

Палеонтологический журнал, 2021, № 4, стр. 55-65
Новый род слоновых химер (Holocephali: Callorhinchidae) из поздней юры Центральной России
Е. В. Попов a, b, *, К. М. Шаповалов c
a Саратовский государственный университет им. Н.Г. Чернышевского
Саратов, Россия
b Казанский (Приволжский) федеральный университет
Казань, Россия
c Независимый исследователь
Москва, Россия
* E-mail: elasmodus74@gmail.com
Поступила в редакцию 02.02.2020
После доработки 17.07.2020
Принята к публикации 17.07.2020
Аннотация
По отдельным мандибулярным, небным и сошниковой зубным пластинам из двух верхнеюрских местонахождений (средневолжский подъярус, зона virgatus) Москвы и Подмосковья описывается новый род слоновых химер Moskovirhynchus robustus gen. et sp. nov. Озубление нового рода морфологически близко к структуре зубной системы современных слоновых химер рода Callorhinchus Lacepède, 1798, но характеризуется мозаичным сочетанием примитивных (на небных пластинах) и продвинутых (на небных и мандибулярных пластинах) признаков – развитием на мандибулярной пластине асимметричной бифуркации срединного тритора и редуцированными триторами небной пластины (продвинутое состояние для каллоринхид) вместе с сильно развитой аборальной частью небной пластины (примитивное состояние). Это древнейший представитель Callorhinchidae s. s., известный по зубной системе.
Первые находки отдельных зубных пластин химеровых рыб (Chimaeroidei) в юрских отложениях Русской плиты были сделаны давно (Боголюбов, 1912), но их целенаправленный сбор и изучение началось лишь с конца XX в. (Аверьянов, 1992). Сейчас на этой территории известно 15 юрских местонахождений (все – в пределах европейской части России) из отложений средней (бат, келловей) и верхней (волжский ярус) юры (Попов, 2003, 2005; Попов, Безносов, 2006; Попов, Шаповалов, 2007). Наиболее многочисленна по числу известных находок и таксономически разнообразна келловейская ассоциация, включающая в себя шесть–семь родовых таксонов каллоринхид и “эдафодонтид” из семи местонахождений (Попов, 2005).
Из отложений бореального волжского яруса рассматриваемой территории известны редкие находки отдельных зубных пластин химероидов “Edaphodontidae” gen. et sp. indet. (возможно, таксон близкий к Elasmodus sp., Е.В. Попов, личн. набл.) и Ischyodus cf. schuebleri Quenstedt, 1858 [оба – (?) нижневолжский подъярус, разрез Каргорт в республике Коми: Попов, Безносов, 2006], Stoilodon sp. (зона nikitini средневолжского подъяруса разреза Городищи в Ульяновской обл.: Попов, Ефимов, 2012) и Brachymylus bogolubovi Averianov, 1992 (средневолжский подъярус Москвы: Боголюбов, 1912; Аверьянов, 1992). С территории Москвы и Подмосковья также были описаны отдельные представители комплекса из средневолжского (зона virgatus) подъяруса верхней юры (Попов, Шаповалов, 2007), включающего в себя двух каллоринхид – Callorhinchus sp. и Brachymulus sp. – и двух “эдафодонтид” – Ischyodus cf. townsendi Buckland, 1835 и Elasmodectes sp. (последний таксон может быть теперь переопределен как Stoilodon sp.; см. также: Попов, Ефимов, 2012, с. 74). Происходящий из этого комплекса Callorhinchus sp. переописывается в настоящей статье как новый род слоновых химер (Callorhinchidae s. s.) – Moskovirhynchus robustus gen. et sp. nov.
Изображенные зубные пластины хранятся в региональном музее землеведения Саратовского государственного ун-та (колл. СГУ № 155).
Авторы благодарны М.А. Рогову (ГИН РАН, Москва) за консультации по стратиграфии волжского яруса европейской части России; анонимным рецензентам – за ценные замечания по рукописи статьи, позволившие ее улучшить. Комплект зубных пластин современного вида Callorhinchus callorhynchus (экз. СГУ 155/103) был любезно предоставлен М.Е. Суаресом (Чили). Работа выполнялась при финансовой поддержке РФФИ (проекты №№ 10-05-00926 и 18-05-01045), а также за счет средств Программы стратегического академического лидерства Казанского (Приволжского) федерального университета.
МЕСТОНАХОЖДЕНИЯ И МАТЕРИАЛ
Описываемые зубные пластины нового рода слоновых химер происходят из двух местонахождений Центральной России в пределах Москвы и Московской области (рис. 1, а):
Рис. 1.
а – Схема расположения местонахождений с остатками Moskovirhynchus gen. nov. в Центральной России (Москва и Подмосковье); местонахождения: 1 – Кунцево; 2 – карьер ЛФР 7-2-бис; б – схема зонального расчленения волжского яруса европейской части России (по: Rogov, 2010, 2014; Киселев и др., 2018), стратиграфическое распространение нового рода химер показано цветом; в – корреляция зоны Virgatitus virgatus со стратиграфическими подразделениями Англии (по: Cope, 1993), Франции и Германии (по: Schweigert, 2007; Bulot et al., 2014).
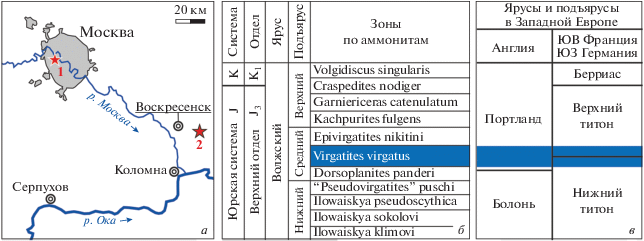
1. Кунцево. Овраги (N 55°44′33.5′′, E 37°26′27.6′′; N 55°44′40.3′′, E 37°26′16.8′′) на правом берегу р. Москвы у Кунцевского городища (Фили–Кунцевский парк) в Москве. Юрские отложения представлены здесь песчанистыми глинами и песками с фосфоритами и большим количеством остатков беспозвоночных средневолжского подъяруса (зоны virgatus и nikitini) (рис. 1, б). Выше залегают в целом сходные по строению, но более песчаные отложения верхневолжского подъяруса (зоны fulgens, catenulatum и nodiger) и готеривского яруса нижнего мела (Герасимов, 1971; Рогов, 2017; Тесакова, 2017). Близлежащие разрезы (Хорошево, Студеный овраг в Мневниках; оба ныне недоступны) по р. Москве как “классические” разрезы подмосковной юры известны с конца XVIII века (Стародубцева, 2006). Они неоднократно изучались геологами (Рулье, 1845; Trautschold 1861; Траутшольд, 1870; Никитин, 1883; Розанов, 1912; и др.) и были обычными объектами геологических экскурсий (Павлов, 1946; Апродов, Апродова, 1963; Gerasimov, 1967). Зубные пластины химер собраны одним из авторов (К.М. Шаповалов) в 1990-е гг. путем промывки на ситах темно-серых глинистых песков кровли зоны virgatus, переполненных окаменелостями (“ракушняк”) (верхняя часть слоя 1 в: Тесакова, 2017).
Кроме нового вида химер, из слоя происходят зубные пластины Brachymylus sp. (экз. СГУ № 155/78) и Stoilodon sp. (экз. СГУ № 155/79). Помимо химер, в этом слое (а также из вышележащих отложений зоны nikitini средневолжского подъяруса и fulgens верхневолжского подъяруса) были собраны многочисленные остатки (зубы, шипы) акул, кости, чешуя и многочисленные отолиты костистых рыб, зубы морских рептилий, разнообразные беспозвоночные – двустворчатые, брюхоногие, лопатоногие и головоногие моллюски, брахиоподы и иглокожие (иглы последних составляют основную массу вмещающего “ракушняка”).
Рассматриваемый интервал разреза также описан во временных разрезах, вскрытых при строительных работах близлежащих районов Москвы (Маленкина, Школин, 2009; Школин, Маленкина, 2015) и ряда сохранившихся естественных разрезов Подмосковья (Рогов, 2017).
2. ЛФР 7-2-бис. Заброшенный ныне карьер № 7-2-бис (N 55°23′36′′, E 38°51′02.6′′) Лопатинского месторождения фосфоритового сырья, в Воскресенском р-не Московской области. Волжские отложения (общей мощностью около 3 м) на карьерах месторождения представлены песчано-глинистыми породами небольшой мощностью (1.5–2 м), содержащими фосфориты, средневолжского подъяруса (зоны panderi, virgatus и nikitini), и песчаными (пески, песчаники с фосфоритами) породами верхневолжского подъяруса (Митта, 1993; Герасимов и др., 1995; Рогов, 2017). Подстилаются волжские отложения глинами оксфорда, а перекрываются песчаниками рязанского яруса нижнего мела (Митта, 2005).
Материал по Moskovirhynchus robustus gen. et sp. nov. из местонахождения Кунцево представлен шестью зубными пластинам разной сохранности, в их числе – голотип СГУ № 155/75 и сошниковая пластина (экз. СГУ № 155/74) ювенильной особи; из карьера ЛФР № 7-2-бис – двумя зубными пластинам: сравнительно крупной левой небной пластиной (экз. СГУ № 155/64), а также неполной правой мандибулярной пластиной ювенильной особи (экз. СГУ № 155/73). Из этого местонахождения также определены другие таксоны химер (Попов, Шаповалов, 2007): Brachymylus sp. и “эдафодонтиды” “Ischyodus” cf. townsendi Buckland, 1835 и Elasmodectes sp. [последний таксон был позже переопределен как (?) Stoilodon sp.: см. Попов, Ефимов, 2012)]. Весь материал происходит из отвалов горно-вскрышного комплекса (ex situ) и датируется, наиболее вероятно, средневолжским подъярусом, зоной virgatus, судя по частоте находок аммонитов зонального комплекса в местах сбора пластин химер (К.М. Шаповалов, личн. наблюдение, 1990-е гг.). Сборы К.М. Шаповалова, 1997–1999 гг.
Зона virgatus средневолжского подъяруса сейчас сопоставляется (Rogov, 2010, 2014; Киселев и др., 2018) с нижней частью портландского яруса Англии (зона Glaucolithites glaucolithus полностью и зоны Galbanites okusensis и Progalbanites albani частично) (Cope et al., 1980; Cope, 2013), или нижней частью верхнего титона (нижняя часть зоны Micracanthoceras ponti) и верхами нижнего титона Юго-Западной Германии и Юго-Восточной Франции (Schweigert, 2007; Bulot et al., 2014) (рис. 1, в; см. также схему зональной корреляции волжского яруса в: Arkhangelsky et al., 2019, рис. 2).
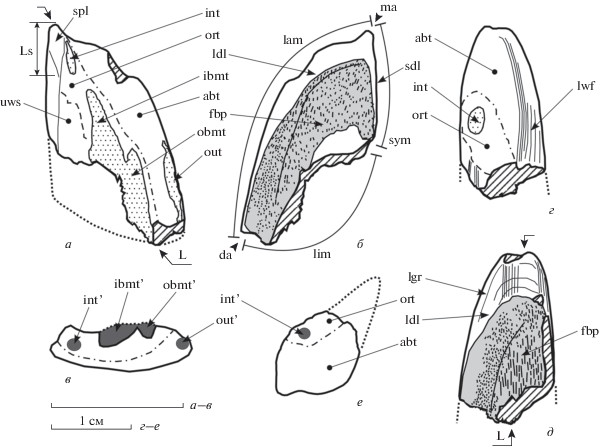
Рис. 2.
Морфологическая интерпретация мандибулярных и небных зубных пластин Moskovirhynchus robustus gen. et sp. nov. и терминология, использованная в тексте; а–в – экз. СГУ, № 155/65, правая мандибулярная (L18, Ls3.2) пластина: а – симфизно-окклюзивно, б – базально, в – лингвальный профиль; Моcква, Кунцево; верхняя юра, средневолжский подъярус, зона virgatus; г–e – экз. СГУ, № 155/66, левая небная (L19.8) пластина: г – окклюзивно, д – базально, е – лингвальный профиль; Московская обл., Лопатинский фосфоритовый рудник (ЛФР) № 7-2-бис; верхняя юра, средневолжский подъярус, зона ?virgatus. Обозначения: косой штриховкой показаны поврежденные поверхности или части пластины; точечный крап – васкулярный плеромин триторов; темно-серая заливка – васкулярный плеромин; пунктир – граница между нестертой частью окклюзивной поверхности и поверхностью прикуса; точечный пунктир – направления отсутствующих частей зубных пластин. Сокращения: (’), int’, out’ и др. – тела плеромина соответствующих триторов в лингвальной профиле; abt – аборальная часть пластины; da – дистальный угол; fbp – поле базальной перфорации (светло-серая заливка); ibmt – внутренняя ветвь срединного тритора; int – внутренний тритор; L – медио-дистальная длина; lam – лабиальный край; ldl – латеральная нисходящая пластинка; lgr – линии роста на поверхности “компактной глянцевой ткани”, lim – лингвальный край; Ls – длина симфизной площадки мандибулярной пластины; lwf – лабиальная фасетка стирания; ma – медиальный угол; obmt – внешняя ветвь срединного тритора; ort – оральная часть пластины; out – наружный тритор; sdl – симфизная нисходящая пластинка; spl – симфизная площадка; sym – симфизный край; uws – нестертая часть окклюзивной поверхности.
ТЕРМИНОЛОГИЯ ЗУБНЫХ ПЛАСТИН
Описательная терминология и измерения были предложены первым автором (Попов, 1999, 2003; Попов, Безносов, 2006; Попов, Шаповалов, 2007; Popov, Machalski, 2014; Popov et al., 2019), ключ к основным описательным терминам и измерениям приведен на рис. 2 и 3.
Рис. 3.
Зубные пластины ювенильных особей позднеюрского Moskovirhynchus robustus gen. et sp. nov.: а, б – экз. СГУ, № 155/73, неполная правая мандибулярная (L8.3, Ls2.3) пластина: а – базально, б – окклюзивно; Московская обл., ЛФР- 7-2-бис; средневолжский подъярус, зона ?virgatus; в–з – экз. СГУ, № 155/74, правая сошниковая (L 6.7) пластина: в – медиально, г – симфизно-окклюзивно, д – базально, е – лингвально, ж – парабазально, з – апикально; Моcква, Кунцево; средневолжский подъярус, зона virgatus. Масштабная линия универсальна для всех фигур. Обозначения: pbs – парабазальная поверхность сошниковой пластины; knb – шишки (без плеромина); остальные см. рис. 2.

При отсутствии на мандибулярных пластинах каллоринхид передне-наружного тритора и отсутствии возможности, таким образом, контролировать положение пластины в онтогенетической серии, как это делается для “эдафодонтид” (Попов, Безносов, 2006; Попов, Шаповалов, 2007; Popov, Machalski, 2014), использовано измерение длины симфизной площадки (Ls) контакта левых и правых мандибулярных пластин (рис. 2, а). В случае небных и сошниковых пластин как относительный показатель использована максимальная медио-дистальная длина (L) образца (рис. 2, д), хотя этот показатель и менее надежен.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Система химеровых рыб (уровень надсемейства и выше) приведена по Дж. Нельсону (Nelson, 2006). Состав семейства Callorhinchidae Garman, 1901 дан по представлениям авторов (Попов, 2003; Popov et al., 2019).
КЛАСС CHONDRICHTHYES
ПОДКЛАСС HOLOCEPHALI
НАДОТРЯД HOLOCEPHALOMORPHA
ОТРЯД CHIMAERIFORMES
ПОДОТРЯД CHIMAEROIDEI
НАДСЕМЕЙСТВО CALLORHINCHOIDEA GARMAN, 1901
СЕМЕЙСТВО CALLORHINCHIDAE GARMAN, 1901
Род Moskovirhynchus Popov et Shapovalov, gen. nov.
Название рода – по р. Москве, на берегах которой был найден материал, и от rhynchus греч. – клюв; м.р.
Типовой вид – Moskovirhynchus robustus gen. et sp. nov.
Диагноз (по зубной системе; знаком (*) обозначены апоморфные признаки). Каллоринхид, известный по некрупным мандибулярным, небным и сошниковой зубным пластинам. Поле базальной перфорации хорошо развито на всех пластинах. Мандибулярные пластины имеют васкулярные триторы: срединный тритор широкий, раздвоенный, его внутренняя ветвь хорошо развита (*). Внутренний тритор развит умеренно. Наружный тритор узкий. Небные пластины робустные и высокие, с сильно развитой аборальной частью, в полтора–два раза превышающей высоту оральной части пластины; единственный округлый внутренний тритор из васкулярного плеромина смещен к симфизному краю (*). Сошниковые пластины субтреугольные, латерально сжатые, с развитым симфизом, развернутым окклюзивно и формирующим симфизный гребень в окклюзии; триторы отсутствуют (*).
Видовой состав. Типовой вид из Центральной России, зона virgatus, средневолжский подъярус верхней юры.
Сравнение. От наиболее близкого современного рода Callorhinchus Lacepède, 1798, известного также в ископаемом состоянии (Stahl, 1999; Otero et al., 2013; Cicimurri, Ebersole, 2015), новый род отличается выраженной бифуркацией срединного тритора с более развитой внутренней ветвью мандибулярной зубной пластины, большей массивностью небной пластины со слабо развитым единственным мелким тритором, более высокой нисходящей пластинкой и, в целом, более развитой аборальной частью небной пластины; более латерально сжатой сошниковой пластиной с развернутой окклюзивной симфизной поверхностью и полностью лишенной триторов (у каллоринхов обычно присутствует один тритор: Аверьянов, 1997; Herman et al., 2001).
От среднеюрского Pachymylus Woodward, 1892 московиринх отличается наличием внутреннего тритора и бифуркации срединного тритора мандибулярной пластины; более мелким внутренним тритором небной пластины, большим удалением этого тритора от симфизного края и отсутствием симфизных отростков плеромина. Сошниковые зубные пластины для Pachymylus не известны.
От среднеюрского Duffinodus Popov, 2003 новый род отличается меньшей высотой симфиза и меньшей робустностью мандибулярной пластины, более выраженной на ней внутренней ветвью срединного тритора (у даффинодуса наиболее выражена наружная ветвь, а внутренняя более короткая и медиально сегментированная), медиальным положением внутреннего тритора; более латерально сжатой сошниковой пластиной с развернутым окклюзивно симфизом, формирующим симфизный гребень, и без окклюзивных триторов. Небные пластины для даффинодуса не известны.
От среднеюрского Brachymylus Woodward, 1892 московиринх отличается отсутствием симфизного тритора на мандибулярной пластине, бифуркацией на ней срединного тритора и лучшим развитием симфизной половины латеральной нисходящей пластинки; наличием только одного тритора небной пластины (у брахимилуса имеются два развитых васкулярных тритора). Сошниковые зубные пластины для Brachymylus не известны.
От раннеюрского “Brachymylus” latus Duffin, 1996, известного только по мандибулярной пластине, новый род отличается меньшей степенью редукции окклюзивных триторов, присутствием наружного тритора и иной структурой бифуркации срединного тритора, с более развитой и глубокой внутренней ветвью, меньшей робустностью мандибулярной пластины, менее редуцированной по высоте симфизной частью латеральной нисходящей пластинки.
От раннеюрских родов Bathytheristes Duffin, 1995 и Eomanodon Ward et Duffin, 1989, известных только по небным зубным пластинам, Moskovirhynchus gen. nov. отличается более робустной структурой небной пластины, отсутствием в структуре тритора пластинчатого плеромина (батитеристес имеет один, а эоманодон – два более крупных пластинчатых тритора), лучше развитой аборальной частью пластины.
Замечания. Мандибулярные пластины московиринха очень близки к таковым каллоринха, отличаясь в основном структурой бифуркации срединного тритора, которая у современных и ископаемых видов последнего отсутствует даже как морфологическая аберрация (Stahl, 1999; Kriwet, Gazdzicki, 2003; Otero et al., 2013). Учитывая, что мандибулярные пластины химер являются наиболее диагностичными структурами озубления у химеровых рыб (Stahl, 1999; Попов, 2003), московиринха и каллоринха можно считать наиболее близкими среди всех родов каллоринхид в принимаемом здесь объеме (Попов, 2003; Popov et al., 2019). Состав семейства Callorhinchidae требует дальнейшей ревизии, которая не входит в задачу настоящей статьи, но указанную пару родов (Callorhinchus и Moscovirhinchus) можно считать таксонами Callorhinchidae sensu stricto. Московиринх, таким образом, является наиболее древним представителем современных слоновых химер, известных по зубным пластинам. Следующим за ним по времени существования является Callorhinchus borealis Nessov et Averianov, 1996 из позднего альба (ранний мел) Центральной России (Несов, Аверьянов, 1996) – древнейший из известных видов слоновых химер рода Callorhinchus.
Moskovirhynchus robustus Popov et Shapovalov, sp. nov.
Табл. IX , фиг. 1–6 (см. вклейку)
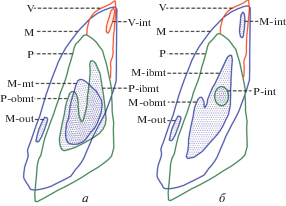
Рис. 4.
Схематическая реконструкция левостороннего перекрытия (прикуса) зубных пластин для двух видов каллоринхид, без масштаба: a – Callorhinchus callorhynchus (L., 1758), экз. СГУ, № 155/103; восточная часть Тихого океана, к западу от Сан-Антонио, Чили; современный; б – Moskovirhynchus robustus gen. et sp. nov.; поздняя юра Московской обл. Обозначения: контуры зубных пластин и префиксы в обозначении триторов: M (M-) – мандибулярная зубная пластина (синий цвет); P (P-) – небная зубная пластина (зеленый цвет); V (V-) – сошниковая зубная пластина (красный цвет); контуры триторов: ibmt – внутренняя ветвь срединного тритора (с бифуркацией); int – внутренний тритор; mt – срединный тритор; obmt – внешняя ветвь срединного тритора (с бифуркацией); out – наружный тритор. Точечным крапом показан васкулярный плеромин триторов.
Callorhinchus sp.: Попов, Шаповалов, 2007, рис. 3, фиг. а–е, табл. 1 , фиг. 1–3; Popov et al., 2019, рис. 4C.
Название вида robustus лат. – крепкий, робустный; характеристика небной пластины в сравнении с таковыми ископаемых и современных слоновых химер рода Callorhinchus.
Голотип – СГУ, № 155/75, левая мандибулярная (L 19.4) пластина; г. Москва, местонахождение Кунцево; верхняя юра, средневолжский подъярус, зона virgatus.
Описание (рис. 2, 3). Для вида известны мандибулярные, небные и сошниковая зубные пластины. Мандибулярные пластины взрослых особей (рис. 2, а–в; табл. IX , фиг. 1–4) характеризуются выпуклой симфизной поверхностью, с нечеткими краями без развитых гребней; поверхность расширяется медиально за счет симфизной нисходящей пластинки, в медиальной же части несет симфизную площадку, которая развернута окклюзивно. Базальная поверхность поперечно выпуклая, ее большую часть занимают нисходящие пластинки с сопряженным полем базальной перфорации. Нисходящие пластинки невысокие, длинные, смыкаются около медиального угла пластины. Латеральная нисходящая пластинка утолщается дистально, формируя неглубокую дисто-базальную борозду. Поле базальной перфорации хорошо развито и занимает почти всю базальную поверхность, исключая небольшой лингвальный сектор, параллельный латеральной пластинке. Лабиальная поверхность узкой полосой окаймляет латеральную пластинку и несет слабо выраженные линии роста по поверхности “компактной глянцевой ткани”. Лабиальный край слабовыемчатый и формирует один (реже – два) асимметричный лингвальный вырез. На окклюзивной поверхности развиты васкулярные триторы: внутренний, наружный и составной срединный. Внутренний тритор узкий, умеренно развитый, смещен к самому медиальному углу пластины; наружный тритор узкий, длинный и сопоставим по ширине с внутренним, он протягивается вдоль лабиального края и заметно отставлен от срединного тритора. Срединный тритор раздвоенный: внутренняя ветвь широкая (в два–три раза шире наружной), длинная и протянутая медиально; наружная ветвь узкая, короткая, не выступает в плане за медиальное окончание наружного тритора. В лингвальном профиле ветви хорошо обособлены, ось внутренней ветви смещена в сторону симфизного края. Граница оральной и аборальной частей пластины проходит вдоль срединного тритора; наружный тритор размещен на аборальной части, которая окклюзивно имеет вид широкой полосы. Между внутренней ветвью срединного тритора и симфизным краем сохранился нестертый участок с глянцевой покровной тканью, линии роста заметны очень слабо.
Мандибулярная пластина ювенильной особи (экз. СГУ, № 155/73; рис. 3, а–в) представляет собой сохранившуюся центральную часть пластины с внутренним и срединным тритором. Срединный тритор приострен медиально, бифуркация почти не выражена, но разрушенный васкулярный плеромин показывает, что тритор является составным – с более глубокой и медиально протянутой внутренней ветвью и более мелкой и немного более широкой наружной ветвью. Срединный тритор медиально доходит до внутреннего тритора; последний имеет субтреугольную форму и показывает неясную структуру в виде сохранившихся трех косых и медиально направленных перегородок в пластинчатой структуре, при отсутствии заполнения плеромином. Дистальная часть пластины и наружный тритор не сохранились. На лабиальной поверхности сохранился медиальный фрагмент латеральной нисходящей пластинки. Базальная поверхность в сохранившейся части полностью покрыта полем базальной перфорации.
Небные пластины взрослых особей (рис. 2, г–е; табл. IX , фиг. 5 ) массивные и высокие, с сильно развитой аборальной частью, охватывающей в лингвальном профиле в 2–2.5 раза более низкую оральную часть. Симфизная поверхность высокая, с невыраженным симфизно-окклюзивным краем; она скошена окклюзивно и постепенно переходит в нестертую часть окклюзивной поверхности. Около внутреннего тритора симфизная поверхность неровная, осложнена парой слабо выраженных борозд и продольным триторным валиком. В базальной части симфизной поверхности заметны прямые и скошенные медиально линии роста. В базальном плане пластина рельефная с сильно развитыми и хорошо перфорированными нисходящими пластинками. Латеральная пластинка очень высокая (максимум до 4 мм), смыкается с менее развитой симфизной пластинкой в средней части лабиальной поверхности. От места соединения пластинки расходятся по дуге дистально (латеральная пластинка) и к симфизно-базальному углу (симфизная пластинка); в последнем случае дуга более крутая. Поле базальной перфорации развито хорошо и на всей видимой части базальной поверхности. Лабиальная поверхность несет умеренно развитую борозду и осложнена дугообразными линиями роста пластины. Окклюзивная поверхность прикуса протягивается медио-дистально, равномерно и резко скошена медиально; ее лабиальный край занимает хорошо развитая лабиальная фасетка стирания, расширяющаяся дистально. В средней части пластины, около симфизно-окклюзивного края расположен небольшой, короткий и овальный (круглый в профиле) васкулярный внутренний тритор. Граница оральной и аборальной частей пластины выражена на окклюзивной поверхности швом, медиально огибающим внутренний тритор и выходящим к симфизной поверхности.
Единственная сошниковая пластина (экз. СГУ, № 155/74; рис. 3, в–з) принадлежит, судя по размеру, ювенильной особи. Пластина субтреугольная при виде на симфизно-окклюзивную поверхность. Лабиальная поверхность выпуклая в дистальной половине и слабо S-образная в профиле. Латеральная нисходящая пластинка хорошо выражена, сплошная, формирует большую часть лабиальной поверхности. В медиальной трети она прижата к базальной поверхности и увеличивает высоту дистально. Поле базальной перфорации хорошо выражено и полностью покрывает всю сохранившуюся часть базальной поверхности. Симфизная поверхность лежит в плоскости окклюзивной и сходится с лабиальной поверхностью под углом около 15°; их контакт формирует выраженный апикальный гребень. Вдоль симфизно-окклюзивного края располагаются две шишки без плеромина, дистальная чуть более крупная; обе не выходят за пределы нестертой части пластины. Парабазальная площадка высокая. Окклюзивные триторы отсутствуют окклюзивно; следов тел плеромина на лингвальной поверхности также не наблюдается.
| Экз. № | L | Ls | Hpb |
|---|---|---|---|
| СГУ 155/64 [P] | 27.5 | – | – |
| СГУ 155/65 (паратип) [M] | 18 | 3.2 | – |
| СГУ 155/66 (паратип) [P] | 19.8 | – | – |
| СГУ 155/73 [M] | 8.3 | 2.3 | – |
| СГУ 155/74 [V] | 6.7 | – | 2.7 |
| СГУ 155/75 (голотип) [M] | 19.4 | 4.5 | – |
| СГУ 155/76 [M] | 13.9 | – | – |
| СГУ 155/77 [M] | 10.2 | 7.2 | – |
Возрастные изменения. Крайние по размерам мандибулярные пластины (экз. СГУ, №№ 155/73 и 77) различаются по промеру Ls в три раза. Самая маленькая пластина (экз. СГУ, № 155/73) показывает при этом хорошо развитый срединный тритор с глубокой внутренней ветвью. Ветвь также хорошо протянута медиально, доходя до дистального окончания внутреннего тритора, в отличие от более крупных пластин, где это не наблюдается. Вместе с тем, бифуркация срединного тритора слабо выражена окклюзивно, а внутренний тритор по структуре более близок к пластинчатому плеромину, чем к васкулярному. Последнее, видимо, связано с очень мелкими размерами зубной пластины, которую можно считать ювенильной, а структуру внутреннего тритора – недоразвитой васкулярной. Более крупные мандибулярные пластины (экз. СГУ, №№ 155/65, 75–77), несмотря на небольшую размерную дифференциацию, выраженных возрастных изменений в структуре не показывают.
Небные пластины (экз. СГУ, №№ 155/64 и 66), несмотря на полуторакратную разницу в размерах, также не показывают серьезных морфологических различий, включая сходную степень развития лабиальной фасетки стирания. Можно предположить, что для этого вида характерна стабильная морфология зубной системы в онтогенезе и, возможно, сходный тип обработки пищи и единый спектр питания для установленных по размеру зубных пластин онтогенетических стадий рыб.
Замечания. Пластины отнесены к озублению одного вида на основании корреляции морфологических признаков (Попов, 2003), размерности пластин, а также сходного развития поля базальной перфорации на всех пластинах, которое полностью отсутствует у другого рода каллоринхид (Brachymylus), известного в средневолжском комплексе химеровых рыб Подмосковья (Попов, Шаповалов, 2007, табл. I , фиг. 5, 6 ).
Распространение. Верхняя юра, волжский ярус, средневолжский подъярус (зона virgatus) Москвы и Московской обл., Центральная Россия.
Материал: пять мандибулярных, две небных и одна сошниковая пластины разной сохранности из двух разрезов: Кунцево (экз. СГУ, № 155/64-65, 74-77), и ЛФР 7-2-бис (экз. СГУ, № 155/66 и 73). Сб. К.М. Шаповалова, 1997–1999 гг.
ОБСУЖДЕНИЕ
Степень развития аборальной части небной пластины московиринха (в полтора–два раза больше оральной части пластины) аналогична таковой для мандибулярной пластины среднеюрского рода Ottangodus из байоса Франции (Popov et al., 2019) и больше ни у каких каллоринхид не встречается. Предполагается, что аборальная часть пластин (несущая у мандибулярных пластин каллоринхид также наружный тритор) является примитивной структурой для химероидей и может представлять рудимент альтернативного зуба (Didier et al., 1994). В связи с этим, можно считать, что по этому признаку небные пластины московиринха устроены более примитивно, чем у Callorhinchus и других родов химеровых рыб. С другой стороны, для небных пластин каллоринха типичен крупный составной срединный тритор с двумя медиальными ветвями разной степени выраженности, который присутствует у всех современных и ископаемых видов рода (Stahl, 1999; Kriwet, Gazdzicki, 2003; Otero et al., 2013). Локализация на небной пластине Moskovirhynchus gen. nov. только одного внутреннего тритора (возможно, аналога внутренней ветви срединного тритора каллоринха), но более мелкого и смещенного симфизно, можно также считать продвинутым состоянием (эволюционный тренд на полную редукцию окклюзивных триторов в зубной системе наиболее выражен у современных химер рода Rhinochimaera), равно как и полное отсутствие тритора на сошниковой пластине, что для каллоринхид не характерно (Stahl, 1999; Herman et al., 2001). Это показывает общую редукцию гиперминерализованой ткани (плеромина) в верхнечелюстной части зубной системы Moskovirhynchus и усиление дробящей функции (увеличение площади плеромина за счет бифуркации срединного тритора) в нижнечелюстной части озубления.
Реконструкция функционального перекрытия (окклюзии) зубных пластин для Moskovirhynchus и современного Callorhinchus callorhynchus показывает (рис. 4), что озубление первого характеризуется меньшим перекрытием окклюзивных срединных триторов. При этом у московиринха развитые ветви срединного тритора мандибулярной пластины контактировали с более развитой и более плотной по структуре аборальной частью небной пластины (что формировало у последней характерно выраженную лабиальную фасетку стирания и медиально сильно скошенную окклюзивную поверхность; см. фиг. 2, г; табл. IX , фиг. 5 а, 5б, 6а, 6в). Такая структура окклюзии у Moskovirhynchus могла компенсировать недостаток гиперминерализованной ткани (триторы) и/или являться адаптацией к использованию менее прочных кормов (тонкораковинные беспозвоночные и т.п.).
Аналогичное современному каллоринху функционирование медиальной части зубной системы [захват (?) пищевых объектов] у Moskovirhynchus обеспечивал смещенный на клюв мандибулярной пластины внутренний тритор. У современного каллоринха аналогичный эффект достигается сходным по форме внутренним тритором сошниковой пластины (у ископаемых Callorhinchus функция (?) захвата еще более развита при наличии внутренних триторов и на мандибулярной, и на сошниковой пластинах, чередующихся при окклюзии; Е.В. Попов, личн. набл.). Общая функциональность зубной системы (при отсутствии режущих или чередующихся триторов, иных типов плеромина и т.п.) у двух рассматриваемых родов, видимо, в целом была сходной.
Химерообразные рыбы (Chimaeriformes) конца юрского периода известны в мире в составе всего нескольких ассоциаций (Stahl, 1999). Самая известная из них происходит из литографских известняков Южной Германии (верхний кимеридж Нусплингена, земля Баден-Вюртемберг; верхний кимеридж и нижний титон “архипелага Золенгофен” в Баварии). Эта ассоциация происходит из “консерват-лагерштетта” (Arratia et al., 2015) и известна по многочисленным скелетным остаткам исключительной сохранности химероидов Ischyodus quenstedti Wagner, 1857 и Elasmodectes avitus (von Meyer, 1862) (см. Popov et al., 2013; Kriwet, Klug, 2015). В комплексе также присутствуют единичные находки каллоринхид – Callorhinchidae gen. et sp. indet. (Duffin, 2018), ринохимерид (неописанная мелкая Harriotta sp. в частной коллекции в Германии; Е.В. Попов, личн. наблюдение, 2013; этот же образец изображен как “ювенильный экземпляр химеры Ischyodus sp.”: Mäuser, 2015, с. 525, рис. 1013 ) и химеропсидная рыба Chimaeropsis paradoxa Zittel, 1887 – реликт раннемезозойской группы мириакантовидных цельноголовых (Riess, 1887; Woodward, 1891; Lauer et al., 2018). В Англии из отложений портланда (Portland Beds), а также, в переотложенном состоянии, в базальном горизонте меловой системы (Neocomian Bone-Beds) известны разрозненные зубные пластины лишь одного вида химер – Ischyodus townsendi Buckland, 1835 (Woodward, 1891), отличающиеся, впрочем, очень крупными размерами зубных пластин (Newton, 1878, табл. XI , фиг. 1–7 ; Woodward, 1891, с. 64–65). В целом, к титону разнообразие химерообразных рыб в Западной Европе сокращается до пяти родов (четыре рода химероидей и один род химеропсид), уменьшаясь с известных максимально для юры девяти родов в келловее (комплекс “оксфордской глины”; Popov et al., 2012) и позже – шести родов в кимеридже (Popov et al., 2009).
Бореальные отложения волжского яруса Русской плиты содержат более редкий и разрозненный материал по химерам, но он позволяет провести сравнение западно- и восточноевропейских ассоциаций. Бореальный волжский ярус территории европейской части России показывает сходное разнообразие (пять родов), но иное распределение таксонов – отсутствие мириакантовидных цельноголовых и носатых химероидей, демонстрируя при этом большее разнообразие “эдафодонтид” (три рода) и каллоринхид (два рода). Это указывает на возможное проявление палеобиогеографической дифференциации в ассоциациях химерообразных рыб в конце юрского периода.
Список литературы
Аверьянов А.О. Новые юрские химеры России // Палеонтол. журн. 1992. № 3. С. 57–62.
Аверьянов А.О. Редкая находка сошниковой пластинки слоновой химеры (Holocephali, Callorhinchidae) в позднем мелу России // Палеонтол. журн. 1997. № 1. С. 78–80.
Апродов В.А., Апродова А.А. Движения земной коры и геологическое прошлое Подмосковья (Учебные геологические экскурсии в окрестностях Москвы). М.: Изд‑во Моск. ун-та, 1963. 268 с.
Боголюбов Н.Н. Следы химер в московском портланде // Ежегодн. по геол. и минералогии России. 1912. Т. 14. № 2. С. 25–28.
Герасимов П.А. Юрская система // Геология СССР. Т. IV. Центр европейской части СССР. Геологическое описание. М.: Недра, 1971. С. 373–416.
Герасимов П.А., Митта В.В., Кочанова М.Д. Ископаемые волжского яруса Центральной России. М.: ВНИГНИ-МосГорСЮН, 1995. 116 с.
Киселев Д.Н., Рогов М.А., Захаров В.А. Зона Volgidiscus singularis терминальной части волжского яруса европейской части России и ее значение для межрегиональной корреляции и палеогеографии // Стратигр. Геол. корреляция. 2018. Т. 26. № 2. С. 87–114.
Маленкина С.Ю., Школин А.А. Новые данные о келловейских и оксфордских отложениях г. Москвы // Юрская система России: проблемы стратиграфии и палеогеографии: Третье Всеросс. совещ.: научные материалы / Ред. Захаров В.А. Саратов: Наука, 2009. С. 133–136.
Митта В.В. Аммониты и зональная стратиграфия средневолжских отложений Центральной России. Киев: Геопрогноз, 1993. 132 с.
Митта В.В. Новые данные о возрасте подошвы рязанского яруса // Стратигр. Геол. корреляция. 2005. Т. 13. № 5. С. 51–59.
Несов Л.А., Аверьянов А.О. Древние химерообразные рыбы России, Украины, Казахстана и Средней Азии. I. Некоторые экологические особенности химер и обзор местонахождений // Вестн. СПбГУ. Сер. 7. 1996. Вып. 1. № 21. С. 11–19.
Никитин С.Н. Заметка по вопросу о последовательности волжского яруса московской юры. СПб., 1883. 8 с. Отд. оттиск.
Павлов А.П. Геологический очерк окрестностей Москвы. Пособие для экскурсий и для краеведов. 5-е изд. М.: МОИП, 1946. 84 с.
Попов Е.В. Новые данные по морфологии зубных пластин химеровых рыб рода Ischyodus из мела и палеогена Центральной России и Поволжья // Матер. по истории фаун Евразии / Ред. Даревский И.С., Аверьянов А.О. СПб., 1999. С. 67–82 (Тр. Зоол. ин-та РАН. Т. 277).
Попов Е.В. Новый род слоновых химер (Holocephali: Callorhinchidae) из верхнего келловея Саратовского Поволжья, Россия // Палеонтол. журн. 2003. № 5. С. 59–66.
Попов Е.В. Новые данные по химеровым рыбам (Chondrichthyes, Holocephali) из юры Европейской России // Юрская система России: проблемы стратиграфии и палеогеографии: Тез. докл. Всеросс. совещ. (21–22 ноября 2005 г., Москва). М.: ГИН РАН, 2005. С. 199–200.
Попов Е.В., Безносов П.А. Остатки химер (Holocephali: Chimaeroidei) из верхнеюрских отложений Республики Коми // Современная палеонтология: классические и новейшие методы – 2006: Сборн. науч. статей / Ред. Розанов А.Ю., Лопатин А.В., Пархаев П.Ю. М.: ПИН РАН, 2006. С. 55–64.
Попов Е.В., Ефимов В.М. Новые находки химер рода Stoilodon Nessov et Averianov, 1996 (Holocephali, Chimaeroidei) в поздней юре и раннем мелу европейской части России // Изв. Саратовск. ун-та. Нов. сер. Науки о Земле. 2012. Т. 12. Вып. 1. С. 66–79.
Попов Е.В., Шаповалов К.М. Новые находки химеровых рыб (Holocephali, Chimaeroidei) в юре европейской части России // Современная палеонтология: классические и новейшие методы – 2007: Сборн. науч. статей / Ред. Розанов А.Ю., Лопатин А.В., Пархаев П.Ю. М.: ПИН РАН, 2007. С. 25–47.
Рогов М.А. Аммониты и инфразональная стратиграфия кимериджского и волжского ярусов юга Московской синеклизы // Тр. Геол. ин-та РАН. 2017. Вып. 615. С. 7–160.
Розанов А.Н. О зонах подмосковного портланда и о вероятном происхождении портландских фосфоритных слоев под Москвой // Матер. к познанию геол. строения Росс. Империи. 1912. Вып. 4. С. 17–103.
Рулье К.Ф. О животных Московской губернии. М., 1845. 96 с.
Стародубцева И.А. Эволюция взглядов на стратиграфию юры Центральной России (XIX–XX вв.). М.: Научн. мир, 2006. 212 с.
Тесакова Е.М. Остракоды зоны Virgatites virgatus из разрезов Москвы // Тр. Геол. ин-та РАН. 2017. Вып. 615. С. 301–310.
Траутшольд Г.А. Юго-западная часть Московской губернии с картой. Комментарий на специальную геологическую карту этой части России // Матер. для геол. России. СПб., 1870. С. 211–266.
Школин А.А., Маленкина С.Ю. Сравнение типов разрезов верхней юры (волжский ярус) – нижнего мела юго-востока Московского региона // Юрская система России. Проблемы стратиграфии и палеогеографии. VI Всеросс. совещ.: научн. матер. Махачкала: АЛЕФ, 2015. С. 304–308.
Arkhangelsky M.S., Zverkov N.G., Rogov M.A. et al. Colymbosaurines from the Upper Jurassic of European Russia and their implication for palaeobiogeography of marine reptiles // Palaeobiodiv. and Palaeoenviron. 2019. V. 100. P. 197–218. https://doi.org/10.1007/s12549-019-00397-0
Arratia G., Schultze H.-P., Tischlinger H., Viohl G. (eds.) Solnhofen – Ein Fenster in die Jurazeit. Münich: Verl. Dr. F. Pfeil, 2015. V. 1 + 2.
Bulot L.G., Frau C., Wimbledon W.A.P. New and poorly known Perisphinctoidea (Ammonitina) from the Upper Tithonian of Le Chouet (Drôme, SE France) // Volumina Jurassica. 2014. V. 12. № 1. P. 113–128.
Cicimurri D.J., Ebersole J.A. Paleocene chimaeroid fishes (Chondrichthyes: Holocephali) from the eastern United States, including two new species of Callorhinchus // PaleoBios. 2015. V. 32. № 1. P. 1–29
Cope J.C.W. The Bolonian Stage: an old answer to an old problem // Newsletters on Stratigr. 1993. V. 28. № 2/3. P. 151–156.
Cope J.C.W. Stage nomenclature in the uppermost Jurassic rocks of Britain // Geoscience in South-West England. 2013. V. 13. № 2. P. 216–221.
Cope J.C.W., Duff K.L., Parsons C.F. et al. A correlation of Jurassic rocks in the British Isles. Pt 2: Middle and Upper Jurassic // Geol. Soc. London Spec. Rep. 1980. № 15. P. 1–109.
Didier D.A., Stahl B.J., Zangerl R. Development and growth of compound tooth plates in Callorhinchus milii (Chondrichthyes, Holocephali) // J. Morphol. 1994. V. 333. P. 73–89.
Duffin C.J. A callorhynchid chimaeroid (Pisces, Holocephali) from the Nusplingen Plattenkalk (Late Jurassic, SW Germany) // N. Jb. Geol. Paläontol. Abhandl. 2018. V. 289. № 2. P. 161–175.
Gerasimov P.A. Geological excursion to the Moscow suburbs // Upper Jurassic Stratigraphy. Intern. Symp. in the USSR (June 6–18, 1967). Progr. and guidebooks. Moscow, 1967. 51 p.
Herman J., Hovestadt-Euler M., Hovestadt D.C., Stehmann M. Contributions to the study of the comparative morphology of teeth and other relevant ichthyodorulites in living supraspecific taxa of chondrichthyan fishes. Pt C: Holocephali 1: Order Chimaeriformes – Suborder Chimaeroidei // Bull. Inst. Roy. Sci. Natur. Belg., Biol. 2001. V. 71. P. 5–35.
Kriwet J., Gazdzicki A. New Eocene Antarctic chimeroid fish (Holocephali, Chimaeriformes) // Pol. Polar Res. 2003. V. 24. № 1. P. 29–51.
Kriwet J., Klug S. Knorpelfische (Chondrichthyes) // Solnhofen – Ein Fenster in die Jurazeit / Eds. Arratia G., Schultze H.-P., Tischlinger H., Viohl G. Münich: Verl. Dr. F. Pfeil, 2015. S. 334–359.
Lauer B.H., Popov E.V., Duffin C. et al. A new holomorphic specimen of the rare holocephalian fish, Chimaeropsis paradoxa, from the Late Jurassic Plattenkalk of Germany // SVP 2018 Annual Meeting, October 17–20, 2018. Albuquerque, NM, 2018. P. 165.
Mäuser M. Oberjurassische Fossil-Lagerstätten außefhalb der Südlichen Frankenalb. Die laminierten Plattenkalker von Wattendorf in Oberfranken // Solnhofen – Ein Fenster in die Jurazeit / Eds. Arratia G., Schultze H.-P., Tischlinger H., Viohl G. Münich: Verl. Dr. F. Pfeil, 2015. S. 515–535.
Nelson J.S. Fishes of the World. 4th edn. Hoboken: Wiley, 2006. 601 p.
Newton E.T. The chimaeroid fishes of the British Cretaceous rocks // Mem. Geol. Surv. U. K. 1878. Monogr. 4. 62 p.
Otero R.A., Rubilar-Rogers D., Yury-Yanez R.E. et al. A new species of chimaeriform (Chondrichthyes, Holocephali) from the uppermost Cretaceous of the López de Bertodano Formation, Isla Marambio (Seymour Island), Antarctica // Antarctic Sci. 2013. V. 25. № 1. P. 99–106.
Popov E.V., Delsate D., Felten R. A new callorhinchid genus (Holocephali, Chimaeroidei) from the early Bajocian of Ottange-Rumelange, on the Luxembourg-French border // Paleontol. Res. 2019. V. 23. № 3. P. 220–230. https://doi.org/10.2517/2018PR021
Popov E.V., Duffin C.J., Tischlinger H., Atuchin A. Reconstructions of the German Plattenkalk (Late Jurassic) chimaeroid fishes (Holocephali, Chimaeroidei) // 6th Intern. Meeting on Mesozoic Fishes: Diversification and Diversity Patterns. Abstracts / Eds. Schwarz C., Kriwet J. Münich: Verl. Dr. F. Pfeil, 2013. P. 56.
Popov E.V., Machalski M. Late Albian chimaeroid fishes (Holocephali, Chimaeroidei) from Annopol, Poland // Cret. Res. 2014. V. 47. P. 1–18.
Popov E., Ward D., Lepage G. The diversity and nomenclatural revision of the Holocephalian fishes (Chimaeriformes) from the Kimmeridgian (Late Jurassic) of Western Europe // J. Vertebr. Paleontol. 2009. V. 29. Suppl. to № 3. P. 166A.
Popov E.V., Ward D.J., Matheau-Raven E. A revision of the chimaeroid fishes (Chimaeroidei) from the Lower Oxford Clay (Middle Jurassic, Callovian) of Cambridgeshire, England // 56th Annual Meeting of the Palaeontol. Assoc. (December 16–18). Progr. and Abstracts. Dublin, 2012. P. 85.
Riess J. Ueber einige fossile Chimaeriden-Reste im Münchener palaeontologischen Museum // Palaeontogr. 1887. Bd 34. S. 1–28.
Rogov M.A. A precise ammonite biostratigraphy through the Kimmeridgian-Volgian boundary beds in the Gorodischi section (middle Volga area, Russia), and the base of the Volgian Stage in its type area // Volumina Jurassica. 2010. V. 8. P. 103–130.
Rogov M. Infrazonal subdivision of the Volgian Stage in its type area using ammonites and correlation of the Volgian and Tithonian stages // STRATI 2013. First Intern. Congr. on Stratigr. At the Cutting Edge of Stratigraphy. Springer Geol., 2014. P. 577–580.
Schweigert G. Ammonite biostratigraphy as a tool for dating Upper Jurassic lithographic limestones from South Germany – First results and open questions // N. Jb. Geol. Paläontol. Abhandl. 2007. V. 245. P. 117–125.
Stahl B.J. Chondrichthyes III – Holocephali // Handbook of Paleoichthyology. V. 4. / Ed. Schultze H.-P. Münich: Verl. F. Pfeil, 1999. 164 p.
Trautschold H. Recherches géologiques aux environs d’Moscou. Couche jurassique de Mnevniki // Bull. Soc. Natur. Moscou. 1861. V. 34. № 1. P. 64–94.
Woodward A.S. Catalogue of the fossil fishes in the British Museum (Natural History). Part 2. L.: Taylor and Francis, 1891. 567 p.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал