

Палеонтологический журнал, 2021, № 6, стр. 3-14
Сравнение кристаллографической текстуры раковин современных, ископаемых и субфоссильных двустворчатых моллюсков
А. В. Пахневич a, b, *, Д. И. Николаев b, Т. А. Лычагина b
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Объединенный институт ядерных исследований, Лаборатория нейтронной физики им. И.М. Франка
Дубна, Россия
* E-mail: alvpb@mail.ru
Поступила в редакцию 12.05.2020
После доработки 10.02.2021
Принята к публикации 20.02.2021
Аннотация
С помощью нейтронной дифракции изучена кристаллографическая текстура раковин современных, субфоссильных и ископаемых (верхний плейстоцен) двустворчатых моллюсков Mytilus galloprovincialis, современных и субфоссильных M. trossulus, а также современных и ископаемых (верхний плейстоцен) Ostrea edulis. Выявлено, что для раковин видов мидий различного возраста характерна высокая упорядоченность кристаллов кальцита – острая кристаллографическая текстура. Кристаллы арагонита распределены слабо упорядоченно, в зависимости от формы створок. В сухом и теплом климате у раковин M. galloprovincialis происходит расслоение раковинного вещества, что нарушает кристаллографическую текстуру. В условиях суши упорядоченность кристаллов кальцита в раковинах M. galloprovincialis падает. У субфоссильных M. trossulus и ископаемых M. galloprovincialis и O. edulis за сотни и тысячи лет изменяется интенсивность кристаллографической текстуры кальцита и арагонита.
ВВЕДЕНИЕ
Для современной палеонтологии остается актуальным вопрос изменения минерального и органического вещества в процессе перехода остатков организмов из биосферы в литосферу. Поскольку минеральное вещество гораздо чаще сохраняется в виде раковин, костей, панцирей, то и возможностей изучать его значительно больше. У двустворчатых моллюсков раковины образуются клетками мантии. Эти клетки формируют микроструктурные элементы раковины – призмы, фибры, пластины (Золотарев, 1989; Попов, 1992). Если в прижизненном состоянии минеральное вещество контролируется клетками (Checa et al., 2016), то после смерти контроль исчезает, и изменения могут произойти в различных направлениях. Возможна перекристаллизация раковинного вещества с изменением направления кристаллов, которая наблюдается визуально. Это было хорошо проиллюстрировано на примере эоценовых фораминифер Танзании и Центральной Пацифики (Pearson et al., 2015). Диагенез вмещающей породы может привести к ее уплотнению, литификации, цементации. Это сопровождается ростом кристаллов внутри породы, образованием конкреций, изменением содержания воды и т.д. Зачастую меняется исходный минеральный состав осадка, его плотность. С увеличением плотности возрастает давление на окаменевшие остатки, что тоже может влиять на кристаллографическую текстуру. В дальнейшем может происходить метаморфизация горной породы, что значительно изменит текстуру как ископаемых остатков, так и ее самой. С окаменелостями в породе происходят различные преобразования, помимо перекристаллизации. Хорошо известны случаи замещения остатков диагенетическими минералами, растворение. Малой посмертной устойчивостью обладают арагонитовые раковины (Жирков, 2017). Часто после смерти животных они растворяются. При замещении диагенетическими минералами часто сохраняется форма скелета, при этом замещаются минералы с одной кристаллической решеткой минералами с другой, например, тригональный кальцит – кубическим пиритом. Иногда химический состав раковинного вещества остается тот же, но меняется его кристаллическая решетка, например, ромбический арагонит переходит в тригональный кальцит.
Чтобы интерпретировать особенности вымерших организмов, важно знать, всегда ли происходит изменение минерального вещества и, если есть изменения, то как быстро после смерти организма они начинаются. Не исключено, что исходное состояние остается устойчивым и сохраняется в течение миллионов лет. Современные методы исследования позволяют значительно расширить диапазон знаний о строении минерального вещества. Помимо традиционных подходов, с помощью которых изучается морфология объектов (микроструктура с разрешением в микро- и нанометры), томографических методов, существуют возможности исследования объектов на уровне ангстремов. Например, это атомно-силовая микроскопия, дифракция обратно рассеянных электронов (EBSD), текстурный анализ на основе рентгеновской дифракции, текстурный анализ на основе нейтронной дифракции. Если с помощью первых трех методов анализируется небольшой участок на поверхности образца (Frýda et al., 2010; Jacob et al., 2019), то последним методом – весь образец или сразу несколько скрепленных образцов, т.е. небольшая выборка. По результатам анализа ориентации миллионов кристаллов формируются дифракционные минералогические спектры и полюсные фигуры. По показателям остроты текстуры определяется упорядоченность кристаллов. Метод нейтронной дифракции выбран для анализа текстуры (Brokmeier, 1994) раковин современных, субфоссильных и ископаемых двустворчатых моллюсков одних и тех же видов.
МАТЕРИАЛ И МЕТОДИКА
В качестве материала были выбраны крупные створки взрослых двустворчатых моллюсков родов Mytilus и Ostrea, часто встречающихся как в современных, так и в голоценовых и древних плейстоценовых отложениях.
Створки современных и живших 40 лет назад моллюсков Mytilus galloprovincialis Lamarck, 1819 были собраны на берегу Казантипского залива (Азовское море, п-ов Крым) напротив с. Песочное в 2009–2013 и в 2016 гг. А.В. Пахневичем. Современные – собирались на поверхности береговых отложений около уреза воды и на расстоянии до 2 м от него. Створки сорокалетней давности были обнаружены на расстоянии около 500 м от берега под слоем почвы. В 1970-х гг. около с. Песочное находился песчаный карьер (по устным сообщениям местных жителей), засыпанный в целях рекультивации береговыми отложениями, большую часть которых составляют раковины моллюсков. На поверхности отложений были высажены сосны. Информация, полученная от жителей с. Песочное, подтверждается возрастом сосен, который был определен по кольцам роста на свежих спилах деревьев. Таким образом, раковины моллюсков пролежали под почвенным слоем до момента сбора примерно 40 лет (рис. 1, а, б).
Рис. 1.
Исследованный материал: а, б – в засыпанном карьере: а – внешний вид разреза с субфоссильными раковинами моллюсков, мощность 1 м; б – плотные скопления раковин моллюсков, стрелками показаны створки Mytilus galloprovincialis; в – неполные створки субфоссильных M. galloprovincialis с обломанными краями и расслоившейся раковиной; берег Казантипского залива (Азовское море, п-ов Крым) напротив с. Песочное, около 500 м от берега; г – субфоссильная створка M. trossulus; берег бухты Свипер, о. Адак (Алеутские о-ва), древнеалеутское поселение ADK-009, кухонная куча; д – створки современных M. galloprovincialis, подготовленные для исследования с помощью нейтронной дифракции; берег Казантипского залива (Азовское море, п-ов Крым) напротив с. Песочное, от 0 до 2 м от уреза воды; е – ископаемые створки M. galloprovincialis; п-ов Тамань, коса Тузла; карангатские отложения, верхний плейстоцен; ж – створки ископаемых Ostrea edulis; п-ов Тамань, коса Чушка; карангатские отложения, верхний плейстоцен. Размерная линейка 1 см.

Большинство раковин современных и субфоссильных Mytilus trossulus Gould, 1850 происходят с побережья о. Адак (Алеутские о-ва). Современные мидии собраны на литорали узкой каменистой бухты Свипер. На берегу той же бухты было обнаружено и изучено древнеалеутское поселение ADK-009 (Антипушина и др., 2009). Здесь в кухонной куче (midden) найдены раковины субфоссильных M. trossulus. Отложения стоянки формировались с перерывом, слои разделены гумусово-зольными прослойками. Слои с раковинным и костным материалом датированы радиоуглеродным методом в Институте проблем экологии и эволюции им. А.Н. Северцова РАН (ИПЭЭ), в группе исторической экологии. Радиоуглеродные датировки производились по коллагену из костей рыб (Антипушина и др., 2009). Исследованные нами раковины моллюсков отобраны из слоя возрастом около 400 л.н. Материал предоставлен А.Б. Савинецким (ИПЭЭ) и Ж.А. Антипушиной (Государственный биологический музей им. К.А. Тимирязева). Дополнительный материал по M. trossulus из Охотского моря был передан Е.М. Крыловой (Ин-т океанологии им. П.П. Ширшова РАН).
Раковины плейстоценовых моллюсков M. galloprovincialis и Ostrea edulis L., 1758 происходят из карангатских отложений (верхний плейстоцен) п-ова Тамань. Устрицы собраны в отложениях косы Чушки, а мидии – в отложениях косы Тузлы. Они были переданы нам на исследование А.В. Гужовым (лаб. моллюсков, ПИН РАН). Другие образцы O. edulis из карангатских отложений собраны А.В. Пахневичем на побережье Арабатского залива Азовского моря в г. Щелкино (п-ов Крым).
Створки современных O. edulis были собраны Пахневичем на побережье Черного моря, около пос. Малый Утриш, и А.А. Козловой и О.А. Козловой – в береговых выбросах в районе Лагоса (Португалия).
Информация о кристаллографической текстуре − преимущественных ориентировках слагающих поликристалл кристаллитах − может быть получена из полюсных фигур (Bunge, 1982). Полюсная фигура поликристаллического вещества представляет собой проекцию нормалей к данной атомной плоскости, построенную для всех кристаллов исследуемого образца. Если образец изотропен (текстура отсутствует), то выходы нормалей плоскости равномерно покрывают сферу. В текстурированном образце распределение нормалей к плоскости неравномерно и бывает представлено линиями постоянной интенсивности (изолиниями). Если спроектировать такое распределение на экваториальную плоскость сферы, то эта стереографическая проекция и будет полюсной фигурой (рис. 2). Исследования текстуры традиционно проводят с помощью рассеяния рентгеновских лучей или электронов. Однако из-за небольшой глубины проникновения в вещество с помощью этих методов невозможно получить информацию, необходимую для полного описания текстуры. Полные полюсные фигуры могут быть измерены только методом дифракции тепловых нейтронов.
Полюсные фигуры, представленные в данной работе, были измерены в Лаб. нейтронной физики им. И.М. Франка Объединенного ин-та ядерных исследований (г. Дубна, Россия). Использовалась установка СКАТ (спектрометр для количественного анализа текстуры), расположенная на канале 7А-2 импульсного реактора ИБР-2 (Nikolayev et al., 2019). Для анализа применялся метод нейтронной дифракции по времени пролета. С его помощью были получены дифракционные рефлексы, соответствующие минералам, которые входят в состав раковин.
СКАТ состоит из детекторного кольца (диаметр 2 м), на котором расположено 19 детекторно–коллиматорных комплексов под одинаковым углом рассеяния 2θ = 90°. Детекторные модули установлены на окружности кольца таким образом, что шаг на полюсной фигуре имеет величину 5°. Образец вращается с шагом 5° вокруг горизонтальной оси, имеющей угол 45° относительно падающего пучка нейтронов. Вращение осуществляется с помощью гониометра. Таким образом, для каждой полюсной фигуры регистрируется 19 × 72 = 1368 дифракционных спектров. Необходимо отметить, что благодаря времяпролетной методике полюсные фигуры от всех минералов (фаз), имеющихся в образце, измеряются одновременно, т.е. извлекаются из одних и тех же спектров. Сечение пучка нейтронов 50 × 90 мм дает возможность измерять крупные образцы. Наиболее интенсивные обособленные дифракционные рефлексы на каждом из зарегистрированных спектров анализировались с помощью программы Pole Figure Extractor (Nikolayev et al., 2005) для выяснения характера распределения соответствующих кристаллографических плоскостей в створках. Интенсивность одного рефлекса, соответствующего кристаллографической плоскости с определенными индексами Миллера, дает одну точку на полюсной фигуре, обозначенной этими индексами. Все полюсные фигуры были отнормированы и сглажены с одинаковым параметром (Nikolayev, Ullemeyer, 1994). Анализировались наиболее интенсивные дифракционные рефлексы, соответствующие кристаллографическим плоскостям с индексами Миллера (0006) для кальцита и (012)/(121) для арагонита. Чем более упорядоченно располагались кристаллы минералов, тем выше была интенсивность на полюсной фигуре (полюсная плотность), выражающаяся в единицах изотропного распределения (multiple random distribution, mrd – это сокращение используется в англоязычной литературе без расшифровки и не имеет русского аналога). Увеличение интенсивности на полюсной фигуре интерпретируется как усиление текстуры. Значение полюсной плотности, равное единице, означает, что соответствующие кристаллографические плоскости равномерно распределены в образце по всем направлениям.
Для исследования отбирались створки размером не менее 30 мм длиной и массой не менее 30 г. Створки соединялись по три–четыре с помощью специального двухкомпонентного клея. Это делалось для того, чтобы набрать объем материала, достаточный для проведения измерений. Далее подготовленный таким образом образец прикреплялся к стеклянному штифту и закреплялся в гониометре установки (рис. 1, д). Время измерения каждого образца составляло 22 ч.
Рис. 2.
Представление нормалей к плоскостям {100} кубического кристалла на стереографической проекции: а – кристалл в единичной сфере; б – проекция нормалей к плоскостям {100} на плоскость экватора; в – полюсная фигура (100) и определение углов α и β для нормали к плоскости (100). X, Y, Z – обозначения осей.

Использовались или только левые, или только правые створки. Они изучались целиком, а не микрофрагментами на поверхности створок, как это происходит при исследовании с помощью дифракции обратно рассеянных электронов и текстурным анализом на основе рентгеновской дифракции. Таким образом, результаты оказывались более репрезентативными, так как анализировались сотни тысяч и миллионы кристаллов.
Авторы выражают сердечную благодарность всем, кто передал материал на исследование: А.Б. Савинецкому (ИПЭЭ), Ж.А. Антипушиной (Государственный биологический музей им. К.А. Тимирязева), А.В. Гужову (ПИН РАН), Е.М. Крыловой (Ин-т океанологии им. П.П. Шир-шова РАН), А.А. Козловой (Российский национальный исследовательский медицинский университет им. Н.И. Пирогова) и О.А. Козловой (ГБОУ Школа № 1502 “Энергия”).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Условия фоссилизации и сохранность субфоссильных образцов
Современные раковины пролежали на берегу морей не более года или были собраны при жизни моллюска (M. trossulus в бухте о. Адак). Раковины карангатских моллюсков (верхний плейстоцен) были отобраны из песчаных морских отложений (соленость моря – 20–24‰) (Федоров, 1963), т.е., они не захоранивались на берегу. Субфоссильные раковины с побережья Казантипского залива пролежали в условиях суши около 40 лет. При этом на них воздействовали совсем иные абиотические факторы, нежели в морских осадках. Мощность слоев, где залегали раковины, составляет 0.2–1.70 м. Для данного района Керченского п-ова характерны годовые перепады температур от –2 до +25.1°С (в среднем, для самого холодного месяца января и для июля), среднегодовая температура +10…+11°С. Количество осадков небольшое, от 150–200 мм в холодный период (ноябрь–март) до 350–375 мм в теплый летний период (Ведь, 2000; Горбунов и др., 2014; Krivoguz, Bespalova, 2018). Створки M. galloprovincialis почти все с обломанными краями (рис. 1, в). Происходит отслоение перламутрового слоя от верхних слоев, вероятно, из-за разных коэффициентов расширения арагонита и кальцита. При этом органический слой частично сохраняется. На поверхности створок образуются минеральные корки, которые по составу соответствуют карбонату кальция с примесью кремнезема (рис. 3, а, б).
Рис. 3.
Участки раковин современных и субфоссильных двустворчатых моллюсков, изученные с помощью СЭМ: а – створки субфоссильной Chamelea gallina (L., 1758) c минеральной коркой; б – гистограмма элементного состава этой корки; берег Казантипского залива (Азовское море, п-ов Крым) напротив с. Песочное, около 500 м от берега; в – микроструктура современной створки Mytilus galloprovincialis, состоящей из двух слоев карбоната кальция, представленного в верхнем слое кальцитом, а в нижнем арагонитом; берег Казантипского залива (Азовское море, п-ов Крым) напротив с. Песочное, от 0 до 2 м от уреза воды; г – микроструктура современной створки Ostrea edulis, состоящей из двух слоев кальцита; побережье Черного моря, около пос. Малый Утриш.

Иная климатическая ситуация на о. Адак, где мидии пролежали на суше около 400 лет. Раковины M. trossulus располагались в кухонной куче вперемешку с раковинами других моллюсков, усоногих раков, панцирями морских ежей, костями позвоночных животных на глубине около 1 м от поверхности, мощность всего разреза 1.6 м (Антипушина и др., 2009). На о. Адак среднегодовые температуры ниже – +4.7°С, а количество осадков выше: среднегодовой показатель – 1544 мм (Leslie, 1989). Створки, в основном, целые. Органический слой разрушен (рис. 1, г). Перламутр отшелушивается в виде микрофрагментов, образуя белую пудру.
О связи микроструктуры и кристаллографической текстуры
Можно предполагать, что диагенетические изменения ископаемого объекта затронут, прежде всего, микроструктурные элементы раковины, что отразится и на кристаллографической текстуре. Но эта связь не является явной. Ранее с помощью метода рентгеновской дифракции на примере 50 видов современных моллюсков из классов Monoplacophora, Bivalvia, Gastropoda и Cephalopoda было показано (Chateigner et al., 2000), что одному из девяти упомянутых в цитируемой статье типов микроструктуры могут соответствовать несколько типов кристаллографической текстуры, так как кристаллы в одних и тех же микроструктурных элементах, например, в призмах, могут быть ориентированы по-разному. Поскольку не существует прямой строгой связи между типом микроструктуры и кристаллографической текстурой, мы далее никак не связываем эти два различных уровня организации биоминеральных объектов: микронный и ангстремный.
Анализ минерального состава и текстуры створок
С помощью нейтронной дифракции в современных, субфоссильных и ископаемых раковинах M. galloprovincialis и M. trossulus обнаружены две минеральные фазы: кальцит и арагонит. Они формируют слои различных микроструктурных элементов (рис. 3, в). В современных и ископаемых раковинах O. edulis выявлен только кальцит (микроструктура скола створки современной O. edulis изображена на рис. 3, г). Замещения иными минералами не обнаружены.
Полюсные фигуры кальцита и арагонита в раковинах M. galloprovincialis сильно отличаются. Для кальцита характерно очень упорядоченное расположение кристаллов. Кристаллографическая текстура в створках этих мидий из Казантипского залива очень острая – 11.50 mrd (табл. I , фиг. 1). Это означает, что большая часть кристаллов кальцита ориентирована в одном направлении. Такие высокие значения возможны только у металлов, подвергнутых интенсивной деформации (Piane, Burlini, 2008). Острота текстуры у кальцита в мраморе 0.06 mrd – 2.62 mrd (Zeisig et al., 2002). Подобные высокие значения, как у M. galloprovincialis, получены для створок современной мидии M. trossulus с побережья о. Адак – 13.10 mrd. Таким образом, текстура кальцита видов рода Mytilus оказалась очень упорядочена и сходна (табл. I , фиг. 1–5 ). Остается вопрос о функциональности такой упорядоченной кристаллографической текстуры.
Рис. 4.
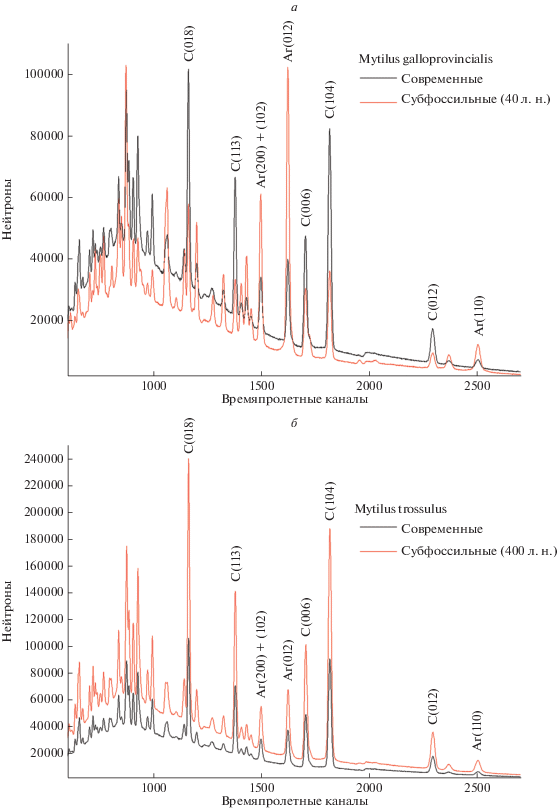
Дифракционные спектры кальцита и арагонита в створках современных (черные) и субфоссильных мидий (красные): а – Mytilus galloprovincialis, берег Казантипского залива (Азовское море, п-ов Крым) напротив с. Песочное; б – M. trossulus, литораль и берег бухты Свипер, о. Адак (Алеутские о-ва), древнеалеутское поселение ADK-009, кухонная куча. Обозначения: C (006) – кальцит (кристаллографическое направление), Ar (012) – арагонит (кристаллографическое направление).
Объяснение к таблице I
Полюсные фигуры кальцита в раковинах Mytilus galloprovincialis, M. trossulus и Ostrea edulis.
Фиг. 1. Створки современных M. galloprovincialis; берег Казантипского залива (Азовское море, п-ов Крым) напротив с. Песочное, от 0 до 2 м от уреза воды.
Фиг. 2. Створки субфоссильных M. galloprovincialis; берег Казантипского залива (Азовское море, п-ов Крым) напротив с. Песочное, около 500 м от берега.
Фиг. 3. Створки современных M. trossulus; литораль бухты Свипер, о. Адак (Алеутские о-ва).
Фиг. 4. Створки субфоссильных M. trossulus; берег бухты Свипер, о. Адак (Алеутские о-ва), древнеалеутское поселение ADK-009, кухонная куча.
Фиг. 5 . Створки ископаемых M. galloprovincialis; п-ов Тамань, коса Тузла; карангатские отложения, верхний плейстоцен.
Фиг. 6 . Створки ископаемых O. edulis; п-ов Тамань, коса Чушка; карангатские отложения, верхний плейстоцен.
Фиг. 7 . Створки современных O. edulis; побережье Черного моря, около п. Малый Утриш.
Фиг. 8 . Створки ископаемых O. edulis; п-ов Крым, Арабатский залив; карангатские отложения, верхний плейстоцен.

Иная ситуация с арагонитом. Его кристаллы ориентированы в различных направлениях, в зависимости от формы створок. Поэтому и текстура арагонита выглядит не так компактно, как это было на полюсных фигурах кальцита (табл. II ). Она менее острая. У M. galloprovincialis (Казантипский залив) текстура достигает значения 2.48 mrd, а у M. trossulus (побережье о. Адак) – 2.46 mrd.
Кристаллографическая текстура кальцита в створках O. edulis с побережья Малого Утриша, сходная по остроте с текстурой арагонита у мидий, только 2.49 mrd. На примере устриц видно, что кристаллы кальцита могут образовывать текстуру разной степени остроты. Она отличается в семействах Mytilidae и Ostreidae. Для подтверждения этого необходимы дальнейшие сравнения текстур мидий и устриц различных родов и видов.
Интересные результаты получены для субфоссильных M. galloprovincialis и M. trossulus. В обоих случаях выявлена высокая острота текстуры кальцита, как и у современных моллюсков. Полюсные фигуры и кальцита, и арагонита выглядят сходно с таковыми для современных представителей видов.
Объяснение к таблице II
Полюсные фигуры арагонита в раковинах Mytilus galloprovincialis и M. trossulus.
Фиг. 1. Створки современных M. galloprovincialis; берег Казантипского залива (Азовское море, п-ов Крым) напротив с. Песочное, от 0 до 2 м от уреза воды.
Фиг. 2. Створки субфоссильных M. galloprovincialis; берег Казантипского залива (Азовское море, п-ов Крым) напротив с. Песочное, около 500 м от берега.
Фиг. 3. Створки современных M. trossulus; литораль бухты Свипер, о. Адак (Алеутские о-ва).
Фиг. 4. Створки субфоссильных M. trossulus; берег бухты Свипер, о. Адак (Алеутские о-ва), древнеалеутское поселение ADK-009, кухонная куча.
Фиг. 5 . Створки ископаемых M. galloprovincialis; п-ов Тамань, коса Тузла; карангатские отложения, верхний плейстоцен.

В случае M. galloprovincialis по результатам дифракции было обнаружено изменение соотношения кальцита и арагонита в створках в сторону увеличения арагонита (рис. 4). Оказалось, что эта особенность никак не связана с перекристаллизацией кальцита в арагонит. Из-за расслоения раковин в изученных створках частично потерян отслоившийся кальцит. Вероятно, этот же процесс привел и к незначительному уменьшению остроты его текстуры. Она достигает 10.86 mrd. А острота текстуры арагонита – 2.76 mrd (табл. II , фиг. 2). Если острота текстуры кальцита уменьшилась, то острота арагонита, наоборот, возросла. Для дальнейшего изучения трансформации кристаллографической текстуры в субфоссильном состоянии необходимы дополнительные измерения. Однако можно сделать заключение, что расслаивание створок моллюсков, содержащих и кальцит, и арагонит, может быть признаком захоронения в условиях суши с сухим и теплым климатом.
У створок M. trossulus происходил иной процесс. Острота текстуры и кальцита, и арагонита возросла. В первом случае она стала 15.45 mrd, а во втором – 2.73 mrd. Можно предположить, что она могла увеличиться за счет тяжести вышележащих отложений, которые давят на раковины, находящиеся внизу. Но порода в верхних слоях разреза не плотная, мощность ее слоя только около 1 м. Она близка к мощности слоя, где были собраны субфоссильные M. galloprovincialis.
У карангатских мидий M. galloprovincialis (рис. 1, е) острота кристаллографической текстуры кальцита и его полюсные фигуры сходны с теми же характеристиками для современных и субфоссильных мидий этого вида. Створки, пролежавшие в отложениях десятки тыс. лет, имеют бóльшую остроту текстуры, чем современные и субфоссильные – 15.72 mrd (табл. I, фиг. 5). Внешне сходны и полюсные фигуры арагонита. Острота текстуры выросла до 3.31 mrd (табл. II, фиг. 5 ).
У современных и карангатских устриц O. edulis (рис. 1, ж) полюсные фигуры кальцита незначительно отличаются. Уже по ним видно, что упорядоченность кристаллов кальцита падает у ископаемых раковин. Линии текстуры “расплываются” по полюсной фигуре. Острота текстуры этого минерала уменьшается до 1.86 mrd у устриц с косы Чушка и до 2.12 mrd, собранных у Щелкино (табл. I , фиг. 6, 8 ).
На примере ископаемых раковин можно предположить, что давление уплотнившихся за тысячи лет вышележащих осадочных пород может оказывать влияние на кристаллографическую текстуру.
Влияние вышележащих осадков на текстуру кальцита в раковинах Mytilus пока представляется наиболее вероятной причиной ее усиления. Но это не подтверждается для раковин O. edulis. Ископаемые устрицы, наоборот, теряют упорядоченность кристаллов кальцита. Поэтому предположение о давлении вышележащих пород, как о возможной причине увеличения остроты кристаллографической текстуры, требует новых доказательств.
Этот метод с использованием нейтронной дифракции очень редко применялся для биологических объектов. Данные, с которыми можно было бы сравнить полученные результаты, отсутствуют. Сравнения с результатами исследований с помощью рентгеновской дифракции и дифракции обратно рассеянных электронов невозможны, поскольку в обоих случаях рассматривается лишь небольшой фрагмент створки, и не во всех ее слоях. Поэтому нельзя с полной уверенностью утверждать о происходящих посмертных текстурных процессах в створках. Мы не можем пока делать окончательные выводы, насколько значимы те временные изменения, которые были выявлены. В дальнейшем предстоит набор статистики измерений. Однако некоторые тенденции просматриваются уже сейчас.
Возникает законное предположение, что на кристаллографическую текстуру могут влиять условия обитания, и тогда вся разница в остроте текстуры кальцита карангатских мидий и устриц и современных черноморских O. edulis и азовских M. galloprovincialis связана с экологическими особенностями. По этому вопросу почти отсутствуют публикации, или же они касаются газового состава, кислотности морской воды (Fitzer et al., 2014). Мы изучили кристаллографическую текстуру створок современных M. galloprovincialis, М. trossulus, O. edulis из разных мест обитания с различными температурами воды и значениями солености. Полученные данные приведены в табл. 1. Текстура кальцита в створках современных M. galloprovincialis варьирует от 11.40 до 12.53 mrd. И тогда можно с большей уверенностью говорить о диагенетических изменениях кристаллографической текстуры кальцита у данного моллюска. Текстура арагонита варьирует в диапазоне 2.48–3.05 mrd. И это тоже подтверждает ее возможные диагенетические изменения. Для М. trossulus из Охотского моря, к сожалению, удалось получить только дополнительные данные по арагониту – 3.44 mrd. Диапазон варьирования остроты кристаллографической текстуры кальцита у O. edulis – 2.49–2.53 mrd. Эти данные могут быть дополнительным аргументом в пользу уменьшения остроты кальцита у ископаемых раковин O. edulis.
Таблица 1.
Максимальная острота текстуры на полюсных фигурах (0006) кальцита и (012)+(121) арагонита в раковинах мидий и устриц. Ископаемые экземпляры выделены полужирным шрифтом
| Образцы | Острота кальцита, mrd | Острота арагонита, mrd |
|---|---|---|
| M. galloprovincialis, Азовское море, Казантипский залив | 11.50 | 2.48 |
| M. galloprovincialis, Адриатическое море | 11.40 | 2.62 |
| M. galloprovincialis, побережье у г. Лангебан, бухта Салданья, Южная Африка | 12.53 | 2.75 |
| M. galloprovincialis, черноморское побережье Румынии | 11.67 | 3.05 |
| M. galloprovincialis, субфоссильные, побережье Казантипского залива | 10.86 | 2.76 |
| M. galloprovincialis, верхний плейстоцен, коса Тузла | 15.72 | 3.31 |
| М. trossulus, Алеутские острова, о. Адак | 13.10 | 2.46 |
| М. trossulus, Охотское море | Кальцита почти нет | 3.44 |
| М. trossulus, субфоссильные, Алеутские острова, о. Адак | 15.45 | 2.73 |
| O. edulis, Португалия, Лагос | 2.53 | – |
| O. edulis, Черное море, Малый Утриш | 2.49 | – |
| O. edulis, верхний плейстоцен, коса Чушка | 1.86 | – |
| O. edulis, верхний плейстоцен, Арабатский залив, Щелкино | 2.12 | – |
ВЫВОДЫ
В результате нашего исследования мы пришли к следующим выводам.
1. Кристаллографические текстуры кальцита раковин моллюсков родов Mytilus и Ostrea отличаются. Текстура кальцита моллюска O. edulis, принадлежащего к семейству Ostreidae, имеет другой характер распределения и не является такой острой, как у моллюсков семейства Mytilidae.
2. Характер текстуры кальцита и арагонита у современных, субфоссильных и ископаемых M. galloprovincialis, а также у современных и субфоссильных M. trossulus практически не меняется, в то время как наблюдаются изменения в ее интенсивности. Такой же вывод можно сделать и при сравнении текстур полностью состоящих из кальцита современных и ископаемых раковин O. edulis.
Список литературы
Антипушина Ж.А., Киселева Н.К., Хасанов Б.Ф. и др. Динамика фауны беспозвоночных литорали о. Адак (Алеутские о-ва) в позднем голоцене по археозоологическим данным // Экология. 2009. № 2. С. 140–148.
Ведь И.П. Климатический атлас Крыма. Симферополь: Таврия-Плюс, 2000. 120 с., 110 карт.
Горбунов Р.В., Горбунова Т.Ю., Кононова Н.К. Климатические нормы температуры воздуха на территории полуострова Крым // Культура народов Причерноморья. 2014. № 278. Т. 2. С. 89–94.
Жирков И.А. Биогеография. Общая и частная: суши, моря и континентальных водоемов. М.: КМК, 2017. 568 с.
Золотарев В.Н. Склерохронология морских двустворчатых моллюсков. Киев: Наук. думка, 1989. 112 с.
Попов С.В. Микроструктура раковины некоторых групп двустворчатых моллюсков. М.: Наука, 1992. 46 с. (Тр. Палеонтол. ин-та РАН. Т. 245).
Федоров П.В. Стратиграфия четвертичных отложений Крымско-Кавказского побережья и некоторые вопросы геологической истории Черного моря. М.: Изд-во АН СССР, 1963. 164 с. (Тр. Геол. ин-та АН СССР. Вып. 88).
Brokmeier H.-G. Texture analysis by neutron diffraction // Mat. Sci. Forum. 1994. V. 157–162. P. 59–70.
Bunge H.J. Texture Analysis in Material Science, Mathematical Methods. L.: Butterworths publ., 1982. 595 p.
Chateigner D., Hedegaard C., Wenk H.-R. Mollusc shell microstructures and crystallographic textures // J. Struct. Geol. 2000. V. 22. P. 1723–1735.
Checa A.G., Macías-Sanchez E., Harper E.M., Cartwright J.H.E. Organic membranes determine the pattern of the columnar prismatic layer of mollusc shells // Proc. Roy. Soc. B. 2016. V. 283. Iss. 1830. https://doi.org/10.1098/rspb.2016.0032
Fitzer S.C., Phoenix V.R., Cusack M., Kamenos N.A. Ocean acidification impacts mussel control on biomineralisation // Sci. Rep. 2014. V. 4. № 6218. P. 1–7. https://doi.org/10.1038/srep06218
Frýda J., Klicnarová K., Frýdová B., Mergl M. Variability in the crystallographic texture of bivalve nacre // Bull. Geosci. 2010. V. 85. № 4. P. 645–662.
Jacob D.E., Ruthensteiner B., Trimby P. et al. Architecture of Anoteropora latirostris (Bryozoa, Cheilostomata) and implications for their biomineralization // Sci. Rep. 2019. V. 9. № 11439. https://doi.org/10.1038/s41598-019-47848-4
Krivoguz D., Bespalova L. Analysis of Kerch peninsula’s climatic parameters in scope of landslide susceptibility // Вестн. Керченского гос. морского технол. ун-та. Биол. науки. 2018. № 2. С. 5–12.
Leslie L.D. Alaska Climate Summaries. Alaska Climate Center Technical Note 5, 2nd ed. Arctic Environ. Information and Data Center, Univ. of Alaska, Anchorage. 1989. 478 p.
Nikolayev D.I., Lychagina T.A., Nikishin A.V., Yudin V.V. Study of error distribution in measured pole figures // Solid State Phenomena. 2005. V. 105 P. 77–82. https://doi.org/10.4028/www.scientific.net/SSP.105.77
Nikolayev D., Lychagina T., Pakhnevich A. Experimental neutron pole figures of minerals composing the bivalve mollusc shells // Springer Nature Appl. Sci. 2019. V. 1. Iss. 4. № 344. https://doi.org/10.1007/s42452-019-0355-1
Nikolayev D.I., Ullemeyer K. A note on preprocessing of diffraction pole density data // J. Appl. Cryst. 1994. V. 27. P. 517–520.
Pearson P.N., Evans S.L., Evans J. Effect of diagenetic recrystallization on the strength of planktonic foraminifer tests under compression // J. Micropalaeontol. 2015. V. 34. P. 59–64.
Piane C.D., Burlini L. Influence of strain history on the mechanical and micro-fabric evolution of calcite rocks: insights from torsion experiments // Swiss J. Geosci. 2008. V. 101. P. 361–375.
Zeisig A., Siegesmund S., Weiss T. Thermal expansion and its control on the durability of marbles // Geol. Soc. Spec. Publ. 2002. V. 205. P. 64–79.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал