

Палеонтологический журнал, 2021, № 4, стр. 66-73
Сребробрюшки (Acanthuriformes, Leiognathidae) из раннего миоцена Карпатского бассейна
А. Н. Ковальчук a, b, *, Э. Швидницка b, К. Стефаняк b
a Национальный научно-природоведческий музей НАН Украины
Киев, Украина
b Вроцлавский университет
Вроцлав, Польша
* E-mail: biologiest@ukr.net
Поступила в редакцию 06.11.2020
После доработки 26.11.2020
Принята к публикации 26.11.2020
Аннотация
Сребробрюшки (Leiognathidae) представлены в палеонтологической летописи Европы, Африки и Азии скелетными остатками и изолированными отолитами. Описаны отпечатки скелетов этих рыб из нижнего миоцена (бурдигала) местонахождения Харта (Польша). Эти экземпляры морфологически идентичны и сходны по меристическим параметрам с Leiognathoides minutus (Daniltshenko), известным из Кавказского бассейна. Изученные остатки обладают рядом признаков, позволяющих отнести этот вид к лиогнатидам. Среди них пять остеологических синапоморфий семейства Leiognathidae, а также признаки, дающие основания поместить L. minutus в состав Acanthuriformes. Мы предполагаем, что Западный Тетис мог быть центром происхождения сребробрюшек, откуда они мигрировали в разные части Паратетиса. Используя морской путь между Тетисом и Индийским океаном, лиогнатиды проникли в Индийский и западную часть Тихого океана и стали там разнообразными, а в Паратетисе вымерли в течение среднего миоцена. Наши данные подтверждают гипотезу о связи между ихтиофаунами Карпатского и Кавказского бассейнов и возможности обмена фаунистическими элементами между различными частями Паратетиса в раннем миоцене.
ВВЕДЕНИЕ
Leiognathidae (сребробрюшки) – немногочисленное семейство рыб, включающее около 48 видов, входящих в состав девяти родов (Ikejima et al., 2004; Chakrabarty, Sparks, 2008; Kimura et al., 2008; Abraham et al., 2011; Sparks, Chakrabarty, 2015; Nelson et al., 2016). Для этих небольших рыб со сжатым с боков телом характерны выдвижной рот, наличие запирающего механизма для шипов спинного и анального плавников, а также циркумэзофагеальный (окружающий пищевод) световой орган с бактериальным свечением (Sparks, Dunlap, 2004; Sparks et al., 2005; Chakrabarty et al., 2011).
Cребробрюшки распространены в тропических и субтропических регионах Индийского и западной части Тихого океана, часто встречаются в морской и солоноватоводной среде, где они, как правило, обитают на мелководье, однако некоторые виды могут заходить в пресные воды. Представители этой группы впервые появляются в палеонтологической летописи предположительно в среднем палеоцене, будучи представлены Leiognathidarum tashlikensis – формой, описанной на основании отолитов из местонахождения Лузановка на Украине (Schwarzhans, Bratishko, 2011). Находки ископаемых остатков эоценового и олигоценового возраста, относящихся к лиогнатидам, включают Eoleiognathus dorsalis (Agassiz, 1838) из ипра Италии (Bannikov, 2014), Leiognathoides altapinna (Weiler in Hess et Weiler, 1955) из рюпеля Швейцарии (Hess, Weiler, 1955), Франции (Pharisat, 1991) и России (Данильченко, 1960, 1980; Банников, 2001, 2010; Prokofiev, 2002), L. cf. altapinna из рюпеля Германии (Micklich et al., 2017), а также таксоны, описанные на основании отолитов из Бельгии (Nolf, 1985), Франции (Steurbaut, 1982) и Венгрии (Nolf, Brzobohatý, 1994).
Палеонтологическая летопись сребробрюшек включает также Gazza hilberi (Weinfurter, 1952) из миоцена Австрии (Weinfurter, 1952) и Leiognathoides minutus (Daniltshenko, 1980) из Азербайджана и России (Банников, 2001, 2010). Несколько экземпляров из миоценовых отложений Чехии (бурдигал) и Турции (серравалий) были идентифицированы как Leiognathoides sp. (Rückert-Ülkümen, 1994; Reichenbacher et al., 2018). Ископаемые остатки лиогнатид за пределами Европы представлены Euleiognathus tottori из миоцена (бурдигала–лангия) Японии (Yabumoto, Uyeno, 1994, 2011) и Secutor africanus из бурдигала–тортона Западной Африки (Schwarzhans, 2013).
Э. Гилл и С. Михальски (Gill, Michalski, 2020) рассмотрели остеологические признаки сребробрюшек и объединили их в три группы. Первая группа представляет собой 10 синапоморфий, поддерживающих монофилию семейства Leiognathidae, вторая – 14 признаков взрослых особей, позволяющих поместить лиогнатид в состав отряда Acanthuriformes, а третья группа включает в себя дополнительные признаки, общие для хирургообразных и представителей других групп, однако полезные для идентификации сребробрюшек (Gill, Michalski, 2020). Не все эти признаки могут быть определены на ископаемом материале из-за его обычной неполноты и плохой сохранности. Согласно этим авторам (Gill, Michalski, 2020), только Euleiognathus обладает остеологическими синапоморфиями, позволяющими поместить его в состав семейства Leiognathidae. Систематическое положение других ископаемых родов (в частности, Leiognathoides) проблематично, поскольку ранее они не поддерживались достоверными синапоморфными признаками.
Целью настоящего исследования является детальный морфологический анализ ископаемых остатков сребробрюшек из раннего миоцена Центральной Европы, выяснение их остеологических синапоморфий и уточнение их систематического положения. Кроме того, мы кратко обсуждаем палеонтологическую летопись этой группы рыб и приводим предварительную схему их прошлого распространения.
МАТЕРИАЛ И МЕТОДЫ
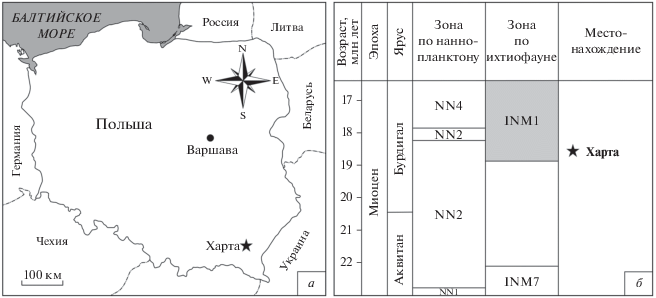
Описанные здесь ископаемые образцы лиогнатид происходят из местонахождения Харта. Оно расположено в центральной части Сколевской толщи, во флишевой зоне Внешних Карпат. Местонахождение было обнаружено в 1976 г. (Rajchel, 1989) в пределах синклинали Харта, заполненной кросненскими слоями (Rajchel, 1989; Jerzmańska et al., 2001). Изучаемые рыбные остатки были собраны на левом берегу руч. Харта (49°51′ с.ш., 22°12′ в.д.), в обнажении, расположенном вдоль дороги, ведущей в дер. Пекло, в административной зоне Подкарпатского воеводства на юго-востоке Польши (рис. 1). Разрез имеет мощность около 15 м и сложен кросненскими сланцами и песчаниками с прослоями менилитовых сланцев и аргиллитов (Jerzmańska et al., 2001). Ископаемые остатки происходят из пепельно-серых и коричневатых сланцев, расположенных на глубине от 5 м ниже до 2 м выше слоя туфа (Добжанский туфовый горизонт по: Kotlarczyk et al., 2006). Этот уровень соответствует пятому туфовому горизонту, установленному В. Сикорой с соавт. (Sikora et al., 1959). Его возраст (ранний миоцен, бурдигал) был установлен Й. ван Куверингом с соавт. (Van Couvering et al., 1981), а позже определен радиометрически Т. Визером (Wieser, 1985) как соответствующий 20.5 ± 0.9 млн лет (Olszewska, 1982; Rajchel, 1989; Garecka, Olszewska, 1998; Jerzmańska et al., 2001; Rajchel et al., 2001). Слои с ископаемыми остатками рыб (в том числе сребробрюшек) отвечают зоне NN2 известкового наннопланктона по Э. Мартини (Martini, 1971) и зоне N5 по фораминиферам (Blow, 1969). Комплекс INM 1 Харты представляет неритическую ихтиофауну, обитавшую на небольшой глубине.
Рис. 1.
Географическое расположение (а) и стратиграфическое положение (б) местонахождения Харта. Ихтиофаунистическое зонирование и его корреляция со стратиграфическими подразделениями приведены по И. Котлярчику и др. (Kotlarczyk et al., 2006).
Описываемый материал представлен сочлененными скелетами, их частями, а также отдельными костями. Всего исследовано 22 экз., хранящихся на каф. палеозоологии Вроцлавского ун-та (Вроцлав, Польша). Число полных, почти полных, частично сохранившихся скелетов и разрозненных костей составляет 2, 5, 12 и 3 соответственно. Серия отпечатков, предварительно идентифицированных как Leiognathus? cf. minutus из Харты, была получена А. Ежманьской, Я. Швидницким и Э. Швидницкой во время полевых исследований в 1982–1984 гг. Их определение проводилось с учетом известных диагностических признаков, детально охарактеризованных в литературе (Данильченко, 1960, 1980; Yabumoto, Uyeno, 1994, 2011; Банников, 2001, 2010; Bannikov, 2014; Sparks, Chakrabarty, 2015; Micklich et al., 2017). Систематика соответствует таковой у Э. Гилла и Дж. Лиса (Gill, Leis, 2019). Фотографии были сделаны с помощью стереомикроскопа Olympus ZSX 12, оснащенного цифровой камерой. Образцы измеряли с помощью штангенциркуля Mitutoyo с точностью до 0.1 мм. Морфометрические признаки приведены по Н. Миклиху и др. (Micklich et al., 2017). Скелетная терминология соответствует таковой для сребробрюшек (James, 1985; Банников, 2001, 2010; Gill, Michalski, 2020).
Морфометрические сокращения: HL – длина головы; lAI–III – длина колючих лучей анального плавника; lDI–III – длина колючих лучей спинного плавника; lVI – длина шипа брюшного плавника; pu4 – четвертый преуральный позвонок; SL – стандартная длина.
Институциональные сокращения: ПИН – Палеонтологический ин-т им. А.А. Борисяка РАН (Москва, Россия); ZPALWr. – каф. палеозоологии Вроцлавского ун-та (Вроцлав, Польша).
Работа выполнена при грантовой поддержке проекта 0121U110402 НАН Украины для исследовательских групп.
Авторы благодарны П. Сохе (Вроцлавский ун-т, Польша) за помощь с подготовкой фотографий исследуемых образцов. Мы искренне признательны А.Ф. Банникову (ПИН РАН) и Н. Миклиху (Hessisches Landesmuseum Darmstadt, Германия) за обсуждение отдельных аспектов темы исследования, а также М.В. Назаркину (Зоологический ин-т РАН) и Дж. Карневале (Ун-т Турина, Италия) за важные замечания, качественно улучшившие рукопись настоящей работы.
ОТРЯД ACANTHURIFORMES
СЕМЕЙСТВО LEIOGNATHIDAE GILL, 1893
Род Leiognathoides Bannikov, 2001
Leiognathoides minutus (Daniltshenko, 1980)
Leiognathus minutus: Данильченко, 1980, с. 133, табл. IV , фиг. 2.
Рис. 2.
Leiognathoides minutus (Daniltshenko, 1980): а – экз. ZPALWr. H/104; б – экз. ZPALWr. H/105; в – экз. ZPALWr. H/111; г – экз. ZPALWr. H/113; д – экз. ZPALWr. H/114; Ю.-В. Польша, местонахождение Харта; нижний миоцен, бурдигал.

Leiognathus? cf. minutus Daniltshenko: Jerzmańska et al., 2001, рис. 5B .
Leiognathoides minutus (Daniltshenko, 1980): Prokofiev, 2002, с. 214, рис. 7 ; Банников, 2010, с. 66, табл. Х, фиг. 5 ; Micklich et al., 2017, рис. 9b .
Голотип – ПИН, № 2180/3; Азербайджан, Апшеронский п-ов, Перекишкюль; нижний миоцен, верхний майкоп.
Описание (рис. 2). Исследованные экземпляры характеризуются различной степенью сохранности. Стандартная длина тела этих рыбок колеблется в пределах 14.6–47.9 мм. Тело высокое (45.6–58.3% SL), его дорсальный край плавно закругленный, каудальная часть постепенно сужается. Голова довольно крупная (43.0–45.0% SL), мозговая коробка составляет 74.8–86.9% длины черепа. Затылочный гребень высокий, заостренный. Орбита большая, ее горизонтальный диаметр (2.1–6.6 мм) составляет 31.6–39.4% HL. Нижняя челюсть соединяется с черепом под передней третью орбиты и несет мелкие конические зубы одинаковой величины, расположенные в один ряд. По крайней мере, у одного образца есть клыкообразные зубы, что мы считаем доказательством постеролатеральной замены зубов (Gill, Michalski, 2020). Supramaxillare отсутствует. Praemaxillare имеет расширенную переднюю часть и характеризуется наличием длинного восходящего отростка, ориентированного вверх, и выступающего постмаксиллярного отростка. Maxillare S‑образное, со сложной головкой. Вентральный край praeoperculum лишен шипов. Operculum умеренно удлиненное, отношение его длины к ширине составляет 1.7–2.1 (в среднем 1.9). Имеется пять лучей жаберной перепонки. Позвоночный столб состоит из 24 позвонков, в том числе 10 туловищных и 14 хвостовых. Три supraneuralia расположены в первом межневральном промежутке (между невральными отростками первого и второго позвонков). В грудных плавниках имеется по 16 лучей. Брюшной плавник состоит из одного внешнего шипа и пяти мягких лучей. В протяженном неразделенном спинном плавнике имеется девять шипов (один из которых сверхштатный), поддерживаемых восемью сильными птеригиофорами, и 14 мягких лучей. Первый дорсальный птеригиофор располагается в первом межневральном промежутке; он имеет очень сильное шиповидное расширение, ориентированное антеродорсально и нависающее над вторым и третьим supraneuralia. Первый шип спинного плавника самый короткий, второй – самый длинный (в 3.3 раза длиннее первого и в 1.4 раза длиннее третьего). Пятый межневральный промежуток вакантный. Анальный плавник расположен напротив мягкой части спинного плавника и содержит три сильных шипа, из которых второй самый длинный (в 2.2 раза длиннее первого шипа анального плавника и лишь в 1.2 раза короче второго шипа спинного плавника), и 14 мягких лучей. Вдоль переднего края шипов спинного и анального плавников есть зазубрины, свидетельствующие о возможном наличии замыкающего механизма (Siegel, 1982). Хвостовой плавник усеченный, содержит 15–17 главных лучей. Хвостовой скелет состоит из одной пары uroneuralia, трех epuralia, а также слитых в две пластинки hypuralia 1–2 и 3–4. Вершины неврального и гемального отростков pu4 лопатовидные. Основания внешних главных лучей хвостового плавника расширены. Тело покрыто циклоидной аксиллярной чешуей.
Размеры. Промеры образцов представлены в табл. 1.
Таблица 1.
Меристические параметры Leiognathoides minutus из Харты (ZPALWr. H/104-H/125) и типовой серии
| Параметр | H/104–H/125 | ПИН (колл. 2180*) |
||
|---|---|---|---|---|
| диапазон | среднее | |||
| Стандартная длина, мм | 14.6–47.9 | 16.2 | До 40 | |
| Максимальная высота тела (BD), мм | 6.4–27.9 | 7.4 | – | |
| Максимальная высота тела (% SL) | 45.6–58.3 | 47.8 | 45–52 | |
| Минимальная BD, мм | 1.9–5.8 | 2.1 | – | |
| Минимальная BD (% SL) | 12.1–13.8 | 13.3 | – | |
| Длина головы (HL), мм | 6.4–20.9 | 7.3 | – | |
| Длина головы (% SL) | 43.0–45.0 | 44.1 | 38–43 | |
| Длина мозговой капсулы, мм | 4.8–16.2 | 5.7 | – | |
| Длина мозговой капсулы (% HL) | 74.8–86.9 | 77.2 | – | |
| Диаметр орбиты, мм | 2.1–6.6 | 2.7 | – | |
| Диаметр орбиты (% HL) | 31.6–39.4 | 37.2 | 35–40 | |
| Operculum | Длина (L), мм | 4.4–4.7 | 4.6 | – |
| Ширина (W), мм | 2.1–2.8 | 2.4 | – | |
| L/W | 1.7–2.1 | 1.9 | – | |
| Шипы спинного плавника | lDI, мм | 0.8–2.4 | 1.1 | – |
| lDII, мм | 3.3–8.3 | 3.6 | – | |
| lDIII, мм | 2.4–6.7 | 2.6 | – | |
| Шип брюшного плавника (lVI), мм | 4.5–4.7 | 4.6 | – | |
| Шипы анального плавника | lAI, мм | 0.8–1.8 | 1.3 | – |
| lAII, мм | 2.5–3.2 | 2.9 | – | |
| lAIII, мм | 1.7–2.5 | 2.2 | – | |
Замечания. Исследованные экземпляры морфологически идентичны Leiognathoides minutus из типовой серии (Данильченко, 1980, с. 133–134; Prokofiev, 2002, с. 214–216). Они хорошо вписываются в ряд морфометрических признаков этого вида, за исключением более длинной головы, а также более короткого и широкого operculum.
Распространение. Азербайджан (Апшеронский п-ов, Перекишкюль), Россия (Дагестан, р. Уллучай), Польша (местонахождение Харта); нижний миоцен, бурдигал.
Материал. 22 экз. (ZPALWr. H/104–H/125) удовлетворительной сохранности из местонахождения Харта.
ОБСУЖДЕНИЕ
Систематическое положение Leiognathoides minutus по остеологическим признакам. Образцы из нижнего миоцена Польши, отнесенные к Leiognathoides minutus, обладают рядом признаков, позволяющих поместить этот вид в состав лиогнатид. В частности, на исследуемом материале установлено наличие пяти остеологических синапоморфий семейства Leiognathidae (Gill, Michalski, 2020): S1 – ребра перекрывают переднее расширение первого анального птеригиофора; S2 – замыкающий механизм, о чем свидетельствует наличие небольших зазубрин вдоль переднего края третьего и четвертого шипов спинного плавника и второго шипа анального плавника; S8 и S9 – hypuralia 1–2 и 3–4 в виде двух недифференцированных пластинок; S10 – расширение оснований нескольких главных лучей хвостового плавника. Постеролатеральное замещение зубов, отсутствие supramaxillare, наличие вакантного межневрального промежутка под передней частью спинного плавника, расположение первого птеригиофора спинного плавника в первом межневральном пространстве, наличие пары uroneuralia, пяти лучей жаберной перепонки, а также лопатообразных вершин неврального и гемального шипов pu4, т.е., признаки A1, A5, A11, A12 и A14 по Гиллу и Михальски (Gill, Michalski, 2020), позволяют поместить исследуемые экземпляры в состав отряда Acanthuriformes. Что касается дополнительных признаков (O1–O9 по: Gill, Michalski, 2020), все они распознаются на наших образцах. Некоторые из них (наличие трех supraneuralia и девяти шипов в спинном плавнике) примитивны и сближают Leiognathoides minutus с Eoleiognathus dorsalis (Agassiz, 1838), однако большинство других характерны для Euleiognathus и современных лиогнатид (Gill, Michalski, 2020).
Прошлое распространение и палеобиогеография лиогнатид. Самые древние известные находки представителей семейства Leiognathidae (как скелетные остатки, так и отолиты) приурочены к западной части Тетиса. Шельф этого морского бассейна был центром формирования мелководной фауны (Harzhauser et al., 2007; Попов и др., 2009). Принимая во внимание присутствие Eoleiognathus dorsalis в раннем эоцене Италии, а также родов олигоценовых сребробрюшек (Leiognathoides, Gazza) в различных частях Европы и их отсутствие в других регионах в это же время, мы предполагаем, что Западный Тетис можно рассматривать в качестве центра происхождения лиогнатид.
Палеоокеанографические изменения, открытие и закрытие морских путей между океанами влияли на обмен фаунистическими элементами. Ф. Рёгл (Rögl, 1998) считал эти факторы основными движущими силами эволюции морских экосистем. В конце эоцена Тетис был соединен с Индийским океаном и западной частью Тихого океана большим морским путем (Rögl, 1998, 1999). Эта связь была использована рыбами и другими морскими организмами для расселения на запад из Индо-Пацифики (Harzhauser et al., 2007; Банников, 2010). В то же время, сребробрюшки могли использовать этот морской путь, чтобы расселиться в противоположном (восточном) направлении. В раннем олигоцене эти рыбы (напр., Leiognathoides altapinna) были распространены в центральной и восточной частях Паратетиса. Во второй половине олигоцена лиогнатиды, возможно, проникли в Индийский океан (и западную часть Тихого океана), используя морской путь, существовавший до бурдигала (Rögl, 1999; Piller et al., 2007). Мы предполагаем, что видообразование этой группы происходило здесь в миоцене и в последующие эпохи, что привело к высокому разнообразию современных лиогнатид. Представителям этой группы удалось выжить и впоследствии процветать в новом регионе, однако они вымерли в Паратетисе, вероятно, в течение среднего миоцена.
Климатические условия в Карпатском бассейне в раннем миоцене были оптимальными, о чем свидетельствует наличие богатой мелководной теплолюбивой фауны (Kotlarczyk et al., 2006; Попов и др., 2009; Garecka, 2012). Сообщество ископаемых рыб Харты было разнообразным и включало в себя 11 таксонов, относящихся к девяти семействам (Jerzmańska et al., 2001), среди которых преобладающей группой были сельдевые (52.8%). Сребробрюшки находятся на втором месте по обилию остатков, в то время как мерлузовые (Merlucciidae) и тресковые (Gadidae) представлены вдвое меньшим числом особей (Jerzmańska et al., 2001; Kovalchuk et al., 2019). Тесная связь между ихтиофаунами Карпатского и Большого Кавказского бассейнов (Попов и др., 2009; Банников, 2010; Kovalchuk et al., 2019, 2020) подтверждается наличием Leiognathoides minutus в составе сообщества Харты. Полученные данные позволяют значительно расширить известный ранее ареал этого вида в западном направлении и подтверждают возможность обмена фауной между различными частями Паратетиса в раннем миоцене.
Список литературы
Банников А.Ф. Новый ископаемый род сребробрюшковых рыб (Perciformes, Leiognathidae) // Вопр. ихтиол. 2001. Т. 41. № 1. С. 122–124.
Банников А.Ф. Ископаемые позвоночные России и сопредельных стран. Ископаемые колючеперые рыбы (Teleostei, Acanthopterygii). М.: ГЕОС, 2010. 243 с.
Данильченко П.Г. Костистые рыбы майкопских отложений Кавказа. М.: Изд-во АН СССР, 1960. 208 с. (Тр. Палеонтол. ин-та АН СССР. Т. 78).
Данильченко П.Г. Отряд Perciformes (кроме Percidae) // Ископаемые костистые рыбы СССР. М.: Изд-во АН СССР, 1980. С. 125–169 (Тр. Палеонтол. ин-та АН СССР. Т. 178).
Попов С.В., Ахметьев М.А., Лопатин А.В. и др. Палеогеография и биогеография бассейнов Паратетиса. Поздний эоцен–ранний миоцен. М.: Научн. мир, 2009. 194 с. (Тр. Палеонтол. ин-та РАН. Т. 292).
Abraham K.J., Joshi K.K., Murty V.S.R. Taxonomy of the fishes of the family Leiognathidae (Pisces, Teleostei) from the West coast of India // Zootaxa. 2011. V. 2886. P. 1–18.
Agassiz L. Recherches sur les poissons fossiles. T. 4. Neuchâtel: Petitpierre, 1833–1844. 296 p.
Bannikov A.F. The new genus Eoleiognathus for the percoid fish Pygaeus dorsalis Agassiz from the Eocene of Bolca in northern Italy, a putative ponyfish (Perciformes, Leoignathidae) // Studi e Richerche sui Giacimenti Terziari di Bolca. V. 15. Verona, 2014. P. 35–42.
Blow W.H. Late middle Eocene to Recent planktonic foraminiferal biostratigraphy // Proc. of the First Intern. Conf. on Planktonic Microfossils. Leiden: Brill, 1969. P. 199–422.
Chakrabarty P., Sparks J.S. Diagnoses for Leiognathus Lacépède 1802, Equula Cuvier 1815, Equulites Fowler 1904, Eubleekeria Fowler 1904, and a new ponyfish genus (Teleostei: Leiognathidae) // Amer. Mus. Novit. 2008. V. 3623. P. 1–11.
Chakrabarty P., Davis M.P., Smith W.L. et al. Evolution of the light organ system in ponyfishes (Teleostei: Leiognathidae) // J. Morphol. 2011. V. 272. P. 704–721.
Garecka M. Record of changes in the Oligocene-Miocene sediments of the Menilite-Krosno Series of the Skole Unit based on calcareous nannoplankton studies – biostratigraphy and palaeogeographical implications (Polish Outer Carpathians) // Biul. Państw. Inst. Geol. 2012. V. 453. P. 1–22.
Garecka M., Olszewska B. Biostratigraphy of the Early Miocene of the Southern Poland based on planktic foraminifera and calcareous nannoplankton // Przegl. Geol. 1998. V. 46. № 8/2. P. 712–721.
Gill A.C., Leis J.M. Phylogenetic position of the fish genera Lobotes, Datnioides and Hapalogenys, with a reappraisal of acanthuriform composition and relationships based on adult and larval morphology // Zootaxa. 2019. V. 4680. № 1. P. 1–81.
Gill A.C., Michalski S. Osteological evidence for monophyly of the Leiognathidae (Teleostei: Acanthomorpha: Acanthuriformes) // Zootaxa. 2020. V. 4732. № 3. P. 409–421.
Gill T.N. Families and subfamilies of fishes // Mem. Nat. Acad. Sci. Wash. 1893. V. 6. P. 127–138.
Harzhauser M., Kroh A., Mandic O. et al. Biogeographic responses to geodynamics: A key study all around the Oligo-Miocene Tethyan Seaway // Zool. Anz. 2007. V. 246. P. 241–256.
Hess H., Weiler W. Untersuchungen an Fischen aus dem Alttertiär der Umgebung von Basel // Ecl. Geol. Helv. 1955. V. 48. № 9. P. 427–447.
Ikejima K., Ishiguro N.B., Wada M. et al. Molecular phylogeny and possible scenario of ponyfish (Perciformes: Leiognathidae) evolution // Mol. Phylog. Evol. 2004. V. 31. № 3. P. 904–909.
James P.S.B.R. Comparative osteology of the fishes of the family Leiognathidae. Part I. Osteology // Ind. J. Fish. 1985. V. 32. P. 309–358.
Jerzmańska A., Rajchel J., Świdnicka E. Nowe stanowisko ichtiofauny z warstw krośnieńskich jednostki skolskiej (Karpaty fliszowe) // Geologia. 2001. V. 27. № 1. P. 21–37.
Kimura S., Kimura R., Ikejima K. Revision of the genus Nuchequula with descriptions of three new species (Perciformes: Leiognathidae) // Ichthyol. Res. 2008. V. 55. P. 22–42.
Kotlarczyk J., Jerzmańska A., Świdnicka E. et al. A framework of ichthyofaunal ecostratigraphy of the Oligocene–early Miocene strata of the Polish Outer Carpathian basin // Ann. Soc. Geol. Pol. 2006. V. 76. P. 1–111.
Kovalchuk O., Baykina E., Stefaniak K. et al. A systematic revision of herrings (Teleostei, Clupeidae, Clupeinae) from the Oligocene and early Miocene of the Eastern Paratethys and the Carpathian Basin // J. Vert. Paleontol. 2020. V. 40. № 2. https://doi.org/10.1080/02724634.2020.1778710
Kovalchuk O.M., Świdnicka E., Stefaniak K. A new record of Gobius jarosi (Teleostei, Gobiidae) from the Early Miocene of Poland with inference to palaeogeography and palaeoecology of the Carpathian Basin // Hist. Biol. 2019. V. 31. № 10. P. 1394–1401.
Martini E. Standard Tertiary and Quaternary calcareous nannoplankton zonation // Proc. of the II Planktonic Conference, Roma, 1970. V. 2. Rome: Edizioni Tectoscienza, 1971. P. 739–785.
Micklich N., Bannikov A.F., Yabumoto Y. First record of ponyfishes (Perciformes: Leiognathidae) from the Oligocene of the Grube Unterfeld (“Frauenweiler’’) clay pit // Paläontol. Z. 2017. V. 91. P. 375–398.
Nelson J.S., Grande T.C., Wilson M.V.H. Fishes of the World. 5th ed. Hoboken: Wiley, 2016. 707 p.
Nolf D. Otolithi piscium // Handbook of Palaeoichthyology. V. 10. Stuttgart; N.Y.: Fischer, 1985. 145 p.
Nolf D., Brzobohatý R. Fish otoliths from the Late Oligocene (Eger and Kiscell Formations) in the Eger area (northeastern Hungary) // Bull. Inst. Roy. Sci. Natur. Belg., Sci. Terre. 1994. V. 64. P. 225–252.
Olszewska B. Otwornice warstw krośnieńskich w pobliżu V-tego horyzontu tufowego w Radziszowie (Polskie Karpaty zewnętrzne) // Kwart. Geol. 1982. V. 26. P. 627–645.
Pharisat A. La paléoichthyofaune du Rupélien marin de Froidefontaine (Territoire de Belfort). Taxinomie et populations, genèse du gisement, implications paléobiogéographiques // Ann. Sci. Univ. Besançon, Géol. 4 sér. 1991. Fasc. 11. P. 13–97.
Piller W.E., Harzhauser M., Mandic O. Miocene Central Paratethys stratigraphy – current status and future directions // Stratigraphy. 2007. V. 4. № 2/3. P. 151–168.
Prokofiev A.M. New and little known perciform fishes from the Upper Oligocene–Lower Miocene boundary deposits of the Caucasus (Osteichthyes, Perciformes) // Zoosyst. Ross. 2002. V. 11. № 1. P. 209–217.
Rajchel J. Budowa geologiczna doliny Sanu w rejonie Dynów – Dubiecko // Biul. Państw. Inst. Geol. 1989. V. 361. P. 11–53.
Rajchel J., Franus W., Franus M. Mineralogical characteristics of the main tuffite horizon from Harta in the Skole Unit (Flysch Carpathians) // Biul. Państw. Inst. Geol. 2001. V. 396. P. 128–129.
Reichenbacher B., Gregorová R., Holcová K. et al. Discovery of the oldest Gobius (Teleostei, Gobiiformes) from a marine ecosystem of Early Miocene age // J. Syst. Palaeontol. 2018. V. 16. № 6. P. 493–513.
Rögl F. Palaeogeographic considerations for Mediterranean and Paratethys seaways (Oligocene to Miocene) // Ann. Naturhist. Mus. Wien. Ser. A. 1998. Bd 99. P. 279–310.
Rögl F. Mediterranean and Paratethys. Facts and hypotheses of an Oligocene to Miocene paleogeography (short overview) // Geol. Carpath. 1999. V. 50. № 4. P. 339–349.
Rückert-Ülkümen N. Neue Funde von Capros breviventralis Rückert-Ülkümen (Pisces, Zeiformes) aus dem Tertiär von Thrakien, Türkei // N. Jb. Geol. Paläontol. Monatsh. 1994. № 7. P. 443–448.
Schwarzhans W. Otoliths from the Miocene of West Africa, primarily from the Mandorové Formation of Gabon // PalaeoIchthyol. 2013. V. 13. P. 151–184.
Schwarzhans W., Bratishko A. The otoliths from the middle Paleocene of Luzanivka (Cherkasy district, Ukraine) // N. Jb. Geol. Paläontol. Abh. 2011. V. 261. № 1. P. 83–110.
Siegel J. Median fin-spine locking in the ponyfishes (Perciformes: Leiognathidae) // Copeia. 1982. P. 202–205.
Sikora W., Wieser T., Żgiet J. et al. Tuff horizons in the Menilite-Krosno Series of the Flysch Carpathians // Bull. Acad. Pol. Sci., Sér. Sci. Chim. Geol. Géogr. 1959. V. 7. P. 497–503.
Sparks J.S., Chakrabarty P. Description of a new genus of ponyfishes (Teleostei: Leiognathidae), with a review of the current generic-level composition of the family // Zootaxa. 2015. V. 3947. № 2. P. 181–190.
Sparks J.S., Dunlap P.V. A clade of non-sexually dimorphic ponyfishes (Teleostei: Perciformes: Leiognathidae): phylogeny, taxonomy, and description of a new species // Amer. Mus. Novit. 2004. V. 3459. P. 1–21.
Sparks J.S., Dunlap P.V., Smith W.L. Evolution and diversification of a sexually dimorphic luminescent system in ponyfishes (Teleostei: Leiognathidae), including diagnoses for two new genera // Cladistics. 2005. V. 21. P. 305–327.
Steurbaut E. Les otolithes de téléostéens du gisement de Peyrère à Peyrehorade (couches de passage de l’Oligocène au Miocène d’Aquitaine méridionale, France) // Meded. Werkgr. Tert. Kwart. Geol. 1982. V. 19. № 2. P. 35–57.
Van Couvering J.A., Aubry M.-P., Berggren W.A. et al. The terminal Eocene event and the Polish connections // Palaeogeogr., Palaeoclimatol., Palaeoecol. 1981. V. 36. P. 321–362.
Weinfurter E. Die Otolithen der Wetzelsdorfer Schichten und des Florianer Tegels (Miozän, Steiermark) // Sitz. Österr. Akad. Wiss., Math.-Naturwiss. Cl. 1952. Bd 161/7. S. 455–498.
Wieser T. The Teschinite Formation and other evidences of magmatic activity in the Polish Flysch Carpathians and their geotectonic and stratigraphic significance // Fundamental Researches in the Western Part of the Polish Carpathians. Guide to Excursion 1. Carpatho-Balkan Geological Association 13 Congress. Cracow, 1985. P. 23–36.
Yabumoto Y., Uyeno T. A new Miocene ponyfish of the genus Leiognathus (Pisces, Leiognathidae) from Tottori Prefecture, Japan // Bull. Nat. Sci. Mus. Tokyo. Ser. C. 1994. V. 20. № 2. P. 66–77.
Yabumoto Y., Uyeno T. Euleiognathus, a new genus proposed for the Miocene ponyfish, Leiognathus tottori Yabumoto and Uyeno, 1994 (Perciformes: Leiognathidae) from Japan // Ichthyol. Res. 2011. V. 58. P. 19–23.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал