

Палеонтологический журнал, 2021, № 4, стр. 41-49
Новые стрекозы семейства Kennedyidae (Odonata: Protozygoptera) из верхнепермских отложений Вологодской области
А. С. Фелькер *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: felkafelka95@gmail.com
Поступила в редакцию 09.06.2020
После доработки 22.06.2020
Принята к публикации 22.06.2020
Аннотация
Из северодвинских отложений местонахождения Исады (Мутовинская линза; Вологодская обл., Великоустюгский р-н) описан новый вид стрекоз семейства Kennedyidae: Kennedya suchonensis sp. nov. Несмотря на преемственность основных особенностей жилкования крыльев, исадские стрекозы имеют необычное для протозигоптер строение дискоидальной области стебелька, позволяющее на новом уровне рассмотреть вопрос о морфогенезе этой структуры. Обсуждаются эволюционные связи исадских стрекоз с остальными кеннедиидами.
ВВЕДЕНИЕ
Kennedyidae Tillyard, 1925 – вымершее семейство стебельчатокрылых стрекоз, представители которого известны из пермских и триасовых отложений Северного полушария (Tillyard, 1925; Martynov, 1930; Carpenter, 1931, 1933, 1939, 1947; Притыкина, 1981; Nel et al., 2012; Felker, 2020). Признаки строения тела пока не описаны детально, но по особенностям жилкования кеннедиид относят к инфраотряду Protozygoptera Tillyard, 1925. В состав семейства включают небольших по размеру стрекоз с редуцированным жилкованием крыльев, основными признаками которых являются: наличие стебелька, заметно укороченная анальная жилка и редкое поперечное жилкование.
Самые разнообразные комплексы кеннедиид происходят из кунгурских местонахождений Эльмо, Мидко (США) (Tillyard, 1925; Carpenter, 1931, 1933, 1939, 1947) и Чекарда (Россия) (Nel et al., 2012; Фелькер, Василенко, 2017), северодвинского Исады (Россия) (Василенко, 2013; Фелькер, Василенко, 2015, 2017), индского Бабий Камень (Россия) (Фелькер, 2021) и ладин–карнийского Джайлоучо (Киргизия) (Притыкина, 1981). В настоящей статье описывается часть материала из местонахождения Исады, представленная новым видом рода Kennedya.
Исады – одно из самых богатых позднепалеозойских местонахождений ископаемых насекомых (Rasnitsyn et al., 2013). За период целенаправленных палеоэнтомологических сборов c 2005 г. по настоящее время из него известно уже более 7000 остатков ископаемых насекомых разной степени сохранности: от неполных крыльев до отпечатков целых насекомых. Наиболее перспективные насекомоносные слои находятся в пределах Мутовинской линзы, сложенной сероцветными породами кичугской и каликинской пачек полдарсской свиты северодвинского яруса (Golubev, 2013). Сама линза вместе с вмещающими красноцветами верхней перми расположена на левом берегу р. Сухоны, примерно в 500 м выше по течению от дер. Исады, находящейся на противоположной стороне реки (Вологодская обл., Великоустюгский р-н). Данные по геологическому строению, истории изучения и палеонтологическому составу комплекса местонахождения Исады подробно представлены в литературе (Синиченкова, 2013; Aristov et al., 2013; Karasev, 2013; Kopylov, 2013; Arefiev et al., 2015).
Отметим, что комплекс стрекоз из Исад довольно необычен, так как представлен преимущественно мелкими кеннедиеобразными формами, сближаемыми по облику и жилкованию крыла с родом Progoneura, который наиболее обилен в нижнепермских отложениях местонахождения Мидко (Василенко, 2013; Фелькер, Василенко, 2015). В то же время, найдены единичные остатки стрекоз, сходных с триасовыми протомирмелеонтоидами Terskeja Pritykina, 1981. Так, из 40 известных на сегодняшний день экземпляров стрекоз 14 экз. относится к крупным и среднеразмерным формам семейств Permepallagidae и Permagrionidae (по предварительным данным, один новый вид Permepallage и три – Permolestes), пять экз. – к описанному ниже новому виду рода Kennedya, 19 экз. – к небольшим прогоневроидным формам сем. Kennedyidae (не менее трех пока не описанных родов и шести видов) и два экз. – по предварительным данным, к протомирмелеонтоидам сем. Voltzialestidae (один пока не описанный род и вид).
Автор выражает искреннюю благодарность Р.А. Ракитову и Д.Е. Щербакову (ПИН РАН) за помощь в фотографировании материала на электронном сканирующем микроскопе TeScan Vega 3, а также Д.В. Василенко (ПИН РАН) за прочтение рукописи и ценные замечания.
МАТЕРИАЛ И МЕТОДЫ
Все описываемые в настоящей работе остатки собраны в период с 2010 по 2012 гг. экспедиционными отрядами лаборатории артропод ПИН РАН в серых алевролитах Мутовинской линзы (местонахождение Исады).
Изображения получены с помощью стереомикроскопа Leica M165C и камеры Leica DFC425. Плохо различимое под оптическим микроскопом строение дискоидальной области стебелька представлено на изображениях, полученных сканирующим микроскопом TeScan Vega 3 в режиме детектора BSE. При обработке изображений применялись графические редакторы Adobe Photoshop CS6 и Adobe Illustrator СС 2020. Уточнение морфологии крыльев и корректировка рисунка проводились с использованием стереомикроскопа Olympus SZX10.
Для большинства экземпляров предложены реконструкции полных крыльев. На рисунках восстановленные элементы жилкования показаны серым. При описании материала использованы следующие обозначения жилок: А – анальная жилка; An2 – вторая антенодальная поперечная жилка; Arc – дужка; Asn – утолщенная поперечная жилка, расположенная между R1 и R2 на интервале от дужки до узелка; br – опорная жилка птеростигмы; C – костальная жилка; cua – дистальная сторона sq; СuP – задняя кубитальная жилка; IR2, IR3 – интеркалярные радиальные жилки; МА – передняя медиальная жилка; MP – укороченная до размера поперечной жилка между MA и СuP, по-видимому, являющаяся сильно редуцированной задней ветвью медиальной жилки; N – узелок; n – нодальная жилка (опорная жилка узелка); Pn – постнодальные поперечные жилки; Psn – постсубнодальные поперечные жилки; Pt – птеростигма; q – дискоидальная ячейка; R1, R2, R3, R4 + 5 – ветви радиуса; Rs – радиальный сектор; Sc – субкостальная жилка; sn – субнодальная поперечная жилка; sq – субдискоидальная ячейка.
Типовой материал хранится в лаборатории артропод ПИН РАН, колл. 3840.
ОТРЯД ODONATA
ИНФРАОТРЯД PROTOZYGOPTERA
СЕМЕЙСТВО KENNEDYIDAE TILLYARD, 1925
Род Kennedya Tillyard, 1925
Kennedya suchonensis Felker, sp. nov.
Табл. VII , фиг. 1–3 (см. вклейку)
Рис. 1.
Kennedya suchonensis sp. nov., фотографии остатков: а – голотип ПИН, № 3840/2605; б – паратип ПИН, № 3840/2607; в – паратип ПИН, № 3840/1699; г – паратип ПИН, № 3840/5003; д – паратип ПИН, № 3840/1306; Россия, местонахождение Исады; северодвинский ярус, полдарсская свита. Масштабный отрезок соответствует 2 мм.

Рис. 2.
Kennedya suchonensis sp. nov.: a–д – прорисовки остатков: а – голотип ПИН, № 3840/2605; б – паратип ПИН, № 3840/5003; в – паратип ПИН, № 3840/1699; г – паратип ПИН, № 3840/2607; д – паратип ПИН, № 3840/1306; е–з – реконструированные крылья (восстановленные элементы жилкования показаны серым): е – голотип ПИН, № 3840/2605; ж – паратип ПИН, № 3840/1699; з – паратип ПИН, № 3840/2607; Россия, местонахождение Исады; северодвинский ярус, полдарсская свита. Обозначения названий жилок и ячеек указаны в работе. Масштабный отрезок соответствует 2 мм.
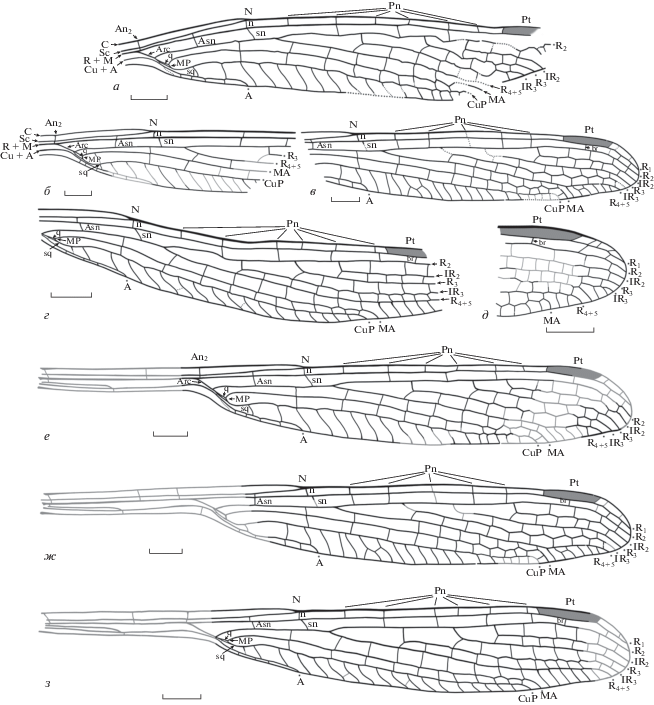
Рис. 3.
Строение дискоидальной области крыльев триасовых стрекоз рода Kennedya: a–д – K. carpenteri, экземпляры типовой серии: а, в – прорисовка и фотография паратипа ПИН, № 2555/616; б, г – прорисовка и фотография паратипа ПИН, № 2240/1798; д – фотография голотипа ПИН, № 2785/2; Киргизия, местонахождение Джайлоучо; ладинский–карнийский ярус; мадыгенская свита; е – фотография K. kedrovkensis, голотип ПИН, № 4887/144; Россия, местонахождение Бабий Камень; индский ярус, мальцевская свита. Обозначения названий жилок и ячеек указаны в работе. Масштабный отрезок соответствует 1 мм.

Название вида – от реки Сухоны, на берегу которой расположено местонахождение.
Голотип – ПИН, № 3840/2605, позитивный отпечаток средней части крыла (от основных структур стебелька до не полностью сохранившейся вершины); Вологодская обл., Великоустюгский р-н, левый берег р. Сухоны, обнажение напротив дер. Исады, местонахождение Исады; верхняя пермь, северодвинский ярус, путятинский горизонт, полдарсская свита, кичугская пачка.
Описание (рис. 1; 2; 4, а). An2 полная, расположена немного дистальнее базального края Arc, субкостальная ее часть без смещения продолжает костальную. N образован примерно равным изгибом C и Sc; полный по строению, по-видимому, расположен немного проксимальнее середины крыла. Перпендикулярная переднему краю крыла n расположена прямо под N. Заметно наклоненная sn находится прямо под n на голотипе (на паратипах ПИН, №№ 3840/1699, 2607 и 5003 немного смещена в дистальном направлении относительно нее). Немного базальнее N в поле между Sc и R1 находится одна практически вертикальная поперечная жилка, без смещения продолжающаяся в следующем поле между R1 и R2 – прямой Asn (на паратипе ПИН, № 3840/2607 Asn слегка наклонена в сторону основания крыла и расположена с небольшим дистальным смещением относительно поперечной в вышележащем поле). Pn жилок не менее пяти, Psn не менее трех, по-видимому, они могут находиться на разном расстоянии друг относительно друга. На голотипе и паратипе ПИН, № 3840/2607 чаще всего постнодальные (Pn – по 6 на обоих экземплярах) не продолжаются постсубнодальными (Psn – не менее 3 на голотипе и 4 на паратипе ПИН, № 3840/2607), в то же время на паратипе ПИН, № 3840/1699 они расположены попарно без смещения (Pn – 5, Psn – 5; первая Pn расположена слегка проксимальнее первой Psn). Pt узкая, трапециевидной формы, отношение ширины к длине – 1 : 7, передний и задний ее края прямые, параллельные друг другу; базальный и дистальный края наклонены в разных направлениях. Жилка br прямая, перпендикулярная R1 и R2, расположена немного проксимальнее уровня середины Pt. По-видимому, в поле между R1 и передним краем крыла дистальнее Pt могут присутствовать слегка наклоненные поперечные жилки (присутствуют на паратипе ПИН, № 3840/1699).
Arc неполный, средней длины, на голотипе равен длине переднего края q (на паратипе ПИН, № 3840/5003 – вдвое длиннее его). На голотипе расстояние между Asn и Arc равно расстоянию между Asn и N (на паратипе ПИН, № 3840/5003 – практически вдвое больше его); q закрытая, небольшого размера, треугольная по форме, дистальный ее край (МР) заметно утолщен и наклонен по направлению к основанию крыла. Субдискоидальная ячейка (sq) четырехугольная (так как основание CuP соединено с А слабой поперечной жилкой в месте ответвления R2). Дистальный край sq (cua) слегка изогнут и наклонен в сторону основания крыла.
Основание R4 + 5 расположено заметно дистальнее N, расстояние между ними практически вдвое меньше расстояния между Asn и N; основание IR3 находится между второй и третьей Pn, немного проксимальнее середины расстояния между N и Pt; основание R3 расположено в дистальной трети того же расстояния. Основание IR2 находится заметно проксимальнее Pt: на голотипе и паратипе ПИН, № 3840/2607 на полторы ячейки проксимальнее Pt, на паратипе ПИН, № 3840/1699 – на одну ячейку. Поле между А и задним краем крыла очень узкое: примерно на уровне N или немного проксимальнее его – в 5 раз уже вышерасположенного поля между A и CuP на голотипе и паратипе ПИН, № 3840/5003; в четыре раза уже того же поля на паратипе ПИН, № 3840/1699; в 6 раз уже на паратипе ПИН, № 3840/2607. Вершина А расположена практически на уровне N: немного проксимальнее его на голотипе и, по-видимому, паратипе ПИН, № 3840/5003, и слегка дистальнее – на паратипах ПИН, №№ 3840/1699 и 2607. Вершина CuP находится немного проксимальнее уровня Pt. Вершина МА расположена под проксимальным концом Pt, кроме паратипа ПИН, № 3840/1306, где она находится под дистальной частью Pt. Вершины остальных основных продольных жилок, по-видимому, расположены дистальнее Pt (на большинстве экз., см. Изменчивость). Между А и задним краем крыла находятся не менее трех поперечных жилок, между вершинами А и CuP находятся не менее 15 ячеек. Между вершинами CuP и MA, МА и R4 + 5 и R4 + 5 и IR3 число ячеек может быть разным (см. Изменчивость). IR3, МА и А слегка зигзагообразные в дистальной части, остальные продольные жилки прямые.
Размеры в мм. Размеры представителей типовой серии K. suchonensis sp. nov. указаны в табл. 1.
Таблица 1.
Размеры представителей K. suchonensis sp. nov.: 1 – длина сохранившегося остатка (полная длина реконструированного крыла); 2 – максимальная ширина (немного проксимальнее уровня вершины CuP); 3 – узелковая ширина; 4 – расстояние от Arc до N; 5 – расстояние от N до Pt; 6 – расстояние от N до вершины крыла; 7 – длина Pt; 8 – ширина Pt
| Экз. ПИН, № | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|
| 3840/2605, голотип | 24.4 (35.0) | 4.5 | 3.8 | 6.1 | 14.3 | 18.1 | 2.2 | от 0.3 до 0.5 |
| 3840/2607 | 20.2 (30.8) | 4.0 | 2.8 | ~5.5 | 12.7 | ~16.8 | ~3.1 | от 0.2 до 0.5 |
| 3840/1699 | 24.2 (36.7) | 4.9 | 3.9 | ~7.0 | 14.8 | 20.6 | 3.4 | от 0.3 до 0.5 |
| 3840/1306 | 5.5 | – | – | – | – | – | 3.3 | от 0.3 до 0.5 |
| 3840/5003 | 19.6 | ~4.8 | 4.1 | 7.2 | – | – | – | – |
Изменчивость. На экземплярах типовой серии ПИН, №№ 3840/1699, 2605 и 2607 наблюдается различное количество и положение Pn и Psn жилок. Так, на голотипе имеется 6 Pn и не менее 3 Psn; на паратипе ПИН, № 3840/2607 – 6 Pn и 4 Psn, причем практически все эти поперечные жилки расположены не попарно; без смещения находится только первая Psn, продолжающая вторую Pn. В то же время, на паратипе ПИН, № 3840/1699 наблюдаются 5 Pn и 5 Psn, расположенные попарно, практически без смещения друг относительно друга.
Кроме того, на экземплярах с сохранившейся дистальной частью крыла (паратипы ПИН, №№ 3840/1306, 1699 и частично 3840/2607) замечено различное положение вершин продольных жилок и, следовательно, количество ячеек между ними. Так, на паратипах ПИН, №№ 3840/1699 и 2607 вершины CuP и МА сближены (расположены через одну ячейку) и находятся проксимальнее Pt или под ее проксимальным краем, вершины MA и R4+5 заметно удалены друг от друга (между ними расположено не менее шести ячеек), а между вершинами R4 + 5 и IR3 также присутствует только одна ячейка (эта часть сохранилась только на паратипе ПИН, № 3840/1699). В свою очередь, на паратипе ПИН, № 3840/1306, представленном дистальной оконечностью крыла, вершина МА находится под дистальной частью Pt и заметно удалена от вершины CuP (между ними насчитываются не менее пяти ячеек – установить точнее не позволяет сохранность), а расстояние между вершинами МА и R4 + 5 (три ячейки) примерно равно расстоянию между вершинами R4 + 5 и IR3 (четыре ячейки).
Отметим также, что размеры паратипа ПИН, № 3840/2607 немного меньше, по сравнению с остальными экземплярами типовой серии.
Сравнение. От всех видов рода Kennedya отличается строением дискоидальной области стебелька: закрытой, треугольной q, четырехугольной sq и соединением CuP с дистальным краем Arc. От K. carpenteri отличается меньшим количеством Pn и Psn, от всех остальных видов – большим количеством этих жилок. C K. carpenteri и K. tyulkinensis сходен длиной IR2, c K. azari и K. kedrovkensis – длиной А. Положением An2 новый вид сходен с видами K. mirabilis, K. azari, K. carpenteri и K. kedrovkensis, узостью поля между А и задним краем крыла – с K. tillyardi, K. fraseri, K. ivensis. От K. mirabilis, K. tillyardi, K. fraseri, K. pritykinae и K. azari новый вид отличается меньшими размерами, от K. tyulkinensis, K. kedrovkensis и K. gracilis – заметно большими размерами крыла.
Замечание. У всех паратипов, за исключением ПИН, № 3840/5003, отсутствует базальная часть крыла. Дистальная часть крыла хорошо сохранилась на паратипах ПИН, №№ 3840/1699 и 1306.
Материал. Пять изолированных крыльев разной сохранности из местонахождения Исады. Наиболее полную сохранность имеет голотип, на котором отсутствует только основание крыла и часть вершины; у паратипа ПИН, № 3840/2607 сохранилась середина крыла примерно от окончания стебелька до дистального края птеростигмы. Паратип ПИН, № 3840/1699 представляет собой около двух дистальных третей крыла; паратип ПИН, № 3840/5003 сохранился от дистальной части стебелька до узелка; паратип ПИН, № 3840/1306 представлен только вершиной крыла (примерно шестая часть от целого крыла).
ОБСУЖДЕНИЕ
Несмотря на заметное преобладание кеннедиид среди стрекоз в ориктоценозе местонахождения Исады, большинство из них относится к мелким прогоневроподобным формам, и только пять экз. несут характерные признаки рода Kennedya (“полный” N, длинную Pt, не полностью редуцированную А, относительно длинные жилки IRs и Rs). По облику и размерам крыла исадские кеннедии больше всего похожи на ладин–карнийских представителей K. carpenteri, имея наряду с относительно расширенным задним краем крыла заметный переход от суженного стебелька к основной крыловой пластинке. По жилкованию крыла новый вид, по-видимому, также следует сближать именно с K. carpenteri, так как от других известных видов кеннедий их отличают такие признаки: присутствие длинных Rs, IRs и учащенное поперечное жилкование, в том числе увеличенное количество Pn и Psn.
Однако самым необычным признаком, до этого не наблюдавшимся как у кеннедиид, так и у всех остальных представителей инфраотряда Protozygoptera, является весьма специфичное строение дискоидальной области стебелька. Оно проявляется в присутствии уменьшенной, закрытой и треугольной по форме q. Причем нижележащая sq видоизменена на том же интервале (табл. VII ). В обычном для протозигоптер состоянии ее проксимальный край образуется плавным разветвлением ствола Cu+A на продольные жилки CuP и А, так что sq имеет треугольную форму. В случае исадских кеннедий, в указанной области между продольными жилками CuP и А, по-видимому, присутствует слабая поперечная жилка, изменяющая форму sq на четырехугольную (табл. VII , фиг. 1а–1е, фиг. 3а; рис. 4, а – отмечена серой стрелкой).
Рис. 4.
Строение дискоидальной области стебелька: a – Kennedya suchonensis sp. nov.; q закрытая, серой стрелкой показана поперечная жилка между CuP и А, преобразующая проксимальный край sq; б – K. kedrovkensis с типично кеннедиидным строением указанной области: q открытая, sq треугольная. Обозначения названий жилок и ячеек указаны в работе. Масштабный отрезок соответствует 2 мм.
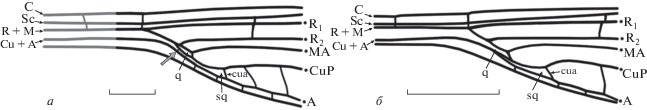
У известных протозигоптер связь между R + M и СuP представлена в стебельке в виде заметно редуцированной (практически до состояния поперечной) жилки MP. Однако на исадском материале (в особенности на паратипе ПИН, № 3840/2605) происходит двойное соединение CuP и R + M. Причем проксимальная часть этого соединения переходит из R + M, как обычное ответвление продольной жилки от общего ствола, а не путем образования поперечной жилки. Сформированная таким образом закрытая q, по-видимому, является особенностью исадских кеннедий (рис. 4, а). Расположение составляющих ее края продольных жилок (CuP и R + M) может быть объяснено заметным сужением стебелька при переходе к основной крыловой пластинке. Такую конфигурацию структур стебелька нельзя считать артефактом сохранности, так как она присутствует как минимум на трех экземплярах – паратипе ПИН, № 3840/5003 (практически не гофрированная часть стебелька; табл. VII , фиг. 1а–1е), голотипе ПИН, № 3840/2605 (жилки дискоидальной области расположены в разных плоскостях, но при “развертке” образуют сходный с плоским крылом рисунок; табл. VII , фиг. 2а, 2б) и паратипе ПИН, № 3840/2607 (присутствует дискоидальная область, Arc не сохранился; табл. VII , фиг. 3а).
Подобное строение дискоидальной и субдискоидальной областей отмечается на K. carpenteri из среднего–верхнего триаса Киргизии, паратип ПИН, № 2555/616. На указанном экземпляре наблюдаются схожие с исадскими признаки: закрытая q и соединение CuP и R + M в дискоидальной зоне (рис. 3, а, в). Однако здесь они, возможно, обусловлены аберрантностью экземпляра, так как на голотипе того же вида представлено вполне “нормальное” кеннедиидное строение стебелька (рис. 3, д), либо диагенетическими преобразованиями, также прослеживающимися на других экземплярах типовой серии K. carpenteri из этого же местонахождения. Так, на экз. ПИН, № 2240/1798 присутствует видоизмененная sq, дистальный край которой (cua) заметно вытянут и сильно наклонен. Его вид вполне соотносится со слегка сглаженной формой заднего края крыла, указывающей на произошедшую деформацию экземпляра (рис. 3, б, г). Помимо этого, промежуточный по возрасту между исадскими и мадыгенскими формами индский вид K. kedrovkensis Felker, 2021 из сибирского местонахождения Бабий Камень характеризуется типично кеннедиидным строением интересующей области: базально открытой трапециевидной q и треугольной sq с относительно короткой и слегка наклоненной cua (рис. 3, е; 4, б) (Фелькер, 2021).
Следует отметить, что базально открытая q – один из признаков инфраотряда (при первоописании – подотряда) Protozygoptera, установленный еще Р. Тильярдом и позже подтвержденный Ф. Карпентером на множестве примеров из раннепермских местонахождений формации Веллингтон (Tillyard, 1925; Carpenter, 1931; Tillyard, Fraser, 1939). Открытая дискоидальная ячейка характерна и для настоящих архизигоптер (по-видимому, более продвинутых, чем протозигоптеры) – сем. Protomyrmeleontidae и Batkeniidae (Притыкина, 1981; Nel et al., 2005), Кроме того, именно такое ее строение наблюдается у некоторых раннемеловых Coenagrionidae (Cretacoenagrion alleni Jarzembowski, 1990) и даже на передних крыльях современных Hemiphlebiidae (Hemiphlebia mirabilis Selys, 1869) и Synlestidae (Chorismagrion risi Morton, 1914) (Jarzembowski, 1990). Основываясь на этих данных, Ф. Фрэзер полагал, что открытая дискоидальная ячейка является плезиоморфным признаком, в основном проявляющимся у ранних форм равнокрылых и остающимся как реликтовый признак только у “живых ископаемых”, наподобие H. mirabilis (Fraser, Tillyard, 1957; Jarzembowski, 1990). Противоположного мнения относительно этой структуры придерживался Ф. Карле, который полагал, что открытая q – апоморфный признак, который мог возникать независимо в разных группах зигоптер (Carle, 1982; Jarzembowski, 1990). Учитывая особенности исадских кеннедий, а также доступный для изучения материал, представленный ранне- и среднепермскими формами из Чекарды и Сояны (все крылья с открытой q), мы полагаем толкование открытого строения q в качестве примитивного признака более приемлемым, во всяком случае, для изученных кеннедиид (рис. 4).
Список литературы
Василенко Д.В. Стрекозы семейства Kennedyidae Tillyard, 1925 (Insecta: Odonata) в пермских одонатофаунах // Гидроэнтомология в России и сопредельных странах: Матер. V Всеросс. симп. по амфибиотическим и водным насекомым. Ярославль: Филигрань, 2013. С. 32–35.
Притыкина Л.Н. Новые триасовые стрекозы Средней Азии // Новые ископаемые насекомые с территории СССР / Ред. Вишнякова В.Н., Длусский Г.М., Притыкина Л.Н. М.: Наука, 1981. С. 5–42.
Синиченкова Н.Д. Новые поденки (Insecta: Ephemerida = Ephemeroptera) из верхнепермского местонахождения Исады, север европейской части России // Палеонтол. журн. 2013. № 2. С. 35–38. https://doi.org/10.7868/S0031031X13020141
Фелькер А.С. Первые стрекозы семейства Kennedyidae (Odonata: Protozygoptera) из пограничных пермо-триасовых отложений Кузнецкого бассейна России // Палеонтол. журн. 2021. № 2. P. 52–59.
Фелькер А.С., Василенко Д.В. Уникальный комплекс стрекоз местонахождения Исады (верхняя пермь Вологодской области) // Современная палеонтология: классические и новейшие методы. Тез. докл. XII Всеросс. науч. школы молодых ученых-палеонтологов. М.: ПИН РАН, 2015. С. 39–40.
Фелькер А.С., Василенко Д.В. К вопросу о систематике пермских и триасовых кеннедиидообразных стрекоз // Современная палеонтология: классические и новейшие методы. Тез. докл. XIV Всеросс. науч. школы молодых ученых-палеонтологов. М.: ПИН РАН, 2017. С. 36.
Arefiev M.P., Golubev V.K., Kuleshov V.N., Pokrovsky B.G. Paleogeographical reorganization of the Moscow syneclise during the Severodvinian (Capitanian–Wuchiapingian) time based on isotopic (δ13C and δ18O) and paleontological data, Permian period, East European platform // Paleontol. J. 2015. V. 49. № 11. P. 1206–1227. https://doi.org/10.1134/S0031030115110027
Aristov D.S., Bashkuev A.S., Vassilenko D.V. Locality of Late Permian animals and plants Mutovino (=Isady) on the Sukhona River in Vologda Region: Review of the Entomofauna // Fossil Insects of the Middle and Upper Permian of European Russia / Ed. Aristov D.S. N.Y., 2013. P. 658–667 (Paleontol. J. V. 47. № 7). https://doi.org/10.1134/S003103011307001
Carle F.L. The wing homologies and phylogeny of the Odonata: a continuing debate // Soc. Int. Odonatol. Rapid. Comm. 1982. V. 4. P. 1–66.
Carpenter F.M. The Lower Permian insects of Kansas. Part 2. The orders Paleodictyoptera, Protodonta and Odonata // Amer. J. Sci. Ser. 5. 1931. V. 21. P. 97–139. https://doi.org/10.2475/ajs.s5-21.122.97
Carpenter F.M. The Lower Permian insects of Kansas. Part 6. Delopteridae, Protelytroptera, Plectoptera and a new collection of Protodonata, Odonata, Megasecoptera, Homoptera and Psocoptera // Proc. Amer. Acad. Arts & Sci. 1933. V. 68. № 11. P. 411–505. https://doi.org/10.2307/20022959
Carpenter F.M. The Lower Permian insects of Kansas. Part 8. Additional Megasecoptera, Protodonata, Odonata, Homoptera, Psocoptera, Protelytroptera, Plectoptera, and Protoperlaria // Proc. Amer. Acad. Arts & Sci. 1939. V. 73. № 3. P. 29–70. https://doi.org/10.2307/25130151
Carpenter F.M. Lower Permian insects from Oklahoma. Part 1. Introduction and the orders Megasecoptera, Protodonata, and Odonata // Proc. Amer. Acad. Arts & Sci. 1947. V. 76. № 2. P. 25–54. https://doi.org/10.2307/20023497
Felker A.S. New damselflies of the family Kennedyidae from Permian deposits of European Russia // Paleontol. J. 2020. V. 54. № 7. P. 734–742.
Fraser F.C., Tillyard R.J. A reclassification of the order Odonata. Sydney: R. Zool. Soc. N.S.W., 1957. 133 p.
Golubev V.K. Locality of Late Permian animals and plants Mutovino (=Isady) on the Sukhona River in Vologda Region: Historical geology // Fossil Insects of the Middle and Upper Permian of European Russia / Ed. Aristov D.S. N.Y., 2013. P. 651–658 (Paleontol. J. V. 47. № 7). https://doi.org/10.1134/S003103011307001
Jarzembowski E.A. Early Cretaceous zygopteroids of southern England, with the description of Cretacoenagrion alleni gen. nov., spec. nov. (Zygoptera: Coenagrionidae; “Anisozygoptera”: Tarsophlebiidae, Euthemistidae) // Odontologica. 1990. V. 19. № 1. P. 27–37.
Karasev E.V. Locality of Late Permian animals and plants Mutovino (= Isady) on the Sukhona River in Vologda Region: Review of Plants // Fossil Insects of the Middle and Upper Permian of European Russia / Ed. Aristov D.S. N.Y., 2013. P. 667–669 (Paleontol. J. V. 47. № 7). https://doi.org/10.1134/S003103011307001
Kopylov D.S. Locality of Late Permian animals and plants Mutovino (=Isady) on the Sukhona River in Vologda Region: History of Studies // Fossil Insects of the Middle and Upper Permian of European Russia / Ed. Aristov D.S. N.Y., 2013. P. 649–651 (Paleontol. J. V. 47. № 7). https://doi.org/10.1134/S003103011307001
Martynov A.V. New Permian insects from Tikhie Gory, Kazan province. 1. Palaeoptera // Тр. Геол. музея АН СССР. 1930. С. 69–86.
Nel A., Petrulevicius Ju.F., Martínez-Delclòs X. New Mesozoic Protomyrmeleontidae (Insecta: Odonatoptera: Archizygoptera) from Asia with a new phylogenetic analysis // J. Syst. Palaeontol. 2005. V. 3. № 2. P. 187–201. https://doi.org/10.1017/S1477201905001549
Nel A., Bechly G., Prokop J. et al. Systematics and evolution of Paleozoic and Mesozoic damselfly like Odonatoptera of the Protozygopteran grade // J. Paleontol. 2012. V. 86. № 1. P. 81–104. https://doi.org/10.2307/41409133
Rasnitsyn A.P., Aristov D.S., Rasnitsyn D.A. Insects of the Permian and Early Triassic (Urzhumian–Olenekian Ages) and the Problem of the Permian–Triassic Biodiversity Crisis // Fossil Insects of the Middle and Upper Permian of European Russia / Ed. Aristov D.S. N.Y., 2013. P. 793–823 (Paleontol. J. V. 47. № 7). https://doi.org/10.1134/S003103011307001
Tillyard R.J. Kansas Permian insects. Part 5. The orders Protodonata and Odonata // Amer. J. Sci. Ser. 5. 1925. V. 10. P. 41–73. https://doi.org/10.2475/ajs.s5-10.55.41
Tillyard R.J., Fraser F.C. A reclassification of the order Odonata based on some new interpretations of the venation of the dragonfly wing. Part II. The suborder Zygoptera (continued), and Protanisoptera // Austr. Zool. 1939. V. 9. P. 195–221.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал