

Палеонтологический журнал, 2021, № 4, стр. 10-17
Новый род Balkhanomeandra (Scleractinia) из нижнего мела Туркменистана и Азербайджана
И. Ю. Бугрова *
Санкт-Петербургский государственный университет, Институт наук о Земле
Санкт-Петербург, Россия
* E-mail: i.bugrova@spbu.ru
Поступила в редакцию 21.11.2019
После доработки 12.12.2019
Принята к публикации 14.02.2020
Аннотация
В составе семейства Latomeandridae Alloiteau, 1952, emend. Roniewicz, 1976 установлен новый род склерактиний из нижнемеловых отложений Западного Туркменистана и Азербайджана. Представители данного рода ранее были изучены автором и Е.И. Кузьмичевой и относились к родам Meandrophyllia d’Orbigny, Microphyllia d’Orbigny и Fungiastraea Alloiteau. Более детальное изучение микроструктуры и изменчивости этих кораллов из коллекции автора позволило пересмотреть их систематическое положение и выделить новый род латомеандрид Balkhanomeandra gen. nov. с типовым видом B. roniewiczae sp. nov.
ВВЕДЕНИЕ
Шестилучевые кораллы отряда Scleractinia относятся к основным рифостроителям раннемеловой эпохи, широко распространенным в Тетической области. Представители подотряда Microsolenina Morycowa et Roniewicz, 1995 составляют одну из наиболее значительных по численности групп породообразующих склерактиний в рифогенных отложениях нижнего мела горных областей Закаспия, Кавказа и Крыма. Очень многочисленными по количеству остатков в этих регионах являются микросоленины родов Microsolena Lamoroux, 1821, Dimorpharea Fromentel, 1861, Meandrarea Etallon, 1858, принадлежащие семейству Microsolenidae Koby, 1890, а также родов Latiastraea Beauvais, 1964, Microphyllia d’Orbigny, 1849, Fungiastraea Alloiteau, 1952, Dimorphastraea Fromentel, 1857 из семейства Latomeandridae Alloiteau, 1952, emend. Roniewicz, 1976 (Бендукидзе, 1961; Сихарулидзе, 1985; Бугрова и др., 1985, 1987; Кузьмичева, 1987, 1988, 2002; Bugrova, 1990; Бугрова, 1995, 1997, 1999, 2005, 2009). Как правило, они представлены широко распространенными видами.
В комплексах рифостроящих склерактиний нижнего мела Западного Туркменистана и азербайджанской части Малого Кавказа автором выявлен новый род микросоленин, который также относится к основным породообразователям. Представители этого рода ранее были описаны И.Ю. Бугровой и Е.И. Кузьмичевой, но были отнесены либо к другим родам и семействам – Meandrophyllia d’Orbigny, 1849 семейства Haplaraeidae Vaughan et Wells, 1943 (Бугрова и др., 1987), Fungiastraea семейства Thamnasteriidae Vaughan et Wells, 1943 (Кузьмичева, 1987), либо к другому роду (Microphyllia d’Orbigny, 1849) того же семейства Latomeandridae Alloiteau, 1952 (Бугрова, 1999). По результатам более детального изучения микроструктуры и изменчивости этих кораллов из обширной коллекции автора пересмотрено их систематическое положение. В составе семейства Latomeandridae они выделены в новый род Balkhanomeandra с типовым видом B. roniewiczae sp. nov. Главными отличиями нового рода от гомеоморф являются: 1) цериоидно-меандроидное строение полипняков с отчетливым преобладанием цериоидных или меандроидных форм; 2) полное или почти полное слияние периферических краев септ (т.е., конфлюэнтные, реже субконфлюэнтные септы); 3) присутствие ложбинных септ; 4) наличие пеннул и их форма, типичная для семейства Latomeandridae; 5) неполная синаптикулотекальная стенка.
МАТЕРИАЛ И МЕТОДЫ
Изученные остатки склерактиний рода Balkhanomeandra gen. nov. происходят из нескольких местонахождений в Закаспии и на Малом Кавказе (рис. 1). Наиболее представительная коллекция (более 40 экз.) собрана автором в 1979–1991 гг. в Закаспии (Западный Туркменистан). В изученных местонахождениях кораллы приурочены к отложениям двух формационных комплексов: карбонатного неокомского и нижней части терригенного среднемелового (Прозоровский, 1991). Кораллы изучены в разрезах южной части хр. Большой Балхан (нижний готерив), в западной части хр. Малый Балхан (верхи баррема), на возвышенности Кельдже в южной части Туаркыра (баррем) и в разрезах Огланлинской гряды (верхняя часть баррема). Материал хранится в Палеонтолого-стратиграфическом музее Санкт-Петербургского государственного ун-та (ПСМ СПбГУ, колл. №№ 347, 361, 367, 372).
Рис. 1.
Схема местонахождений кораллов нового рода Balkhanomeandra на территории Туркменистана и Азербайджана. Обозначения: 1 – хр. Большой Балхан, ущелье Ташлы-Дере; 2 – хр. Большой Балхан, гряда Шахлибурун; 3 – Огланлинская гряда; 4 – горы Туаркыр, возвышенность Кельдже; 5 – хр. Малый Балхан; 6 – бассейн р. Базарчай, с. Аликулиушагы.
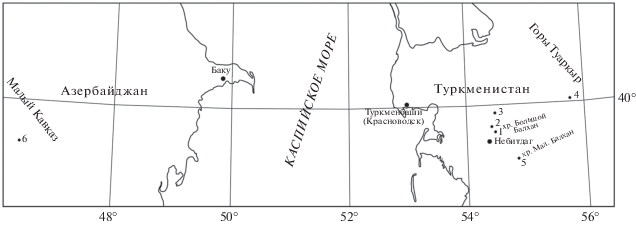
Имеется также два экземпляра из коллекции М.Б. Преображенского, собранные из зейвинской свиты в азербайджанской части Малого Кавказа в бассейне р. Базарчай (баррем) (ПСМ СПбГУ, колл. № 272).
Все изученные кораллы представлены массивными уплощенными полипняками, как небольших, так и крупных размеров (до 60 см диаметром и 25 см в высоту). Степень сохранности материала различна – от хорошей (с фрагментами сохранившейся микроструктуры) до удовлетворительной (частичная перекристаллизация колонии). В коллекцию не включались колонии неудовлетворительной сохранности (т.е., сохранившие внешние признаки, но с полностью разрушенными перекристаллизацией внутренними элементами).
Кораллы изучались с применением традиционных методов полевых геологических, палеонтологических и палеоэкологических исследований с детальным описанием вмещающих пород, характера расположения в них органических остатков и их состава, типа сохранности остатков, с полевым описанием самих коралловых поселений, замером кораллов, отбором целых экземпляров или их фрагментов (при нахождении крупных полипняков). Последующее изучение в камеральных условиях включало в себя: детальное рассмотрение и описание под бинокулярным микроскопом внешней морфологии кораллов, изготовление поперечных и продольных шлифов (всего изготовлено 82 шлифа) и изучение их под оптическим бинокулярным и поляризационным микроскопом для дальнейшего выявления деталей внутреннего строения склерактиний (микроморфологический и микроструктурный анализ) и определения их систематической принадлежности. При изучении кораллов применялись оптический микроскоп МБС-10 и поляризационный Leitz Laborlux 12 POL. Фотографирование шлифов производилось в Ресурсном центре СПбГУ “Рентгенодифракционные методы исследования” с помощью микроскопа Leica DM4500 P с микропозиционным столиком.
МЕСТОНАХОЖДЕНИЯ КОРАЛЛОВ РОДА BALKHANOMEANDRA
В Закаспии кораллы рода Balkhanomeandra встречаются в следующих отложениях карбонатной неокомской формации.
Аликперская рифогенная система. Эта система входит в состав ахалтекинской серии (нижний готерив) хр. Большой Балхан и представляет собой закономерное сочетание разнофациальных рифовых отложений суммарной мощностью 5–20 м (Преображенский, 1987, 1995; Preobrazhensky, 1990). Ранее отложения Аликперской системы описывали в составе арланской свиты, относимой к берриасу–валанжину (Прозоровский, 1970; Решения …., 1977; Бугрова и др., 1985, 1987; Bugrova, 1990; Преображенский, 1995). В разрезе этой системы в южной части Большого Балхана (ущелье Ташлыдере, гряда Шахлибурун) с севера на юг последовательно сменяются фациальные зоны: береговой суши, мелководного прибрежного бассейна с коралловыми биогермами и биостромами, рудистовых банок, зарифового калькаренитового шлейфа, кораллово-водорослевого барьера, предрифового шлейфа (Бугрова и др., 1987; Bugrova, 1990). Многочисленные и разнообразные по систематическому составу кораллы собраны и в органогенных постройках, и в промежутках между ними. Биогермы с основанием 2.5–9 м и достигающие 2 м в высоту сложены биогенными известняками со структурой фреймстоун и байндстоун. Они могут сливаться в нижней части и местами переходить в биостромы. Основу построек составляют крупные массивные уплощенные колонии от 5–60 до 30–60 см (реже до 80 см) в поперечнике и до 15 см (реже до 50 см) в высоту, а также массивно-ветвистые (рамозные) колонии до 100 см в основании и до 50 см в высоту. Пространство между постройками заполнено флаутстоуном и вакстоуном с обломками массивных и фацелоидных колоний. В самих постройках и между ними многочисленны остатки организмов-рифолюбов, таких как водоросли, одиночные склерактинии, брахиоподы, двустворчатые моллюски, гастроподы, правильные морские ежи, серпулы, губки и фораминиферы.
Кумдагская свита. Свита, относимая к баррему, развита в горах Копетдаг и Малый Балхан и сложена преимущественно известняками без примеси терригенного материала (мощностью от 120 до 350 м) (Нижний мел …, 1985; Стратиграфия …, 1986; Прозоровский, 1991; Преображенский, 1995). Верхняя ее часть представлена типичной ургонской фацией (Прозоровский, 1989; Бугрова и др., 1985; Кузьмичева, 2002), для которой характерно развитие кораллово-рудистово-орбитолиновых сообществ. Кораллы собраны в западной части хр. Малый Балхан в верхней пачке свиты (XXI пачка по В.И. Марченко) (Марченко, 1962; Бугрова, 1999; Кузьмичева, 2002), относимой в настоящее время к верхам баррема. Пачка мощностью до 60 м представляет собою цикличное чередование карбонатных (известняки), терригенно-карбонатных (глинистые и алевритистые известняки, мергели) и глинистых отложений. Из известковых и глинисто-карбонатных прослоев мощностью 2–15 м, представленных погребенными водорослево-коралловыми и рудистовыми поселениями, описано богатое сообщество склерактиний и разнообразных рифолюбивых организмов (водорослей, одиночных склерактиний, брахиопод, двустворчатых моллюсков, гастропод, морских ежей, серпул, губок и фораминифер) (Бугрова, 1999, 2005). На четырех уровнях наблюдаются водорослево-коралловые биогермы высотой 2–5 м (реже до 8 м) и шириной основания 3–8 м, расположенные на расстоянии 4–15 м друг от друга. Тела построек сложены фреймстоуном, пространство между ними заполнено вакстоуном и пакстоуном, реже – грейнстоуном. Биогермы образовались путем нарастания массивных, рамозных и фацелоидных колоний склерактиний. Размеры массивных достигают 10–120 см в поперечнике и 20–60 см в высоту, рамозных – до 1.5–8 м в ширину и 2–3 м в высоту. Фацелоидные имеют ширину основания до 2 м и высоту до 50–60 см (Бугрова, 1999). Рамозные колонии часто образуют крупные монотаксонные постройки.
Борджоклинская свита. Свита относится к баррему, развита в северной части хребта Большой Балхан, в горах Кубадаг и Туаркыр, где представлена чередованием различных известняков, мергелей-ракушечников, а также алевролитов и глин, имеющих подчиненное значение (Решения…, 1977; Верба, Прозоровский, 1979; Стратиграфия…, 1986; Прозоровский, 1991). Мощность свиты 12–50 м. Верхняя ее часть выражена ургонской фацией. Изученные кораллы собраны в юго-западной части возвышенности Кельдже (южная часть Туаркыра) вблизи кровли свиты, в трехметровом слое плотных ракушечников. В основании слоя в грейнстоунах и слабоглинистых пакстоунах наблюдаются скопления целых и раздробленных раковин двустворок. На них и рядом с ними расположены многочисленные небольшие по размеру (до 10 см в поперечнике и 8 см в высоту) массивные колонии склерактиний уплощенной и желваковидной формы, не образующие построек. К верхней, более глинистой части слоя приурочено большое количество отдельно стоящих крупных (до 85 см в основании и 30 см в высоту) фацелоидных полипняков.
Терригенная среднемеловая формация западной части Средней (Центральной) Азии. Находки нового рода склерактиний приурочены к ее низам – основанию нижней части большебалханской свиты (Меловые …, 1980; Стратиграфия …, 1986; Прозоровский, 1991). Разрезы последней на Большом Балхане, в Кубадаге и Туаркыре сложены чередованием глауконитовых алевролитов, глин, песчаников с линзами органогенных известняков общей мощностью 5–120 м. Возраст свиты – поздняя часть баррема – начало апта. Остатки склерактиний обнаружены в Огланлинской гряде в линзах известняков мощностью до 0.4 м. Они представлены небольшими (до 15 см в диаметре и 7 см в высоту) разрозненными массивными колониями и одиночными формами (Бугрова, 1995). Совместно с кораллами встречаются брахиоподы, двустворчатые моллюски, морские ежи и фораминиферы.
На Малом Кавказе кораллы собраны из зейвинской свиты (Паффенгольц, 1951; Халилов и др., 1977), распространенной в смежных районах Армении и Азербайджана (восточная часть Зангезурского хребта). Возраст свиты барремский (Схемы …, 1986). Она представлена органогенными, оолитовыми, пелитоморфными и песчанистыми известняками, входящими в состав ургонского комплекса и содержащими органогенные постройки. В Кубатлинском р-не в бассейне р. Базарчай к югу от с. Аликулиушагы свита достигает мощности 224 м и содержит три уровня органогенных построек (Халилов и др., 1977; Кузьмичева, 2002). Биогермы в ее основании достигают ширины 40 м и высоты 25–30 м, причем биогермы небольших размеров часто сливаются в нижней части. Биостромы имеют протяженность до 50 м и высоту 5 м. Колониальные склерактинии слагают тела органогенных построек, наряду с водорослями и строматопоратами, а также встречаются в изобилии в виде изолированных экземпляров в отложениях другого литологического состава. Они представлены массивными сферическими, полусферическими, реже – рамозными и фацелоидными полипняками средних размеров (до 15 см в поперечнике и в высоту), а также одиночными формами (Кузьмичева, 1988, 2002). Сопутствующая богатая ассоциация рифолюбивых организмов состоит из губок, серпул, двустворок (в том числе рудистов), гастропод, брахиопод, крупных фораминифер, морских ежей, морских лилий.
Ниже приведены описания нового рода и типового вида склерактиний. При описании склерактиний автором используется система, принятая в работах Ж. Аллуато (Alloiteau, 1952, 1957) с дополнениями, предложенными Э. Роневич и Э. Морыцовой (Morycowa, Roniewicz, 1995).
СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
ОТРЯД SCLERACTINIA
ПОДОТРЯД MICROSOLENINA
СЕМЕЙСТВО LATOMEANDRIDAE ALLOITEAU, 1952, EMEND. RONIEWICZ, 1976
Род Balkhanomeandra Bugrova, gen. nov.
Название рода – от хребтов Большой и Малый Балханы в Западном Туркменистане.
Типовой вид – Balkhanomeandra roniewiczae sp. nov.
Диагноз. Массивные колонии цериоидно-меандроидного типа (табл. II , фиг. 1а, 1в; см. вклейку). Почкование внутричашечное конечное. Меандры короткие и длинные. Присутствует большое количество обособленных кораллитов (табл. II , фиг. 1а), которые могут резко преобладать в некоторых колониях (табл. III , фиг. 2 а; см. вклейку). Форма изолированных чашек изменчива, обычно округло-полигональная или вытянутая. Ложбины довольно глубокие, гребни закругленные. Септы неравномерно-пористые (пористость увеличивается к осевой части кораллитов) (табл. II , фиг. 1а–г, 2а; табл. III , фиг. 1, 2 а, 2б), сливающиеся или почти сливающиеся периферическими краями с септами соседних кораллитов (конфлюэнтные и субконфлюэнтные). Характерен анастомоз септ. Развиты длинные ложбинные септы. Септы почти равной толщины во всей длине, состоят из простых трабекул (табл. II , фиг. 1в), расположенных в одной дивергентной системе. Дистальный край с небольшими округлыми зубчиками. Хорошо развиты пеннулы с приподнятыми краями (латомеандридного типа) (табл. II , фиг. 1г, 2а, 2б; табл. III , фиг. 3 в). Эндотеку слагают немногочисленные синаптикулы, расположенные главным образом в области гребней, и хорошо развитые диссепименты, слегка вогнутые в осевой части и приподнятые по периферии. Стенка – неполная синаптикулотека. Столбик париетальный.
Видовой состав. Типовой вид из нижнего готерива – баррема Западного Туркменистана и баррема Азербайджана.
Сравнение. Многие признаки, такие как тип почкования, наличие меандр, пористость и анастомоз септ, пеннулярная орнаментация их боковых поверхностей, присутствие ложбинных септ сближают новый вид с представителями рода Microphyllia d’Orbigny, 1849 (сем. Latomeandridae). Отличие Balkhanomeandra gen. nov. состоит в строении колонии – цериоидно-меандроидном, часто с резким преобладанием одиночных кораллитов, а не меандроидном; в наличии сливающихся или почти сливающихся септальных пластинок, более регулярной пористости септ, четкого париетального столбика и неполной синаптикулотекальной стенки.
Замечания. У рода Balkhanomeandra gen. nov. проявляется гомеоморфное сходство с Meandrarea Etallon, 1859 (сем. Microsolenidae Koby, 1890) и Meandrophyllia d’Orbigny, 1849 (сем. Haplaraeidae Vaughan et Wells, 1943). От Meandrarea новый род отличается цериоидно-меандроидным строением колонии, присутствием ложбинных септ, менее равномерной пористостью септ, более отчетливой стенкой, более развитыми диссепиментами и выраженным париетальным столбиком. Главным отличием от чрезвычайно сходного рода Meandrophyllia является развитие пеннул, а не гранул на боковой поверхности септ.
Сравнение морфологических признаков рода Balkhanomeandra gen. nov. и других родов склерактиний приведено в табл. 1.
Таблица 1.
Сравнительная характеристика рода Balkhanomeandra gen. nov. и некоторых других родов отряда Scleractinia
| Подотряд | Microsolenina Morycowa et Roniewicz, 1995 | Fungiina Verrill, 1865 | ||
|---|---|---|---|---|
| Семейство | Latomeandridae | Microsolenidae | Haplaraeidae | |
| Род | Microphyllia | Balkhanomeandra | Meandrarea | Meandrophyllia |
| Тип колонии | Меандроидный | Цериоидно-меандроидный | Меандроидный | Цериоидно-меандроидный |
| Способ почкования | Внутричашечное | Внутричашечное | Внуртичашечное | Внутричашечное |
| Форма гребней | Заостренные | Закругленные | Закругленные | Закругленные, заостренные (?) |
| Микроморфология септ | Пеннулы и менианы | Пеннулы | Пеннулы | Игловидные гранулы |
| Костосепты | Не сливающиеся (реже – сливающиеся или почти сливающиеся) | Сливающиеся (редко – почти сливающиеся) | Сливающиеся (реже – почти сливающиеся) | Сливающиеся, почти сливающиеся |
| Ложбинные септы | Присутствуют | Присутствуют | Отсутствуют | Присутствуют |
| Анастомоз септ | Присутствует | Присутствует | Присутствует | Присутствует |
| Пористость септ | Редкая, чаще на внутренних краях септ | Редкая, чаще на внутренних краях септ | Регулярная | Редкая (септы почти компактные) |
| Синаптикулы | Присутствуют в стенке | Присутствуют главным образом в стенке | Присутствуют ближе к гребням | Присутствуют в стенке, немногочисленные |
| Тип стенки | Синаптикулотекальная | Синаптикулотекальная, неполная | Неотчетливая | Синаптикулотекальная, неполная |
| Эндотека | Диссепименты | Диссепименты выпуклые по периферии кораллита и вогнутые в центре | Редкие табулоидные диссепименты | Табулоидные диссепименты |
| Столбик | Отсутствует или присутствует париетальный (папиллярный), рудиментарный | Париетальный | Монотрабекулярный, или небольшой париетальный, неотчетливый; может отсутствовать | Париетальный |
Признаки нового рода и типового вида наблюдаются у экземпляра, описанного из нижнего баррема Грузии Г.Я. Сихарулидзе (1985, с. 52, табл. XXV , фиг. 3 ) как Microphyllia undans (Koby, 1885). Единственное отличие последнего от Balkhanomeandra gen. nov. в наличии не пеннул, а гранул на боковых поверхностях септ. Из-за отсутствия пеннул этот коралл может быть отнесен и к роду Microphyllia. К сожалению, из-за отсутствия в работе Сихарулидзе изображения продольного сечения колонии и неудовлетворительного изображения поперечного сечения трудно судить о характере орнаментации септ (пеннулы или гранулы?) и сделать вывод о родовой принадлежности этого коралла.
Возможно, к новому роду Balkhanomeandra относится также экземпляр, описанный Р. Барон-Жабо (Baron-Szabo, Fernandes-Mendiola, 1997, с. 48, рис. 4F ) из альба Северной Испании как Meandrarea meandroides Koby, 1898 и отнесенный в дальнейшем этим автором к роду Meandrophyllia (Baron-Szabo et al., 2003, с. 210).
Не исключено, что к роду Balkhanomeandra gen. nov. принадлежит также Meandrophyllia meandroides (Koby), описанный из верхнего апта – нижнего альба Центрального Ирана (Baron-Szabo et al., 2003, с. 210, табл. 37 , фиг. 1).
Барон-Жабо указывает на наличие у этих форм как коротких меандр, так и изолированных чашек, почти равных по толщине; неравномерно пористых септ, которые сливаются или почти сливаются периферическими краями; а также пеннул, париетального столбика, неполной синаптикулотекальной стенки, выпуклых и уплощенных диссепиментов. На приведенном изображении (Baron-Szabo, Fernandes-Mendiola, 1997, рис. 4F ) хорошо заметно присутствие ложбинных септ. Все это соответствует диагнозу рода Balkhanomeandra gen. nov. Наличие пеннул не позволяет относить описанные Барон-Жабо экземпляры к роду Meandrophyllia, входящему в семейство Haplararaeidae. Их признаки не соответствуют и признакам рода Meandrarea, что убедительно показала Морыцова (Morycowa, Masse, 2009, с. 119). К сожалению, в работах не приведены изображения вертикальных сечений колоний, которые бы позволили составить полное представление об орнаментации септ, поэтому систематическая принадлежность форм, описанных Барон-Жабо, требует уточнения.
Balkhanomeandra roniewiczae Bugrova, sp. nov.
Табл. II , фиг. 1, 2 ; табл. III , фиг. 1–3
? Microphyllia undans (Koby, 1885): Сихарулидзе, 1985, с. 52, табл. XXV , фиг. 3 ; Бугрова, 1999, с. 40, табл. III , фиг. 1а–в.
Meandrophyllia sp.: Бугрова в: Бугрова и др., 1987, с. 98, табл. I , фиг. 4 а, 4б, 5.
Fungiastraea tendagurensis (Dietrich, 1925–27): Кузьмичева, 1987, с. 247, табл. V , фиг. 4 ; табл. VI , фиг. 1.
? Meandrarea meandroides Koby, 1898: Baron-Szabo in: Baron-Szabo, Fernandes-Mendiola, 1997, с. 48, рис. 4F .
? Meandrophyllia meandroides (Koby, 1898): Baron-Szabo in: Baron-Szabo et al., 2003, с. 210, табл. 37 , фиг. 1.
Вид назван в честь исследователя кораллов Э. Роневич (Ewa Roniewicz).
Голотип – ПСМ СПбГУ, № 367/60, два шлифа – поперечный и продольный; Западный Туркменистан, хр. Большой Балхан, гряда Шахлибурун; Аликперская рифогенная система, нижний готерив.
Описание. Массивные колонии цериоидно-меандроидного типа. Размеры и форма найденных экземпляров колеблются в широких пределах: высота колоний достигает 25 см, диаметр – 60 см. Почкование внутричашечное конечное. Меандры короткие (по две–три чашки) и длинные (четыре и более чашек), много обособленных кораллитов, в некоторых колониях они преобладают. Форма изолированных чашек изменчива; обычно они округло-полигональные или вытянутые, размер сечения изолированных кораллитов колеблется от 4 до 5–6.5 мм в поперечнике. Ложбины довольно глубокие, гребни закругленные. Расстояние между гребнями составляет (3.5)4–6.5(7) мм, а между центрами чашек в ложбине – (3.5)4–6(8) мм. Септы прямые или слабоизогнутые по направлению к осевой части кораллитов, с неравномерно расположенными порами (пористость увеличивается к осевой части кораллитов), довольно тонкие и почти равной толщины во всей длине, состоящие из простых трабекул, расположенных в одной дивергентной системе. Дистальные края с небольшими округлыми зубчиками, обычно не сохраняются. Периферические края септ соседних кораллитов сливаются или почти сливаются (т.е., развиты септы конфлюэнтные или субконфлюэнтные). По длине различаются септы трех–четырех порядков. Септы первого порядка наименее пористые. В поперечном сечении участки септ без пор имеют четковидную форму, в них насчитываются 4–5 трабекул на 1 мм. Наблюдается анастомоз септ. Хорошо развиты длинные ложбинные септы. На боковых поверхностях септ хорошо развиты пеннулы с приподнятыми краями. Количество септ в изолированных чашках – 26–52. Осевой части кораллита достигают 15(20) септ первого–третьего порядка; выступы трабекул на их внутренних краях, сплетаясь, образуют париетальный столбик. Септы четвертого порядка короткие (до 1/4 длины септ первого порядка). Эндотеку слагают немногочисленные синаптикулы, расположенные, главным образом, в области гребней, и хорошо развитые диссепименты, слегка вогнутые в осевой части и приподнятые по периферии.
В периферической части кораллитов (в том числе, в стенке) на прямолинейных участках на 2 мм насчитывается 6–7(8) септ. Стенка – неполная синаптикулотека.
Размеры в мм элементов и их количество:
| Местонахождение экземпляров | Большой Балхан | Малый Балхан | Малый Кавказ |
|---|---|---|---|
| Число чашек в меандрах | До 4 и более | 2–3 | До 4 |
| Диаметр изолированных кораллитов, мм | 4 до 5–6.5 | 4–5 | |
| Ширина ложбин (расстояние между гребнями), мм | (3.5)4–6.5(7) мм | 3.5–4 | 3.5–4 |
| Расстояние между центрами обособленных чашек, мм | 4–5 | 4–6 | Не установлено |
| Расстояние между центрами чашек в ряду (в меандрах), мм | (3.5)4–6 | 5.5 | 3–5 |
| Число септ в изолированных чашках | 48(52), до центра доходит около 12 (до 20) | 42–48 | 46–48 |
| Число септ в стенке на 2 мм | 6–7(8) | 6–7 | 6–7(8) |
| Толщина септ, мм | До 0.2 | До 0.2 | 0.2(0.25) |
| Частота трабекул на 1 мм | 4–5 | 4–5 | 5 |
Изменчивость. Вид Balkhanomeandra roniewiczae sp. nov. является одним из самых распространенных в кораллосодержащих отложениях нижнего мела Западного Туркменистана. Он обладает значительной экологической изменчивостью, выраженной как в разнообразной внешней форме полипняков, так и в их внутреннем строении. Колонии, собранные на Малом Балхане, отличаются от найденных на Большом Балхане, в Огланлинской гряде и на Кельдже гораздо меньшими размерами и резким преобладанием изолированных чашек при малочисленных и коротких меандрах.
Сравнение. В роде Balkhanomeandra один вид.
Распространение. Нижний готерив – баррем Западного Туркменистана и баррем Азербайджана.
Материал. Колонии из пяти местонахождений на территории Западного Туркменистана и одного – на территории Азербайджана. Колл. № 367 – 1 экз. хорошей сохранности (поперечный и продольный шлифы); колл. № 361 – 1 экз. хорошей сохранности (поперечный и продольный шлифы); колл. № 372 – 36 экз. хорошей и удовлетворительной сохранности (поперечные и продольные шлифы); колл. № 347 – 2 экз. удовлетворительной сохранности (поперечные и продольные шлифы).
Список литературы
Бендукидзе Н.С. К изучению нижнемеловых кораллов Крыма // Тр. ГИН АН ГССР. Сер. геол. 1961. Т. 12(42). 40 с.
Бугрова И.Ю. Фаунистический комплекс ургонской фации Огланлинской гряды гор Большого Балхана // Вопросы стратиграфии и региональной геологии. Межвуз. сб. / Ред. Прозоровский В.А. СПб.: Изд-во СПб. ун-та, 1995. С. 142–152.
Бугрова И.Ю. Кораллы // Атлас меловой фауны Юго-Западного Крыма / Ред. Аркадьев В.В., Богданова Т.Н. СПб.: Пангея, 1997. С. 18–39, 212–235.
Бугрова И.Ю. Новые данные о склерактиниях Юго-Западной Туркмении // Вопросы палеонтологии. Т. XI: Межвуз. сб. / Ред. Степанов Д.Л., Киселев Г.Н. СПб.: Изд-во СПб. ун-та, 1999. С. 33–42.
Бугрова И.Ю. Новые данные о кораллах раннего мела Туркменистана // Меловая система России: проблемы стратиграфии и палеогеографии. Сб. статей Всеросс. совещ. / Ред. В.В. Аркадьев, В.А. Прозоровский. СПб., 2005. С. 136–142.
Бугрова И.Ю. К ревизии кораллов отряда Scleractinia из коллекции к монографии Н.И. Каракаша “Нижнемеловые отложения Крыма и их фауна” в Палеонтолого-стратиграфическом музее кафедры динамической и исторической геологии СПбГУ // Матер. научн. конф., посвященной памяти чл.-корр. Петербургской Акад. наук, основателя кафедры геологии, геологического музея, геологического кабинета, проф. Санкт-Петербургского университета А.А. Иностранцева (СПб., 19–21 ноября 2009 г.) / Ред. В.В. Аркадьев. СПб., 2009. С. 76–79.
Бугрова И.Ю., Верба Ю.Л., Преображенский М.Б., Прозоровский В.А. Нижнемеловой рифовый комплекс Большого Балхана // Геологическое строение Туркменистана / Ред. Аманниязов К.Н. Ашхабад: Ылым, 1987. С. 79–103.
Бугрова И.Ю., Преображенский М.Б., Прозоровский В.А. Раннемеловые рифовые комплексы Западной Туркмении // Вестн. ЛГУ. Сер. Геол., геогр. 1985. Вып. 1. № 7. С. 22–30.
Верба Ю.Л., Прозоровский В.А. Стратиграфическая схема берриас–нижнебарремских отложений Западной Туркмении // Вопросы стратиграфии. Вып. 2. Л., 1979. С. 53–65.
Кузьмичева Е.И. Кораллы из нижнебарремских органогенных построек Малого Балхана и Туаркыра // Геологическое строение Туркменистана / Ред. Аманниязов К.Н. Ашхабад: Ылым, 1987. С. 217–262.
Кузьмичева Е.И. Меловые кораллы Азербайджана // Меловая фауна Азербайджана. Баку: Элм, 1988. С. 153–183.
Кузьмичева Е.И. Морфология скелета, система и эволюция склерактиний. М.: Наука, 2002. 212 с. (Тр. Палеонтол. ин-та РАН. Т. 286).
Марченко В.И. Неоком Копет-Дага. Кн. 1. Стратиграфия, литология и фации неокома Копет-Дага. Сер. Проблемы нефтегазоносности Средней Азии. Вып. 11. Л.: Гостоптехиздат, 1962. 328 с. (Тр. ВСЕГЕИ. Нов. сер. Т. 78).
Меловые отложения обрамления Каспийского моря / Ред. Бененсон В.А., Лебедев Л.И. М.: Наука, 1980. 244 с.
Нижний мел юга СССР / Ред. Бененсон В.А. М.: Наука, 1985. 224 с.
Паффенгольц К.Н. К стратиграфии меловых отложений восточной части Малого Кавказа // Изв. АН СССР. Сер. геол. 1951. № 1. С. 60–80.
Преображенский М.Б. О зональности и стадиях формирования нижнемелового рифового комплекса южного склона хребта Большой Балхан // Вестн. Ленингр. ун-та. 1987. Сер. 7. Вып. 2(№ 14). С. 94–96.
Преображенский М.Б. Копетдагская складчатая система // Рифогенные и сульфатоносные формации фанерозоя СССР / Ред. Беленицкая Г.А., Задорожная Н.М. М.: Недра, 1990. С. 102–106.
Преображенский М.Б. Фациальные зоны и этапы формирования карбонатных отложений верхней юры и нижнего мела Балхано-Копетдагского бассейна // Вопросы стратиграфии и региональной геологии. СПб.: Изд-во СПб. ун-та, 1995. С. 64–87.
Прозоровский В.А. О расчленении берриас–валанжинских отложений Западной Туркмении // Вестн. Ленингр. ун-та. Сер. Геол. и геогр. 1970. Вып. 4. № 24. С. 142–147.
Прозоровский В.А. Ургонская фация нижнего мела в Средней Азии // Вестн. Ленингр. ун-та. Сер. 7. 1989. Вып. 4. № 28. С. 20–25.
Прозоровский В.А. Верхняя юра и нижний мел запада Средней Азии. Стратиграфия и история геологического развития. Л.: Изд-во Ленингр. ун-та, 1991. 254 с.
Решения Межведомственного стратиграфического совещания по мезозою Средней Азии (Самарканд, 1971 г.). Л.: Изд-во ВСЕГЕИ, 1977. 48 с.
Сихарулидзе Г.Я. Гексакораллы ургонской фации Дзирульского массива и его северного обрамления // Тр. Геол. ин-та АН ГрузССР. 1985. Нов. сер. Вып. 88. 80 с.
Стратиграфия СССР. Меловая система (полутом I). М.: Недра, 1986. 340 с.
Схемы стратиграфии меловых отложений Азербайджана / Ред. Али-заде А., Халилов А.Г. Баку: Элм, 1986. 68 с.
Халилов А.Г., Алиев Г.А., Кузьмичева Е.И., Аскеров Р.Б. Ургонский комплекс бассейна р. Базарчай (Малый Кавказ) // Бюлл. МОИП. Отд. Геол. 1977. Т. 52. № 4. С. 85–93.
Alloiteau J. Madréporaires post-paléozoïques // Traité de Paléontologie. T. 1 / Ed. Piveteau J. P.: Masson, 1952. P. 539–684.
Alloiteau J. Contribution a la Systématique des Madréporaires Fossiles. P.: C.N.R.S., 1957. 426 p.
Baron-Szabo R.C., Fernandez-Mendiola P.A. Cretaceous scleractinian corals from the Albian of Cabo de Ajo (Cantabria Province, N-Spain) // Paläontol. Z. 1997. Bd 71. № 1–2. P. 35–50.
Baron-Szabo R.C., Hamedani A., Senowbari-Daryan B. Scleractinian corals from Lower Cretaceous deposits North of Esfahan (Central Iran) // Facies. 2003. V. 48. № 1. P. 199–215.
Bugrova I.J. The facies zonation and scleractinians of the Early Hauterivian reef complex of Bolshoy Balkhan // Cret. Res. 1990. V. 11. № 3. P. 229–236.
Morycowa E., Masse J.P. Lower Cretaceous Microsolenina (Scleractinia) from Provence (southern France) // Ann. Soc. Geol. Pol. 2009. V. 79. P. 97–140.
Morycowa E., Roniewicz E. Microstructural disparity between Recent fungaiine and Mesozoic microsolenine scleractinians // Acta Palaeontol. Pol. 1995. V. 40. № 4. P. 361–385.
Preobrazhensky M.B. Urgonian lithofacies and fauna of the Kopetdag Basin, southern USSR // Cret. Res. 1990. V. 11. № 3. P. 247–252.
Roniewicz E. Les Scléractiniaires du Jurassique supérieur de la Dobrogea Centrale, Roumanie // Palaeontol. Pol. 1976. V. 34. P. 17–118.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал