

Палеонтологический журнал, 2022, № 2, стр. 109-116
Распространение рода Dzergalanella Genkina (членистостебельные) в пермских и мезозойских отложениях Сибири и Европы
Г. Н. Садовников *
Российский государственный геолого-разведочный университет
117997 Москва, Россия
* E-mail: sadovnikov.gennady@yandex.ru
Поступила в редакцию 28.12.2020
После доработки 20.09.2021
Принята к публикации 20.09.2021
- EDN: ZQJIPL
- DOI: 10.31857/S0031031X22020118
Аннотация
Род Dzergalanella Genkina (членистостебельные) первоначально был описан из Киргизии. Он широко распространен на севере Сибири и отмечен в Европе. Для этого рода характерны мутовчатые листья с многочисленными жилками. Сейчас в большинстве случаев поверхность высоких листовых влагалищ принимают за стебель и относят эти растения к Neocalamites и другим родам. Автор включает в Dzergalanella виды, описанные ранее как Arthropitys (?) prynadae, A. (?) tunguskanus, и дополняет диагноз информацией об анатомическом строении стебля.
ВВЕДЕНИЕ
Dzergalanella установлен Р.З. Генкиной (1966) в нижней юре Киргизии. Растение, по облику вполне соответствующее членистостебельным, имело членистый стебель и расположенные мутовками узкие листья. Но в листе была не одна жилка, как обычно бывает у членистостебельных, а несколько параллельных жилок.
В 1971 г. автор описал из вулканогенных отложений верхов перми Средней Сибири два вида членистостебельных в анатомической сохранности и условно отнес их к роду Arthropitys Goeppert (Садовников, 1971). Растения с близким типом стелы были позднее описаны из Антарктиды (Osborn, Taylor, 1989; Ryberg et al., 2008).
Один из видов – Arthropitys (?) prynadae Sadovnikov – представлял собой безлистные стебли, и применение к нему этого родового названия было вполне правомерным. Второй вид – A. (?) tunguskanus Sadovnikov – был представлен побегами, и применение для него названия рода, предложенного для стеблей, вряд ли было вполне корректно. Нужно отметить, что у обоих видов были весьма многочисленные (до 90) проводящие пучки (что не свойственно роду Arthropitys). Листья располагались мутовками, высота которых могла не менее чем втрое превышать длину междоузлий, и полностью срастались, образуя узко-конические листовые влагалища.
К.-П. Кельбер опубликовал фотографии Neocalamites merianii (Brongiart) Halle из нижнего кейпера (вероятно, ладинского яруса) Германии (Brunner, Kelber, 1988; Kelber, Hansch, 1995; Kelber, 2015). На одной из фотографий (Kelber, Hansch, 1995, рис. 105 ) видно, что листья растения в большей нижней части срастаются, образуя очень высокое трубчатое листовое влагалище, которое ошибочно принимают за стебель (рис. 1). Подобные ошибки при изучении ископаемых членистостебельных выявлялись и ранее (Meyen, 1971; Мейен, 1990).
Рис. 1.
Схема строения побега Neocalamites merianii (по фотографии из: Kelber, Hansch, 1995, × 1): а – ядро, черные линии – четкие хорошо видные тонкие ребра – выполнения породой каринальных лакун; б – фитолейма стебля, белые линии – своды пологих продольных волнообразных поднятий на поверхности стебля; в – фитолейма трубчатого листового влагалища (внизу) и фрагментов листьев (зубцов) (выше), белые линии – своды пологих продольных волнообразных поднятий на поверхности стебля, деформирующие листовое влагалище; красные линии – жилки (видны плохо); г – вмещающая порода.

Растения, которые идентифицировались как Neocalamites merianii, широко распространены в триасовых отложениях севера России (Коломийченко и др., 2001; Садовников и др., 2002). При подготовке к публикации каталога триасовых растений Таймыра (Садовников, 2007, 2015а) автор установил, что у представителей этого вида с п-ова Таймыр имеет место то же самое, что видно на фотографиях К.-П. Кельбера и В. Ханша в Германии: листья в большей нижней части срастаются, образуя очень высокое трубчатое листовое влагалище (Садовников, 2015а, с. 66–69). Но, кроме того, на таймырском материале было установлено, что в каждом листе имеется не одна жилка, а несколько параллельных жилок (или тяжей механической ткани). Такое явление не свойственно Neocalamites, но именно оно, как было сказано, характерно для рода Dzergalanella (Генкина, 1966).
Генкина причислила род Dzergalanella к членистостебельным условно, предложив для него следующий диагноз (Генкина, 1966, с. 54–55): “Листья (?) одиночные, либо изолированные лопасти (?) мутовок, крупные и средних размеров. Удлиненно-эллипсовидные в очертании, 50–80 мм длиной и наибольшей шириной 12–18 мм в средней части, с тупо-округлой верхушкой и несколько клиновидносуженным основанием. Жилкование двоякого типа: четыре–шесть сильных жилок выходят из основания листа (или лопасти), слегка дугообразно изгибаясь, идут параллельно краям пластинки и вновь изгибаются по направлению к верхушке. Между основными сильными жилками расположены три–пять более тонких, но ясных жилок, которые выходят из основания, затем пробегают параллельно друг другу и в верхней части изгибаются по направлению к притупленной верхушке. Пластинка имеет тенденцию расщепляться по основным сильным жилкам на отдельные узкие “листья”.
Далее в разделе “Сравнение” Генкина указала: “Возможно, что отмеченные нами сильные жилки, по которым пластинка имеет тенденцию расщепляться на узкие “листья”, представляют собой швы срастания смежных узких листьев, подобных тем, какие имеют место у представителей рода Schizoneura …”. Из двух предложенных автором рода трактовок остатков Dzergalanella нам представляется малоубедительной первая (широкие листья с двумя типами жилок, различающихся по толщине), и мы, напротив, полностью согласны со второй (лопасти мутовок из сросшихся узких листьев).
Автор (Садовников, 2001, 2015а) предложил причислить, по крайней мере, печорские и сибирские, а может быть, и часть западноевропейских растений, определявшихся как Neocalamites merianii, к роду Dzergalanella, скорректировав его диагноз. В этот же род могут быть включены Arthropitys (?) tunguskanus и A. (?) prynadae. Вполне возможно, что к этому же роду относятся некоторые Equisetites, в частности, E. arenaceus (Jaeger) Schenk. Но этот вопрос требует специального рассмотрения.
ОПИСАНИЕ ВИДОВ
Род Dzergalanella Genkina, 1966, emend. Sadovnikov, emend. nov.
Dzergalanella Genkina, 1966: Генкина, 1966, с. 54–55; Садовников, 2015а, с. 61.
Arthropitys Goeppert, 1864 (part.): Садовников, 1971, с. 830.
Neocalamites Halle, 1908 (part.): Kelber, Hansch, 1995, с. 48–52; Садовников, 2013б, с. 92; Садовников и др., 2002, с. 125.
Типовой вид – Dzergalanella dzergalanensis Genkina, 1966; нижняя юра Киргизии.
Diagnosis. The stem is rounded, sometimes obtusely tetrahedral, with a broad smooth-walled core cavity. The surface of the stem in cross-section is smooth or gently wavy. Vascular bundles are numerous. Primary xylem is mesarch. The protoxylem and the part of the metaxylem facing the core are usually destroyed. Then on the surface of the core, clear sharp thin longitudinal ribs are visible, corresponding to the carinal channels. Leaves are linear, arranged in whorls. In most of the lower part, they coalesce and form barrel-shaped (divided into blades), conical or tubular leaf sheaths. The teeth (previously taken for full leaves) are long.
Диагноз. Стебель округлый, иногда тупо-четырехгранный, с широкой гладкостенной сердцевинной полостью. Поверхность стебля в поперечном сечении ровная или широко-волнистая. Проводящие пучки многочисленные с мезархной первичной ксилемой. Протоксилема и часть метаксилемы, обращенная к сердцевине, обычно разрушаются. Тогда на поверхности ядра видны четкие резкие тонкие продольные ребра, отвечающие каринальным каналам. Листья линейные, расположены мутовками. Они срастаются и образуют очень высокие бочонковидные, конические или трубчатые листовые влагалища (первые могут разделяться на широкие лопасти).
Видовой состав. D. dzergalanensis Genkina emend. Sadovnikov (Киргизия, юра), D. prynadae (Sadovnikov) Sadovnikov (Средняя Сибирь, верхняя пермь), D. taimyrica sp. nov. (Таймыр, триас), D. tunguskana (Sadovnikov) Sadovnikov (Средняя Сибирь, верхняя пермь).
Сравнение. От большинства родов членистостебельных (имеющих в листе одну жилку, ветвящуюся или неветвящуюся), Dzergalanella отличается наличием в листе нескольких неветвящихся жилок.
Dzergalanella dzergalanensis Genkina, 1966, emend. Sadovnikov, emend. nov.
Dzergalanella dzergalanensis: Генкина, 1966, с. 55, табл. V, фиг. 4–8 .
Голотип – Обр. 536, место хранения не указано (Генкина, 1966, табл. V фиг. 8 ); Киргизия, южный берег оз. Иссык-Куль, бассейн р. Джергалан; нижний лейас, джильская свита.
Diagnosis (emended). The leaves are narrow, with 3 to 5 thin but clear veins that emerge from the base, then run parallel to each other and bend towards the blunted tip at the top. The leaves are arranged in whorls and coalesce into isolated elongated-ellipsoid lobes with a blunt-rounded tip and a somewhat wedge-shaped narrowed base. The length of the blades is 50–80 mm, the maximum width is 12–18 mm (in the middle part).
Описание. Листья узкие, с тремя–пятью тонкими, но ясными жилками, которые выходят из основания, затем пробегают параллельно друг другу и в верхней части изгибаются по направлению к притупленной верхушке. Листья располагаются мутовками и срастаются в изолированные удлиненно-эллипсовидные лопасти с тупо-округлой верхушкой и несколько клиновидно суженным основанием. Длина лопастей 50–80 мм, наибольшая ширина 12–18 мм в средней части.
Распространение. Нижний лейас; Киргизия, южный берег оз. Иссык-Куль: басс. р. Джергалан, базальная и угленосная свиты; басс. р. Согуты11, джильская свита.
Материал. 15 экз. из типового местонахождения.
Dzergalanella prynadae (Sadovnikov, 1971) Sadovnikov, comb. nov.
Табл. XIV , фиг. 1, 3, 4, 6, 7 (см. вклейку)
Рис. 2.
Схематические поперечные сечения побегов Dzergalanella (по фотографиям, × 1): а–ж – D. рrynadae: а – обр. ГИН 3760/1, б, в – обр. ГДМ 15323, г – обр. ГДМ 18622/1-3, д–ж – обр. ГДМ 18625; з–р – D. tunguskana: з – обр. ГИН 3760/4, и – обр. ГИН 3760/5, к–м – обр. ГДМ 18623, н – обр. ГДМ 18626/3, о – обр. ГДМ 18626/4, п, р – обр. ГДМ 18626/6; р. Нижняя Тунгуска, Гагарий; вятский ярус, дюкинская свита. Зеленой заливкой показана первичная кора. Тонкий или утолщенный коричневый внутренний контур – ксилема. Внешний контур – граница стебля. Зеленые утолщенные линии вне стебля – листовые влагалища или зубцы. Радиальные линии на фиг. б – зона нарастания нитчатых водорослей на листовое влагалище (?).

Рис. 3.
Корневище (?) Dzergalanella рrynadae: вверху – вид в плане (на плоскости напластования), внизу – вид сбоку (по фотографиям, в натуральную величину); р. Нижняя Тунгуска, Кочумдек-2; вятский (?) ярус, эксинская (?) толща.
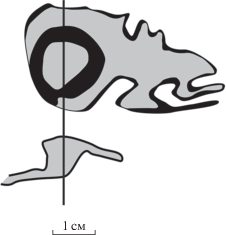
Arthropitys (?) prynadae: Садовников, 1971, табл. 2, фиг. 1–4 ; рис. 1, 2.
Голотип – ГИН, № 3760/1; p. Нижняя Тунгуска, лев. берег в 30 км ниже устья р. Таймура, у о-ва (осередка) Гагарий; верхняя пермь, вятский ярус, дюкинская свита, экозона Quadrocladus pachyphyllum … Prilukiella tomiensis.
Diagnosis (emended). The stem is furrowed (epicycloid), rounded or angular-rounded. The primary cortex is broad. It contains furrow air cavities rounded in cross-section. The stele is rounded or angular-rounded. The core cavity is broad and smooth-walled. There are 70–90 vascular bundles. The thickness of the xylem is about 0.1 mm. Primary xylem is mesarch. There is a poorly developed secondary xylem. The tracheids of the secondary xylem are spiral, stair or porous. The leaves are whorled, about 2 mm wide, with several parallel veins. In the lower part, they coalesce into conical leaf sheaths.
Описание (рис. 2, а–ж; 3). Побеги членистостебельных in situ в анатомической сохранности. Стебель бороздчатый (эпициклоидный), округлый, угловато-округлый или овальный, около 24 мм в поперечнике. Первичная кора широкая (4–5 мм), с округлыми в поперечном сечении ложбиночными воздушными полостями (табл. XIV , фиг. 3). Стела округлая или угловато-округлая (табл. XIV , фиг. 3; рис. 2, а–ж), 12–15 мм в поперечнике. Углы (от трех до восьми) образуются за счет веточных лакун разного размера (до трех в одном сечении). Сердцевинная полость широкая гладкостенная. Проводящих пучков 70–90. Толщина ксилемы около 0.1 мм. Первичная ксилема мезархная. Протоксилема часто разрушается и образуются протоксилемные лакуны. Имеется слаборазвитая вторичная ксилема с многочисленными одно- или двурядными вторичными сердцевинными лучами. Трахеиды вторичной ксилемы спиральные, лестничные, пористые. Первичные сердцевинные лучи широкие, состоят из несколько удлиненных паренхимных клеток (Садовников, 1971). Листья мутовчатые, шириной около 2 мм, с несколькими параллельными жилками, в нижней части срастаются в конические листовые влагалища (рис. 2).
Близ основания стебля от него отходят в одну сторону горизонтальные уплощенные ответвления (корневища?) шириной 11–15 мм с 15–24 проводящими пучками (по-видимому, на каждой стороне, т. е. всего пучков, вероятно, 30–50). Расстояние между проводящими пучками приблизительно 1 мм (рис. 3).
Сравнение. От D. dzergalanensis вид отличается наличием широкой первичной коры с воздушными полостями и нерасчлененными коническими листовыми влагалищами.
Распространение. Р. Нижняя Тунгуска; верхняя пермь, вятский ярус, дюкинская свита, экозона Quadrocladus pachyphyllum … Prilukiella tomiensis (Садовников, 1981, 1987, 1989, 1991, 2003, 2008, 2013а, б, 2014, 2015а, б, 2016, 2017, 2018; Садовников, Орлова, 1990, 1994, 1995, 1997; Sadovnikov, Orlova, 1993; Sadovnikov, 1997).
Материал. Пять экз. из типового местонахождения (ГИН 3760/1; ГДМ НВФ 15323, 18622, 18623, 18625); один экз. из местонахождения Кочумдек-2: р. Нижняя Тунгуска, вятский ярус (?), эксинская (?) толща.
Dzergalanella taimyrica Sadovnikov, sp. nov.
Табл. XV , фиг. 3–4 (см. вклейку)
Dzergalanella merianii: Садовников, 2015а, с. 61–74.
Название вида – по п-ову Таймыр.
Голотип – ГДМ НВФ № 8590, фрагмент побега (Садовников, 2015а, с. 67); Центральный Таймыр, руч. Многовершинный (большой правый приток р. Фадью-Куда в ее верховьях), лев. берег в 8 км от устья (Многовершинный-1); средний–верхний триас, мамоновская свита.
Diagnosis. The stem is up to 50 mm in diameter, elongated, with very long internodes (about 250 mm). They are smooth or with a thin longitudinal ribbing. The leaves are narrow, linear, parallel-edged, 1–4 mm wide, fused into high cylindrical leaf sheaths. The leaves have several parallel veins.
Описание. Фрагменты побегов имеют длину до 250 мм. Стебель удлиненный, с очень длинными междоузлиями (около 250 мм). Они гладкие или с тонкой продольной ребристостью. Диаметр стебля от 6 до 50 мм, преимущественно 10–20 мм. Часто хорошо выражена продольная ребристость (около 12–15 ребер на 5 мм ширины стебля). Нередко встречаются стебли, поверхность которых гладкая или покрыта только тонкими продольными ребрышками (20–35 на 5 мм ширины стебля). Такие стебли в изолированном состоянии можно было бы идентифицировать как Calamoderma Sadovnikov. Но на некоторых образцах видно, что это разные слои одного и того же остатка (табл. XV , фиг. 3). Гладкая поверхность может быть поверхностью внутренней сердцевинной полости стебля. Часто на поверхности стебля, помимо обычных ребер, видны более редкие ребра (от одного до шести на 5 мм ширины стебля). В этом случае, по-видимому, мы наблюдаем поверхность не самого стебля, а листового влагалища из сросшихся нижних частей листьев. Такое явление встречается у нескольких родов членистостебельных, но на каждом листе почти всегда имеется одно ребрышко, отвечающее жилке листа. У таймырских растений в листе несколько одинаковых жилок (табл. XV , фиг. 3, 4 ), что свойственно только роду Dzergalanella Genkina. И на поверхности стебля, и на поверхности листьев помимо обычной ребристости видна и более тонкая, как на гладких стеблях (20–35 на 5 мм ширины стебля). По-видимому, она соответствует рядам клеток эпидермы.
Узлы встречаются крайне редко (обр. ГДМ НВФ № 8716). Это свидетельствует о большой длине междоузлий растения (более 250 мм). Узлы перпендикулярны оси стебля или слабо наклонены к ней. Иногда диаметр узла несколько больше диаметра междоузлия. При переходе через узел ребра обычно слабо изгибаются, но правильного чередования ребер в соседних междоузлиях нет. Четких листовых рубцов нет. Возможно, это связано с тем, что мы видим узлы, “просвечивающие” через листовые влагалища. Листья узкие, линейные, параллельно-крайние, шириной 1–4 мм, сросшиеся в высокие цилиндрические листовые влагалища. В каждом листе имеется несколько параллельных жилок.
В нескольких случаях вместе с остатками Dzergalanella taimyrica встречаются остатки корней, которые могут быть идентифицированы, как Radicites sp. и R. sulcatus Sadovnikov. По-видимому, они принадлежат этим же растениям.
В типовом местонахождении встречен один экземпляр стробила, который может быть идентифицирован как Equisetostachys (табл. XIV , фиг. 2). Стробил эллипсоидальный, длиной 33 мм, шириной 15 мм. Спорофиллы пельтатные. Их расширенные верхние части (около 3–4 мм в поперечнике) округло-полигональные, несут около 12 нечетких радиальных бороздок.
Сравнение. Нерасчлененное цилиндрическое листовое влагалище отличает этот вид от других видов рода.
Распространение. Средний–верхний триас, мамоновская свита Центрального и Восточного Таймыра (Садовников, 2007, 2015а).
Материал. Около 50 экз. из шести местонахождений: Многовершинный-1, Многовершинный-2, Фадью-Куда-9 (басс. р. Фадью-Куда), Тулай-Киряка-1 (возв. Тулай-Киряка), Останцовая-1 (р. Чернохребетная), Цветкова (мыс Цветкова на побережье моря Лаптевых близ устья р. Чернохребетная).
Dzergalanella tunguskana (Sadovnikov, 1971) Sadovnikov, comb. nov., emend. Sadovnikov, emend. nov.
Табл. XIV , фиг. 5 ; табл. XV , фиг. 1, 2
Arthropitys (?) tunguskanus: Садовников, 1971, с. 833–835, табл. I , фиг. 5, 6 , табл. II , фиг. 1–6 .
Голотип – ГИН, № 3760/6; Таймыр, лев. берег р. Нижняя Тунгуска в 30 км ниже устья р. Таймура, у о-ва (осередка) Гагарий; вятский ярус, дюкинская свита, экозона Quadrocladus pachyphyllum … Prilukiella tomiensis.
Diagnosis (emended). Shoots of Аrthrophyta in situ in anatomical preservation. The stem is blunt-tetrahedral to four-furrowed, with a broad smooth-walled core cavity. The primary cortex is thin. The thickness of the xylem is about 0.1 mm. The protoxylem is often destroyed, and protoxylem lacunae are formed in its place. There is a poorly developed secondary xylem with numerous secondary core rays. The tracheids of the primary xylem are spiral, stair or porous. The leaves are whorled, much longer than the internodes, with a well-developed palisade parenchyma. They coalesce into narrowly conical (almost cylindrical) leaf sheaths.
Описание (рис. 2, з–р). Побеги членистостебельных in situ в анатомической сохранности. Имеются пять фрагментов побегов из типового местонахождения диаметром 10–28 мм. На образце, выбранном голотипом, стебель тупо-четырехгранный, около 9 мм в поперечнике, гладкий. Первичная кора шириной около 1 мм. Древесинный цилиндр толщиной около 1 мм. Ширина проводящего пучка около 0.5 мм. Сердцевинные лучи шириной немного менее 1 мм, из двух–трех рядов клеток. Первичная ксилема имеет поперечник около 1 мм. Протоксилема обычно разрушена с образованием каринального канала. Вторичная ксилема толщиной около 1 мм состоит приблизительно из 20 радиальных рядов трахеид. На одном из углов стебля хорошо выражена лакуна (веточная или корневищная) шириной около 4 мм и длиной более 4 мм. На соседнем ребре стебля лакуна той же ширины, но немного короче. На следующем ребре лакуна едва намечается, на четвертом – не выражена. Следовательно, ветви (или корневища) располагались спирально. Имеются три волнисто изгибающиеся листовые влагалища толщиной немного более 1 мм. Расстояние между жилками около 0.4 мм.
На других образцах при том же диаметре стела в поперечном сечении четырехлопастная (обр. ГИН № 3760/4), при меньшем диаметре (т.е., в прикорневой или верхушечной части стебля) – округлая или овальная (обр. ГДМ 18626; рис. 2, з–р).
Сравнение. Угловато-округлый стебель и коническое листовое влагалище отличают этот вид от других видов рода.
Материал. Пять экз.: ГИН № 3760/4, 5, 6; ГДМ 18623, 18626 из типового местонахождения (в соседнем захоронении с побегом Kirjamkenia в полной анатомической сохранности; Садовников, 1983).
ЗАКЛЮЧЕНИЕ
Род членистостебельных Dzergalanella, описанный из Киргизии Генкиной (1966), широко распространен на севере Сибири и установлен в Европе. Для этого рода характерны мутовчатые листья с многочисленными жилками. Сейчас очень часто за поверхность стебля принимают поверхность высоких цилиндрических листовых влагалищ и относят эти растения к другим родам. Роду Dzergalanella принадлежит часть растений, которые ранее в Сибири идентифицировались как Neocalamites и Arthropitys (?).
Список литературы
Генкина Р.З. Ископаемая флора и стратиграфия нижнемезозойских отложений Иссык-Кульской впадины (Северная Киргизия). М.: Наука, 1966. 148 с.
Коломийченко А.В., Садовников Г.Н., Бадамшина Н.О. Особенности разнообразия биоты на рубеже раннего и среднего триаса Таймыра // Биоразнообразие в истории Земли. Тез. докл. 47 сесс. Палеонтол. об-ва при РАН. СПб., 2001. С. 42.
Мейен С.В. Phyllotheca-подобные растения из верхнепалеозойской флоры Ангариды // Теоретические проблемы палеоботаники. М.: Наука, 1990. С. 17–49.
Садовников Г.Н. Окаменелости членистостебельных из тутончанской свиты верхней перми Тунгусского бассейна // Ботан. журн. 1971. Т. 56. № 6. С. 830–836.
Садовников Г.Н. Региональные стратиграфические подразделения верхней перми и нижнего триаса Сибирской платформы и сопредельных районов // Сов. геол. 1981. № 6. С. 74–84.
Садовников Г.Н. Новые данные о морфологии и анатомии рода Kirjamkenia Prynada // Палеонтол. журн. 1983. № 4. С. 76–81.
Садовников Г.Н. К стратиграфии пермотриасовых отложений Тунгусского бассейна // Сов. геол. 1987. № 2. С. 84–89.
Садовников Г.Н. О тутончанском горизонте и вулканогенных пермотриасовых отложениях Тунгусского бассейна // Тр. Ин-та геол. и геофиз. СО АН СССР. 1989. Т. 732. С. 13–19.
Садовников Г.Н. Стратиграфия верхних горизонтов перми и граница перми и триаса в континентальных отложениях Сибири // Междунар. конгр. “Пермская система земного шара”. Тез. докл. Свердловск: Уральск. отд. АН СССР (УрОАН), 1991. С. 74.
Садовников Г.Н. Палеоэкологические причины изменения во времени разнообразия неморских биот // Биоразнообразие в истории Земли. Тез. докл. 47 сесс. Палеонтол. об-ва при РАН. СПб., 2001. С. 86–87.
Садовников Г.Н. Стратиграфия верхнего палеозоя восточного крыла Тунгусской синеклизы // Стратигр. Геол. корреляция. 2003. Т. 11. № 4. С. 41–54.
Садовников Г.Н. Триасовая флора Таймыра в коллекциях Дарвиновского музея // Тр. Гос. Дарвиновского музея. 2007. Вып. 10. С. 206–217.
Садовников Г.Н. О положении “точки глобального стратотипа нижней границы” триаса // Стратигр. Геол. корреляция. 2008. Т. 16. № 1. С. 34–50. https://doi.org/10.1134/S0869593808010036
Садовников Г.Н. К методике полуколичественного и количественного анализа неморских палеоэкосистем палеозоя и мезозоя // Палеонтол. журн. 2011. № 1. С. 97–104.
Садовников Г.Н. Экозоны неморских отложений вишкильского–индского ярусов Средней Сибири // Палеоботанический временник. Приложение к журн. “Lethaea rossica”. 2013а. Вып. 1. С. 70–73.
Садовников Г.Н. Неморские экозоны карбона, перми, триаса и юры севера и центра Евразии // Изв. ВУЗов. Сер. Геол. и разведка. 2013б. № 6. С. 91–94.
Садовников Г.Н. Экозона Quadrocladus pachyphyllum … Prilukiella tomiensis вятского яруса верхней перми // Изв. ВУЗов. Сер. Геол. и разведка. 2014. № 6. С. 6–12.
Садовников Г.Н. Триасовая флора Таймыра. Каталог коллекции Дарвиновского музея. М.: Изд-во ГДМ, 2015а. 157 с.
Садовников Г.Н. Структура экозоны Quadrocladus pachyphyllum … Prilukiella tomiensis вятского яруса верхней перми // Изв. ВУЗов. Сер. Геол. и разведка. 2015б. № 4. С. 8–14.
Садовников Г.Н. Эволюция биома траппового плато Средней Сибири // Палеонтол. журн. 2016. № 5. С. 87–99. https://doi.org/10.7868/S0031031X16050123
Садовников Г.Н. Эволюция растительности и фауны беспозвоночных севера Средней Сибири и сопредельных областей в конце перми и начале триаса // Изв. ВУЗов. Сер. Геол. и разведка. 2017. № 6. С. 5–15. https://doi.org/10.32454/0016-7762-2017-6-7-17
Садовников Г.Н. Экостратоны палеозоя и мезозоя севера и центра Азии // Палеоботанический временник. Приложение к журн. “Lethaea rossica”. 2018. Вып. 3. С. 50–56.
Садовников Г.Н., Базаркина Е.А., Турлова М.А. Экозона Dzergalanella merianii … Borysthenia // Проблемы биохронологии в палеонтологии и геологии. Тез. докл. 48 сесс. Палеонтол. об-ва. СПб., 2002. С. 124–125.
Садовников Г.Н., Орлова Э.Ф. Возраст континентальных вулканогенных отложений севера Средней Сибири // Изв. АН СССР. Сер. геол. 1990. № 3. С. 58–70.
Садовников Г.Н., Орлова Э.Ф. Таймырский ярус – терминальный ярус континентальной перми // Докл. Акад. Наук. 1994. Т. 338. № 5. С. 658–661.
Садовников Г.Н., Орлова Э.Ф. Новое в стратиграфии вулканогенной толщи центральной части Тунгусской синеклизы // Стратигр. Геол. корреляция. 1995. Т. 3. № 1. С. 34–42.
Садовников Г.Н., Орлова Э.Ф. О пограничных отложениях перми и триаса северной и восточной окраин Сибирской платформы // Стратигр. Геол. корреляция. 1997. Т. 5. № 1. С. 14–20.
Brongniart A. Histoire des vegetaux fossiles recherches botaniques et géologiques sur les diversis couches du globe. P., 1828–1838. V. 1, 2.
Brunner H., Kelber K.-P. Eisenerzkonkretionen im württembergisch-fränkischen Unterkeuper - Bemerkungen zum fossilen Environment // Neue Forschungen zur Erdgeschichte von Crailsheim / Ed. Hagdorn H. Stuttgart, 1988. S. 185–205 (Sonderbände Ges. Naturk. in Württemberg. Bd 1).
Halle T.G. Zur Kenntnis mesozoischen Equisetales Schwedens // Kungl. Svenska vet. Acad. Handl. 1908. Bd 43. № 1. S. 1–40.
Kelber K.-P. Die Makroflora des Lettenkeupers // Der Lettenkeuper – Ein Fenster in die Zeit vor den Dinosauriern / Eds. Hagdorn H., Schoch R., Schweigert G. Stuttgart, 2015. S. 51–100 (Palaeodiversity Suppl. Staatl. Mus. Naturk. Stuttgart).
Kelber K.-P., Hansch W. Keuperpflanzen. Die Entraetselungeiner ueber 200 Millionen Jahre alten Flora // Museo. 1995. № 11. 157 s.
Meyen S.V. Phyllotheca-like plants from the Upper Palaeozoic flora of Angaraland // Palaeontogr. Abt. B. 1971. Bd 133. № 1/3. P. 1–33.
Osborn J.M., Taylor T.N. Structurally preserved sphenophytes from the Triassic of Antarctica: vegetative remains of Spaciinodum, gen. nov. // Amer. J. Bot. 1989. V. 76. № 11. P. 1594–1601.
Ryberg P.E., Hermsen E.J., Taylor E.L., Taylor T. N. Development and ecological implications of Dormant Buds in the high-paleolatitude Triassic sphenophyte Spaciinodum (Equisetaceae) // Amer. J. Bot. 2008. V. 95. № 11. P. 1443–1453.
Sadovnikov G.N. Taimyrian stage of the terminal non-marine Permian // Prace Panstw. Inst. Geol. 1997. V. 157. P. 133–136.
Sadovnikov G.N., Orlova E.F. Nonmarine latest Permian stratigraphy and the Permian / Triassic boundary in Siberia // Contribution to Eurasian geology, papers presented at the International Congress on the Permian system of the World, Perm, Russia, 1991. Pt II. Occas. publ. Earth Sci. & resources Inst., Univ. of South Carolina, new ser. 9 (A, B). 1993. P. 119–124.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал