

Палеонтологический журнал, 2021, № 4, стр. 84-93
Новые таксоны птиц (Aves: Galliformes, Gruiformes) из нижнего эоцена Монголии
Н. В. Зеленков *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: nzelen@paleo.ru
Поступила в редакцию 03.10.2020
После доработки 19.11.2020
Принята к публикации 19.11.2020
Аннотация
Описаны новые таксоны птиц из нижнего эоцена пачки бумбан местонахождения Цаган-Хушу в Южной Монголии. Bumbanortyx transitoria gen. et sp. nov. – мелкая курообразная птица, сочетающая морфологическое сходство с ископаемыми семействами Quercymegapodiidae и Gallinuloididae. Несколько более крупный Bumbanipodius magnus gen. et sp. nov. также проявляет сходство с Quercymegapodiidae, но по строению тарсоматетатарсуса близок к Argillipes aurorum из нижнего эоцена Англии. Bumbanipes aramoides gen. et sp. nov. проявляет наибольшее сходство с современными Aramidae (Gruiformes). Bumbaniralla walbeckornithoides gen. et sp. nov. описан по коракоиду, морфологически промежуточному между таковыми Walbeckornis и Messelornithidae (стволовые Gruiformes). Также отмечено присутствие в фауне местонахождения Цаган-Хушу неопределимых представителей Messelornithidae.
Палеонтологическая летопись птиц палеогена Азии очень фрагментарна. Одними из наименее изученных остаются авифауны раннего эоцена, известные всего по нескольким местонахождениям на континенте (см. Mayr, 2009; Зеленков, Курочкин, 2015; Hood et al., 2019). При этом наиболее репрезентативная фауна птиц этого временного интервала в Азии происходит из пачки бумбан местонахождения Цаган-Хушу в Южной Монголии, откуда установлены Presbyornithidae из Anseriformes, Phoenicopterimorphae, Charadriiformes и Strigiformes (Kurochkin, Dyke, 2010; Курочкин, Дайк, 2011; Hood et al., 2019; Зеленков, 2021). Из пачки бумбан также известны мелкие стволовые Galliformes, систематическое положение которых ранее не было установлено (Hwang et al., 2010; Hood et al., 2019). В настоящей статье описываются новые диагностичные материалы по мелким Galliformes из нижнего эоцена местонахождения Цаган-Хушу, собранные в 1980-х гг. отрядами Совместной Советско-Монгольской палеонтологической экспедиции (ССМПЭ) (подробнее см.: Lopatin, 2006) и позволяющие установить присутствие двух новых таксонов родового ранга. Также приведено описание новых таксонов Gruiformes из этого же местонахождения. Эти данные существенно расширяют имеющиеся представления о таксономическом разнообразии раннеэоценовых птиц Центральной Азии.
Настоящая работа следует традиционной линнеевской номенклатуре; стволовые представители включаются в состав современных отрядов. Ориентация сторон костей принята по Nomina Anatomica Avium (Baumel et al., 1993). В соответствии с этим, extremitas omalis коракоида формирует краниальную вершину кости, а его extremitas sternalis – каудальную. Данные по геологии и фауне местонахождения суммированы в ряде предыдущих публикаций (Бадамгарав, Решетов, 1985; Lopatin, 2006; Zelenkov, 2018; Зеленков, 2021 и ссылки в этих работах).
ОТРЯД GALLIFORMES
GALLIFORMES INCERTAE FAMILIAE
Род Bumbanortyx Zelenkov, gen. nov.
Название рода от Bumban – нижнеэоценовая пачка в местонахождении Цаган-Хушу, и Ortyx лат. – перепел; ж. р.
Типовой вид – Bumbanortyx transitoria sp. nov.
Диагноз. В коракоиде cotyla scapularis крупная, неясно-овальная, ориентирована преимущественно дорсально; медиальный край стержня на уровне facies articularis humeralis ориентирован субпараллельно длинной оси кости (слабо наклонен медиально) и формирует неглубокую вырезку вблизи processus acrocoracoideus; pr. acrocoracoideus сильно выступает краниально относительно вершины facies articularis humeralis и медиально – относительно медиального края стержня (за счет медиального сдвига impressio bicipitis); краниальная часть (вершина) impressio lig. acrocoracohumeralis ориентирована дорсокраниально и видна при виде с дорсальной стороны, каудальная часть impressio ориентирована краниально; facies articularis clavicularis расширена краниокаудально; impressio bicipitis заметно выдается вентрально относительно прилегающей вентральной поверхности стержня; facies articularis humeralis короткая (ее длина примерно вдвое превышает ширину), широкая и с тупым краниальным концом; labrum glenoidale сильно выдается вентрально и выпуклая; processus procoracoideus умеренно развит (его ширина примерно соответствует половине ширины cotyla scapularis).
В плечевой кости caput humeri узкая краниокаудально и не формирует вентрального расширения; костный мостик, соединяющий вентральную часть caput и tuberculum ventrale, отсутствует; tuberculum dorsale удлиненный; fossa tricipitalis dorsalis имеется, но не глубокая, caput humeri несильно нависает над ней; fossa tricipitalis не пневматизирована.
В тарсометатарсусе fossae parahypotarsales слабо выражены (латеральная практически не заметна), sulcus extensorius довольно глубокий, огорожен тонким медиальным краем и утолщенным латеральным краем, проксимально переходит в глубокую fossa infracotylaris dorsalis; гипотарсус проксимодистально короткий, дорсоплантарно низкий и по ширине составляет около 2/3 от ширины проксимального эпифиза; канал m. flexor digirotum longus располагается вблизи плантарного края гипотарсуса (не утоплен дорсально); желоба для сухожилий мускулов-сгибателей II пальца и m. flexor hallucis longus ориентированы плантарно, желоб для m. flexor hallucis longus располагается несколько дорсальнее первого. При виде с дорсальной стороны латеральный край кости ориентирован субпараллельно длинной оси кости, в то время как медиальный край формирует перегиб на уровне fossa infracotylaris dorsalis.
Видовой состав. Типовой вид.
Сравнение (рис. 1, в, г, е, к; 2, а, в, д, ж, и; 3, а, б). Коракоид Bumbanortyx общими очертаниями сходен с таковым Quercymegapodius Mourer-Chauviré, 1992, но отличается от него довольно крупным processus procoracoideus (полностью редуцирован у Quercymegapodius: Mourer-Chauviré, 1992), наличием неглубокой, но отчетливой вырезки в медиальном крае стержня кости непосредственно вблизи processus acrocoracoideus (рис. 1, inc), а также тем, что стержень кости не расширяется медиально в краниальном направлении (у Bumbanortyx медиальный край стержня субпараллелен длинной оси кости на уровне facies articularis humeralis). Bumbanortyx отличается от родов Gallinuloides Eastman, 1900 и Paraortygoides Mayr, 2000 (Gallinuloididae) более округлой cotyla scapularis (отчетливо овальная у Gallinuloides и Paraortygoides: Mayr, 2000; Mayr, Weidig, 2004), утоньшенным и более поперечно ориентированным crista acrocoracoidea (утолщенный валикообразный и более продольно ориентированный у Gallinuloides и Paraortygoides; рис. 1, ca) и несколько укороченным extremitas omalis (часть кости, расположенная краниальнее cotyla scapularis, удлинена у Gallinuloides и Paraortygoides; рис. 1, а, б). Bumbanortyx отличается от Paraortygidae более округлой cotyla scapularis (у Paraortygidae, включая Xorazmortyx Zelenkov et Panteleyev, 2019, длинная ось cotyla ориентирована косо по отношению к длинной оси кости: Mourer-Chauviré, 1992; Zelenkov, Panteleyev, 2019; Stidham et al., 2020). Bumbanortyx отличается от типового рода Paraortyx Gaillard, 1908 также тем, что impressio lig. acrocoracohumeralis ориентирован дорсально при виде с дорсальной стороны, и весь processus acrocoracoideus выступает краниально при виде с дорсальной стороны. От Xorazmortyx и Scopelortyx Mourer-Chauviré, Pickford et Senut, 2015, имеющих морфологически отличающийся от Paraortyx коракоид, Bumbanortyx отличается медиально расширенным processus acrocoracoideus при виде с дорсальной стороны (см. Mourer-Chauviré et al., 2015; Zelenkov, Panteleyev, 2019).
Рис. 1.
Коракоиды избранных представителей ископаемых Galliformes: а – Gallinuloides wyomingensis Eastman, 1900, экз. Wyoming Dinosaur Center (Термополис, США), № CGR-012; США; нижний эоцен, формация Грин Ривер (по: Mayr, Weidig, 2004, c изменениями); б – Paraortygoides messelensis Mayr, 2000, голотип Senckenberg Forschungsinstitut und Naturmuseum (Франкфурт-на-Майне, Германия), № ME 1303a (фото автора); Германия, местонахождение Мессель; нижний эоцен; в, г, е, к – Bumbanortyx transitoria gen. et sp. nov., голотип ПИН, № 3104/265, краниальный фрагмент левого коракоида; д, ж, л – Bumbanipodius magnus gen. et sp. nov., экз. ПИН, 3104/195, краниальный фрагмент левого коракоида; Монголия, местонахождение Цаган-Хушу; нижний эоцен; з – Quercymegapodius brodkorbi Mourer-Chauviré, 1992, экз. в колл. Ун-та наук и техник Лангедока (Монпелье, Франция), № BFI 1849, краниальный фрагмент левого коракоида; Франция, местонахождение la Bouffie; верхний эоцен; и – Xorazmortyx turkestanensis Zelenkov et Panteleyev, 2019, голотип Зоол. ин-т РАН (С.-Петербург), № 4991, краниальный фрагмент правого коракоида (отражен); Узбекистан, местонахождение Джерой 2; средний эоцен. а–в – вид с дорсомедиальной стороны, г, д – вид с латеральной стороны; е–и – вид с дорсальной стороны; к, л – вид с вентральной стороны. Обозначения: ca – crista acrocoracoidea; cs – cotyla scapularis; fah – facies articularis humeralis; ib – impressio bicipitis; ila – impressio lig. acrocoracohumeralis; inc – вырезка в медиальном крае стержня; pp – processus procoracoideus.
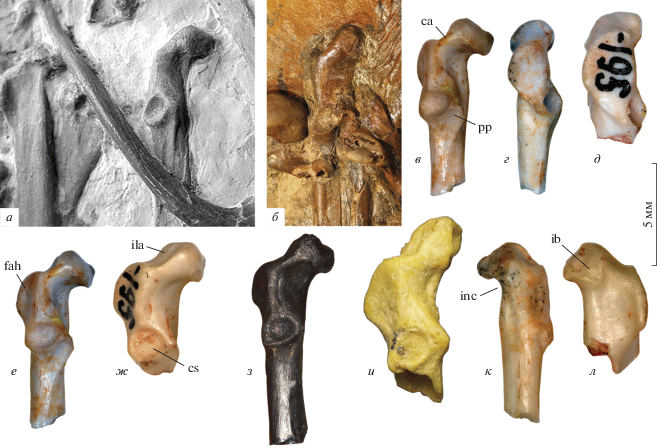
Рис. 2.
Проксимальные фрагменты тарсометатарсусов избранных представителей ископаемых Galliformes: а, в, д, ж, и – Bumbanortyx transitoria gen. et sp. nov., экз. ПИН, № 3104/663; б, г, е, з, к – Bumbanipodius magnus gen. et sp. nov., голотип ПИН, № 3104/183; Монголия, местонахождение Цаган-Хушу; нижний эоцен; л – Argillipes aurorum Harrison et Walker, 1977, голотип Муз. естественной истории в Лондоне, № A 3130; Англия, о-в Шеппи, местонахождение Лондон Клэй; нижний эоцен (по: Mayr, Smith, 2019, с изменениями); м – Ameripodius alexis Mourer-Chauviré, 2000 (Quercymegapodiidae), экз. Нац. муз. Естественной истории в Париже, № SG 9342; Франция, местонахождение Сан-Жеран-ле-Пюи; нижний миоцен (фото автора). а, б – вид с дорсальной стороны; в, г – вид с латеральной стороны; д, е – вид с плантарной стороны; ж, з – вид с медиальной стороны; и–м – вид с проксимальной стороны (не в масштабе). Обозначения: cl – cotyla lateralis; cm – cotyla medialis; fdl – канал для сухожилия m. flexor digitorum longus; fhl – борозда для сухожилия m. flexor hallucis longus; fp2 – борозда для сухожилий m. flexor perforatus digiti II и/или m. flexor perforans et perforatus digiti II; fpl – fossa parahypotarsalis lateralis; fpm – fossa parahypotarsalis medialis; hyp – hypotarsus; lcm – бугорок ligamentum collaterale mediale.
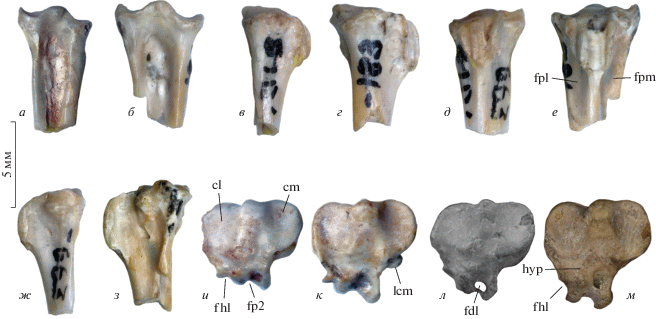
Рис. 3.
Ископаемые Galliformes и Gruiformes: а, б – Bumbanortyx transitoria gen. et sp. nov., экз. ПИН, № 3104/128, проксимальный фрагмент левой плечевой кости; в–д – Bumbanipes aramoides gen. et sp. nov., голотип ПИН, № 3104/665, дистальный фрагмент правого тарсометатарсуса; е–з – Bumbaniralla walbeckornithoides gen. et sp. nov., голотип ПИН, № 3104/264, краниальный фрагмент левого коракоида; и – Messelornithidae indet.; экз. ПИН, № 3104/567, фрагмент левого коракоида; Монголия, местонахождение Цаган-Хушу; нижний эоцен; к, л – Itardiornis hessae Mourer-Chauviré, 1995, экз. Ун-т наук и техник Лангедока (Монпелье, Франция), № ITD 679, краниальный фрагмент левого коракоида; Франция, местонахождение Итардиес; нижний олигоцен (фото автора). а – Вид с каудальной стороны; б – вид с краниальной стороны; в, ж, и, к – вид с дорсальной стороны; г – вид с плантарной стороны; д – вид с дистальной стороны (не в масштабе); е – вид с медиальной стороны; з, л – вид с вентральной стороны. Обозначения: ap – вершина facies articularis humeralis; bpp – каудальный край основания processus procoracoideus; ch – caput humeri; cs – cotyla scapularis; fah – facies articularis humeralis; fd – fossa tricipitalis dorsalis; fns – foramen n. supracoracoidei; h – крючок processus acrocoracoideus; inc – вырезка в медиальном крае стержня; lg – labrum glenoidale; pa – processus acrocoracoideus; tm2–4 – trochleae metatarsorum II–IV; tv – tuberculum ventrale. Длина масштабной линейки – 5 мм.

Плечевая кость Bumbanortyx отличается от таковой всех других семейств курообразных узкой (не расширенной краниокаудально) caput humeri (состояние не известно для Gallinuloides). От раннеэоценового Paraortygoides (Gallinuloididae) также отличается мелкой fossa tricipitalis dorsalis.
Тарсометатарсус Bumbanortyx отличается от такового Paraortygoides более центрально расположенным гипотарсусом (у Paraortygoides он смещен латерально: Mayr, 2016, рис. 2E); от представителей семейств Quercymegapodiidae и Paraortygidae – дорсоплантарно низким гипотарсусом. От Argillipes aurorum Harrison et Walker, 1977 отличается медиолатерально более широким гипотарсусом за счет преимущественно плантарной ориентации желоба для m. flexor hallucis longus (у Argillipes aurorum гипотарсус более узкий за счет того, что желоб для m. flexor hallucis longus ориентирован в значительной степени латерально; Harrison, Walker, 1977; Mayr, Smith, 2019). Помимо этого, медиальный край тарсометатарсуса не формирует расширения при переходе на проксимальный эпифиз у Argillipes aurorum, в то время как перегиб на уровне fossa infracotylaris dorsalis отчетливо выражен у Bumbanortyx. От Percolinus Harrison et Walker, 1977 отличается дорсоплантарно низким гипотарсусом и наличием перегиба в медиальном крае при переходе на проксимальный эпифиз.
Замечания. Bumbanortyx представляет стволовых курообразных, поскольку имеет сильновогнутую округлую cotyla scapularis, как у трех ископаемых семейств – Paraortygidae, Gallinuloididae и Quercymegapodiidae (Mourer-Chauviré, 1992; Mayr, 2000). При этом общими очертаниями и морфологией Bumbanortyx ближе всего к Quercymegapodius, с которым он сближается крупной округлой и расположенной строго дорсально cotyla scapularis (у Paraortygidae и Gallinuloididae cotyla scapularis овальная и несколько смещена латерально), сильно выдающимся краниально processus acrocoracoideus, а также короткой, широкой и сильно выступающей вентрально (формирующей высокую labrum glenoidale) facies articularis humeralis (см. Mourer-Chauviré, 1992; Alvarenga, 1995). У Quercymegapodius полностью редуцирован processus procoracoideus, однако этот отросток имеется у Ameripodius silvasantosi Alvarenga, 1995 из олигоцена Бразилии, который также сближается с Bumbanortyx прямым стержнем в краниальной части кости (см. Alvarenga, 1995). Ранее отсутствие processus procoracoideus рассматривалось как характерная черта Quercymegapodiidae (Mourer-Chauviré, 1992), однако присутствие выраженного processus procoracoideus у Ameripodius silvasantosi и в меньшей степени у A. alexis Mourer-Chauviré, 2000 указывает на изменчивость этого признака у Quercymegapodiidae.
Наличие неглубокой, но отчетливой вырезки в медиальном крае стержня коракоида вблизи processus acrocoracoideus и относительно крупного processus procoracoideus позволяет сближать Bumbanortyx с Gallinuloididae при общем структурном сходстве с Quercymegapodiidae. Таким образом, коракоид Bumbanortyx демонстрирует морфологическую мозаику признаков Gallinuloididae и Quercymegapodiidae и может рассматриваться как промежуточное звено между этими двумя семействами. При этом Gallinuloididae известны из раннего эоцена, а Quercymegapodiidae – только начиная с позднего эоцена (Mourer-Chauviré, 1992; Mayr, 2009), поэтому морфологические черты, общие между Bumbanortyx и Gallinuloididae, могут представлять собой плезиоморфии, а сходство между Bumbanortyx и Quercymegapodiidae, очевидно, является продвинутым. Тем не менее, промежуточная морфология Bumbanortyx указывает на то, что морфологический тип коракоида Quercymegapodiidae сформировался на основе такового Gallinuloididae.
Ранее еще один коракоид из пачки бумбан (экз. IMG, № 100/1371) был описан как предполагаемый представитель семейства Quercymegapodiidae (Hood et al., 2019). Экз. IMG, № 100/1371 в целом характеризуется сходной морфологией, но отличается от описываемого в этой работе несколько более крупными размерами и отсутствием вырезки в медиальном краю стержня, и может представлять отдельный вид.
Наиболее характерной морфологической чертой Bumbanortyx является узкая (не имеющая вентрального расширения) caput humeri плечевой кости, не характерная ни для каких других известных представителей Galliformes. Эта особенность ранее отмечалась для неназванного представителя отряда из пачки бумбан (Hwang et al., 2010), которого я здесь отношу к Bumbanortyx. Наличие такой же caput humeri недавно было отмечено для мелкой фазановой птицы из нижнего эоцена Бельгии, предположительно отнесенной к Argillipes (Mayr, Smith, 2019). Проявление данного признака у ряда раннеэоценовых курообразных может свидетельствовать о том, что они представляют отдельную группу уровня семейства. В то же время, необходимо отметить, что строение caput humeri не известно для Gallinuloides, однако с этим родом Bumbanortyx сближается в строении коракоида – таким образом, нельзя исключить, что Bumbanortyx представляет Gallinuloididae. С другой стороны, общее структурное сходство с коракоидом более молодого рода Quercymegapodius может быть апоморфным – в таком случае Bumbanortyx может оказаться примитивным представителем Quercymegapodiidae.
Тарсометатарсусы, относимые к Bumbanortyx transitoria, общими пропорциями сходны с таковым Argillipes aurorum, но отличаются формой гипотарсуса и наличием перегиба в медиальном крае кости при переходе на проксимальный эпифиз (см. рубрику “Сравнение”). Мелкая курообразная птица, сходная с Argillipes aurorum по строению тарсометатарсуса и также имеющая узкую caput humeri, известна из раннего эоцена Бельгии (Mayr, Smith, 2019). Для бельгийской птицы также характерен указанный перегиб, как у Bumbanortyx, но иная форма гипотарсуса. Таким образом, неназванный бельгийский таксон и Argillipes aurorum, по-видимому, представляют собой формы, близкие к Bumbanortyx transitoria. При этом отличия в строении тарсометатарсуса от Paraortygoides не позволяют сближать Bumbanortyx с Gallinuloididae. “Argillipes” paralectoris Harrison et Walker, 1977, скорее всего, представляет отдельный род, и его отнесение к Galliformes требует подтверждения (Mayr, Smith, 2019).
По общей организации проксимального эпифиза тарсометатарсуса и, в частности, гипотарсуса, Bumbanortyx сближается с современными Cracidae, от которых отличается плантарным положением и небольшим диаметром канала m. flexor digitorum longus, а также почти не выраженными fossae parahypotarsales. Последнее свидетельствует о слабо развитых коротких мускулах стопы и, следовательно, иной локомоторной специализации Bumbanortyx в сравнении с современными Cracidae.
Bumbanortyx transitoria Zelenkov, sp. nov.
Название вида от transitorius лат. – переходный.
Голотип – ПИН, № 3104/265, краниальный фрагмент левого коракоида; Монголия, местонахождение Цаган-Хушу; нижний эоцен, пачка бумбан. Сборы ССМПЭ, 1980 г.
Описание (рис. 1, в, г, е, к; 2, а, в, д, ж, и; 3, а, б). Совпадает с диагнозом рода (который также составляет диагноз нового вида).
Размеры в мм. Коракоид: длина от краниальной вершины кости до каудального края cotyla scapularis 5.6; дорсовентральная высота на уровне cotyla scapularis 2.2. Плечевая кость: ширина проксимального эпифиза 7.8; краниокаудальная высота caput humeri 2.5. Тарсометатарсус: ширина проксимального эпифиза 4.4.
Сравнение. В роде Bumbanortyx один вид.
Замечания. К B. transitoria на основании сходных относительных размеров я отношу плечевую кость и тарсометатарсус. Дорсовентральная ширина caput humeri у экз. ПИН, № 3104/663 и длина гленоидной части коракоида (от вершины facies articularis humeralis до вершины processus procoracoideus) у голотипа соответствуют некрупным экз. современного Coturnix coturnix. Вторая курообразная птица из Цаган-Хушу (Bumbanipodius magnus gen. et sp. nov.) имеет отчетливо большие размеры.
Материал. Кроме голотипа, из типового местонахождения: экз. ПИН, № 3104/128, проксимальный фрагмент левой плечевой кости; экз. ПИН, № 3104/663, проксимальный фрагмент левого тарсометатарсуса.
Род Bumbanipodius Zelenkov, gen. nov.
Название рода от Bumban – нижнеэоценовая пачка в местонахождении Цаган-Хушу, и Megapodius – современный род Galliformes; м. р.
Типовой вид – Bumbanipodius magnus sp. nov.
Диагноз. В коракоиде cotyla scapularis крупная, округлая, ориентирована преимущественно дорсально; processus acrocoracoideus короткий и очень незначительно выступает краниально относительно вершины facies articularis humeralis; impressio lig. acrocoracohumeralis ориентирован в значительной степени дорсально и отчетливо виден при виде с дорсальной стороны; facies articularis clavicularis расширена краниокаудально; impressio bicipitis несущественно выдается вентрально относительно прилегающей вентральной поверхности стержня и слабо выдвинут медиально относительно прилегающей части стержня; facies articularis humeralis короткая (ее длина примерно вдвое превышает ширину), широкая и с тупым краниальным концом; labrum glenoidale сильно выдается вентрально и выпуклая.
В тарсометатарсусе fossa parahypotarsalis medialis умеренно выражена, fossa parahypotarsalis lateralis едва заметна; sulcus extensorius довольно глубокий, огорожен тонким медиальным краем и утолщенным латеральным краем, проксимально переходит в глубокую fossa infracotylaris dorsalis; hypotarsus проксимодистально короткий, умеренно высокий дорсоплантарно, по ширине составляет около 2/3 от ширины проксимального эпифиза; канал m. flexor digitorum longus крупный и несколько утоплен дорсально; желоб для сухожилия длинных сгибателей второго пальца (m. flexor perforatus digiti II и/или m. flexor perforans et perforatus digiti II) ориентирован плантарно и расположен заметно плантарнее, чем желоб m. flexor hallucis longus; желоб m. flexor hallucis longus ориентирован латероплантарно; желоб для m. fibularis longus отчетливо выражен. При виде с дорсальной стороны стержень равномерно расширяется проксимально при переходе на проксимальный эпифиз.
Видовой состав. В роде Bumbanipodius один вид.
Сравнение (рис. 1, д, ж, л; 2, б, г, е, з, к). Коракоид Bumbanipodius отличается от всех других палеогеновых курообразных коротким и слабо выдающимся краниально processus acrocoracoideus, который при этом слабо выдается медиально относительно медиального края стержня. От Paraortygoides и Gallinuloides (сем. Gallinuloididae) помимо этого отличается округлой и дорсально ориентированной cotyla scapularis; от Bumbortyx и Gallinuloides также отличается отсутствием вырезки в медиальном крае стержня непосредственно у processus acrocoracoideus (рис. 1, inc); от Quercymegapodius – также расширенным processus procoracoideus (степень развития отростка у Bumbanipodius не ясна).
Тарсометатарсус Bumbanipodius отличается от всех других палеогеновых курообразных коротким и медиоплантарно ориентированным бугорком для крепления lig. collaterale mediale при виде с проксимальной стороны (у других представителей этот бугорок или не виден, или имеет вид удлиненной площадки; рис. 2, lcm). От Paraortygoides также отличается более центральным расположением гипотарсуса; от Ameripodius – более широким и несколько более низким гипотарсусом с более плантарно расположенным желобом длинных сгибателей второго пальца, а также наличием желоба для m. fibularis longus; от Quercymegapodius – заметно более низким гипотарсусом и дорсальным смещением желоба длинных сгибателей второго пальца. От Argillipes aurorum – также ровным латеральным краем кости при виде с проксимальной стороны и маленьким желобом длинных сгибателей второго пальца (у Argillipes aurorum плантарная часть cotyla lateralis выдается латерально, в результате чего латеральный край кости формирует выступ при виде с проксимальной стороны). От Percolinus также отличается более симметричным гипотарсусом (у Percolinus латеральный край гипотарсуса редуцирован). От Bumbanortyx – также высоким гипотарсусом, расширенным в своей плантарной половине cotyla medialis (у Bumbanortyx плантарная часть cotyla усечена), более плантарно располагающимся желобом длинных сгибателей второго пальца, а также равномерно расширяющимся стержнем при переходе на эпифиз при виде с дорсальной стороны.
Замечания. Коракоид Bumbanipodius сочетает черты Quercymegapodiidae и Gallinuloididae, при этом с последними он сходен утолщенным crista acrocoracoidei. Однако в целом строение плечевой части коракоида не позволяет однозначно сближать Bumbanipodius ни с одним из семейств.
Тарсометатарсус Bumbanipodius в общих чертах сходен с таковым Argillipes aurorum (Harrison, Walker, 1977) и представляется более близкой к Argillipes формой, чем Bumbanortyx. Bumbanipodius и Argillipes aurorum сближаются строением гипотарсуса, сходным уровнем развития fossae parahypotarsales, а также формой стержня при переходе на эпифиз при виде с дорсальной стороны. При этом Bumbanipodius отличается от Argillipes aurorum формой cotyla lateralis, геометрией гипотарсуса и укороченным бугорком для lig. collaterale mediale. Короткий бугорок также имеется у современных Megapodius Gaimard, 1823 (семейство Megapodiidae), где он располагается дорсальнее и ориентирован строго латерально.
Bumbanipodius magnus Zelenkov, sp. nov.
Название вида magnus лат. – крупный.
Голотип – ПИН, № 3104/183, проксимальный фрагмент левого тарсометатарсуса; Монголия, местонахождение Цаган-Хушу; нижний эоцен, пачка бумбан. Сборы ССМПЭ, 1980 г.
Описание (рис. 1, д, ж, л; 2, б, г, е, з, к). Совпадает с диагнозом рода (который также составляет диагноз нового вида).
Размеры в мм. Тарсометатарсус: ширина проксимального эпифиза 5.2. Коракоид: длина от краниальной вершины кости до каудального края cotyla scapularis 6.6; дорсовентральная высота на уровне cotyla scapularis 2.8.
Сравнение. В роде Bumbanipodius один вид.
Замечания. Bumbanipodius magnus – более крупная форма по сравнению с Bumbanortyx transitoria (размером с современного Ptilopachus petrosus Gmelin, 1789). На этом основании к B. magnus отнесен относительно крупный фрагментарный коракоид из типового местонахождения, подтверждающий обособленный родовой статус этого вида.
Материал. Кроме голотипа, из типового местонахождения экз. ПИН, № 3104/195, краниальный фрагмент левого коракоида.
ОТРЯД GRUIFORMES
GRUIFORMES INCERTAE FAMILIAE
Род Bumbanipes Zelenkov, gen. nov.
Название рода от Bumban – нижнеэоценовая пачка в местонахождении Цаган-Хушу, и pes лат. – стопа; м. р.
Типовой вид – Bumbanipes aramoides sp. nov.
Диагноз. Стержень тарсометатарсуса широкий, дистальный эпифиз несущественно расширен относительно стержня; trochlea metatarsi III медиолатерально широкий; trochlea metatarsi IV узкий медиолатерально (заметно уже trochlea metatarsi III), его дорсальный край располагается примерно на одном уровне с дорсальным краем trochlea metatarsi III; trochlea metatarsi II сдвинут заметно проксимальнее относительно trochlea metatarsi IV и умеренно смещен плантарно (его дорсальный край располагается на уровне середины trochlea metatarsi III).
Видовой состав. Типовой вид.
Сравнение (рис. 3, в–д). От всех представителей Gruiformes, кроме Heliornithidae, отличается медиолатерально суженным trochlea metatarsi IV при виде с дистальной стороны. Кроме того, trochlea metatarsi IV очень несущественно отставлен латерально, в результате чего дистальный эпифиз слабо расширен (близкое состояние характеризует некоторых Rallidae s.l.). От Parvigruidae также отличается тем, что при виде с дистальной стороны trochlea metatarsi IV располагается примерно на одном уровне с trochlea metatarsi III (смещен плантарно у Parvigruidae) и его дорсальный край асимметричен (латеральный валик сочленовной поверхности выступает несколько сильнее, чем медиальный), а также тем, что trochlea metatarsi II несильно смещен плантарно.
Замечания. Общими пропорциями дистального эпифиза тарсометатарсуса Bumbanipes сходен с примитивными гусеобразными Presbyornis, но отличается от этого рода и других Anseriformes тем, что trochlea metatarsi III медиолатерально широкий при виде с дистальной и дорсальной сторон (у гусеобразных и Presbyornis – сужен), а при виде с плантарной стороны его суставная поверхность субквадратная (у гусеобразных и Presbyornis – субтреугольная с проксимальным заостренным концом). Также, в отличие от Presbyornis и гусеобразных, стержень кости довольно широкий, поэтому дистальный эпифиз практически не выглядит расширенным по отношению к нему. У Presbyornis и большинства гусеобразных trochlea metatarsi III формирует отчетливую дорсальную выпуклость при виде с медиальной или латеральной стороны, что не выражено у Bumbanipes.
Характерный для Bumbanipes широкий trochlea metatarsi III с субквадратной плантарной сочленовной поверхностью характеризует большинство Gruiformes (за исключением Aramidae и части Rallidae, у которых плантарная поверхность бывает треугольной). При этом из Gruiformes по общим пропорциям Bumbanipes ближе всего к Aramidae и ископаемому семейству Parvigruidae, от которых отличается в первую очередь практически не расширенным дистальным эпифизом и узким trochlea metatarsi IV (от Aramidae – также субквадратной плантарной суставной поверхностью trochlea metatarsi III). Сходство с Aramidae проявляется и в положении trochlea metatarsi II (у Aramidae и Bumbanipes умеренно сдвинут плантарно, у Parvigruidae – сильно). Среди журавлеобразных узкий trochlea metatarsi IV характерен для Heliornithidae, что, несомненно, представляет собой адаптацию к плаванию у этих специализированных птиц, имеющих сильно преобразованный тарсометатарсус. Parvigruidae и некоторые Rallidae имеют относительно зауженный trochlea metatarsi IV, но он все равно шире, чем у Bumbanipes. В то же время, дорсальный край trochlea metatarsi IV располагается дорсальнее у Bumbanipes, чем у Parvigruidae. Крупное плантарное пневматическое отверстие характеризует ряд различных представителей журавлеобразных. Таким образом, положение Bumbanipes в системе Gruiformes пока не ясно и требует новых материалов. При этом узкий trochlea metatarsi IV характеризует разнообразных плавающих птиц и указывает на вероятно сходный образ жизни у Bumbanipes.
Bumbanipes aramoides Zelenkov, sp. nov.
Название вида – от современного рода Aramus Vielillot, 1816.
Голотип – ПИН, № 3104/665, дистальный фрагмент правого тарсометатарсуса; Монголия, местонахождение Цаган-Хушу; нижний эоцен, пачка бумбан. Сборы ССМПЭ, 1984 г.
Описание (рис. 3, в–д). Совпадает с диагнозом рода (который также составляет диагноз нового вида).
Размеры в мм. Ширина дистального эпифиза тарсометатарсуса 10.6, ширина trochlea metatarsi III 4.6, максимальная высота trochlea metatarsi III 6.1.
Сравнение. В роде Bumbanipes один вид.
Материал. Голотип.
Род Bumbaniralla Zelenkov, gen. nov.
Название рода от Bumban – нижнеэоценовая пачка в местонахождении Цаган-Хушу, и rallus лат. – пастушок; ж. р.
Типовой вид – Bumbaniralla walbeckornithoides sp. nov.
Диагноз. Коракоид имеет стройные очертания, медиальный край стержня при виде с вентральной стороны прямой, дорсальная поверхность стержня уплощенная; processus acrocoracoideus существенно выдвинут медиально, крючок практически не выражен; наиболее расширенная часть facies articularis humeralis располагается вблизи cotyla scapularis; cotyla scapularis медиолатерально расширена и глубоковыямчатая; processus procoracoideus с умеренно расширенным основанием, crista procoracoidei (sensu Livezey, Zusi, 2006) оканчивается в средней части стержня; foramen n. supracoracoidei некрупное, расположено вблизи cotyla scapularis.
Видовой состав. Типовой вид.
Сравнение (рис. 3, е–з). От Walbeckornis Mayr, 2007 отличается расширенным основанием processus procoracoideus (рис. 3, bpp), слабо выраженным крючком (рис. 3, h) processus acrocoracoideus и краниокаудально удлиненным facies articularis humeralis, вершина которого (рис. 3, ap) располагается на уровне cotyla scapularis (у Walbeckornis вершина facies сдвинута краниально относительно cotyla: Mayr, 2007). От родов Messelornis Hesse, 1988, Pellornis Bertelli, Chiappe et Mayr, 2011 и Itardiornis Mourer-Chauviré, 1995 (семейство Messelornithidae) отличается краниально смещенным foramen n. supracoracoidei, которое у Bumbaniralla приближено к cotyla scapularis; от Messelornis и Itardiornis – также отсутствием вырезки на медиальном крае стержня коракоида вблизи processus acrocoracoideus (заметно при виде с вентральной стороны: рис. 3, inc); от Pellornis – также менее выдающейся латерально labrum glenoidale при виде с вентральной стороны; от Itardiornis – также тем, что вершина facies articularis humeralis сдвинута каудально и находится на уровне cotyla scapularis (у Itardiornis – сдвинута краниально относительно cotyla). От Songzia Hou, 1990 отличается более широким processus procoracoideus.
Замечания. Общими пропорциями и морфологией коракоид Bumbaniralla сходен с таковым Walbeckornis и Messelornithidae (см. Hesse, 1990; Mourer-Chauviré, 1995; Mayr, 2004, 2007, 2016; Bertelli et al., 2011; Wang et al., 2012). С Walbeckornis новый род сближается краниально расположенным foramen n. supracoracoidei (у Messelornithidae отверстие сдвинуто заметно каудальнее), в то время как расширенное основание processus procoracoideus, напротив, сближает Bumbaniralla с Messelornithidae. Таким образом, коракоид Bumbaniralla демонстрирует в некоторой степени промежуточную морфологию и подтверждает возможную близость Walbeckornis к Messelornithidae (Mayr, 2007, 2017). При этом примечательно, что в сборах из Цаган-Хушу также присутствуют кости неопределимых Messelornithidae (рис. 3, и), что впервые показывает сосуществование этих двух групп примитивных Gruiformes.
Bumbaniralla walbeckornithoides Zelenkov, sp. nov.
Название вида от Walbeckornis, морфологически близкого к новому таксону ископаемого рода птиц.
Голотип – ПИН, № 3104/264, краниальный фрагмент левого коракоида; Монголия, местонахождение Цаган-Хушу; нижний эоцен, пачка бумбан. Сборы ССМПЭ, 1980 г.
Описание (рис. 3, е–з). Совпадает с диагнозом рода (который также составляет диагноз нового вида).
Размеры в мм. Общая длина сохранившегося фрагмента коракоида 9.7, длина от краниальной вершины до каудального края cotyla scapularis 4.2, минимальная ширина стержня 1.7.
Сравнение. В роде Bumbaniralla один вид.
Материал. Кроме голотипа, из типового местонахождения экз. ПИН, №№ 3104/266, 267, краниальные фрагменты правых коракоидов.
Список литературы
Бадамгарав Д., Решетов В.Ю. Палеонтология и стратиграфия палеогена Заалтайской Гоби. М.: Наука, 1985. 104 с. (Тр. Совм. Сов.-Монгол. палеонтол. экспед. Вып. 25).
Зеленков Н.В. Ревизия палеоцен–эоценовых монгольских Presbyornithidae (Aves: Anseriformes) // Палеонтол. журн. 2021. № 3. С. 83–91.
Зеленков Н.В., Курочкин Е.Н. Класс Aves // Ископаемые рептилии и птицы. Часть 3 / Ред. Курочкин Е.Н., Лопатин А.В., Зеленков Н.В. М.: ГЕОС, 2015. С. 86–290.
Курочкин Е.Н., Дайк Г.Дж. Первые ископаемые совы (Aves: Strigiformes) из палеогена Азии и обзор палеонтологической летописи отряда совообразных // Палеонтол. журн. 2011. № 4. С. 85–97.
Alvarenga H.M. Um primitive membro de ordem Galliformes (Aves) do Terciário médio de Bacia de Taubaté, Estado de São Paulo, Brasil // An. Acad. Bras. Ciênc. 1995. V. 67. P. 33–44.
Baumel J.J., King A.S., Breazile J.E. et al. Handbook of Avian Anatomy: Nomina Anatomica Avium. Cambridge, Mass.: Nuttall Ornithological Club, 1993. 779 p.
Bertelli S., Chiappe L.M., Mayr G. A new Messel rail from the Early Eocene Fur Formation of Denmark (Aves, Messelornithidae) // J. Syst. Palaeontol. 2011. V. 9. P. 551–562.
Harrison C.J.O., Walker C.A. Birds of the British Lower Eocene // Tert. Res. Spec. Pap. 1977. № 3. P. 1–52.
Hesse A. Die Beschreibung der Messelornithidae (Aves: Gruiformes: Rhynocheti) aus dem Alttertiär Europas und Nordamerikas // Cour. Forsch. Senckenb. 1990. Bd 128. S. 1–176.
Hood S.C., Torres C.R., Norell M.A., Clarke J.A. New fossil birds from the earliest Eocene of Mongolia // Amer. Mus. Novit. 2019. № 3934. P. 1–24.
Hwang S.H., Mayr G., Bolortsetseg M. The earliest record of a galliform bird in Asia, from the late Paleocene–early Eocene of the Gobi desert, Mongolia // J. Vertebr. Paleontol. 2010. V. 30. P. 1642–1644.
Kurochkin E.N., Dyke G.J. A large collection of Presbyornis (Aves, Anseriformes, Presbyornithidae) from the late Paleocene and early Eocene of Mongolia // Geol. J. 2010. V. 45. P. 375–387.
Livezey B.C., Zusi R.L. Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. I. // Bull. Carn. Mus. Natur. Hist. 2006. № 37. P. 1–544.
Lopatin A.V. Early Paleogene insectivore mammals of Asia and establishment of the major groups of Insectivora // Paleontol. J. 2006. V. 40. Suppl. 3. P. 205–405.
Mayr G. A new basal galliform bird from the middle Eocene of Messel (Hessen, Germany) // Senckenb. Leth. 2000. Bd 80. P. 45–57.
Mayr G. Phylogenetic relationships of the early Tertiary Messel rails (Aves, Messelornithidae) // Senckenb. Leth. 2004. Bd 84. P. 317–322.
Mayr G. The birds from the Palaeocene Fissure filling of Walbeck (Germany) // J. Vertebr. Paleontol. 2007. V. 27. P. 394–408.
Mayr G. Paleogene fossil birds. Berlin, Heidelberg: Springer, 2009. 262 p.
Mayr G. Variations in the hypotarsus morphology of birds and their evolutionary significance // Acta Zool. 2016. V. 97. P. 196–210.
Mayr G. Avian Evolution. The Fossil Record of Birds and Its Paleobiological Significance. Chichester, West Sussex: J. Wiley & Sons, 2017. 293 p.
Mayr G., Smith T. A diverse bird assemblage from the Ypresian of Belgium furthers knowledge of early Eocene avifaunas of the North Sea Basin // N. Jb. Geol. Paläontol. Abh. 2019. Bd 291. P. 253–281.
Mayr G., Weidig I. The early Eocene bird Gallinuloides wyomingensis – a stem group representative of Galliformes // Acta Palaeontol. Pol. 2004. V. 49. P. 211–217.
Mourer-Chauviré C. The Galliformes (Aves) from the Phosphorites du Quercy (France): systematics and biostratigraphy // Natur. Hist. Mus. Los Angeles Cty Sci. Ser. 1992. № 36. P. 67–95.
Mourer-Chauviré C. The Messelornithidae (Aves: Gruiformes) from the Paleogene of France // Cour. Forsch. Senckenb. 1995. Bd 181. P. 95–105.
Mourer-Chauviré C., Pickford M., Senut B. Stem group galliform and stem group psittaciform birds (Aves, Galliformes, Paraortygidae, and Psittaciformes, family incertae sedis) from the Middle Eocene of Namibia // J. Ornithol. 2015. V. 156. P. 275–286.
Stidham T.A., Townsend K.E.B., Holroyd P.A. Evidence for wide dispersal in a stem galliform clade from a new small-sized middle Eocene pangalliform (Aves: Paraortygidae) from the Uinta Basin of Utah (USA) // Diversity. 2020. V. 12: 90. https://doi.org/10.3390/d12030090
Wang M., Mayr G., Zhang J., Zhou Z. Two new skeletons of the enigmatic, rail-like avian taxon Songzia Hou, 1990 (Songziidae) from the early Eocene of China // Alcheringa. 2012. V. 36. P. 487–499.
Zelenkov N.V. A swan-sized anseriform bird from the late Paleocene of Mongolia // J. Vertebr. Paleontol. 2018. V. 38: e1531879.
Zelenkov N.V., Panteleyev A.V. A small stem-galliform bird (Aves: Paraortygidae) from the Eocene of Uzbekistan // C.R. Palevol. 2019. V. 18. P. 517–523.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал