

Палеонтологический журнал, 2021, № 6, стр. 63-75
Пермские триаплиды (Coleoptera, Triaplidae), еще одна мезозойская группа жуков в терминальной перми
А. Г. Пономаренко *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: aponom@paleo.ru
Поступила в редакцию 23.05.2020
После доработки 28.09.2020
Принята к публикации 06.10.2020
Аннотация
Пересмотрено систематическое положение жуков Tomiaplus gen. nov., Т. sibiricus (Volkov, 2013), T. minimus sр. nov., Tunguskagyrus Yan, Beutel et Lawrence, 2018, T. planus Yan et al., 2018, T. уani sp. nov., описанных из отложений, близких к пермо-триасовой границе в Тунгусском и Кузнецком бассейнах; эти таксоны предлагается рассматривать принадлежащими к семейству Triaplidae, ранее известному только из верхнего триаса. Поддерживается мнение о том, что Tunguskagyrus не принадлежит к семейству вертячек (Kirejtshuk, Prokin, 2018). Отвергается синонимизация Triaplidae и Coleocatiniidae (Kirejtshuk, Prokin, 2018). Жук, рассматривавшийся как один из триаплид, описывается как Permocatinus gen. nov., P. tomiensis sp. nov. в семействе Coleocatiniidae Ponomarenko et Prokin, 2015. Многочисленность пермских триаплид подтверждает представления о том, что новые группы появляются не после и в результате пермо-триасового кризиса, а до него. На горном плато сибирских траппов нет падения разнообразия ни растений (Садовников, 2016), ни животных, и именно горные сооружения были наиболее вероятными рефугиями сохранения биоразнообразия во время кризиса.
ВВЕДЕНИЕ
Семейство Triaplidae было предложено (Пономаренко, 1977) для новых жуков из верхнего триаса Мадыгена (Средняя Азия). Название отражало определенное сходство новых жуков с жуками семейства Haliplidae, которых традиционно относят к адефагам, хотя они имеют и целый ряд признаков, характерных для архостемат, особенно в строении крыльев (Пономаренко, 1977). У Haliplidae и Triaplidae задние тазики имеют необычно длинные медиальные отростки и большие бедренные покрышки. Неожиданно оказалось, что многие древнейшие адефаги, карабоиды и дитискоиды, также имели большие бедренные покрышки. Появилась возможность по-новому представить взаимоотношение адефаг и архостемат, что и было сделано в указанной выше публикации. В момент выделения семейства Triaplidae высшие подотряды жуков, Polyphaga и Adephaga, были известны только с позднего триаса, и предполагалось, что они возникли в триасе (Пономаренко, 1969). К настоящему времени стало известно, что это предположение было ошибочным, и высшие подотряды уже существовали в терминальной перми. В терминальной перми (Кузнецкий бассейн, Бабий Камень; мальцевская свита, кедровские слои) были найдены и Haliplidae (Ponomarenko, Prokin, 2015), к сожалению, еще не описанные.
Из этого же местонахождения был описан как новый вид Triaplus sibiricus Volkov, 2013, довольно крупный жук посредственной сохранности с большими бедренными покрышками (Волков, 2013). Затем в том же местонахождении был найден остаток жука лучшей сохранности, он был сходного внешнего вида и примерно того же размера. Он рассматривался как принадлежавший к тому же виду, и в дальнейшем обсуждение строения и систематического положения T. sibiricus в основном основывалось не на голотипе, а на этом экземпляре. Так, утверждалось, что жуки этого вида не имели больших бедренных покрышек задних тазиков (Kirejtshuk, Prokin, 2018), тогда как на обратном отпечатке голотипа бедренные покрышки видны. При электронно-микроскопическом изучении голотипа на его надкрылье были обнаружены маленькие (длиной около 0.1 мм) черточки, расположенные по большей части неправильными поперечными рядами. Предполагается, что это волоски, от которых сохранилась лишь часть. Такие же структуры могут быть видны на электронных изображениях нескольких изолированных надкрылий из того же местонахождения. Кажется, что они расположены на той же поверхности, что и фрамбоиды пирита. Сохранение фрамбоидов более вероятно под надкрыльем, возможно, что и волоски были расположены на его нижней поверхности. В местонахождении было найдено несколько крупных (до 1 см) изолированных надкрылий с гладкой верхней поверхностью и продольным вдавлением на боковом крае (схиза). Поскольку крупные жуки в местонахождении очень редки, эти надкрылья рассматривались как принадлежащие жукам того же вида. Изучение голотипа и дополнительного экземпляра показало существенные различия между ними и отличия от типового вида рода Triaplus Ponomarenko, 1977. Об этом свидетельствуют большие поперечные передние тазики, разделенные у голотипа T. sibiricus переднегрудным отростком, доходящим до их вершины. У дополнительного экземпляра контакт передних тазиков виден плохо, и нельзя решить, разделены тазики переднегрудным отростком или нет. У него задние тазики оказались много короче, чем у голотипа. На голотипе можно видеть многочисленные фрамбоиды пирита, тогда как на втором экземпляре их нет. Эти два остатка отличаются по размерам, ширине лба, форме переднеспинки и метэпистерна, относительной длине последнего стернита брюшка, и должны быть описаны как представители разных видов, хотя частично эти отличия могут зависeть от условий захоронения. Голова и переднеспинка голотипа направлены прогнатно; у второго экземпляра они повернуты вбок на правую сторону и голова немного отогнута вниз, с чем частично могут быть связаны различия в форме этих частей тела. Оба экземпляра имеют на среднегрудке большие округлые выемки для вкладывания передних тазиков; на отпечатках между ними и тазиками значительное расстояние, так что прижизненное положение переднегруди и головы должно было быть наклонным, или даже ортогнатным. На новом экземпляре бедренные покрышки видны лишь фрагментарно, они часто вообще не видны, если налегают на брюшко, и об их присутствии можно судить только по разной пунктировке базальных стернитов брюшка с правой и левой стороны.
Поэтому ниже эти жуки описываются как представители разных видов, и для T. sibiricus Volkov, 2013 описывается особый род, который, тем не менее, можно оставить в семействе Triaplidae, поскольку различия между T. sibiricus и остальными Triaplidae не больше, чем между разными родами семейства Schizophoridae. Для второго экземпляра (ПИН, № 4887/348) описывается новый род в семействе Coleocatiniidae Ponomarenko et Prokin, 2015 (=Catiniidae Ponomarenko, 1968).
Наконец, недавно еще одна экспедиция на местонахождении Бабий Камень (начальник А.В. Храмов) собрала значительную коллекцию, почти вдвое увеличив число жуков. Среди них оказалось два экземпляра, возможно, принадлежащих жукам того же рода. Они много меньше по размерам, чем голотип T. sibiricus, так что не могут принадлежать к тому же виду. К сожалению, оба жука неполной сохранности. Присутствует только один признак, сближающий этих жуков с Tomiaplus. Это характерные короткие черточки (или волоски) на надкрыльях. У одного (экз. ПИН, № 4887/704) видны длинные срединные отростки задних тазиков. Большие поперечные передние тазики, разделенные отростком, сближают его с голотипом T. sibiricus. Поскольку второй экземпляр много меньше, ниже он описывается как второй вид рода Tomiaplus. Второй экземпляр (ПИН, № 4887/626) еще меньше, и у него не видны достаточно отчетливо структуры нижней стороны. У него отличное от всех остальных триаплид строение переднегруди, которая не вырезана спереди. В настоящее время предпочтительнее не рассматривать этот вид среди триаплид.
Похожие жуки найдены в местонахождении Анакит-3 в Тунгусском бассейне (Средняя Сибирь, Красноярский край; верхняя пермь, таймырский ярус, лебедевский горизонт). Они также имеют длинные медиальные отростки задних тазиков, длина которых лишь немного уступает ширине тазика. Они были описаны как новые род и вид Tunguskagyrus planus Yan, Beutel et Lawrence, 2018 и отнесены к вертячкам (Yan et al., 2018). Еще до выхода этой публикации они были переизучены, объединены в один род с Triaplus sibiricus Volkov, 2013 и помещены в семейство Triaplidae, которое было переведено в схизофороидные архостематы в (Kirejtshuk, Prokin, 2018). При этом интерпретация строения была радикально изменена; признаки, по которым данный жук был отнесен к вертячкам, были сочтены артефактом: глаза были описаны как простые, а не двойные, строение базальных члеников антенн, как не соответствующее таковому вертячек, брюшко описано как имеющее только пять видимых стернитов, ноги как ходильные (Kirejtshuk, Prokin, 2018). Предложенная интерпретация строения мало отличалась от той, что получилась у меня в результате изучения этого же материала. Есть, однако, и важные отличия.
В обеих публикациях голова голотипа считается прогнатной, однако с самого начала я считал, что она ортогнатная. На это указывает то, что голова поперечная, и на образце, по моему мнению, не видно структур ни верхней, ни нижней стороны головы. Ни в одной публикации не отражено то, что на голове есть несколько фрамбоидов пирита, весьма похожих на фасеточные глаза. В последней публикации утверждается, что у голотипов Triaplus sibiricus и Tunguskagyrus planus нет больших бедренных покрышек, однако на первом из них они видны достаточно отчетливо, почему он и был помещен в род Triaplus. Задние тазики описаны как “сильно поперечные” (Kirejtshuk, Prokin, 2018, с. 879), но на рис. 5 в той же работе их длина составляет четыре пятых ширины, тогда как у рода Hadeocoleodes лишь менее двух пятых. Это отличие не позволяет относить Tunguskagyrus к схизофороидам и подтверждает справедливость сближения Triaplus sibiricus и Tunguskagyrus planus с Triaplus.
Наконец, недавно А.С. Башкуевым среди немаркированного материала из местонахождения Анакит, без указания конкретной точки сбора, был обнаружен еще один остаток жука, похожий на голотип Tunguskagyrus planus, но существенно большего размера и отличающийся пропорциями тела (экз. ПИН, № 2362/22). Этот остаток имеет лучшую сохранность, и на нем ноги сохранились как объемные структуры. При этом границы склеритов видны плохо, и на фотографиях, сделанных при косом освещении, лучше видна тень от склерита, чем его граница. На изображениях, сделанных при большом увеличении на СЭМ видно, что граница размывается из-за того, что остаток состоит из пеплового материала, частицы которого уплощены в чешуйки, слегка смещенные друг относительно друга. Из-за этого не удается получить ни качественных фотографий, ни прорисовок. Даже границы стернитов брюшка, обычно видимые хорошо, на отпечатке точно установить невозможно. Наравне с ними видны и другие поперечные структуры, вероятно, границы тергитов. В зависимости от характера освещения образца лучше видны то одни, то другие наборы поперечных структур, почему число и пропорции склеритов трудно установить. Те же трудности встречаются и при определении числа и формы члеников лапок. Под надкрыльями видны и продольные структуры, по-видимому, жилкование сложенных крыльев, но его строение восстановить не удалось. Можно видеть только овальные структуры, очень похожие на oblongum Hydradephaga. Поскольку они видны на одном и том же месте под обоими надкрыльями и хуже, но все же видны на голотипе; гипотеза, что это все же oblongum, представляется весьма вероятной. На некоторых электронных изображениях, возможно, видны структуры, похожие на длинные щетинкоподобные образования на задних голенях. Возможно, это остатки плавательных волосков.
Авторы рода Tunguskagyrus опубликовали статью (Beutel et al., 2019), содержащую некоторые новые результаты переизучения голотипа T. planus. Отвергается большая часть соображений А.Г. Кирейчука и А.А. Прокина (Kirejtshuk, Prokin, 2018), причем главным недостатком их работы признается то, что не были использованы кладистические методы. Не могу с этим согласиться. За 60 лет занятий филогенией жуков я не могу припомнить ни одного случая, когда кладистическими методами был бы получен результат, существенно превосходящий результаты, полученные традиционными методами. Обвинение филогенетиков, работающих традиционными методами, в том, что их гипотезы не могут быть фальсифицируемы, несправедливо. Единственный метод фальсификации филогенетических древес – это накопление палеонтологических фактов, и он применим к филогенетическим гипотезам, полученным любым способом. Приходится еще раз остановиться на характере глаз – цельные они или разделенные, как по-прежнему утверждают авторы; это мне, несмотря на тщательные поиски, найти не удалось. Зато ноги действительно присутствуют, но сохранились они настолько плохо, что их можно изобразить только предположительно.
Описываемые остатки интересны не только сами по себе как дающие важный материал для понимания эволюции жуков на одном из базальных этапов, но и как значимые свидетельства характера прохождения пермо-триасового экологического кризиса. Оба местонахождения, где они найдены, находятся во временной близости к границе палеозоя и мезозоя, и образовывались во время пермо-триасового события. Ныне большинство исследователей считает, что причиной этого события было колоссальное извержение, давшее вулканогенную толщу Тунгусских траппов. Климатические и ландшафтные последствия этого извержения преобразовали всю экосистему Земли, однако детальное изучение межтрапповых отложений, проведенное, главным образом, Г.Н. Садовниковым, показало, что на самом базальтовом плато картина вовсе не была апокалиптической и существовали достаточно нормальные для этого времени фитоценозы. Многочисленность и разнообразие остатков насекомых также подтверждает выводы Садовникова. Местонахождения Тунгуски и Кузбасса, возможно, образовались в результате единого гигантского вулканического события (Svetlitskaya, Nevolko, 2016): Анакит – в конце туфогенно-осадочного этапа строительства плато, а Бабий Камень, в разрезе которого также присутствуют дайки и покровы базальтов, возник несколько позже, перед массовым излиянием базальтов, когда они распространились далеко от центра извержения. Возможно, что дайки и покровы базальтов Кузнецкого бассейна – это тоже часть базальтов Сибирских траппов и они синхронны. Считается, что высота вулканогенного плато была более 1.5 км, и на этом горном сооружении существовали условия, приемлемые для достаточно обильной жизни и во время термического максимума в середине индского времени (Sun et al., 2012). М.П. Арефьев в важной работе о распределении изотопов углерода и кислорода в карбонатах Русской платформы в конце перми – начале триаса выразил сомнение о связи вымирания с излиянием базальтов, поскольку близ границы имело место не потепление, а значительное похолодание (Арефьев и др. 2015), однако следует заметить, что в начале события была стадия взрывного вулканизма с выбросом в атмосферу огромного количества пепла и соединений серы, что не могло не вызвать заметного похолодания. Представляется, что образование вулканогенных отложений Тунгусского бассейна было однократным событием и заняло ничтожное в геологическом смысле время. Само по себе однократное извержение такой чудовищной мощности вне вулканических областей требует специального объяснения. Оно было предложено в виде воздействия ударной волны в земном ядре после падения крупного астероида, способного пробить земную кору. Подходящие астроблемы известны в южном полушарии. Находки метеоритного вещества известны в верхневятских отложениях Русской платформы (Арефьев, Цельмович, 2017). Более подробно эти соображения изложены в специальной статье (Пономаренко, 2017), хотя новые факты и концепции появляются так быстро, что она уже несколько устарела.
МАТЕРИАЛ И МЕТОДЫ
Изученный материал хранится в коллекции лаборатории артропод Палеонтологического ин-та им. А.А. Борисяка РАН, Москва (ПИН).
Были использованы сканирующий электронный микроскоп Tescan Vega XMU и световой стереоскопический микроскоп Leica M165C с цифровой камерой Leica DFC420 (ПИН РАН).
Рисунки по фотографиям и фототаблицы выполнены в программе Corel Draw. Условные обозначения: сплошные линии – границы склеритов и частей тела, волоски; пунктирные линии – неясные границы склеритов, частей тела и борозд, места повреждения отпечатка, структуры, перекрывающие друг друга.
Работа выполнена при финансовой поддержке Программы Президиума РАН I.22П “Эволюция органического мира и планетарных процессов”. Автор благодарит Р.А. Ракитова за электронно-микроскопическое исследование остатков, Е.В Яна и А.А. Прокина за помощь в подготовке публикации.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
СЕМЕЙСТВО TRIAPLIDAE PONOMARENKO, 1977
Род Tomiaplus Ponomarenko, gen. nov.
Название рода – от р. Томи (им. пад.: р. Томь) и рода Triaplus.
Типовой вид – Triaplus sibiricus Volkov, 2013.
Диагноз. Голова слабо поперечная, почти прогнатная. Передние тазики большие, поперечные, косые, разделены переднегрудным отрезком. Щиток округло-треугольный. Среднегрудка довольно длинная, спереди с парными округлыми вырезками для вхождения вершин передних тазиков. Средние тазики большие, округлые. Метэпистерн сильно расширен вперед. Заднегрудка поперечная, округло сужена вперед. Длина медиального отростка заднего тазика почти равна его ширине на переднем крае. Голени и бедра с короткими редкими волосками, плавательные волоски отсутствуют. Надкрылья гладкие, без точечных борозд; при большом увеличении на них видны короткие черточки, по-видимому, это маленькие волоски. На некоторых надкрыльях видны кольцевые структуры, которые обычно интерпретируются как отверстия колумелл.
Видовой состав. Два–три вида из местонахождения Бабий Камень, терминальная пермь Кузбасса. Помимо двух описанных ниже видов, в местонахождении присутствуют более крупные надкрылья с характерными черточками.
Сравнение. От других родов семейства отличается почти прогнатной головой и разделенными отростком переднегруди тазиками.
Tomiaplus sibiricus (Volkov, 2013)
Triaplus sibiricus: Волков, 2013, с. 74, рис. 1, 2.
Рис. 1.
Tomiaplus sibiricus (Volkov, 2013), голотип ПИH, № 4887/72: а – обратный, б – прямой отпечатки; в, г – рисунок, вид сверху и снизу; д – надкрылье с характерными структурами: насечки или волоски и fr – фрамбоиды пирита; Бабий Камень; верхняя пермь, вятский ярус. Масштабная линейка – 1 мм.

Рис. 2.
Tomiaplus minimus sр. nov., голотип ПИH, № 4887/704: а, б – сверху, в – снизу; Бабий Камень; верхняя пермь, вятский ярус. Масштабная линейка – 1 мм.
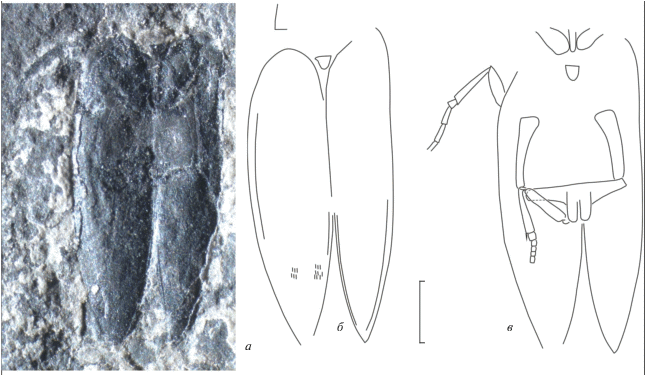
Гoлoтип – ПИH, № 4887/72, неполный прямой и обратный деформированные отпечатки тела жука с головой и переднеспинкой, без антенн и конечностей, стороны остатка асимметричны; Кемеровская обл., Новокузнецкий р-н, правый берег р. Томь в 10 км ниже пос. Усть-Нарык, местонахождение Бабий Камень; мальцевская свита, кедровские слои.
Описание (рис. 1). Голова почти треугольная, ее длина примерно в 1.5 раза меньше ширины. Глаза большие, овальные, боковые. Переднеспинка сильно поперечная, ширина переднеспинки втрое больше длины, спереди вырезанная, передние углы слабо выступают вперед. Переднегрудь короче головы, длина переднегрудки не больше длины передних тазиков. Проплевры у заднего края вдвое шире, чем у переднего. Передние тазики большие, поперечные, косые, вдвое больше в ширину, чем в длину; разделены переднегрудным отрезком, почти совсем не сужающимся назад. Щиток округло-треугольный. Среднегрудка довольно длинная. Средние тазики большие, округлые, расставленные. Метэпистерн сильно расширен вперед. Заднегрудка поперечная, округло сужена вперед (ее длина в 2.4 раза меньше ширины), несет отросток между средними тазиками, на заднем крае с поперечным швом. Задние тазики немного сужены вбок, на заднем крае вырезаны. Бедренные покрышки меньше в длину, чем в ширину. Медиальный отросток заднего тазика лишь немого короче его ширины. Средние бедра расширены, примерно такой же длины, как задние, которые не расширенные. Задняя голень такой же длины, как заднее бедро, но немного уже.
Размеры в мм. Длина 7.1, ширина 3.4, длина надкрылья 6.4.
Сравнение. От T. minimus sр. nov. отличается более короткой заднегрудкой, более длинными медиальными отростками задних тазиков и большими размерами.
Материал. Голотип.
Tomiaplus minimus Ponomarenko, sр. nov.
Название вида minimus лат. – меньший.
Гoлoтип – ПИH, № 4887/704, неполный прямой и обратный деформированные отпечатки тела жука без головы и большей части переднеспинки, без антенн и передней конечности; из-за захоронения в наклонном положении стороны остатка асимметричны, правое надкрылье на четверть шире левого; Кемеровская обл., Новокузнецкий р-н, правый берег р. Томь в 10 км ниже пос. Усть-Нарык, местонахождение Бабий Камень; мальцевская свита, кедровские слои.
Описание (рис. 2). Передние тазики большие, поперечные, косые, разделены переднегрудным отрезком, почти совсем не сужающимся назад. Щиток продольный, округло-треугольный. Среднегрудка длинная, лишь немного короче заднегрудки. Средние тазики округлые, расставленные. Метэпистерн не сильно расширен вперед. Заднегрудка поперечная, несильно округло сужена вперед; ее длина на одну пятую меньше ширины. Метэпистерн втрое шире у переднего края, чем у заднего. Задние тазики немного сужены вбок, их задние отростки немного короче ширины тазика. Бедренные покрышки больше в длину, чем в ширину. Средние бедра расширены, примерно такой же длины, как линейные голени; первый членик средней лапки втрое короче второго. Задняя голень такой же длины, как заднее бедро, расширена к вершине; первый членик задней лапки втрое короче второго.
Размеры в мм. Длина надкрылья 3.97.
Сравнение. От типового вида отличается более длинной заднегрудкой, менее длинными медиальными отростками задних тазиков и меньшими размерами.
Материал. Голотип.
Род Tunguskagyrus Yan, Beutel, Lawrence, 2018
Tunguskagyrus: Yan et al., 2018, с. 2.
Типовой вид – Tunguskagyrus planus Yan et al., 2018, по первоначальной мононимии; Красноярский край, Тунгусский бассейн; верхняя пермь, таймырский ярус, лебедевский горизонт.
Диагноз. Средних размеров жуки с вальковатым телом, ортогнатной головой, отогнутой вниз переднегрудью, надкрыльями без следов жилкования, довольно длинными ногами. Переднеспинка сильно поперечная, спереди вырезанная, передние углы выступают вперед. Щиток округло-треугольный. Переднегрудь короче головы. Передние тазики большие, входят в округлые ямки на среднегруди. Среднегрудка довольно длинная, спереди с парными округлыми вырезками для вхождения вершин передних тазиков. Средние тазики большие, округлые. Метэпистерн расширен вперед. Заднегрудка поперечная, округло сужена вперед, несет отросток с округлым передним краем между средними тазиками, на заднем крае с поперечным швом. Задние тазики немного сужены вбок, на заднем крае вырезаны. Длина медиального отростка заднего тазика не меньше его ширины на переднем крае. Бедренные покрышки меньше в длину, чем в ширину. Ноги довольно длинные. Голени и бедра почти равной длины, с короткими редкими волосками. Надкрылья гладкие, без точечных борозд, при большом увеличении на них видны короткие черточки, по-видимому, это маленькие волоски. На некоторых надкрыльях видны кольцевые структуры, которые обычно интерпретируются как отверстия колумелл.
Видовой состав. Два вида из местонахождения Анакит-3.
Сравнение. От других родов семейства отличается широкими и короткими соприкасающимися передними тазиками.
Tunguskagyrus planus Yan et al., 2018
Tunguskagyrus planus: Yan et al., 2018, с. 2, рис. 1–3.
Рис. 3.
Tunguskagyrus planus Yan et al., 2018, голотип ПИH, № 4887/72: а – фото; б, г, д – электронное изображение, объект на переднегруди – раковина остракоды; в – рисунок, вид снизу; Анакит-3; верхняя пермь, вятский ярус. Масштабная линейка – 1 мм.

Гoлoтип – ПИН, № 3381/32, прямой и обратный отпечатки жука без антенн и большей части ног, переднегрудь и голова сдвинута вперед и повернута налево, так что переднегрудь выглядит асимметричной; Красноярский край, Тунгусский бассейн, Анакит-3; верхняя пермь, таймырский ярус, лебедевский горизонт.
Описание (рис. 3). Овальный жук, сильнее суженный назад. Длина жука вдвое больше ширины. Переднеспинка в длину равна толщине головы, сильно поперечная, спереди вырезанная, передние углы выступают вперед. Передние тазики большие, поперечные, косые, их длина примерно вдвое меньше ширины. Среднегрудка без крестообразного шва. Средние тазики большие, округлые. Метэпистерн на переднем крае в 5 раз шире, чем на заднем. Длина заднегрудки между тазиками втрое меньше ширины на заднем крае, ширина переднего края составляет ¾ заднего. Передний отросток между средними тазиками почти треугольный, поперечный шов не доходит до границ заднегрудки. Задние тазики сильно сужены вбок, длина медиального отростка примерно равна ширине тазика на переднем крае. Бедренные покрышки в длину примерно равны медиальному отростку. Последний вентрит брюшка вдвое длиннее предпоследнего. Задние ноги, вытянутые назад, не доходят до вершины брюшка. Задние бедра длинные, выходят за границы тела, голени короче бедер, первый и пятый членики лапок длиннее почти равных остальных. Надкрылья тонкие, через них видны основания жилок крыла, в том числе и oblongum.
Размеры в мм. Длина 9.48, ширина 4.66, длина надкрылья 7.1.
Сравнение. Вид отличается меньшим размером, более широким телом, сильнее расширенными вперед метэпистернами, более короткими ногами.
Материал. Голотип.
Tunguskagyrus уani Ponomarenko, sp. nov.
Вид назван в честь палеоэнтомолога Е. Яна.
Гoлoтип – ПИН, № 2362/22, прямой и обратный отпечатки жука без антенн и части ног, голова и переднегрудь подогнуты; Красноярский край, Тунгусский бассейн, Анакит; верхняя пермь, таймырский ярус, лебедевский горизонт.
Описание (рис. 4). Овальный жук, сильнее суженный назад. Длина жука в 2.5 раза больше ширины. Переднеспинка сильно поперечная, спереди вырезанная, передние углы выступают вперед. Передние тазики большие, длинные. Среднегрудка с крестообразным швом. Средние тазики большие, округлые. Метэпистерн на переднем крае вдвое шире, чем на заднем. Длина заднегрудки между тазиками в 2.2 раза меньше ширины на заднем крае; ширина переднего края заднегрудки вдвое меньше, чем заднего. Передний отросток между средними тазиками почти треугольный, поперечный шов доходит до границ заднегрудки. Задние тазики сильно сужены вбок, длина медиального отростка почти в 1.4 раза меньше ширины тазика на переднем крае. Бедренные покрышки в длину больше медиального отростка. Последний вентрит брюшка длиннее предпоследнего. Задние ноги, вытянутые назад, заходят за вершину брюшка. Задние бедра длинные, доходят до границы тела, голени длиннее бедер, первый и пятый членики лапок длиннее почти равных остальных. Надкрылья тонкие, через них видны основания жилок крыла, в том числе и oblongum.
Рис. 4.
Tunguskagyrus уani sp. nov., голотип ПИH, № 2362/22: а – фото; б, г, д – электронное изображение; в – рисунок, вид снизу; Анакит-3; верхняя пермь, вятский ярус. Масштабная линейка – 1 мм.

Рис. 5.
Permocatinus tomiensis sр. nov., голотип ПИН, №3446/348: а–в, е – электронное изображение; г, д – рисунки, вид снизу; е – средняя нога; Анакит-3; верхняя пермь, вятский ярус. Масштабная линейка – 1 мм.

Размеры в мм. Длина 12.5–13.3, ширина 5.0–5.26, длина надкрылья 9.6–10.7.
Сравнение. Вид отличается от типового большим размером, более вытянутым телом, слабее расширенными вперед метэпистернами, более длинными ногами.
Материал. Голотип и, возможно, экз. ПИН, №3381/110, Анакит-3, с повернутой вперед головой.
СЕМЕЙСТВО COLEOCATINIIDAE PONOMARENKO ET PROKIN, 2015 (=CATINIIDAE PONOMARENKO, 1968)
Род Permocatinus Ponomarenko, gen. nov.
Название рода – от пермского периода и рода Catinus; м.р.
Типовой вид – Permocatinus tomiensis sр. nov.
Диагноз. Средних размеров жуки с овальным уплощенным телом, прогнатной головой, слегка отогнутой вниз переднегрудью, довольно длинными ногами. Голова поперечная, слабо сужена вперед. Глаза небольшие, боковые. Переднеспинка сильно поперечная, округло сужена вперед, спереди вырезанная, передние углы выступают вперед. Щиток поперечно-треугольный. Переднегрудь много длиннее головы, округло сужена вперед. Передние тазики большие, поперечные. Среднегрудка довольно короткая, спереди с парными округлыми вырезками для вхождения вершин передних тазиков, с крестообразным швом. Средние тазики большие, округлые, расставленные. Метэпистерн сильно расширен вперед. Заднегрудка поперечная, округло сужена вперед, несет отросток с округлым передним краем между средними тазиками, на заднем крае с прерванным в боковой части поперечным швом. Задние тазики немного сужены вбок, на заднем крае вырезаны. Медиальный отросток заднего тазика широкий и короткий. Голени и бедра с короткими редкими волосками, плавательные волоски отсутствуют. Брюшко с пятью вентритами, последний вентрит узкий и длинный.
Видовой состав. Род монотипический.
Сравнение. От других родов семейства отличается каплевидной формой тела, округло суженной вперед переднеспинкой и большими поперечными передними тазиками.
Permocatinus tomiensis Ponomarenko, sр. nov.
Название вида от р. Томи.
Гoлoтип – ПИН, № 4887/348, прямой и обратный отпечатки жука без надкрылий и большей части ног; Кемеровская обл., Новокузнецкий р-н, правый берег р. Томь в 10 км ниже пос. Усть-Нарык, местонахождение Бабий Камень; мальцевская свита, кедровские слои.
Описание (рис. 5). Средних размеров жуки с овальным уплощенным телом, прогнатной головой, слегка отогнутой вниз переднегрудью, довольно длинными ногами. Голова поперечная, слабо сужена вперед. Глаза небольшие, боковые. Переднеспинка поперечная, сильно округло сужена вперед, спереди вырезанная, передние углы выступают вперед. Щиток поперечно-треугольный. Переднегрудь много длиннее головы, округло сужена вперед. Передние тазики большие, поперечные. Среднегрудка довольно короткая, спереди с парными округлыми вырезками для вхождения вершин передних тазиков, с крестообразным швом. Средние тазики большие, округлые, расставленные. Метэпистерн сильно расширен вперед. Заднегрудка поперечная, округло сужена вперед, несет отросток с округлым передним краем между средними тазиками, на заднем крае с прерванным в боковой части поперечным швом. Задние тазики немного сужены вбок, на заднем крае вырезаны. Медиальный отросток заднего тазика широкий и короткий. Голени и бедра с короткими редкими волосками. Брюшко с пятью вентритами, последний вентрит узкий и длинный.
Размеры в мм. Длина 7.1, ширина 3.4, длина надкрылья 6.4.
Материал. Голотип.
ОБСУЖДЕНИЕ
В современной систематике сосуществуют два достаточно далеких по идеологии подхода. Мейнстримом является кладизм – позитивистский номинализм. Набирается как можно больше признаков, которыми заряжается машина, определяющая, кто с кем более сходен. Приспособительный и корреляционный анализы, как правило, не используются. Достоинство этого подхода – его формализованность и однозначность результатов. Но однозначность в рамках компьютерной программы еще не означает истинность полученных результатов, их соответствие реальному эволюционному процессу. Тем не менее, кладизм притязает на истину в последней инстанции. С помощью кладистического анализа было установлено, что вертячки должны быть противопоставлены остальным Adephaga. При этом более раннее представление о том, что первыми отщепились Haliplidae (Пономаренко, 1977), просто не было рассмотрено и дезавуировано. Оно было основано на сходстве Haliplidae со схизофороидными архостематами в строении задних крыльев и специфическом строении задних тазиков и основания брюшка. В ископаемом состоянии далеко не всегда можно видеть задние тазики с большими бедренными покрышками. Они хорошо видны на голотипе Triaplus macroplatus, поскольку у него оторвано брюшко. Но о паратипе этого же вида утверждалось, что он не имеет бедренных покрышек и не может считаться принадлежащим к триаплидам (Kirejtshuk, Prokin, 2018). В действительности бедренные покрышки у него присутствуют, но, по-видимому, при захоронении они были отогнуты и стояли торчком. Они видны в виде узкой полоски на рис. 10 (с. 67) в работе Кирейчука и Прокина, поскольку при захоронении объемных объектов мы обычно видим не реальную форму склеритов, а ее проекцию на плоскость захоронения. Конспецифичность голотипа и паратипа T. macroplatus доказывается наличием у них длинных медиальных отростков задних тазиков.
Подробное изучение голотипа Tunguskagyrus planus Yan et al., 2018 не дало мне никаких признаков его принадлежности к вертячкам. Голова действительно ортогнатная, чего не бывает у вертячек. Глаз мне удалось найти только два, и они боковые и не того строения, как у Spanglerogyrus. Внутри головы смутно просматриваются контуры большой мандибулы, не похожей на мандибулы вертячек.
Бедренные покрышки задних тазиков часто встречаются и у Adephaga, и у карабоидов, и у дитискоидов, и у Polyphaga (Mesocinetidae), от терминальной перми до поздней юры. Позднее, в том числе и у современных жуков, они встречаются много реже. Они известны и у водных жуков, и у жуков с наземным обликом. У коптоклавид и эвдромеин можно видеть, что триасовые и юрские формы почти сплошь имеют увеличенные задние тазики, в мелу преобладают жуки с умеренными покрышками или вовсе без них. Удлинение медиальных отростков задних тазиков и выдвижение из-под них первого видимого (второго истинного) брюшного стернита рассматривалось как приспособление для увеличения проведения вперед бедра с увеличением амплитуды гребка (Пономаренко, 1977). Маловероятно, что возникновение больших бедренных покрышек связано с их использованием для хранения воздуха, поскольку они присутствуют и у наземных форм.
Триаплиды рассматривались как возможный предок галиплид, связывающий последних со схизофороидными архостематами. Однако ныне древнейшая галиплида найдена в том же местонахождении Бабий Камень, что и рассматриваемые жуки, так что они все должны рассматриваться как разные линии процесса адаптации жуков к водному образу жизни. Из-за значительной разницы в размерах триаплиды и галиплиды должны были реализовать разные экологические ниши.
Увеличение разнообразия жуков с длинными задними тазиками и большими бедренными покрышками не облегчило трудности их классификации, скорее наоборот, эти трудности увеличились. Не только триаплиды и галиплиды не принадлежат к единой филогенетической ветви, но и триаплиды с новоописанными жуками также лишь параллельные формы. По-видимому, в позднепермское время, возможно лишь в вятское, полифаги и адефаги происходят не двумя стволами, а целым “эволюционным газоном”, представляющим собой архаическое разнообразие. Конечно, не только эволюция задних тазиков и основания брюшка эволюирует таким способом. Подобным образом формирование гладких надкрылий и надкрылий с точечными бороздками происходит в многочисленных параллельных стволах.
Точное систематическое положение жуков, описанных выше как Tomiaplus, не может быть определено. В палеонтологической практике имеется тенденция помещать формы, сочетающие признаки архаичных и продвинутых групп, в последние. Следуя ей, Triaplus можно помещать среди адефаг, так же как галиплид и триаплид. Можно их рассматривать как архостемат, но в этом случае туда же следует перемещать и галиплид, что приведет к значительному изменению таксономических стандартов в адефагах.
В то же время, предположения А.Г. Пономаренко (1977) о водном образе жизни Triaplus не лишены оснований, так как вытянутые вдоль средней линии тела медиальные отростки задних тазиков и крупные бедренные покрышки, действительно, могли обеспечивать продвижение вперед задних ног для более эффективного гребка, подобно таковому у Haliplidae.
Многочисленность пермских триаплид подтверждает представления о том, что новые группы появляются не после и в результате пермо-триасового кризиса, а до него. На горном плато сибирских траппов нет падения разнообразия ни растений (Садовников, 2016), ни животных, и именно горные сооружения были наиболее вероятными рефугиями сохранения биоразнообразия во время кризиса.
Резюмируем сказанное: Триаплиды представляют собой особую ветвь жуков, специфическим признаком которых являются длинные медиальные отростки задних тазиков, и слияние их со схизофоридами нецелесообразно. Требуется изучение основания брюшка на лучше сохранившемся материале для решения вопроса о том, принадлежат ли триаплиды к архостематам или к адефагам.
Список литературы
Арефьев М.П., Кулешов В.Н., Покровский Б.Г. Изотопный состав углерода и кислорода в континентальных карбонатах поздней перми и раннего триаса Русской плиты: глобальный экологический кризис на фоне меняющегося климата // Докл. Акад. наук. 2015. Т. 460. № 2. С. 193–197.
Арефьев М.П., Цельмович В.А. Признаки присутствия импактного материала в вятском ярусе Малой Северной Двины // Палеострат-2017. Годичн. собрание (научн. конф.) секции палеонтологии МОИП и Московского отделения Палеонтол. об-ва при РАН. Тез. докл. М., 2017. С. 7–8.
Волков А.Н. Новый вид семейства Triaplidae из местонахождения Бабий Камень (Кузнецкий бассейн) // Палеонтол. журн. 2013. № 1. С. 72–75.
Пономаренко А.Г. Историческое развитие жесткокрылых-архостемат. М.: Наука, 1969. 239 с. (Тр. Палеонтол. ин-та АН СССР. Т. 125).
Пономаренко А.Г. Состав и экологическая характеристика мезозойских жесткокрылых. Подотряд Adephaga. Инфраотряд Staphyliniformia // Мезозойские жесткокрылые. М.: Наука, 1977. С. 8–119 (Тр. Палеонтол. ин-та АН СССР. Т. 161).
Пономаренко А.Г. Экология наземной биоты во временных окрестностях Р/Т кризиса // Палеонтол. журн. 2017. № 6. С. 53–58.
Садовников Г.Н. Эволюция траппового плато Средней Сибири // Палеонтол. журн. 2016. № 5. С. 87–99.
Beutel R., Yan E., Lawrence J. Phylogenetic methods applied to extinct beetles – the case of Tunguskagyrus (Gyrinidae or Triaplidae) // Palaeoentomology. 2019. V. 2. № 4. https://doi.org/10.11646/palaeoentomology.2.4.11
Kirejtshuk A.G., Prokin A.A. The position of the Paleozoic genus Tunguskagyrus Yan, Beutel et Lawrence in the family Triaplidae sensu n. (Coleoptera, Archostemata: Schizophoroidea) // Entomol. Rev. 2018. V. 98. № 7. P. 872–882.
Ponomarenko A.G., Prokin A.A. Review of paleontological data on the evolution of aquatic beetles (Coleoptera) // Paleontol. J. 2015. V. 49. № 13. P. 1383–1412.
Sun Yadong, Joachimowski M.M., Wignail P.B. et al. Lethally hot temperatures during the Early Triassic Greenhouse // Science. 2012. V. 338. № 6105. P. 366–370. https://doi.org/10.1126/Science.1224126
Svetlitskaya N.V., Nevolko P.A. Late Permian-Early Triassic traps of the Kuznetsk Basin, Russia: Geochemistry and petrogenesis in respect to an extension of the Siberian Large Igneous Province // Gondvana Res. 2016. V. 39. P. 57–76.
Yan E., Beutel R., Lawrence J. Whirling in the late Permian: ancestral Gyrinidae show early radiation of beetles before Permian-Triassic mass extinction // BMC Evol. Biol. 2018. 18:33. P. 1–10. https://doi.org/10.1186/s12862-018-1139-8
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал