

Палеонтологический журнал, 2022, № 1, стр. 51-66
Инкрустирующие мшанки из верхнего мела Среднего Поволжья и Крыма
А. В. Коромыслова a, *, Е. Ю. Барабошкин b, c, **
a Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
b Московский государственный университет имени М.В. Ломоносова
119991 Москва, Россия
c Геологический институт РАН
119017 Москва, Россия
* E-mail: koromyslova.anna@mail.ru
** E-mail: ejbaraboshkin@mail.ru
Поступила в редакцию 03.02.2021
После доработки 14.04.2021
Принята к публикации 28.04.2021
- EDN: MXORFT
- DOI: 10.31857/S0031031X22010093
Аннотация
Изучены мшанки, инкрустирующие ростры белемнитов, из верхнего мела Среднего Поволжья (окрестности гг. Вольск и Шиханы) и Центрального Крыма (г. Ак-Кая). Из-за плохой сохранности колоний в них отсутствуют многие диагностические признаки, поэтому большинство видов приведено в открытой номенклатуре. Изученные мшанки из Среднего Поволжья происходят из сантона–маастрихта и представлены видами Cyclostomata gen. et sp. indet. (класс Stenolaemata), Herpetopora cf. anglica Lang, Hillmeropora sp., ?Marginaria sp., Pyriporella sp., Tyloporella sp., Rhagasostoma gibbosulum Brydone и Aechmellina cf. anglica (Brydone) (класс Gymnolaemata, отряд Cheilostomata). Мшанки г. Ак-Кая происходят из кампана–маастрихта и включают виды Plagioecia sp. (отряд Cyclostomata), ?Electra sp., ?Conopeum sp. и Anornithopora sp. (отряд Cheilostomata). Сведения о хейлостомных мшанках г. Ак-Кая, а также о сантон-кампанском комплексе мшанок Среднего Поволжья приводятся впервые. Изученные виды мшанок из обоих регионов принадлежат к родам, широко распространенным в верхнемеловых отложениях Евразии и США.
ВВЕДЕНИЕ
Мшанки широко распространены в верхнемеловых отложениях Среднего Поволжья и Крыма (Фогт, 1962; Вискова, 1965, 1972, 2004, 2005; Kvachko, 1995; Koromyslova, Seltser, 2020). Тем не менее, новые находки мшанок из этих регионов свидетельствуют о том, что к настоящему времени их таксономический состав полностью не выявлен.
В данной работе описаны новые комплексы мшанок, установленные в сантоне–маастрихте Саратовской обл. (окрестности двух городов: Вольск и Шиханы, правый берег р. Волги) и кампане–маастрихте Центрального Крыма (гора Ак-Кая, Белогорский р-н). Цель данной работы – расширить наши знания о таксономическом разнообразии и палеобиогеографии позднемеловых мшанок Среднего Поволжья и Крыма.
МАТЕРИАЛ И МЕТОДЫ
Коллекция ростров белемнитов с инкрустирующими их мшанками была собрана в середине XX в. Д.П. Найдиным [геологический факультет Московского государственного ун-та им. М.В. Ломоносова (МГУ)] и хранится в Музее землеведения МГУ (МЗ МГУ), №№ 142 и 144.
Все колонии мшанок, обнаруженные на рострах, были изучены на сканирующих электронных микроскопах (СЭМ) Tescan Vega 2 и Tescan Vega 3 в Палеонтологическом ин-те им. А.А. Борисяка Российской академии наук (ПИН РАН), Москва. Образцы изучались без покрытия с использованием детектора обратного рассеяния (BSE), работающего в низком вакууме (10 Па) при напряжении 20 и 30 кВ. Замеры элементов колоний мшанок были получены с СЭМ-изображений. Приведенные размеры в тексте даны в мкм в следующей последовательности: наблюдаемый диапазон, затем (в скобках) среднее арифметическое ± стандартное отклонение, количество образцов (N) и общее количество измерений (n).
Выделены и измерены следующие элементы колоний: АвД – авикулярий, длина; АвШ – авикулярий, ширина; АзД – автозооид, длина; АзШ – автозооид, ширина; АпД – апертура автозооида, длина; АпШ – апертура автозооида, ширина; КзД – кенозооид, длина; КзШ – кенозооид, ширина; ОвД – овицелла, длина; ОвШ – овицелла, ширина; ОпД – опезия автозооида, длина; ОпШ – опезия автозооида, ширина; ОрД – орифис автозооида, длина; ОрШ – орифис автозооида, ширина; ОпAвД – опезия авикулярия, длина; ОпAвШ – опезия авикулярия, ширина.
ГЕОГРАФИЧЕСКОЕ И СТРАТИГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ ИНКРУСТИРОВАННЫХ РОСТРОВ БЕЛЕМНИТОВ
Изученные образцы белемнитов происходят из двух регионов (рис. 1), из близких к кампану–маастрихту стратиграфических интервалов, хотя их точное положение в разрезах неизвестно.
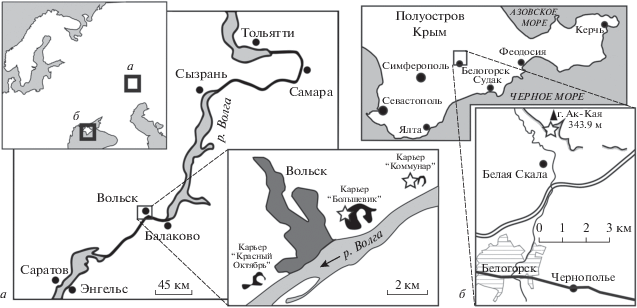
Первая группа местонахождений относится к карьерам, расположенным в окрестностях Вольска и Шихан в Среднем Поволжье (Саратовская обл., Россия). Из окрестностей Шихан в коллекции присутствует один инкрустированный мшанкой обломок белемнита (экз. МЗ МГУ, № 142/8), который может быть условно определен только до семейства Belemnitellidae Pavlow, 1914 (рис. 2, а); происходит он из интервала от сантона до маастрихта.
Рис. 2.
Ростры белемнитов, инкрустированные мшанками, из верхнего сантона–маастрихта окрестностей Шихан (а), Вольска (б–е) и г. Ак-Кая (ж–и): а – ?Belemnitellidae Pavlow, 1914, экз. МЗ МГУ, № 142/8; б, в – Belemnitellidae Pavlow, 1914, экз. МЗ МГУ, № 144/1; г – Belemnitella sp. ex gr. praecursor Stolley, 1897, экз. МЗ МГУ, № 42/4; д – Belemnella licharewi Jeletzky, 1941, экз. МЗ МГУ, № 142/7; е – B. lanceolata (von Schlotheim, 1813), экз. МЗ МГУ, № 144/2; ж – B. lanceolata (von Schlotheim, 1813), экз. МЗ МГУ, № 144/3; з – Belemnitella mucronata (von Schlotheim, 1813), экз. МЗ МГУ, № 144/4; и – Belemnitellidae Pavlow, 1914, экз. МЗ МГУ, № 144/5. Масштабная линейка: 1 см.

Стратиграфическая последовательность и распределение белемнитов в разрезах карьеров “Коммунар” и “Большевик” окрестностей Вольска, откуда происходят находки, подробно описаны ранее (Олферьев и др., 2009а, б, 2014; Барабошкин и др., 2019; Сельцер и др., 2020). Разрез представлен известняками, мергелями и писчим мелом общей мощностью около 40–50 м, и охватывает интервал от турона до маастрихта.
Два образца происходят из карьера “Коммунар”. Экз. МЗ МГУ, № 144/1 (рис. 2, б, в) представлен обломками альвеолярной части белемнита; он не может быть определен точнее, чем представитель семейства Belemnitellidae Pavlow, 1914, и поэтому относится к интервалу от сантона до маастрихта. Экз. МЗ МГУ, № 142/4 (прежний номер 5381/3) (рис. 2, г) имеет более полную сохранность и может быть определен как Belemnitella ex gr. praecursor Stolley, 1897. Эти белемниты распространены в интервале от верхнего сантона до нижнего кампана.
В каком карьере был найден экз. МЗ МГУ, № 142/7 (рис. 2, д), неизвестно. Однако сам образец имеет сравнительно хорошую сохранность и отличается веретеновидной уплощенной в спинно-брюшном направлении формой, характерной для Belemnella licharewi Jeletzky, 1941 и для одноименной зоны верхнего кампана.
Наконец, экз. МЗ МГУ, № 144/2 (прежний номер 6309-5) (рис. 2, е), происходящий из карьера “Большевик”, также имеет достаточно полную сохранность. Он представлен крупным ростром веретеновидной формы и может быть отнесен к Belemnella lanceolata (von Schlotheim, 1813). Как было показано ранее (Барабошкин и др., 2019), данный вид белемнитов характеризует верхнюю часть кампанского яруса и нижнюю часть маастрихтского яруса.
Другое местонахождение – гора Ак-Кая (Белая Скала), расположенная к северу от г. Белогорска (Центральный Крым). Несмотря на то, что этот разрез хорошо обнажен и посещался многими поколениями геологов, детали его строения изучены недостаточно полно. Сведения о строении интересующего нас стратиграфического интервала и распределении белемнитов (по данным А.С. Алексеева) приведены в: Барабошкин и др. (2016). Разрез верхнего кампана–маастрихта представлен здесь толщей мергелей и песчаников (в верхней части) мощностью около 140–150 м, из которых на редких уровнях встречаются ростры белемнитов.
Экз. МЗ МГУ, № 144/3 (прежний номер 5165/9) (рис. 2, ж) имеет более полную сохранность и, вероятно, относится к виду Belemnella lanceolata, который был распространен в позднем кампане и раннем маастрихте.
Экз. МЗ МГУ, № 144/4 (прежний номер 5143/1) (рис. 2, з), вероятно, является кампанским представителем с коротким скульптированным ростром, определенным как Belemnitella mucronata (von Schlotheim, 1813).
Экз. МЗ МГУ, № 144/5 (прежний номер 2548/2) (рис. 2, и) представляет собой фрагмент апикальной части ростра и не может быть определен точнее, чем представитель семейства Belemnitellidae. С учетом строения разреза, этот образец характеризует верхний кампан–маастрихт.
ХАРАКТЕРИСТИКА МШАНОК ИЗ ОКРЕСТНОСТЕЙ ВОЛЬСКА И ШИХАН
Новые находки мшанок из окрестностей Вольска (карьеры “Большевик” и “Коммунар”) и Шихан происходят из интервала сантона–маастрихта. Они принадлежат к восьми видам из классов Stenolaemata (отряд Cyclostomata) и Gymnolaemata (отряд Cheilostomata).
К первому классу отнесен один вид, Cyclostomata gen. et sp. indet. (рис. 3, а), экз. МЗ МГУ, № 144/6, белемнит Belemnitella sp. ex gr. praecursor (экз. МЗ МГУ, № 142/4); верхний сантон–нижний кампан. Фрагмент вертикально-растущей булавовидной колонии, 1 мм высотой и 2 мм шириной, с широким основанием, представлен начальной стадией роста, поэтому необходимые диагностические признаки для определения до родового и видового уровня отсутствуют.
Рис. 3.
Мшанки из верхнего сантона‑маастрихта окрестностей Вольска: а – Cyclostomata gen. et sp. indet., экз. МЗ МГУ, № 144/6; б, в – Herpetopora cf. anglica Lang, 1914, экз. МЗ МГУ, № 142/4b, на в стрелкой показан автозооид с разрушенной закрывающей пластиной; г, д – Hillmeropora sp., экз. МЗ МГУ, № 144/2a; е–и – ?Marginaria sp., экз. МЗ МГУ, № 142/7d. Обозначения: Ав: авикулярий; ?Ан: анцеструла; Гм: гимноциста; К: кауда; Кр: криптоциста; Ов: овицелла; ОШ: основания шипов; ПК: поровая камера. Масштабные линейки: а, в, е – 500 мкм; б, г – 1 мм; д, ж – 200 мкм; з, и ‒ 100 мкм.

Ниже приводится характеристика мшанок отряда Cheilostomata. Из-за плохой сохранности колоний до видового уровня были идентифицированы только три образца, тогда как остальные таксоны приводятся в открытой номенклатуре.
Herpetopora cf. anglica Lang, 1914 (рис. 3, б, в), семейство Electridae Stach, 1937; экз. МЗ МГУ, № 142/4b, белемнит Belemnitella sp. ex gr. praecursor (экз. МЗ МГУ, № 142/4); верхний сантон–нижний кампан. Колония инкрустирующая, представлена ветвящимися цепочками с унисериально расположенными автозооидами. Автозооиды грушевидные, часто с длинной проксимальной гимноцистой (кауда). По размеру опезии автозооидов (Д/Ш = 2.09) колония может быть предположительно отнесена к виду H. anglica (см. Thomas, Larwood, 1960; Taylor, McKinney, 2006). Этот вид имеет широкое распространение и известен из отложений верхнего турона–маастрихта Западной Европы и Северной Америки (Taylor, 1988, 2002; Martha et al., 2017). В Европе H. anglica встречается с верхнего турона до низов кампана (Taylor, McKinney, 2006). Другой близкий вид, H. laxata (d’Orbigny, 1853), встречается, в основном, в кампане и маастрихте и отличается от H. anglica более короткими и узкими апертурами автозооециев (Thomas, Larwood, 1960).
Размеры основных элементов колонии: АзД (с кауда) 470–850 мкм (646 ± 171 мкм; N = 1, n = 5); АзШ 200–340 мкм (274 ± 52 мкм; N = 1, n = 5); ОпД (= ‘ha’ в: Thomas, Larwood, 1960) 380–575 мкм (437 ± 92 мкм; N = 1, n = 4); ОпШ (= ‘la’ в: Thomas, Larwood, 1960) 160–265 мкм (209 ± 43 мкм; N = 1, n = 4).
Hillmeropora sp. (рис. 2, е; 3, г, д), семейство Calloporidae Norman, 1903; экз. МЗ МГУ, № 144/2a, белемнит Belemnella lanceolata (экз. МЗ МГУ, № 144/2); верхний кампан–нижний маастрихт. Колония инкрустирующая мультисериальная; поровые камеры крупные. Автозооиды яйцевидной формы и радиально расходятся от анцеструлы (в изученной колонии она разрушена), опезия крупная (Д/Ш = 1.45). Гимноциста полностью окружает криптоцисту: в проксимолатеральных частях ее ширина варьирует от 30 до 150 мкм, очень узкая в дистальной части. Криптоциста погруженная, узкая в дистальной части и расширяется к проксимальной, однако ее границы в проксимальной части трудно различимы. Овицеллы образованы гимноцистой дистального зооида, эндооеций разрушен. Основания шипов, авикулярии и кенозооиды отсутствуют. Изученная колония может быть отнесена к недавно установленному роду Hillmeropora Martha, Niebuhr et Scholz, 2017, поскольку имеет признаки, отвечающие его диагнозу (Martha et al., 2017). От типового вида этого рода (Hillmeropora pavonina Martha, Niebuhr et Scholz, 2017 из верхнего турона Германии) вольский экземпляр отличается более узкой и погруженной криптоцистой, более короткими автозооидами (вместо АзД 505–651 мкм у H. pavonina), более длинными и узкими опезиями (вместо ОпД 233–296 мкм и ОпШ 215–271 у H. pavonina) и более мелкими овицеллами (вместо ОвД 117–172 мкм и ОвШ 187–240 у H. pavonina).
Размеры основных элементов колонии: АзД (с гимноцистой) 410–540 мкм (485 ± 48 мкм; N = 1, n = 12); АзШ (с гимноцистой) 320–600 мкм (400 ± ± 80 мкм; N = 1, n = 12); ОпД 290–320 мкм (307 ± ± 15 мкм; N = 1, n = 3); ОпШ 190–240 мкм (213 ± 25 мкм; N = 1, n = 3); ОвД 130–170 мкм (143 ± 12 мкм; N = 1, n = 12); ОвШ 120–160 мкм (138 ± 13 мкм; N = 1, n = 12).
?Marginaria sp. (рис. 2, д; 3, е–и), семейство Calloporidae; экз. МЗ МГУ, № 142/7d, белемнит Belemnella licharewi (экз. МЗ МГУ, № 142/7); верхний кампан. Колония инкрустирующая мультисериальная; поровые камеры крупные. Автозооиды грушевидные с крупной опезией (Д/Ш = 1.37). Гимноциста наиболее развита в проксимальной части и полностью окружает узкую криптоцисту. Присутствуют две пары дистальных оснований шипов. Овицеллы образованы гимноцистой дистального зооида, эндооеций разрушен. Авикулярии овальные, с хорошо развитой гимноцистой (?), образуют скопления на отдельных участках колонии. Изученный экземпляр предположительно может быть отнесен к роду Marginaria Römer, 1840, так как в колонии присутствуют скопления мелких авикуляриев, неравномерно разбросанные между автозооидами. Также для представителей этого рода характерно развитие опезиальных шипов (Voigt, 1989). У ?Marginaria sp. присутствуют две пары дистальных оснований шипов, как и у некоторых представителей рода Wilbertopora Cheetham, 1954, напр., W. ostiolatoides Martha, Niebuhr et Scholz, 2017 из верхнего турона Германии. Однако мелкие авикулярии W. ostiolatoides межзооидальные без широкой гимноцисты и скоплений не образуют (Martha et al., 2017). От представителей рода Flustrellaria d’Orbigny, 1853, у которых также могут быть развиты мелкие авикулярии, ?Marginaria sp. отличается развитием только дистальных шипов, а не по всему краю опезии. От описанных видов рода Marginaria из кампана–маастрихта США (Taylor, McKinney, 2006) и маастрихта Мадагаскара (Di Martino et al., 2018) отличается присутствием опезиальных шипов.
Размеры основных элементов колонии: АзД (с гимноцистой) 260–310 мкм (292 ± 22 мкм; N = 1, n = 5); АзШ 160–240 мкм (206 ± 32 мкм; N = 1, n = 5); ОпД 160–180 мкм (170 ± 10 мкм; N = 1, n = 5); ОпШ 110–140 мкм (124 ± 13 мкм; N = 1, n = 5); ОвД 90–110 мкм (100 ± 10 мкм; N = 1, n = 3); ОвШ 90–110 мкм (97 ± 11 мкм; N = 1, n = 3).
Pyriporella sp. (рис. 2, д; 4, а–в), семейство Calloporidae; экз. МЗ МГУ, № 142/7c, белемнит Belemnella licharewi (экз. МЗ МГУ, № 142/7); верхний кампан. Колония инкрустирующая мультисериальная; поровые камеры крупные. Автозооиды яйцевидной формы с крупной опезией (Д/Ш = 1.31). Гимноциста хорошо развита в проксимолатеральной части, криптоциста узкая. Присутствует пара дистолатеральных и пара латеральных оснований шипов. Овицеллы отсутствуют. Авикулярии редкие межзооидальные столбовидные. Изученная колония представлена начальной стадией роста и отличается от других видов рода Pyriporella Canu, 1911 очень редкими межзооидальными авикуляриями, тогда как у P. pedunculata (Shaw, 1967) и P. lacucarinatensis Taylor et McKinney, 2006 из кампана и маастрихта США (Taylor, McKinney, 2006), P. sagittaria (Brydone, 1910) из кампана Англии (Taylor, 2002) и типового вида P. ameghinoi Canu, 1911 из датского яруса Аргентины (Brezina et al., 2021) авикулярии адвентивные, многочисленные, часто развиты на гимноцисте автозооидов. Количество оснований шипов, а также наличие редких авикуляриев на начальной стадии роста сближает Pyriporella sp. с видом P. lacucarinatensis. Однако у изученной Pyriporella sp. автозооиды короткие и узкие (вместо АзД 458–582 мкм, АзШ 242–431 мкм), а опезии более мелкие (вместо ОпД 268–359 мкм, ОпШ 123–260 мкм у P. lacucarinatensis). Кроме того, у изученного экземпляра овицеллы отсутствуют, а у P. lacucarinatensis они гиперстомиальные. Несмотря на указанные отличия, фрагментарность изученной колонии не позволяет устанавливать новые виды.
Размеры основных элементов колонии: АзД 330–460 мкм (408 ± 58 мкм; N = 1, n = 6); АзШ 220–280 мкм (248 ± 20 мкм; N = 1, n = 6); ОпД 210–270 мкм (240 ± 22 мкм; N = 1, n = 5); ОпШ 170–200 мкм (184 ± 11 мкм; N = 1, n = 5); АвД 250–290 мкм (270 ± 28 мкм; N = 1, n = 2); АвШ 100–120 мкм (110 ± 14 мкм; N = 1, n = 2); ОпAвД 110–120 мкм (150 ± 7 мкм; N = 1, n = 2); ОпAвШ 60–80 мкм (70 ± 14 мкм; N = 1, n = 2).
Tyloporella sp. (рис. 4, г, д), семейство Onychocellidae Jullien, 1882; экз. МЗ МГУ, № 142/4c, белемнит Belemnitella sp. ex gr. praecursor (экз. МЗ МГУ, № 142/4); верхний сантон–нижний кампан. Колония крупная, инкрустирующая, мультисериальная. Автозооиды с редуцированной гимноцистой, хорошо развитой криптоцистой и крупной опезией (Д/Ш = 0.95). Авикулярии мелкие, дистолатерально направленные, расположены по одному дистально над каждой опезией автозооида. Овицеллы не наблюдались. Почти вся поверхность колонии, за исключением ее дистального края, стерта, и в связи с этим данный экземпляр может быть определен только до рода. Мшанка Tyloporella sp. отличается от типового вида рода Tyloporella Voigt, 1989, T. reussi Voigt, 1989, происходящего из сеномана Германии (Voigt, 1989; Martha et al., 2017), а также от ?T. lata (Canu, 1911) из маастрихта Мадагаскара (Di Martino et al., 2018) дистолатерально направленными авикуляриями, тогда как у перечисленных выше видов авикулярии направлены дистально. От T. smithi Di Martino et Taylor, 2013 из позднего кампана–маастрихта Объединенных Арабских Эмиратов (Di Martino, Taylor, 2013) и T. cretacea (Canu et Bassler, 1926) из маастрихта США (Taylor, McKinney, 2006) изученный экземпляр отличается дистолатеральным направлением авикуляриев вместо проксимолатерального.
Рис. 4.
Мшанки из сантона–маастрихта окрестностей Вольска: а–в – Pyriporella sp., экз. МЗ МГУ, № 142/7c; г, д – Tyloporella sp., экз. МЗ МГУ, № 142/4c; е–и – Rhagasostoma gibbosulum Brydone, 1936, экз. МЗ МГУ, № 144/1a. Обозначения: Ав: авикулярий; ?Ан: и Ан: анцеструла; Гм: гимноциста; ЗП: закрывающая пластина; Кр: криптоциста; ОШ: основания шипов; ПК: поровые камеры. Масштабные линейки: а, ж, з – 500 мкм; г – 1 мм; б, в, д – 200 мкм; е – 2 мм; и – 100 мкм.

Размеры основных элементов колонии: АзД 650–840 мкм (767 ± 81 мкм; N = 1, n = 6); АзШ 390–490 мкм (432 ± 39 мкм; N = 1, n = 6); ОпД 280–300 мкм (292 ± 9 мкм; N = 1, n = 4); ОпШ 260–360 мкм (310 ± 48 мкм; N = 1, n = 4); АвД 160–180 мкм (167 ± 10 мкм; N = 1, n = 4); АвШ 100–110 мкм (102 ± 5 мкм; N = 1, n = 4).
Rhagasostoma gibbosulum Brydone, 1936 (рис. 2, б; 4, е–и), семейство Onychocellidae; экз. МЗ МГУ, № 144/1a, белемнит Belemnitellidae (экз. МЗ МГУ, № 144/1); сантон–маастрихт. Колония инкрустирующая, мультисериальная. Анцеструла округлая, диаметром 370 мкм. Автозооиды с терминальной D-образной опезией (Д/Ш = 0.98), редуцированной проксимальной гимноцистой и хорошо развитой криптоцистой. Авикулярии межзооидальные с асимметричным рострумом. Овицеллы не наблюдались. Р. Брайдоном (Brydone, 1936) в качестве голотипа R. gibbosulum была описана инкрустирующая колония из среднего кампана Англии, тогда как позже к этому виду были отнесены стержневидные колонии из кампанского отторженца Белоруссии и нижнего маастрихта Германии (Koromyslova et al., 2018b). Вольский экземпляр отличается от ранее описанных экземпляров более крупными опезиями автозооидов.
Размеры основных элементов колонии: АзД 400–560 мкм (494 ± 50 мкм; N = 1, n = 12); АзШ 290–430 мкм (367 ± 40 мкм; N = 1, n = 12); ОпД 120–165 мкм (136 ± 13 мкм; N = 1, n = 12); ОпШ 120–160 мкм (140 ± 13 мкм; N = 1, n = 12); АвД 290–365 мкм (326 ± 37 мкм; N = 1, n = 4); АвШ 80–120 мкм (99 ± 17 мкм; N = 1, n = 4); ОпAвД 60 мкм (60 ± 0 мкм; N = 1, n = 2); ОпAвШ 50–60 мкм (55 ± 7 мкм; N = 1, n = 2).
Aechmellina cf. anglica (Brydone, 1909) (рис. 2, а; 5), семейство Onychocellidae; экз. МЗ МГУ, № 142/8b, белемнит ?Belemnitellidae (экз. МЗ МГУ, № 142/8); сантон–маастрихт. Колония инкрустирующая, мультисериальная. Анцеструла не наблюдалась. Автозооиды яйцевидной формы. Гимноциста отсутствует; криптоциста занимает большую часть фронтальной поверхности, выпуклая. Опезия терминальная округло-треугольная, узкая (Д/Ш = 0.40), с короткими проксимолатеральными опезиулярными выемками; дистальный край приподнят, ростральный, с одним центральным основанием шипа (рис. 5, г, д); проксимальный край прямой, утолщенный, гладкий. Авикулярии межзооидальные, мелкие, каплевидной формы, с симметричным рострумом. Овицеллы не наблюдались. Ранее эта мшанка была ошибочно определена как Cheethamia cf. aktolagayensis Koromyslova, Baraboshkin et Martha, 2018 (см. Коромыслова, Барабошкин, 2021). Однако дальнейшее исследование показало, что изученный экземпляр имеет большое сходство с видом Aechmellina anglica, описанным из нижнего маастрихта Англии (Brydone, 1909). Присутствие дистальных шипов у типовых экземпляров этого вида, а также у экземпляров, описанных Э. Фогтом из нижнего маастрихта г. Хвалынска, Саратовская обл., Россия (Фогт, 1962), и позднего мела Легердорфа, Германия (Voigt, 1949), не указано. Тем не менее, отмечается, что некоторые экземпляры A. anglica могут иметь дистальные основания шипов (Voigt, 1959). От мшанок A. anglica, описанных Фогтом (1962) из нижнего маастрихта г. Хвалынска, A. cf. anglica отличается более крупными автозооидами.
Рис. 5.
Мшанки из сантона–маастрихта окрестностей Шихан: а – Aechmellina cf. anglica (Brydone, 1909), экз. МЗ МГУ, № 142/8b и Voigtella sp., экз. МЗ МГУ, № 142/8a (сверления мшанок показаны стрелками), фотография сделана на микроскопе Leica M165C; б–д – A. cf. anglica, экз. МЗ МГУ, № 142/8b. Обозначения: En: сверления, оставленные губками Entobiai sp.; ОШ: основания шипов. Масштабные линейки: а, в – 1 мм; б – 2 мм; г – 500 мкм; д – 200 мкм.

От типового вида рода Aechmellina Taylor, Martha et Gordon, 2018, A. falcifera (Voigt, 1949), происходящего из нижнего кампана Германии, изученный экземпляр отличается узкой округло-треугольной опезией [у A. falcifera она полуэллиптическая (Taylor et al., 2018)] и выпуклой криптоцистой. Форма опезии и выпуклая криптоциста сближает A. cf. anglica с видами A. stenostoma (Voigt, 1930) из маастрихта о. Рюген, Западного Копетдага, р. Эмбы, плато Актолагай и плато Устюрт (Voigt, 1930, 1967; Никулина, 2001; Koromyslova et al., 2018a); A. lahuseni (Voigt, 1967) из маастрихта плато Устюрт, р. Эмбы и п-ова Мангышлак (Voigt, 1967); A. seriata (Levinsen, 1925) из маастрихта островов Мён и Рюген (Levinsen, 1925; Voigt, 1959), а также с A. viskovae Koromyslova, Baraboshkin et Martha, 2018 из нижнего маастрихта плато Актолагай (Koromyslova et al., 2018a). Однако A. cf. anglica отличается от этих четырех видов мелкими каплевидными авикуляриями, тогда как у A. stenostoma авикулярии длинные со стреловидным симметричным рострумом, у A. lahuseni авикулярии длинные с лопатовидным симметричным рострумом, а у A. seriata и A. viskovae авикулярии мелкие овальные.
Размеры основных элементов колонии: АзД 570–765 мкм (668 ± 61 мкм; N = 1, n = 8); АзШ 410–510 мкм (462 ± 38 мкм; N = 1, n = 8); ОпД 70–90 мкм (79 ± 8 мкм; N = 1, n = 8); ОпШ 180–210 мкм (200 ± 11 мкм; N = 1, n = 8); АвД 200–260 мкм (226 ± 20 мкм; N = 1, n = 8); АвШ 90–115 мкм (106 ± 10 мкм; N = 1, n = 8).
ХАРАКТЕРИСТИКА МШАНОК ГОРЫ АК-КАЯ
Новые находки мшанок из кампана–маастрихта г. Ак-Кая принадлежат классам Stenolaemata и Gymnolaemata. Из-за плохой сохранности колоний все виды приводятся в открытой номенклатуре. Представителями стенолемат является вид Plagioecia sp. (отряд Cyclostomata), гимнолемат – виды ?Electra sp., ?Conopeum sp. и Anornithopora sp. (отряд Cheilostomata). Ниже приводится характеристика этих мшанок.
Plagioecia sp. (рис. 2, и; 6, а–ж), семейство Plagioeciidae Canu, 1918; экз. МЗ МГУ, №№ 144/5a, 144/5b, 144/5c, белемнит Belemnitellidae (экз. МЗ МГУ, № 144/5); верхний кампан–маастрихт. Несколько мультисериальных однослойных колоний инкрустировали ростр белемнита и внутреннюю поверхность раковины моллюска, прикрепленной к тому же ростру. Мшанки отнесены к роду Plagioecia Canu, 1918 на основании обнаруженного у одной из колоний крупного серповидного гонозооида (875 мкм длиной и 1985 мкм шириной), крыша которого пронизана перистомами автозооидов (рис. 6, а, б), что является характерным для представителей этого рода (Harmelin, 1976; Taylor, Sequeiros, 1982; Taylor, McKinney, 2006; Martha et al., 2019). Однако оэциопора не была выявлена. Некоторые апертуры автозооидов закрыты терминальными диафрагмами (рис. 6, ж). Псевдопоры не наблюдались. Особенностью Plagioecia sp. из кампана–маастрихта г. Ак-Кая, также как и некоторых других представителей этого рода, P. cristata Taylor et McKinney, 2006 из маастрихта США и ‘Plagioecia’ sp. из нижнего маастрихта плато Актолагай (Taylor, McKinney, 2006; Koromyslova et al., 2018а), является наличие срединного киля вдоль передней стенки автозооидов, достигающего проксимального края апертуры (рис. 6, в, д, ж). Изученный экземпляр отличается от P. cristata большими размерами апертур (вместо 45–90 мкм в диаметре у P. cristata), а от ‘Plagioecia’ sp. меньшими размерами апертур (вместо АпД 90–190 мкм и АпШ 80–110 мкм у ‘Plagioecia’ sp.). Кроме того, гонозооид актолагайского вида не пронизан апертурами автозооидов.
Рис. 6.
Мшанки из кампана–маастрихта г. Ак-Кая: а–ж – Plagioecia sp.: а–в – экз. МЗ МГУ, № 144/5b; г, д – экз. МЗ МГУ, № 144/5c; е, ж – экз. МЗ МГУ, № 144/5a; з, и – ?Electra sp., экз. МЗ МГУ, № 144/3a. Обозначения: Г: гонозооид; К: киль; П: перистом; РАС: ранняя астогенетическая стадия; ТД: терминальная диафрагма. Масштабные линейки: а, и – 2 мм; б, г, е, з – 1 мм; в, д, ж – 200 мкм.

Размеры основных элементов колонии: АпД 100–150 мкм (128 ± 18 мкм; N = 3, n = 13); АпШ 80–100 мкм (85 ± 7 мкм; N = 3, n = 13).
?Electra sp. (рис. 2, ж; 6, з, и; 7, а, б), семейство Electridae; экз. МЗ МГУ, № 144/3a, белемнит Belemnella lanceolata (экз. МЗ МГУ, № 144/3); верхний кампан–нижний маастрихт. Колония инкрустирующая, мультисериальная. Автозооиды с хорошо развитой проксимальной гимноцистой; криптоциста узкая. Опезии крупные, овальные; присутствует два дистолатеральных шипа и один проксимальный. Овицеллы, авикулярии и кенозооиды не наблюдались. Вид предположительно отнесен к роду Electra Lamouroux, 1816. Особенностью этого рода является хорошо развитая гимноциста, которая может иметь разное количество шипов, включая медиальный шип, расположенный под проксимальным краем опезии. Однако плохо сохранившаяся ранняя астогенетическая стадия (рис. 6, з) не позволяет определить, имела ли изученная колония “сложную многорядную” форму роста, характерную для представителей этого рода (Silén, 1987; Taylor, McKinney, 2006). Кроме того, виды, обладающие схожей морфологией, недавно были описаны в составе рода Conopeum Gray, 1848 (см. Gordon et al., 2020).
Вид ?Electra sp. вместе с E. everretti Taylor et McKinney, 2006 из маастрихта Северной Каролины США, очевидно, являются самыми древними представителями этого рода. Изученный вид отличается от E. everretti более крупными автозооидами и их опезиями (вместо АвД 300–420, АвШ 120–210, ОпД 165‑240, ОпШ 90–150 у E. everretti).
Размеры основных элементов колонии: АзД 670–810 мкм (720 ± 48 мкм; N = 1, n = 5); АзШ 370–435 мкм (399 ± 24 мкм; N = 1, n = 6); ОпД 430–480 мкм (457 ± 21 мкм; N = 1, n = 6); ОпШ 270–330 мкм (295 ± 23 мкм; N = 1, n = 6).
?Conopeum sp. (рис. 2, з; 7, в, г), семейство Electridae; экз. МЗ МГУ, № 144/4a, белемнит Belemnitella mucronata (экз. МЗ МГУ, № 144/4); кампан. Колония инкрустирующая, мультисериальная, анцеструла и ранняя астогенетическая стадия не сохранились. Автозооиды с редуцированной гимноцистой и узкой криптоцистой. Опезия крупная, овальная. Мелкие треугольные зооиды, предположительно, кенозооиды, нерегулярно развиты между автозооидами. Поровые камеры, основания шипов, овицеллы и авикулярии не наблюдались. Изученный экземпляр предположительно отнесен к роду Conopeum, поскольку автозооиды имеют морфологию, характерную для представителей этого рода, а также в колонии присутствуют структуры, подобные кенозооидам, которые часто наблюдаются у представителей этого рода (Taylor, McKinney, 2006; Gordon et al., 2020; Taylor, Rogers, 2021). Имеющийся фрагмент колонии имеет сходство с родом Eokotosokum Taylor et Cuffey, 1992 из маастрихта Канады (Taylor, Cuffey, 1992), выражающееся в морфологии автозооидов и присутствии структур, подобных кенозооидам. Однако от типового вида этого рода E. bicystosum (Allan et Sanderson, 1945) мшанка ?Conopeum sp. отличается более узкой криптоцистой, а также отсутствием дистальной пары оснований шипов. Кроме того, для рода Eokotosokum характерна “сложная многорядная” форма роста колонии (Taylor, Cuffey, 1992), о чем нельзя сказать с уверенностью для ?Conopeum sp. из-за фрагментарности колонии.
Размеры основных элементов колонии: АзД 460–600 мкм (526 ± 43 мкм; N = 1, n = 12); АзШ 240–330 мкм (297 ± 33 мкм; N = 1, n = 12); ОпД 310–400 мкм (349 ± 27 мкм; N = 1, n = 12); ОпШ 150–250 мкм (192 ± 28 мкм; N = 1, n = 12); КзД 230–340 мкм (278 ± 47 мкм; N = 1, n = 5); КзШ 110–150 мкм (130 ± 19 мкм; N = 1, n = 5).
Anornithopora sp. (рис. 2, и; 7, д–к), семейство Cribrilinidae Hincks, 1879; экз. МЗ МГУ, №№ 144/5d, 144/5f, 144/5g, белемнит Belemnitellidae (экз. МЗ МГУ, № 144/5); верхний кампан‑маастрихт. Колония инкрустирующая, мультисериальная, дистальная поровая камера небольшая. Предполагаемая анцеструла округлая, около 190 мкм диаметром (рис. 7, д). Автозооиды с редуцированной гимноцистой и выпуклым фронтальным щитом, который состоит из 10–14 ребер, соединенных четырьмя–пятью перекладинами, формирующими мелкие межреберные поры; дистальная пара ребер сливается, образуя слегка приподнятую апертурную перемычку с центральным выступом (рис. 7, з, и). Орифис D-образный с пятью или больше шипами по дистальному краю (рис. 7, з). Овицеллы гиперстомиальные со срединным швом (рис. 7, и, к). Авикулярии и кенозооиды не наблюдались. Несколько плохо сохранившихся колоний, обрастающих один ростр белемнита, отнесены к роду Anornithopora Lang, 1916. Согласно П. Тэйлору и Ф. Маккини (Taylor, McKinney, 2006), к этому роду следует относить крибриморфных мшанок, которые имеют крошечные зооиды и относительно небольшое количество ребер, а также один тип межзооидальных авикуляриев независимо от их встречаемости. Изученный образец наиболее близок по морфологии автозооидов к A. spooneri (Butler et Cheetham, 1958) из кампана–маастрихта США (Taylor, McKinney, 2006), однако отличается от него отсутствием авикуляриев и более крупными автозооидами и их орифисами (вместо АзД 260–340 мкм и АзШ 170–270 мкм, ОрД 40–70 мкм и ОрШ 60–90 мкм у A. spooneri).
Рис. 7.
Мшанки из кампана‑маастрихта г. Ак-Кая: а, б – ? Electra sp., экз. МЗ МГУ, № 144/3a; в, г – ?Conopeum sp., экз. МЗ МГУ, № 144/4a; д–к – Anornithopora sp.: д, е – экз. МЗ МГУ, № 144/5f; ж, з – экз. МЗ МГУ, № 144/5d; и, к – экз. МЗ МГУ, № 144/5g. Обозначения: Ан: анцеструла; ?Кз: предполагаемые кенозооиды; ОШ: основания шипов; ПК: поровая камера; Ш: шов. Масштабные линейки: а, ж, г – 500 мкм; б, д, и, к – 200 мкм; в – 1 мм; е, з – 100 мкм.

Размеры основных элементов колонии: АзД 340–470 мкм (396 ± 44 мкм; N = 3, n = 8); АзШ 220–310 мкм (274 ± 29 мкм; N = 3, n = 8); ОрД 70–90 мкм (82 ± 8 мкм; N = 3, n = 5); ОрШ 90–110 мкм (100 ± 10 мкм; N = 3, n = 5).
ОБСУЖДЕНИЕ
Комплекс мшанок из сантона–кампана Среднего Поволжья описан впервые. Он включает в себя виды: Cyclostomata gen. et sp. indet., Herpetopora cf. anglica Lang, 1914, ?Marginaria sp., Pyriporella sp. и Tyloporella sp. Кроме того, в интервале сантона–маастрихта встречены виды Rhagasostoma gibbosulum и Aechmellina cf. anglica, а в интервале верхнего кампана–нижнего маастрихта обнаружен вид Hillmeropora sp. Ранее из Среднего Поволжья были известны мшанки только из верхнего турона и маастрихта (Фогт, 1962; Вискова, 1965, 1972, 1992, 2004, 2005; Kvachko, 1995; Koromyslova, Seltser, 2020), которые не включают рассмотренных в статье таксонов.
Виды Herpetopora cf. anglica и Rhagasostoma gibbosulum имеют широкое географическое распространение. Первый происходит из отложений верхнего турона–маастрихта Западной Европы и США (Taylor, 1988, 2002; Taylor, McKinney, 2006; Martha et al., 2017), второй вид – из среднего кампана–нижнего маастрихта Англии, Белоруссии и Германии (Brydone, 1936; Koromyslova et al., 2018b). Возможно, к виду R. gibbosulum принадлежит мшанка, описанная Г. Пугачевской (Pugaczewska, 1965) как Onychocella sp., инкрустирующая ростр Belemnitella sp. из верхнего кампана–маастрихта окрестностей Мельника (Польша). Вид Hillmeropora sp. относится к роду, известному ранее только из верхнего турона Германии (Martha et al., 2017). Экземпляр ?Marginaria sp. отнесен к роду, известному из позднего мела Европы (Voigt, 1989; Martha et al., 2017), кампана–маастрихта США (Taylor, McKinney, 2006), маастрихта Мадагаскара (Di Martino et al., 2018) и миоцена Индии (Guha, Gopikrishna, 2007). Род Pyriporella известен из кампана Англии (Taylor, 2002), кампана–маастрихта США (Taylor, McKinney, 2006), датского яруса Аргентины (Canu, 1911; Brezina et al., 2021) и неогена Индии (Sonar, Pawar, 2016). Род Tyloporella встречен в сеномане Германии (Voigt, 1989; Martha et al., 2017), маастрихте Мадагаскара (Di Martino et al., 2018), верхнем кампане–маастрихте ОАЭ (Di Martino, Taylor, 2013) и маастрихте США (Taylor, McKinney, 2006). К недавно установленному роду Aechmellina отнесены виды, широко распространенные в позднем мелу Евразии и Северной Америки (Taylor et al., 2018).
Многочисленные сверления мшанок Voigtella sp. (отряд Ctenostomata из класса Gymnolaemata) были обнаружены на ростре, инкрустированном мшанкой Aechmellina cf. anglica (рис. 5, а). На других позднемеловых рострах белемнитов из Среднего Поволжья ранее были встречены сверления, принадлежащие мшанкам ?Voigtella prima (Voigt, 1962) и ?V. cf. prima (окрестности Вольска), а также V. regalis Pohowsky, 1978 (окрестности г. Сенгилей, Ульяновская обл.) (Фогт, 1962; Voigt, Soule, 1973; Pohowsky, 1978; Вискова, 1992; Коромыслова, Барабошкин, 2021).
Изученный комплекс мшанок из кампана–маастрихта г. Ак-Кая включает в себя виды Plagioecia sp. (отряд Cyclostomata), ?Electra sp., ?Conopeum sp. и Anornithopora sp. (отряд Cheilostomata). Сведения о хейлостомных мшанках для этого района приводятся впервые. Ранее только два представителя отряда Cyclostomata, Osculipora (=Desmepora) semicylindrica (Rӧmer, 1840) и Crisina macropora (Marsson, 1887), были описаны из верхнего маастрихта г. Ак-Кая (Вискова, 1972, 1992). За исключением O. semicylindrica и Anornithopora sp., все остальные виды изученного комплекса принадлежат к родам (Crisina, Plagioecia, Conopeum, Electra), существующим в современных морях. Род Anornithopora известен из кампана Англии (Lang, 1916), кампана–маастрихта США (Taylor, McKinney, 2006), маастрихта Мадагаскара (Di Martino et al., 2018), палеоцена Дании (Berthelsen, 1962) и Германии (Voigt, 1925). Род Plagioecia имеет широкое стратиграфическое и географическое распространение, начиная с байоса (Taylor, McKinney, 2006). Меловые представители рода Conopeum известны из верхнего сеномана Туркменистана (Фаворская, 1990, 1996), верхнего турона Германии (Martha et al., 2017), кампана–маастрихта США (Taylor, McKinney, 2006) и ОАЭ (Di Martino, Taylor, 2013), верхнего мела Индии (Guha, Nathan, 1996). ?Electra sp., отмеченный здесь, вместе с E. everretti из маастрихта США, вероятно, являются самыми древними представителями этого рода.
ВЫВОДЫ
Впервые приведены сведения о комплексах мшанок из сантона–кампана Среднего Поволжья и кампана–маастрихта г. Ак-Кая (Центральный Крым), поселявшихся на рострах белемнитов, которые формировали твердые субстраты позднемеловых бассейнов. Изученные формы принадлежат к родам, широко распространенным в верхнемеловых отложениях Евразии и США. Представители родов мшанок, описанных из кампана–маастрихта г. Ак-Кая, кроме того, встречаются и в современных морях.
Наиболее полные данные имеются по позднемеловым мшанкам Европы, Среднего Поволжья и плато Актолагай (Западный Казахстан), выявляющие сходство комплексов. Из первых двух регионов известно 14 общих видов: Phormopora irregularis Marsson, 1887, P. langethalii (Marsson, 1887), Sulcocava cristata Orbigny, 1854, Stomatoporopsis multigemmans (Illies, 1974), Diplosolen pavonius Voigt, 1929, Herpetopora cf. anglica, Dionella trifaria (von Hagenow, 1846), Hoplitaechmella nitescens (Brydone, 1914), H. vespertilio (von Hagenow, 1839), Stichomicropora biconstricta (von Hagenow, 1839), Rhagasostoma gibbosulum, R. cf. saltans (Brydone, 1930), Aechmellina anglica и Luganella goldfussi (von Hagenow, 1839) (Фогт, 1962; Вискова, 1972, 1992, 2004; Koromyslova, Seltser, 2020). Из верхнего мела Среднего Поволжья и плато Актолагай известно четыре общих вида – Voigtella regalis, ?V. cf. prima, Cheethamia aktolagayensis и R. cf. saltans (Pohowsky, 1978; Koromyslova et al., 2018a; Коромыслова, 2020; Koromyslova, Seltser, 2020).
Список литературы
Барабошкин Е.Ю., Аркадьев В.В., Копаевич Л.Ф. Опорные разрезы меловой системы Горного Крыма // Путеводитель полевых экскурсий Восьмого Всероссийского совещания 26 сентября–3 октября 2016 г. / Ред. Е.Ю. Барабошкин. Симферополь: Издат. Дом ЧерноморПресс, 2016. 90 с.
Барабошкин Е.Ю., Беньямовский В.Н., Гужиков А.Ю. и др. Комплексное изучение пограничного интервала кампана / маастрихта в Поволжье (Россия) и на плато Актолагай (Западный Казахстан) Русской плиты // Проблемы геологии и расширение минерально-сырьевой базы стран Евразии: Матер. междунар. науч. конф. / Ред. Г.Ж. Жолтаев. Алматы: ТОО “Ин-т геол. наук К.И. Сатпаева”, 2019. С. 69–79.
Вискова Л.А. Пoзднeмeлoвыe мшанки рода Meliceritites Среднего Поволжья // Палеонтол. журн. 1965. № 3. С. 49–58.
Вискова Л.А. Позднемеловые мшанки Cyclostomata Поволжья и Крыма. М.: Наука, 1972. 96 с. (Тр. Палеонтол. ин-та АН СССР. Т. 132).
Вискова Л.А. Морские постпалеозойские мшанки. М.: Наука, 1992. 187 с. (Тр. Палеонтол. ин-та РАН. Т. 250).
Вискова Л.А. Мшанки рода Stomatoporopsis Illies, 1981 (Tubuliporina, Stenolaemata), и их первые находки в маастрихте Среднего Поволжья и Мангышлака // Палеонтол. журн. 2004. № 6. С. 64–72.
Вискова Л.А. Новые мшанки (Tubuliporina, Stenolaemata) из верхнего мела Среднего Поволжья (Россия) // Палеонтол. журн. 2005. № 4. С. 56–62.
Коромыслова А.В. Новые данные о мшанках нижнего маастрихта плато Актолагай (Западный Казахстан) // Меловая система России и ближнего зарубежья: проблемы стратиграфии и палеогеографии: Матер. X Всерос. совещ. / Ред. Е.Ю. Барабошкин, А.Ю. Гужиков. Магадан: ОАО “МАОБТИ”, 2020. С. 120–121.
Коромыслова А.В., Барабошкин Е.Ю. Сверлящие мшанки и другие следы биоэрозии в позднемеловых белемнитах из Среднего Поволжья и Западного Казахстана // Палеонтология, биостратиграфия и палеогеография мезозоя и кайнозоя бореальных районов: Матер. науч. онлайн-сессии, 19–22 апреля 2021 г. [электронный ресурс] / Ред. Н.К. Лебедева, А.А. Горячева, О.С. Дзюба, Б.Н. Шурыгин. Новосибирск: ИНГГ СО РАН, 2021. С. 101–106.
Никулина Е.А. Изменение закономерностей развития зооидных почек в эволюции мшанок отряда Cheilostomata и появление многорядных форм // Палеонтол. журн. 2001. № 5. С. 58–63.
Олферьев А.Г., Беньямовский В.Н., Иванов А.В. и др. Верхнемеловые отложения севера Саратовской области. Статья 1. Разрез карьера ‘‘Большевик” в окрестностях Вольска // Бюлл. МОИП. Отд. геол. 2009а. Т. 84. Вып. 2. С. 5–22.
Олферьев А.Г., Беньямовский В.Н., Иванов А.В. и др. Верхнемеловые отложения севера Саратовской области. Статья 2. Биостратиграфическое расчленение разреза карьера “Большевик” в окрестностях Вольска // Бюлл. МОИП. Отд. геол. 2009б. Т. 84. Вып. 4. С. 29–46.
Олферьев А.Г., Сельцер В.Б., Алексеев А.С. и др. Верхнемеловые отложения севера Саратовской области. Статья 3. Биостратиграфическое расчленение разреза карьера “Красный Октябрь” на южной окраине г. Вольска // Бюлл. МОИП. Отд. геол. 2014. Т. 89. Вып. 6. С. 45–76.
Сельцер В.Б., Первушов Е.М., Калякин Е.А. О сантонских отложениях вольских меловых карьеров // Недра Поволжья и Прикаспия. 2020. Вып. 102. С. 61–81.
Фаворская Т.А. Хейлостомные мшанки из позднего сеномана юго-западных отрогов Гиссарского хребта // Ежегодн. ВПО. 1990. Т. 33. С. 51–63.
Фаворская Т.А. Практическое руководство по макрофауне России и сопредельных территорий. Мшанки мезокайнозоя. СПб.: ВСЕГЕИ, 1996. 81 с.
Фогт Э. Верхнемеловые мшанки европейской части СССР и некоторых сопредельных областей. М.: Изд-во Моск. ун-та, 1962. 125 с.
Berthelsen O. Cheilostome Bryozoa in the Danian deposits of east Denmark // Danmarks Geol. Unders. 1962. V. 83. P. 1–290.
Brezina S.S., Taylor P.D., Romero M.V. et al. Upper Maastrichtian and Danian bryozoans from Northern Patagonia, Argentina // Cret. Res. 2021. V. 125. https://doi.org/10.1016/j.cretres.2021.104845
Brydone R.M. Notes on new or imperfectly known Chalk Bryozoa (Polyzoa) // Geol. Mag. N. S. 1909. V. 6. P. 337–339.
Brydone R.M. Further notes on new or imperfectly known Chalk Polyzoa. Part III. (Semieschara, Micropora, Cryptostoma, etc.). London: Privately published, 1936. P. 61–90.
Canu F. Iconographie des Bryozoaires fossiles de l’Argentine // An. Museo Nac. Hist. Natur. Buenos Aires. Ser. 3. 1911. V. 14. P. 215–288.
Di Martino E., Taylor P.D. First bryozoan fauna from a tropical Cretaceous carbonate: Simsima Formation, United Arab Emirates–Oman border region // Cret. Res. 2013. V. 43. P. 80–96.
Di Martino E., Martha S.O., Taylor P.D. The Madagascan Maastrichtian bryozoans of Ferdinand Canu–Systematic revision and scanning electron microscopic study // Ann. Paléontol. 2018. V. 104. P. 101–128.
Gordon D.P., Sutherland J.E., Perez B.A et al. The bryozoan genus Conopeum (Electridae) in New Zealand, with description of a new species and discussion of the morphological and genetic characters of Conopeum seurati (Canu, 1928) // J. Natur. Hist. 2020. V. 54. P. 947–970.
Guha A.K., Nathan D.S. Bryozoan fauna of the Ariyalur Group (Late Cretaceous). Tamilnadu and Pondicherry, India // Palaeontol. Ind. (N. Ser.). 1996. V. 49. P. 1–217.
Guha A.K., Gopikrishna K. New calloporid (Bryozoa, Cheilostomata) species from Tertiary sequences of western Kachchh, Gujarat // J. Geol. Soc. India. 2007. V. 70. P. 121–130.
Harmelin J.-G. Le sous-ordre des Tubuliporina (bryozoaires cyclostomes) en Méditerranée. Écologie et systématique // Mém. Inst. Océanogr. 1976. V. 10. P. 1–326.
Koromyslova A.V., Baraboshkin E.Y., Martha S.O. Late Campanian to late Maastrichtian bryozoans encrusting on belemnite rostra from the Aktolagay Plateau in western Kazakhstan // Geobios. 2018a. V. 51. P. 307–333.
Koromyslova A.V., Taylor P.D., Martha S.O., Riley M.. Rhagasostoma (Bryozoa) from the Late Cretaceous of Eurasia: Taxonomic revision, stratigraphy and palaeobiogeography // Eur. J. Taxonomy. 2018b. V. 490. P. 1–66.
Koromyslova A.V., Seltser V.B. Early Maastrichtian cheilostome bryozoans from the middle Volga River region // Paläontol. Z. 2020. V. 94. P. 697–714.
Kvachko V.I. Late Cretaceous and Paleocene bryozoans of the genus Lunulites from the middle Volga, Crimea, and Mangyshlak // Paleontol. J. 1995. V. 29. № 4. P. 36–45.
Lang W.D. A revision of the “cribrimorph” Cretaceous Polyzoa // Ann. Mag. Natur. Hist. Ser. 8. 1916. V. 18. P. 81–112, 381–410.
Levinsen G.M.R. Undersøgelser over Bryozoerne i den danske Kridtformation. Efter Forfatterens død udg. af K. Brünnich Nielsen og Th. Mortensen. Det Kongelige Danske Videnskabernes Selskabs Skrifter // Naturvid. Math. Afdeling. 1925. № 7. P. 283–445.
Martha S.O., Niebuhr B., Scholz J. Cheilostome Bryozoen // Geol. Saxonica. 2017. V. 62. P. 11–52.
Martha S.O., Taylor P.D., Rader W.L. Early Cretaceous cyclostome bryozoans from the early to middle Albian of the Glen Rose and Walnut formations of Texas, USA // J. Paleontol. 2019. V. 93. № 2. P. 244–259.
Pohowsky R.A. The boring ctenostomate Bryozoa: Taxonomy and paleobiology based on cavities in calcareous substrata // Bull. Amer. Paleontol. 1978. V. 73. № 301. P. 1–192.
Pugaczewska H. Les organismes sédentaires sur les rostres des bélemnites du Crétacé supérieur // Acta Palaeontol. Pol. 1965. V. 10. P. 73–95.
Silén L. Colony growth pattern in Electra pilosa (Linnaeus) and comparable encrusting bryozoans // Acta Zool. (Stockholm). 1987. V. 68. Iss. 1. P. 17–34.
Sonar M.A., Pawar R.V. Some calloporid (Bryozoa: Cheilostomata) species from the Cenozoic sediments of Western Kachchh, Gujarat // J. Geol. Soc. India. 2016. V. 88. P. 47–54.
Taylor P.D. Colony growth pattern and astogenetic gradients in the Cretaceous cheilostome bryozoan Herpetopora // Palaeontology. 1988. V. 31. P. 519–549.
Taylor P.D. Bryozoans // Fossils of the Chalk. 2nd ed. / Eds A.B. Smith, D.J. Batten. L.: Palaeontol. Assoc., 2002. C. 53–75.
Taylor P.D., Cuffey R.J. Cheilostome bryozoans from the Upper Cretaceous of the Drumheller area, Alberta, Canada // Bull. Brit. Museum (Natur. Hist.), Geol. 1992. V. 48. P. 13–24.
Taylor P.D., Martha S.O., Gordon D.P. Synopsis of ‘onychocellid’ cheilostome bryozoan genera // J. Natur. Hist. 2018. V. 52. P. 1657–1721.
Taylor P.D., McKinney F.K. Cretaceous Bryozoa from the Campanian and Maastrichtian of the Atlantic and Gulf Coastal Plains, United States // Scripta Geol. 2006. V. 132. P. 1–346.
Taylor P.D., Rogers R.R. A new cheilostome bryozoan from a dinosaur site in the Upper Cretaceous (Campanian) Judith River Formation of Montana // J. Paleontol. 2021. V. 95. № 5. P. 965–973.
Taylor P.D., Sequeiros L. Toarcian bryozoans from Belchite in north-east Spain // Bull. Brit. Museum (Natur. Hist.), Geol. 1982. V. 36. P. 117–129.
Thomas H.D., Larwood G.P. The Cretaceous species of Pyripora d’Orbigny and Rhammatopora Lang // Palaeontology. 1960. V. 3. P. 370–386.
Voigt E. Neue cribrimorphe Bryozoen aus der Familie der Pelmatoporidae in Kreidegeschieben Anhalts // Z. Geschiebeforsch. 1925. Bd 1. P. 97–104.
Voigt E. Morphologische und stratigraphische Untersuchungen über die Bryozoenfauna der oberen Kreide. I. Teil. Die cheilostomen Bryozoen der jüngeren Oberkreide in Nordwestdeutschland, im Baltikum und in Holland // Leopoldina: Ber. Kais.-Dtsch. Akad. Naturforsch. Halle. 1930. № 6. S. 379–579.
Voigt E. Cheilostome Bryozoen aus der Quadratenkreide Nordwestdeutschlands // Mitt. Geol. Staat. Hamburg. 1949. H. 19. S. 1–49.
Voigt E. Revision der von F. v Hagenow 1838-1850 aus der Schreibkreide von Rügen verӧffentlichten Bryozoen // Geologie. 1959. V. 8. S. 1–80.
Voigt E. Oberkreide-Bryozoen aus den asiatischen Gebieten der UdSSR // Mitt. Geol. Staat. Hamburg. 1967. H. 36. S. 5–95.
Voigt E. Beitrag zur Bryozoen-Fauna des sächischen Cenomaniums // Abh. Staat. Mus. Miner. Geol. Dresden. 1989. Bd 36. P. 8–87.
Voigt E., Soule J.D. Cretaceous burrowing bryozoans // J. Paleontol. 1973. V. 47. № 1. P. 21–33.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал