

Палеонтологический журнал, 2021, № 4, стр. 35-40
Новые данные о меловых ручейниках семейства Vitimotauliidae (Insecta: Trichoptera) из Бурятии (местонахождение Хасурты)
И. Д. Сукачева a, Д. С. Аристов a, b, *
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Череповецкий государственный университет
Череповец, Россия
* E-mail: danil_aristov@mal.ru
Поступила в редакцию 13.08.2020
После доработки 16.10.2020
Принята к публикации 16.10.2020
Аннотация
Описан новый представитель семейства Vitimotauliidae, Purbimodus khramovi sp. nov. На основании нового материала переописаны род Purbimodus Sukatsheva et Jarzembowski, 2001 и вид P. parvulus Sukatsheva et Aristov, 2020 из нижнего мела России (Республика Бурятия, местонахождение Хасурты). Приведены уточненные определительные таблицы родов семейства Vitimotauliidae и видов рода Purbimodus. Геологический возраст местонахождения Хасурты определяется не моложе раннего мела.
Новый материал, описываемый в настоящей работе, собран экспедиционными отрядами Палеонтологического ин-та им. А.А. Борисяка РАН (ПИН) 2014 и 2019 гг. из нижнемелового местонахождения Хасурты (Россия, Республика Бурятия, Закаменский р-н, окрестности хут. Хасурты, в 25 км восточнее г. Закаменска). Ручейники, как и другие ископаемые насекомые Хасурты, происходят из отложений гусиноозерской серии и датируются ранним мелом (Kopylov, Rasnitsyn, 2020).
Типовой материал хранится в коллекции лаб. артропод ПИН РАН.
Авторы выражают глубокую благодарность А.П. Расницыну и А.Г. Пономаренко (ПИН РАН), В.Д. Иванову и С.И. Мельницкому (СПбГУ) за ценные советы и консультации, а также Н.С. Гороховой (ООО “Издательство АСТ”) за техническую помощь. Работа поддержана Программой Президиума РАН “Эволюция органического мира. Роль и влияние планетарных процессов” и грантом РФФИ № 18-04-00322.
Vitimotauliidae Sukatsheva, 1968 – это наиболее широко распространенное в нижнем мелу семейство ископаемых ручейников. Оно известно из Южной Сибири, Забайкалья, Монголии, Китая и Южной Англии (Сукачева, Аристов, 2020). Как отдельное семейство оно было выделено на основании жилкования передних крыльев, сохранившихся достаточно хорошо, в отличие от морфологических признаков головы и тела. В 2009 г. из местонахождения Исянь (Китай) в семействе Vitimotauliidae был описан род Sinemodus Wang, Liang, Ren et Shin, 2009 (Wang et al., 2009). Благодаря хорошей сохранности материала, в описание семейства и нового рода, помимо рассмотрения жилкования крыльев, авторами были добавлены морфологические детали головы, груди и ног. В расширенное переописание семейства Vitimotauliidae, данное китайскими учеными, нами добавлены новые детали, полученные благодаря сборам 2019 г. в Хасурты и опубликованые нами ранее (Сукачева, Аристов, 2020).
К настоящему времени семейство Vitimotauliidae насчитывает 29 видов пяти родов (см. определительную таблицу родов). Интересно, что Vitimotauliidae известны только из Северного полушария. В Южном полушарии не сделано ни одной находки, хотя в таких нижнемеловых местонахождениях ископаемых насекомых, как Кунварра (Австралия) и Сантана (Бразилия), они могли бы присутствовать. Тем более, что из этих местонахождений известны находки ручейников (Jell, Duncan, 1986; Grimaldi, 1990; Bechly, 2007). Важно, что в нижнем мелу Южного Полушария пока не найдены передвижные домики ручейников, широко представленные в Евразии, где они часто сопутствуют находкам имаго Vitimotauliidae.
Определительная таблица родов семейства Vitimotauliidae
1 (2) Развилок F5 расположен на одном уровне с развилком F1. Развилки F3 и F4 короткие, начинаются на границе четвертой четверти длины крыла............................................................. ...................Sinemodus Wang, Liang et Ren, 2009.
2 (1) Развилок F5 расположен проксимальнее развилка F1. Развилки F3 и F4 длинные, начинаются в третьей четверти длины крыла.
3 (8) Насекомые крупные (длина передних крыльев 10–13 мм), с темным рисунком по краям крыльев и по вершинам апикальных развилков.
4 (7) Ячея MC короче ячеи DC в 1.4–1.7 раз.
5 (6) Субкостальное поле равной ширины с костальным. Ячея MC в 1.7 раз короче ячеи DC. Ячея TC короткая, в 1.3 раза короче ячеи MC. CuP и A1 впадают в задний край крыла проксимальнее середины длины крыла........................ ..................Burimodus Sukatsheva et Aristov, 2020
6 (5) Субкостальное поле уже костального. Ячея MC в 1.4–1.5 раза короче ячеи DC. Ячея TC длинная, намного длиннее ячеи MC. CuP и A1 впадают в задний край крыла на середине его длины или немного дистальнее…...................... ..............................Multimodus Sukatsheva, 1968.
7 (4) Ячея MC в 1.2 раза короче ячеи DC................. ............................Vitimotaulius Sukatsheva, 1968.
8 (3) Насекомые мелких и средних размеров (6–11 мм). Рисунка на крыльях нет или он в виде темной окраски со светлыми пятнами по всему крылу.................................................................. .....Purbimodus Sukatsheva et Jarzembowski, 2001.
Список родов семейства Vitimotauliidae с местонахождениями
Sinemodus Wang et al., 2009; нижний мел, Исянь, Ляонин (Китай).
Burimodus Sukatsheva et Aristov, 2020; нижний мел, Хасурты (Бурятия, Россия).
Multimodus Sukatsheva, 1968; нижний мел, Исянь (Ляонин, Китай), Бон-Цаган (Монгольский Алтай, Монголия), Байса, Бурея, Хасурты (Бурятия, Россия); нижняя юра, Бичектуй (Бурятия, Россия).
Vitimotaulius Sukatsheva, 1968; верхняя юра, Унда; нижний мел, Байса, Ундурга (Забайкальский край, Россия).
Purbimodus Sukatsheva et Jarzembowski, 2001; нижний мел, Южная Англия, Хасурты (Бурятия, Россия).
Большое разнообразие и обилие представителей Vitimotauliidae до сих пор встречалось только в таких крупнейших меловых лагерштеттах, как Бон-Цаган (Баацагаан, Монголия) и, особенно, Байса. В Бон-Цагане в нижнем мелу наблюдается достаточно плоский рельеф с постепенной сменой речных и озерных отложений, а Байсинское местонахождение образовалось на месте межгорной впадины. Всего в местонахождении Байса было собрано более 1000 экз. ручейников, из них более 300 экз. имаго и более 700 экз. личиночных домиков. Кроме того, в небольшом количестве найдены куколки и личинки ручейников. Лагерштетт Хасурты не уступает ни по обилию, ни по разнообразию материала. В этом местонахождении, как и в Байсе, имеются серии экземпляров, принадлежащих одному виду. К настоящему времени из Хасурты известно более 6000 экз. ископаемых насекомых (Синиченкова, 2011; Сукачева, Василенко, 2019; Сукачева, Аристов, 2020; Kopylov et al., 2020). Ручейников из Хасурты найдено 241 экз. из 15 семейств [из которых имаго более 170 экз., 41 экз. представлен преимагинальными стадиями (куколки и передвижные личиночные домики)]. Интересно, что в Байсе домиков найдено вдвое больше, чем имаго, а в Хасурты домиков в четыре раза меньше, чем имаго. Это связано, скорее всего, с экологическими и ландшафтными условиями, т.к. в Хасурты не было обычных для мезозойских озер плавающих растительных агрегатов (матов), на которых могли обитать и широко разноситься личинки ручейников в своих домиках, как это встречается у современных лимнефилид. При разрушении матов домики падали и захоранивались на дне водоема (Kopylov et al., 2020). Отсутствие матов также объясняет малочисленность домиков в глубоких частях озера, где образовывались отложения лагерштетта Хасурты. Скорее всего, в данном случае многочисленные витимотаулииды выводились по большей части из личинок, развивавшихся в близлежащих реках, текущих по довольно залесенным предгорьям, а затем имаго ветром переносились к озеру, где и захоранивались.
По результатам изучения геологического строения и фауны беспозвоночных можно судить достаточно обоснованно о палеоэкологии местонахождения Хасурты. Скорее всего, оно находилось в небольшой долине между низкими холмами с крутыми склонами вблизи озера. По остаткам фауны насекомых и остаткам растительности можно предположить прохладные условия существования в сезонном гумидном климате (Kopylov et al., 2020).
Полученный новый материал из Хасурты позволяет дать более развернутое описание рода Purbimodus и подробное переописание вида P. parvulus Sukatsheva et Aristov, 2020, что необходимо для обоснованного сравнения его с новым видом Purbimodus.
ОТРЯД TRICHOPTERA
СЕМЕЙСТВО VITIMOTAULIIDAE SUKATSHEVA, 1968
Род Purbimodus Sukatsheva et Jarzembowski, 2001
Purbimodus: Sukatsheva, Jarzembowski, 2001, c. 688; Сукачева, Аристов, 2020 с. 56.
Типовой вид – Purbimodus minor Suka-tsheva et Jarzembowski, 2001.
Диагноз. Насекомые средних размеров, передние крылья 6–11 мм. Костальное поле шире субкостального. Sc средней длины, почти всегда оканчивается в третьей четверти длины крыла на переднем крае. Ячея DC, как правило, больше ячеи MС, редко равна ей по длине и начинается всегда во второй четверти длины крыла. R без развилка. Апикальные развилки F1 и F2 длинные и начинаются на одном или почти на одном уровне. Развилки F3 и F4 также могут начинаться на одном уровне, или F3 начинается дистальнее F4. Развилок F5 расположен проксимальнее начала развилка F1. Ячеи DC, MC и TC закрыты. CuP и A1 впадают в задний край крыла в середине его длины или вблизи нее. CuP перед окончанием почти всегда образует коленчатый изгиб.
Видовой состав. Шесть видов из нижнего мела Забайкальского края (Россия) и Южной Англии (см. определительную таблицу видов рода).
Сравнение. См. определительную таблицу родов семейства Vitimotauliidae.
Определительная таблица видов рода Purbimodus Sukatsheva et Jarzembowski, 2001
1 (4) CuP и A1 впадают в задний край крыла в разных точках.
2 (3) RS в 1.2 раза короче ячеи DC. Ячея DC немного длиннее ячеи MC..................................... .....P. rasnitsyni Sukatsheva et Jarzembowski, 2001.
3 (2) RS более чем на половину короче ячеи DC. Ячея DC равна по длине ячее MC...................... .........P. medius Sukatsheva et Jarzembowski, 2001.
4 (1) CuP и A1 впадают в задний край крыла в одной точке.
5 (6) Sc впадает в передний край крыла значительно дистальнее вершин развилков F1 и F2........... ..........P. minor Sukatsheva et Jarzembowski, 2001.
6 (5) Sc впадает в передний край крыла почти на уровне вершин развилков F1 и F2.
7 (8) Ячея DC больше ячеи MC в 1.5 раза. Поперечная жилка cua2–cup длинная........................ ........P. saxosus Sukatsheva et Jarzembowski, 2001.
8 (7) Ячея DC больше ячеи MC в 1.1–1.4 раза или равна ей.
9 (10) Вершины развилков F3 и F4 расположены на одном уровне...................P. khramovi sp. nov.
10 (9) Вершины развилков F3 и F4 расположены на разных уровнях.............................................. ..................P. parvulus Sukatsheva et Aristov, 2020
Purbimodus parvulus Sukatsheva et Aristov, 2020
Purbimodus parvulus: Сукачева, Аристов, 2020, с. 58, рис. 1, б, табл. VI , фиг. 2.
Рис. 1.
Purbimodus parvulus Sukatsheva et Aristov, 2020, голотип ПИН, № 5026/1498: а – фотография, б – прорисовка жилкования. Длина масштабной линейки 1 мм.
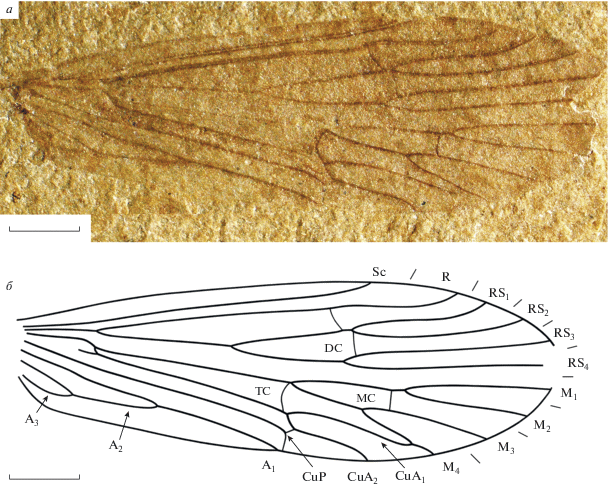
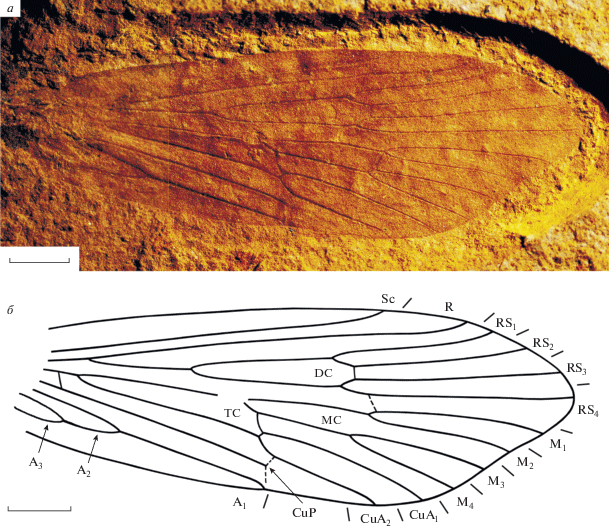
Рис. 2.
Purbimodus khramovi sp. nov.; голотип ПИН, № 5340/2281: а – фотография, б – прорисовка жилкования. Длина масштабной линейки 1 мм.
Голотип – ПИН, № 5026/1498, прямой и обратный отпечатки полного переднего крыла; Россия, Республика Бурятия, местонахождение Хасурты; нижний мел, гусиноозерская серия.
Описание (рис. 1, а, б). Крылья довольно узкие, длина больше ширины в 2.4–3 раза. Передний край крыла прямой, вершина закруглена. Наибольшая ширина крыла на уровне впадения CuA2 в задний край крыла. Sc короткая, прямая, заканчивается в районе середины третьей четверти длины крыла несколько дистальнее вершин развилков F1 и F2. Субкостальное поле в 1.1–1.3 раза уже костального. R с длинным, слабым вершинным ковшеобразным изгибом, иногда с горбообразной выпуклостью на уровне вершины F1. Имеется слабая косая жилка r-rs1 + 2 немного проксимальнее вершины F1. Ячея DC равна по длине или короче своего ствола, длиннее или равна ячее MC и начинается в районе середины второй четверти длины крыла. F1 начинается на уровне или почти на уровне развилка F2. Поперечная жилка rs1 + 2–rs3 прямая или очень слабо изогнута. Жилка rs4–m1 + 2 прямая. Развилки F3 и F4 короче развилков F1 и F2, широкие. Вершины развилков F3 и F4 начинаются на разных уровнях. F3 со стебельком или прямая поперечная m1 + 2–m3 сдвинута к вершинам развилков. Ячея TC длинная. Поперечная жилка m-cua дугообразно изогнутая. Жилка m-cu образует ложное основание переднего кубитуса. CuP и A1 впадают в задний край крыла в одной точке примерно на середине длины крыла. Поперечная cua2-cup четкая, довольно короткая. CuP коленообразно изгибается к своему окончанию. A2 довольно короткая, составляет почти половину длины A1. Анальное поле различной ширины (с учетом паратипов).
Размеры в мм: длина крыла 6–10, ширина 2.7–3.3.
Сравнение. См. определительную таблицу видов.
Материал. Кроме голотипа, еще девять экземпляров ПИН, №№ 5340/1462, 1465, 2280, 2284, 2290, 2297, 2300, 2304, 2305 из того же местонахождения.
Purbimodus khramovi Sukatsheva et Aristov, sp. nov.
Название вида – в честь палеоэнтомолога А.С. Храмова.
Голотип – ПИН, № 5340/2281, прямой и обратный отпечатки переднего крыла; Россия, Республика Бурятия, местонахождение Хасурты; нижний мел, гусиноозерская серия.
Описание (рис. 2, а, б). Крыло узкое, длина больше ширины в 2.4–3.3 раза. Передний край крыла прямой, вершина вытянута. Наибольшая ширина крыла на уровне впадения CuA2 в задний край крыла. Sc короткая, прямая, заканчивается на уровне третьей четверти длины крыла, несколько дистальнее вершин развилков F1 и F2. Субкостальное поле в 1.6–2.6 раза уже костального. R прямой, с очень слабым вершинным ковшеобразным изгибом и иногда с горбообразной выпуклостью на уровне вершины F1. Поперечная жилка r-rs1 прямая или слабо косая. Ячея DC в 1.1–1.6 раза больше ствола RS, в 1.1–1.4 раза больше ячеи MC и начинается вблизи середины второй четверти длины крыла. Развилки F1 и F2, так же как F3 и F4, начинаются на одном уровне. Поперечные жилки rs2-rs3 и rs1-m1 + 2 прямые. Развилки F3 и F4 короче развилков F1 и F2, довольно узкие. Все апикальные развилки сидячие. Поперечная жилка m1 + 2-m3 прямая или слабокосая, поперечная m3 + 4-cua прямая или слабо дугообразно изогнута. Ячея ТС длинная. CuP и A1 впадают в задний край крыла в одной точке на середине его длины. Поперечная cua2-cup длинная. CuP резко изгибается к своему окончанию, десклеротизуясь и образуя коленообразный изгиб. A2 довольно короткая, составляет почти половину длины A1. Анальное поле довольно узкое.
Размеры в мм: длина крыльев 5.7–11, ширина 2.1–3.7.
Сравнение. См. определительную таблицу видов.
Материал. Кроме голотипа, паратипы ПИН, №№ 5026/586; 5340/2279, 2283, 2291, 2295, 2325 из того же местонахождения.
Список литературы
Синиченкова Н.Д. Новые веснянки (Insecta: Perlida = Plecoptera) из позднемезозойского местонахождения Хасуртый, Забайкалье // Палеонтол. журн. 2011. № 5. С. 76–81.
Сукачева И.Д. Мезозойские ручейники (Trichoptera) Забайкалья // Палеонтол. журн. 1968. № 2. С. 59–75.
Сукачева И.Д., Аристов Д.С. Новые ручейники (Insecta: Trichoptera) семейств Vitimotauliidae и Philopotamidae из местонахождения Хасурты (нижний мел России) с кратким обзором ископаемой мировой фауны // Палеонтол журн. 2020. № 4. С. 54–63.
Сукачева И.Д., Василенко Д.В. Новые ручейники семейства Dysoneuridae (Insecta, Trichoptera) из мезозоя Забайкалья (местонахождение Хасуртый) // Палеонтол. журн. 2019. № 5. С. 63–68.
Bechly G. Trichoptera and Lepidoptera: caddisflies and butterflies // The Crato Fossil Beds of Brazil. Window into an Ancient World. Cambridge: Cambridge Univ. Press, 2007. P. 387–393.
Grimaldi D.A. (ed.) Insects from the Santana Formation, Lower Cretaceous, of Brasil // Bull. Amer. Mus. Natur. Hist. 1990. V. 195. 191 p.
Jell P.A., Duncan P.M. Invertebrates, mainly insects, from the freshwater, Lower Cretaceous, Koonwarra Fossil Bed (Korumburra Group), south Gippsland, Victoria // Mem. Assoc. Australas. Palaeontol. 1986. № 3. P. 111–205.
Kopylov D.S., Rasnitsyn A.P. (eds.) The Khasurty fossil insect Lagerstätte // Paleontol. J. 2020. V. 54. № 11. P. 1221–1394.
Sukatsheva I.D., Jarzembowski E.A. Fossil caddisflies (Insecta: Trichoptera) from the Early Cretaceous of Southern England. II // Cret. Res. 2001. V. 22. P. 685–694.
Wang M., Liang J., Ren D., Shin Ch. New fossil Vitimotauliidae (Insecta: Trichoptera) from the Jehol Biota of Liaoning Province, China // Cret. Res. 2009. V. 30. P. 592–598.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал