимущественно удовлетворительной сохранности из типового местонахождения.
Список литературы
Бяков А.С. Новая зональная схема пермских отложений Северо-Востока Азии по двустворчатым моллюскам. Ст. 1. Зональное расчленение // Тихоокеанск. геол. 2012. Т. 31. № 5. С. 13–40.
Бяков А.С. Морские двустворчатые моллюски как индикаторы климатических изменений в перми Северо-Востока Азии // Уч. зап. Казанск. ун-та. Сер. Естеств. науки. 2020. Вып. 2. С. 550–570.
Бяков А.С., Горячев Н.А., Ведерников И.Л. и др. Новые результаты U-Pb SHRIMP датирования цирконов из верхневучапинских (верхняя пермь) отложений Северо-Востока России // Докл. Акад. наук. 2017. Т. 477. № 3. С. 331–336.
Бяков А.С., Кутыгин Р.В. Новая находка иноцерамоподобных двустворок рода Atomodesma в Южном Верхоянье и инвазии внебореальных моллюсков в перми Северо-Востока Азии // Палеонтол. журн. 2020. № 5. С. 20–25.
Бяков А.С., Кутыгин Р.В., Горячев Н.А. и др. Открытие позднечансинского комплекса двустворок и два эпизода вымирания фауны в конце перми на северо-востоке Азии // Докл. Акад. наук. 2018. Т. 480. № 1. С. 121–124.
Дагис А.А., Корчинская М.В. Первые находки конодонтов в отоцерасовых слоях Свальбарда // Бореальный триас. М.: Наука, 1987. С. 110–113.
Дагис А.С., Дагис А.А., Казаков А.М. и др. Биостратиграфия нижнего инда Восточного Верхоянья // Биостратиграфия мезозоя Сибири и Дальнего Востока. Новосибирск: Наука, 1986. С. 21–31.
Кипарисова Л.Д. Нижнетриасовые пластинчатожаберные Уссурийского края // Тр. ГИН АН СССР. 1938. Т. 7. С. 197–283.
Кипарисова Л.Д., Бычков Ю.М., Полуботко И.В. Позднетриасовые двустворчатые моллюски Северо-Востока СССР. Магадан, 1966. 312 с.
Кипарисова Л.Д., Курбатов В.С. О наличии триасовых отложений в Туар-кыре // Изв. АН СССР. Сер. геол. 1952. № 6. С. 76–81.
Кутыгин Р.В., Будников И.В., Бяков А.С. и др. Первые находки цератитов рода Otoceras в Кобюминской зоне Южного Верхоянья, Северо-Восток России // Уч. зап. Казанск. ун-та. Сер. Естеств. науки. 2019. Т. 161. № 4. С. 550–570.
Муромцева В.А., Гуськов В.А. Пермские морские отложения и двустворчатые моллюски Советской Арктики. Л.: Недра, 1984. 208 с.
Невесская Л.А., Попов С.В., Гончарова И.А. и др. Двустворчатые моллюски России и сопредельных стран в фанерозое. М.: Научн. мир, 2013. 524 с.
Ядренкин А.В., Бяков А.С., Кутыгин Р.В., Копылова А.В. Новые находки и стратиграфическое распределение фораминифер в пограничных пермско-триасовых отложениях Южного Верхоянья // Тихоокеанск. геол. 2020. Т. 39. № 5. С. 69–82.
Biakov A.S., Shi G.R. Palaeobiogeography and palaeogeographical implications of Permian marine bivalve faunas in Northeast Asia (Kolyma-Omolon and Verkhoyansk-Okhotsk regions, northeastern Russia) // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2010. V. 298. № 1–2. P. 42–53.
Budnikov I.V., Kutygin R.V., Shi G.R. et al. Permian stratigraphy and paleogeography of Central Siberia (Angaraland) – A review // J. Asian Earth Sci. 2020. V. 196. Article 104365. P. 1–21.
Ciriacks K.W. Permian and Eotriassic bivalves of the Middle Rockies // Bull. Amer. Mus. Natur. Hist. 1963. V. 125. № 1. 99 p.
Chu D.L., Yu J.X., Tong J.N. et al. Biostratigraphic correlation and mass extinction during the Permian-Triassic transition in continental-marine siliciclastic settings of South China // Global and Planetary Change. 2016. V. 146. P. 67–88.
Farabegoli E., Perri M.C., Posenato R. Environmental and biotic changes across the Permian-Triassic boundary in western Tethys: the Bulla parastratotype, Italy // Global and Planetary Change. 2007. V. 55. P. 109–135.
Feng X., Chen Z.Q., Bottjer D.J. et al. Unusual shallow marine matground-adapted benthic biofacies from the Lower Triassic of the northern Paleotethys: Implications for biotic recovery following the end-Permian mass extinction // Earth-Sci. Rev. 2019. V. 189. P. 194–219.
Foster W.J., Danise S., Twitchett R.J. A silicified Early Triassic marine assemblage from Svalbard // J. Syst. Palaeontol. 2017. V. 15. P. 851–877.
Foster W.J., Lehrmann D.J., Hirtz J.A. et al. Early Triassic benthic invertebrates from the Great Bank of Guizhou, South China: systematic palaeontology and palaeobiology // Papers in Palaeontol. 2019. V. 5. № 4. P. 613–656.
Hofmann R., Hautmann M., Brayard A. et al. Recovery of benthic marine communities from the end-Permian mass extinction at the low latitudes of eastern Panthalassa // Palaeontology. 2014. V. 57. P. 547–589.
Hofmann R., Hautmann M., Bucher H. A new paleoecological look at the Dinwoody Formation (Lower Triassic, western USA): intrinsic versus extrinsic controls on ecosystems recovery after the end-Permian mass extinction // J. Paleontol. 2013. V. 87. P. 854–880.
Hofmann R., Hautmann M., Bucher H. Recovery dynamics of benthic marine communities from the Lower Triassic Werfen Formation, northern Italy // Lethaia. 2015. V. 48. P. 474–496.
Kumagae T., Nakazawa K. Bivalves // The Lower Triassic System in the Abrek Bay Area, South Primorye, Russia / Eds. Shigeta Y., Zakharov Y.D., Maeda H., Popov A.M. Tokyo: Nat. Mus. of Nature and Sci., 2009. P. 156–173.
Posenato R. Survival patterns of macrobenthic marine assemblages during the end-Permian mass extinction in the western Tethys (Dolomites, Italy) // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2009. V. 280. P. 150–167.
Song T., Tong J., Tian L. et al. Taxonomic and ecological variations of Permian-Triassic transitional bivalve communities from the littoral clastic facies in southwestern China // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2019. V. 519. P. 108–123.
Spath L.F. Addition to the Eo-Triassic invertebrate fauna of East Greenland // Meddel. Grønland. 1935. V. 98. № 2. 115 p.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал