

Палеонтологический журнал, 2022, № 2, стр. 93-108
Юго-Восточная Европа как арена эволюции позвоночных в позднем миоцене
Н. В. Зеленков a, *, Е. В. Сыромятникова a, К. К. Тарасенко a, В. В. Титов b, А. С. Тесаков c
a Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
b Южный научный центр РАН
344006 Ростов-на-Дону, Россия
c Геологический институт РАН
119017 Москва, Россия
* E-mail: nzelen@paleo.ru
Поступила в редакцию 19.04.2021
После доработки 23.04.2021
Принята к публикации 26.04.2021
- EDN: HDOHLN
- DOI: 10.31857/S0031031X22020143
Аннотация
Рассмотрены основные черты фаун наземных позвоночных и морских млекопитающих позднего миоцена Юго-Восточной Европы и их общая динамика в связи с изменением ландшафтно-климатических обстановок. Отмечено, что регион современного Северо-Западного и Северного Причерноморья с прилегающими территориями на протяжении позднего миоцена был областью интенсивного формообразования. Выявленная биогеографическая значимость региона связывается нами с тем, что в позднем миоцене здесь проходила граница между лесным биомом и областью распространения открытых и/или полуоткрытых ландшафтов. Ускоренная морфологическая эволюция в данной переходной (экотонной) зоне связывается с фрагментацией ареалов в условиях выраженной пространственной дифференциации растительных ассоциаций и широкой представленности мозаичных местообитаний на фоне существенных климатических колебаний. Обсуждается эволюционный потенциал периферийных сообществ применительно к позвоночным гиппарионовых фаун Юго-Восточной Европы.
Позднемиоценовый этап эволюции биоты – один из важнейших в кайнозойской истории органического мира. Эта эпоха характеризовалась глобальными экосистемными перестройками в низких и умеренных широтах Евразии, где, благодаря глобальному тренду на похолодание и иссушение климата (Zachos et al., 2001), широкое распространение получили открытые или полуоткрытые ландшафты саванноподобного типа, простиравшиеся в отдельные временные интервалы от Пиренейского п-ова на западе до Восточного Китая на востоке. Населявшие эти ландшафты ассоциации позвоночных, ядром которых можно считать представителей так называемых “гиппарионовых” фаун наземных млекопитающих, также имели самое широкое распространение в позднем миоцене; именно они легли в основу более молодых плиоценовых сообществ, в свою очередь являющихся предшественниками как плейстоценовых, так и ряда современных фаунистических ассоциаций Африки и, отчасти, внутренних регионов Азии (Kaya et al., 2018).
История становления позднемиоценовых саванных фаунистических ассоциаций млекопитающих в общих чертах охарактеризована; представляется, что ядрами формирования этих фаун послужили: с одной стороны – восточно-средиземноморский (греко-иранский), а с другой – центральноазиатский (тунгурский) фаунистические комплексы (Mirzaie Ataabadi et al., 2013). Объединение форм, длительное время развивавшихся в этих двух очагах аридности, и привело к формированию относительно цельных евразийских саванноподобных фаун, которые в преобразованном виде продолжали существовать на территории Евразии вплоть до конца раннего плейстоцена. В то же время таксономический состав гиппарионовых фаун не был однородным и менялся как во времени, так и в пространстве, во многом отражая преобразования локальных ландшафтно-климатических обстановок.
Область обрамления Восточного Паратетиса – один из классических регионов распространения гиппарионовой фауны позднего миоцена. Здесь выделяются как минимум четыре этапа трансформации позднемиоценовых фаунистических сообществ (Короткевич, 1988; Лунгу, 1990). При этом преобразование биоты периферических регионов саванного биома (таких как область современного Северного Причерноморья) рассматривается исключительно в контексте смещения соотношения между автохтонными формами, связанными с более закрытыми лесными местообитаниями, и вселенцами из афро-азиатского саванного ядра – обитателями открытых ксерофитных ландшафтов (Короткевич, 1988; Лунгу, 1990). В конце миоцена (конец туролия, понт/мессиний) на гиппарионовые сообщества Европы оказало влияние расселение североамериканских вселенцев, связанное с периодом похолодания и еще более значительным расширением области распространения открытых ландшафтов (Короткевич, 1988). Увеличение числа вселенцев маркирует эпохи более аридного климата, но вопрос локальной эволюции этих форм в данном регионе обычно не ставится.
Недавнее исследование палеоразнообразия страусов (Aves: Struthionidae) – характерных представителей гиппарионовых фаун и при этом относящихся к ядру саванных ассоциаций – неожиданным образом выявило уникальную роль Северо-Западного Причерноморья в эволюционной истории этой группы (Mikhailov, Zelenkov, 2020). Для Северного Причерноморья и Восточной Европы в целом страусы являются типичными аллохтонными вселенцами, впервые появившимися здесь одновременно с гиппарионами в начале позднего миоцена (валлезии) и просуществовавшими по меньшей мере до раннего плейстоцена. Существование нескольких таксонов страусов в регионе на протяжении миоцена–плейстоцена обычно рассматривалось как результат последовательного вселения форм из анатолийских или центрально-азиатских фаун (Курочкин, Лунгу, 1970; Михайлов, Курочкин, 1988; но см. Бурчак-Абрамович, 1953). Однако было показано, что в одну из подобных волн (в мэотисе) вселение страусов было сопряжено с автохтонной эволюцией, результатом которой стало формирование филогенетической линии, сохранившейся в саванных фаунах до настоящего времени (Mikhailov, Zelenkov, 2020). Впервые появившись в палеонтологической летописи в мэотисе Северо-Западного Причерноморья, страусы современного типа затем расселились по обширным пространствам Азии и Африки, постепенно вытеснив или заместив существовавшие в этих очагах саванного биома более древние линии рода Struthio (Mikhailov, Zelenkov, 2020).
Вопрос о том, насколько выявленная картина эволюции страусов отображает общую биогеографическую значимость региона Северного и Западного Причерноморья в эволюции гиппарионовых фаун, ранее не поднимался. В то же время, обозначенный биогеографический паттерн может иметь более общий характер: в частности, было показано, что формирование общей восточноевропейской пикермийской фауны крупных млекопитающих (мэотис) в значительной степени основано на расширении ареалов именно автохтонных балканских форм (Kostopoulos, 2009). В настоящей статье нами предпринята попытка анализа оригинальных и литературных данных по позвоночным с целью показать, что область современного Западного и Северного Причерноморья, а также Юго-Восточная Европа в целом, могли быть регионом, к которому в позднем миоцене было приурочено интенсивное формообразование. Биогеографическая важность данного региона может быть связана с нестабильностью ландшафтно-климатических и палеогеографических обстановок в условиях экотонной зоны.
В данной статье мы рассматриваем регион северного и северо-западного обрамления Восточного Паратетиса, что в целом соответствует современным Северным Балканам, а также современному региону Западного и Северного Причерноморья (рис. 1). По мере регрессии Восточного Паратетиса в этот регион включался также и Северный Кавказ. Этот регион в целом соответствует северной части выделявшейся ранее восточноевропейской палеопровинции (Tobien, 1967; Лунгу, 1990), или же северной части греко-иранской палеопровинции (Fortelius et al., 1996) по фаунам млекопитающих. Хронологически фаунистическая последовательность региона тесно связана с этапами развития солоноватоводных бассейнов Восточного Паратетиса – сармата (херсония и бессарабия), мэотиса и понта (рис. 2). Датировки границ этих морских региоярусов, а также их корреляция с зонами и веками млекопитающих неогена, составляющими европейскую биохронологическую шкалу, продолжают дискутироваться в литературе. Мы ориентируемся на последние ревизии границ MN зон (Hilgen et al., 2012) и палеогидрологических событий Восточного Паратетиса (Palcu, 2018; Palcu et al., 2019). Комплексы млекопитающих неогена Восточной Европы и их корреляция с региоярусами Восточного Паратетиса синтезированы по Е.Л. Короткевич (1988) и В.А. Несину (2013).
Рис. 1.
Географическое положение основных обсуждаемых в работе местонахождений позднемиоценовых наземных позвоночных. 1 – Бужор, Калфа, 2 – Грицев, 3 – Михайловка-1, 4 – Шкодова Гора; 5 – Морская 2; 6 – Гавердовский, Фортепьянка-2; 7 – Солнечнодольск; 8 – Бешпагир.
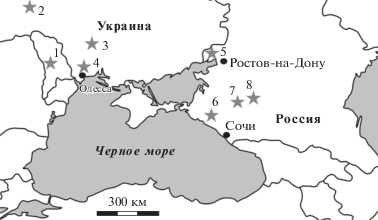
Рис. 2.
Стратиграфическое положение основных обсуждаемых местонахождений позднемиоценовых позвоночных.

Исследование поддержано грантом РНФ 18-74-10081.
ФОРМИРОВАНИЕ ПОЗДНЕМИОЦЕНОВЫХ (ГИППАРИОНОВЫХ) ФАУН НАЗЕМНЫХ ПОЗВОНОЧНЫХ ЕВРАЗИИ
Важнейшее биохронологическое событие, маркирующее начало формирования позднемиоценовых сообществ наземных позвоночных Евразии – вселение американских лошадиных Hipparion s.l., древнейшие европейские представители которых зачастую рассматриваются в составе рода Hippotherium (Woodburne et al., 1996). Появление гиппарионов в составе комплексов наземных позвоночных маркирует начало валлезийского века верхнего миоцена по биохронологии наземных млекопитающих Европы, или бахейского (“Bahean”) века в Китае (Qui et al., 2013). Точное время миграции гиппарионов в Евразию долгое время оставалось предметом дискуссии и до недавних пор чаще всего конвенционально определялось как 10.8 млн лет (Hilgen et al., 2012) – заметно позднее границы серравалия и тортона (т.е., выше границы среднего и верхнего миоцена; 11.63 млн лет). При этом самыми древними из известных евразийских гиппарионов признавались находки из ряда местонахождений зоны С паннона венского бассейна Центральной Европы (Woodburne et al., 1996). Однако недавние работы убедительно показали, что основание зоны С в венском бассейне датируется 11.6 млн лет, т.е., очевидно совпадает хронологически с границей среднего и верхнего миоцена (напр., Lirer et al., 2009; Borgh et al., 2013). При этом древнейшие местонахождения с остатками гиппарионов из зоны С паннонского бассейна (Гайзельберг и Атцельдорф) имеют возраст 11.4–11.0 млн лет, что в целом соответствует возрасту древнейших гиппарионов и в более западных областях – на Иберийском полуострове (Bernor et al., 2017). Таким образом, можно говорить об относительно единовременном появлении и расселении гиппарионов в самом начале позднего миоцена Центральной и Западной Европы.
Поскольку гиппарионы – несомненные американские мигранты (Bernor et al., 2017, 2021), представляется важным проследить их возможное расселение по Евразии, однако данные на этот счет весьма отрывисты. Э.А. Вангенгейм с соавт. (2006) привязывали первое появление гиппарионов в Восточной Европе (в среднем сармате Восточного Паратетиса Молдовы) к отметке 11.8 млн лет, что формально соответствует верхнему серравалию, то есть, концу среднего миоцена. Это могло бы указывать на более раннее появление гиппарионов в Восточной Европе, чем в области Центрального Паратетиса, что в целом соответствует более раннему появлению в этом регионе полуоткрытых стаций саванного типа (Fortelius et al., 1996). В то же время, верхний возрастной предел местонахождений Молдовы с остатками древнейших гиппарионов (таких, как Калфа, Бужор-1 и др.) во многом определялся нижней границей верхнего сармата Восточного Паратетиса, которая в обсуждаемой работе (Вангенгейм и др., 2006) датировалась 9.6 млн лет. Впоследствии нижняя граница верхнего сармата Восточного Паратетиса была оценена на уровне 8.6 млн лет, а позднее – около 9.1 млн лет (Palcu, 2018). Таким образом, возраст молдавских гиппарионов из отложений бессарабия может отвечать возрастному интервалу позднего бессарабия, например, на уровне 10.8 млн лет (Vasiliev et al., 2011) или моложе (Sinitsa, Delinschi, 2016). В этом случае древнейшими азиатскими (и возможно, евроазиатскими) гиппарионами могут оказаться таковые из формации Донсянь (Dongxiang) в бассейне Линся в Китае, датируемые около 11.5 млн лет (Fang et al., 2016; Zhang et al., 2019). В то же время, обычно время появления гиппарионов в Китае принимается как 11.1–10.0 млн лет (Wang et al., 2013).
Хотя появление гиппарионов в фаунах наземных позвоночных, несомненно, является маркером важнейших биотических процессов, обусловивших эволюцию наземной биоты на протяжении последующих нескольких миллионов лет, все же нужно учесть, что становление позднемиоценовых фаун не было единомоментным на всей территории Евразии. Было показано (Fortelius et al., 1996), что в Центральной Европе еще долгое время (по меньшей мере, вплоть до конца мессиния) сохранялись лесные палеообстановки, позволяющие говорить о существовании здесь отдельной палеобиогеографической провинции. В начале позднего миоцена лесные обстановки сохранялись и в других регионах Евразии, таких как Иберийский полуостров или современное Северо-Западное Причерноморье (Молдова). Тем не менее, гиппарионы уже в начале тортона заселили и эти лесные местообитания, но поначалу их появление не вызвало серьезных перестановок в структурах сообществ – так, ранневаллезийские фауны Иберийского полуострова и древние среднесарматские гиппарионовые фауны Молдовы (калфинский фаунистический комплекс) еще имеют всецело среднемиоценовый облик (Лунгу, 1984; Agustí et al., 2001). Другая ситуация могла наблюдаться на южной границе палео-палеарктической палеогеографической провинции в Южном Китае в условиях активно вздымающегося Тибетского плато. В бассейне Линся в самом начале позднего миоцена гиппарионовые и предшествующие им анхитериевые (среднемиоценовые) фауны, по-видимому, сосуществовали во временном отрезке 11.6–11.1 млн лет, будучи разобщены топографически благодаря сложному рельефу (Zhang et al., 2019).
Субсинхронное появление гиппарионов на обширных пространствах умеренных и южных широт Евразии (примерно 11.5–11.1 млн лет), по всей видимости, связано с палеогеографическими изменениями вблизи границы среднего и верхнего миоцена. Широкое распространение гиппарионов как изначальных обитателей полуоткрытых лесостепных стаций в умеренных и отчасти южных областях Евразии (за исключением ориентальной провинции), а также Африки, оказалось возможным, благодаря тому, что уже в конце среднего миоцена в этом регионе сформировался обширный пояс аридности (Flower, Kenneth, 1994), подразумевающий распространение открытых или полуоткрытых местообитаний. Глобальный тренд аридизации в Евразии – во многом определяющая черта именно позднемиоценовых палеообстановок (Fortelius et al., 2006), поэтому вселение гиппарионов в Евразию на границе верхнего миоцена, их последующая позднемиоценовая диверсификация и несомненный эволюционный успех этой группы могут считаться одним из маркеров этой эпохи кайнозоя.
Важным биохронологическим рубежом конца позднего миоцена, отчасти завершающим формирование общего облика гиппарионовых фаун крупных млекопитающих Евразии, можно считать вселение из Северной Америки в Евразию через Берингийский мост верблюдов Paracamelus (Camelidae) и волчьих Eucyon. Представители этих родов быстро расселились по Евразии, занимая ландшафты саванноподобного типа – их остатки известны из мессинийских отложений (MN 13) Испании и Италии (Sotnikova, Rook, 2010; Colombero et al., 2017; Caballero et al., 2021). Уже в мессинское время началась независимая эволюция Camelidae на территории Евразии, и только в плиоцене потомки миоценовых евроазиатских верблюдов, как и предки современных страусов (см. выше), попали в Африку (Titov, Logvinenko, 2006). В Северном Причерноморье и Крыму остатки верблюдов известны из ряда местонахождений таврического фаунистического комплекса, приуроченных к нижнепонтическим отложениям (Одесса, Синявская, Новочеркасск, Мамайские каменоломни). Вероятно, миграция этих животных из Азии в Южную Европу проходила вдоль северного побережья Понтического моря (Titov, Logvinenko, 2006). Остатки Eucyon на территории обсуждаемого региона известны, только начиная с раннего плиоцена (Sotnikova, Rook, 2010), но необходимо отметить, что позднетуролийские фауны крупных животных с территории Восточной Европы изучены еще недостаточно (Короткевич, 1988; Ковальчук и др., 2017).
САМОБЫТНОСТЬ И ЭВОЛЮЦИОННЫЙ ПОТЕНЦИАЛ НАЗЕМНЫХ ФАУН ЮГО-ВОСТОЧНОЙ ЕВРОПЫ
Валлезий. Ярким примером эволюционной значимости обсуждаемого региона являются непосредственно гиппарионы. Обитавшие в валлезии в областях Восточного и Центрального Паратетиса Hippotherium представляются потомками рода Cormohipparion, восходящего еще к среднему миоцену Северной Америки и сохранившемуся в малоизмененном виде в валлезии аридного пояса южной Палеарктики (в частности, Анатолия и Северная Индия; Bernor et al., 2021). В более облесенных обстановках Европы автохтонная дифференциация гиппарионов наблюдается в конце среднего сармата Молдовы и юга Украины, что связывается с преобразованием ландшафтов и постепенным распространением в этом регионе более открытых стаций (Лунгу, 1984). Дифференцированные гиппарионы известны уже в среднесарматских фаунах местонахождений Бужор-2, Грицев и Севастополь (Вангенгейм и др., 2006). Помимо гиппарионов, автохтонная эволюция в конце среднего и, особенно, в позднем сармате обсуждаемого региона регистрируется и для ряда других групп млекопитающих (в частности, безрогих носорогов Aceratherium и Chilotherium; Лунгу, 1984, 1990; Короткевич, 1988). В более западных частях Европы дифференциация гиппарионов не происходила, поскольку там сохранялись консервативные влажно-лесные местообитания (Лунгу, 1984).
Автохтонная эволюция в ряде филогенетических линий и преобразование ландшафтно-экологических условий обусловили формирование фаунистических комплексов наземных млекопитающих, не имеющих аналогов в других регионах. Примечательно сообщество мелких млекопитающих валлезийского местонахождения Грицев (зона МN 9) на западе Украины, характеризующееся преобладанием архаичных среднемиоценовых форм с доминированием насекомоядных на фоне появления ряда характерных позднемиоценовых таксонов грызунов (роды Glis и Muscardinus из сонь, первых Lophocricetus среди Dipodidae и Cricetulodon среди Cricetodontidae; Несин, 2013). При этом отсутствие других характерных позднемиоценовых форм, Muridae и Ochotonidae, потенциально указывающее на относительную древность этой фауны (иногда считается самой древней в валлезии Европы; Несин, 2013), встает в противоречие с продвинутым эволюционным уровнем дифференцированных гиппарионов Грицева (Вангенгейм и др., 2006), т.е., древний облик фауны может быть связан с ее лесным характером и консервативной “валлезийской” фауной лесных местообитаний. Еще одна безаналоговая фауна мелких млекопитающих валлезия Северо-Западного Причерноморья – из раннесарматского (МN 10) местонахождения Михайловка 1 (Несин, 2013). Кластерный анализ сообществ крупных млекопитающих отчетливо указывает на самобытность фаун Украины и Молдовы, датированных биозоной МN 10 (Casanovas-Vilar et al., 2005). Провинциальный характер валлезийских фаун Молдовы проявляется в присутствии ряда эндемичных таксонов (рис. 3, 4), по крайней мере, часть из которых может в действительности быть связана своим происхождением с обсуждаемым регионом.
Рис. 3.
Таксоны земноводных, пресмыкающихся и птиц, для которых самое раннее появление в палеонтологической летописи связано с поздним миоценом Восточной Европы (в т. ч. эндемики).

Рис. 4.
Избранные таксоны млекопитающих, для которых самое раннее появление в палеонтологической летописи связано с поздним миоценом Восточной Европы (в т. ч. эндемики).

Валлезийские фаунистические сообщества птиц пока остаются плохо охарактеризованы не только для Восточной Европы, но и глобально, однако стоит отметить, что в раннем валлезии Молдовы (местонахождения Кишинев и Голбочика) впервые в палеонтологической летописи регистрируются несколько родов водно-болотных птиц (журавль Grus, баклан Microcarbo), два из которых – вымершие эндемики региона: крупная цапля Ardeagrandis и пастушковая птица Mioporphyrula (Зеленков, Курочкин, 2015). Кроме того, для валлезия Молдовы и Украины известно несколько видов птиц, пока не найденных за пределами региона, в т.ч. древнейший в регионе страус Struthio orlovi (Зеленков, Курочкин, 2015). При этом фаунистические сообщества утиных как Центральной (Венгрия), так и Восточной Европы в валлезийское время еще были составлены из среднемиоценовых таксонов, имевших самое широкое распространение по всей Евразии (от Франции до Китая; Zelenkov, 2017; Zelenkov et al., 2018; Зеленков, 2019).
Валлезийские герпетофауны Восточной Европы крайне плохо известны по сравнению с таковыми Западной и Центральной Европы. В герпетокомплексе раннего сармата Молдовы уже появляются элементы открытых пространств, однако в основе комплекса еще лежат среднемиоценовые реликты лесных биотопов (Зерова, 1987). Лучше охарактеризованная герпетофауна Грицева (Украина), вместе с рядом архаичных элементов, таких как хвостатые амфибии Palaeoproteus и удавы Bransateryx, включает в себя безногих ящериц Pseudopus pannonicus (Roček, 2019), которые становятся фоновым видом в Европе, только начиная с конца миоцена.
Позднесарматские местонахождения бериславского териокомплекса (Бешпагир, Спасское) с территории Предкавказья (Ставропольский край) имеют, в общем, сходный между собой облик. Индекс-видами в этих сообществах являлись, наряду с Hipparion giganteum и H. verae, такие формы, как Deinotherium cf. giganteum, Tetralophodon longirostris, Cherolophodon aff. pentelici, Tragocerus cf. frolovi, Procapreolus sp., Cervavitus sp. (Швырева, Титов, 2020; Волокитин и др., 2021). Характерная для местонахождения Грицев ассоциация жвачных Euprox и Protragocerus была распространена и на Северном Кавказе, например, в позднесарматском местонахождении Фортепьянка 2 (Вислобокова, Тарасенко, 2019), которое можно отнести к раннему туролию. Своеобразен состав фаун млекопитающих раннего туролия Северного Кавказа – местонахождения Гавердовский и Волчья Балка. Они коррелируются с концом позднего сармата. Териофауна имеет отчетливый мезофильный облик и в основном связана с местообитаниями закрытого типа. Специфику фаунам придают разнообразие околоводных хвостатых и бесхвостых амфибий и присутствие валлезийских элементов в микротериофауне, например, сонь Paraglirulus (Tesakov et al., 2017).
Туролий. Для второй половины позднего миоцена характерно самое широкое распространение довольно сходных по таксономическому составу саванноподобных фаун в умеренном и субтропическом поясах Палеарктики (напр., Fortelius et al., 1996; Mirzaie Ataabadi et al., 2013; Kaya et al., 2018). В туролийских (Европа) и современным им баодейских (Китай) сообществах полностью отсутствуют среднемиоценовые реликты, интенсивную морфологическую эволюцию претерпевают гиппарионы (Bernor et al., 2021). В частности, на территории Восточной Европы на смену Hipparion giganteum приходит H. moldavicum (Крахмальная, 1996).
В целом, туролийские фауны юга Восточной Европы имеют значительное сходство с таковыми Балкан и Греции, на что указывают, например, многочисленность гребнезубых мастодонтов и дейнотериев, а также наличие зайцеобразных Prolagus. Помимо видов и родов, имеющих большие транспалеарктические ареалы, присутствие лофодонтных тушканчиков Sibirosminthus, полевкозубых хомяков и верблюдов Paracamelus указывает на возросшее азиатское влияние на сообщества северного побережья мэотического и понтического бассейнов Восточного Паратетиса (Titov, Tesakov, 2013; Титов и др., 2017). Ряд характерных таксонов крупных млекопитающих из греко-иранской палеофаунистической провинции (Gazella aff. pigrimi, ?Nisidorcas, Tragoportax gaudryi, Protoryx и Palaeoryx) на рубеже валлезия и туролия или в раннем туролии проникают даже в облесенную Центральную Европу (Вислобокова, 2006).
В последние годы получены достаточно представительные данные по развитию туролийских ассоциаций наземных позвоночных (зоны MN 11–12) на территории Приазовья и Предкавказья (Титов и др., 2006; Подвинцев и др., 2016; Tesakov et al., 2017). Они дополняют имеющиеся сведения о белкинском и черевычанском фаунистических комплексах, описанных на основании материалов из местонахождений с территории Западного Причерноморья. Но если представления о сходстве фаун млекопитающих этой эпохи на обширных пространствах Евразии давно разработаны, то картина палеобиогеографии других групп только начинает проясняться.
Так, материалы по птицам отчетливо указывают на отличие туролийских фаун птиц от таковых валлезия (Zelenkov, 2016). Примечательно появление в туролийских фаунах Евразии (и Восточной Европы, в частности) ряда саванных таксонов птиц – таких как марабу, птицы-носороги, трехперстки (Mlíkovský, 2002; Boev, Kovachev, 2007; Zelenkov et al., 2016). При этом обмен между Африкой и Евразией в туролии заметно сокращается по сравнению с валлезием (Bibi, 2011), поэтому проникновение саванных элементов в сообщества Северного Причерноморья, очевидно, связано с расширением ареалов форм, обитавших в анатолийском и центральноазиатском очагах аридности. Саванные элементы авифауны, такие как жаворонковые трехперстки Ortyxelos, в настоящее время населяющие сахельскую зону в Африке, оказываются широко распространенными в Евразии и даже проникают в Центральную Европу (Zelenkov et al., 2016). Показательно широкое распространение сходных форм страусов (исходя из строения скорлупы яиц) в эту эпоху – от Иберийского п-ова на Западе до Восточного Китая (Mikhailov, Zelenkov, 2020).
Туролийские фауны амфибий и рептилий неоднородны и отличаются высоким экологическим и таксономическим разнообразием. В сравнении с таковыми валлезия, они в целом указывают на большую аридность, однако ряд регионов благоприятствовал влаголюбивым формам. Появление болотных черепах рода Emys в начале туролия (MN 11) в Северном Причерноморье привело к их активному формообразованию (четыре отдельные формы), успешному расселению по территории Восточной Европы и, в последующем, распространению по территории Центральной и Западной Европы в плио-плейстоцене. В мэотисе Северного Причерноморья появляются амфисбены рода Blanus, которые отличаются от миоценовых и всех современных представителей восточноевропейских амфисбен комплекса Blanus strauchi. К концу туролия герпетокомплексы Северного Причерноморья приобретают яркие черты аридности и своеобразия. Высокое разнообразие ящериц Lacertidae в позднем миоцене (MN 13) Предкавказья (местонахождение Солнечнодольск) позволяет оценивать этот регион как наиболее важный в становлении отдельных линий этой группы рептилий. Так, отсюда известны древнейшие находки ящериц Lacerta trilineata и мабуидных сцинков Heremites vittatus (Čerňanský, Syromyatnikova, 2019, 2021), которые ныне являются типичными обитателями открытых ландшафтов Юго-Восточной Европы и Ближнего Востока. При этом присутствие таких архаичных элементов, как Anguis cf. rarus и Ophisaurus cf. spinari, делает эту фауну уникальной по своему составу (Čerňanský et al., 2020). Появление здесь первых Pelobates praefuscus, по-видимому, могло дать начало радиации чесночниц современного типа P. fuscus (Syromyatnikova, 2019).
Несмотря на общее сходство туролийских фаун наземных позвоночных на обширных пространствах Южной Евразии, накапливаются палеонтологические данные, все полнее свидетельствующие о биогеографической важности обсуждаемого региона. Возможно, наиболее показательным примером является уже отмеченное выше формирование в западных окраинах Восточного Паратетиса филогенетической линии современных страусов: строение поровой системы скорлупы причерноморских позднемиоценовых страусов отражает усиленную специализацию к гнездованию в особо аридных условиях (Mikhailov, Zelenkov, 2020). Формирование этого морфотипа, очевидно, приурочено к одному из эпизодов аридизации климата в регионе и может быть связано с началом туролия (биозона MN 11). Именно в это время (~8.6 млн лет), согласно недавнему исследованию (Palcu et al., 2019), в регионе Восточного Паратетиса начинается эпизод значительной позднехерсонской аридификации, приведшей к существенной регрессии херсонского бассейна, пик которой приходится на 7.65 млн лет. Нестабильность палеоклиматических обстановок в регионе в туролии показана для территории Болгарии: относительно аридные фазы отмечены для биозоны MN 11 (7.8–8.2 млн лет), и еще одна (более выраженная) – для биозоны MN 12 (7.4–6.8 млн лет; Böhme et al., 2018). В Южной Украине к первому временному интервалу относится фауна мелких млекопитающих Фрунзовки 2, в которой впервые для региона регистрируются тушканчики Allactaga (Paralactaga) и полевкозубые хомяки Ischymomys. Показательно, что данный комплекс не имеет аналогов в Европе (Несин, 2013). В это же время в Греции впервые регистрируется ряд характерных представителей т. н. пикермийской позднетуролийской фауны крупных млекопитающих, сформировавшейся в результате слияния балканских и анатолийских фаун и существовавшей в интервале 7.3–7.2 млн лет (Kostopoulos, 2009). Очень важно, что формирование пикермийской биоты происходит в основном за счет распространения ареалов балканских видов на юг, но не наоборот (Kostopoulos, 2009). Это указывает на повышенную конкурентоспособность эволюционировавших на северо-западных окраинах Восточного Паратетиса форм млекопитающих и соответствует картине эволюции страусов (сформировавшийся в обсуждаемом регионе страус позднее заселил Евразию и Африку). Распространение пикермийской биоты на Балканах совпадает с резким возрастанием в этом регионе доли приспособленных к аридным условиям С4 трав и появлением принесенной из Сахары пыли (Böhme et al., 2017).
По меньшей мере, у ряда других членов туролийских фаун в условиях нестабильности климата и возрастания аридности на западных границах Восточного Паратетиса интенсифицируется морфологическая эволюция. Так, М. Беме с соавт. (Böhme et al., 2018) отмечают наличие выраженной морфологической эволюции у гиен на территории Болгарии в аридные интервалы туролия. Предполагается, что в Юго-Восточной Европе также могли иметь место важнейшие этапы становления Hominini (Böhme et al., 2017; Fuss et al., 2017). Древнейшим представителем клады Hominini может оказаться Graecopithecus freybergi, известный из отложений возрастом около 7.2 млн лет из Греции и Болгарии, и проявляющий сходство с более ранними валлезийскими греческими (но, что важно – не с анатолийскими) Homininae (Spassov et al., 2012).
Из обсуждаемого региона описано несколько специализированных туролийских таксонов млекопитающих неясного происхождения – весьма вероятно, что они также могут представлять результат автохтонной эволюции в условиях климатических колебаний в конце позднего миоцена. Так, ископаемый заяц Alilepus lascarewi из туролия (MN 12) Украины отличается от всех миоценовых и плиоценовых представителей рода Старого Света (Čermák et al., 2015). Обсуждая необычную морфологию вида, авторы предполагают, что он может представлять собой азиатского мигранта, при этом никаких сходных форм в Азии не известно, в то время как древнейшие несомненные находки рода Alilepus в Европе приурочены именно к низам MN 11 зоны в Молдове (местонахождение Кайнары) и на Украине (местонахождение Палиево). Древнейшие Alilepus найдены и на Северном Кавказе (Гавердовский). Сходная эволюционная история предполагалась и для Vasseuromys tectus из туролия (MN 11–12) Украины – самого специализированного вида этого рода – перед его вымиранием (Sinitsa, Nesin, 2018). Сони рода Vasseuromys – характерный элемент миоценовых микротериофаун Центральной и Восточной Европы и Малой Азии. В позднем туролии Приазовья (низы MN 13) в палеонтологической летописи впервые появляются зайцы рода Hypolagus, получившего широкое распространение в Евразии в плиоцене – раннем плейстоцене (Averianov, 1996; Čermák, 2009). C поздним миоценом юга Восточной Европы связан очаг формообразования и эволюции оленей из рода Cervavitus (Petronio et al., 2007). Автохтонное восточноевропейское происхождение имеет и один из древнейших представителей филогенетической линии большерогих оленей – эндемичный род и вид Neomegaloceros gracilis, известный только из позднего миоцена (предположительно MN 13) юга Украины (Короткевич, 1988; Вислобокова, 2011).
Данные по птицам также подтверждают биогеографическую значимость обсуждаемого региона в туролийское время. Из Западного и Северного Причерноморья описан ряд таксонов птиц, пока не известных из других областей, в том числе несколько форм родового уровня (гусь Proanser major, пастушковые птицы Crexica, Miohypotaenidia, дрофа Miootis, ископаемый род куликов Cherevychnavis и древнейший в палеонтологической летописи представитель бекасов Gallinago azovica, врановая птица Miopica), а также ряд эндемичных видов, как водоплавающих, так и наземных, в том числе нелетающий эргильорнитид Urmiornis ukrainus (Зеленков, Курочкин, 2015; Zelenkov, Panteleyev, 2015; Bochenski et al., 2019). Примечательно, что фауна пастушковых птиц позднего туролия Северного Причерноморья кардинально отличалась на родовом уровне от таковой Центральной Европы (Венгрии; Zelenkov et al., 2017). Фауна наземных птиц также была не однородна – если в мэотисе Украины существовали продвинутые эргильорнитиды Urmiornis, то в континентальной Греции в это время жили более архаичные Amphipelargus (Zelenkov et al., 2016). Высокое таксономическое разнообразие отмечается и для страусов региона, хотя географическое и временное распространение отдельных форм пока остается неясным (Mikhailov, Zelenkov, 2020). Мелкие фазановые птицы из туролия Приазовья первоначально были отнесены к “Plioperdix hungaricus”, известному из одновозрастных отложений Центральной Европы (Zelenkov, Panteleyev, 2015), однако новые материалы и проведенная ревизия (Зеленков, Горобец, 2020) указывают на родовую обособленность приазовской формы.
МОРСКИЕ МЛЕКОПИТАЮЩИЕ
Сообщества морских млекопитающих Восточного Паратетиса в позднем миоцене приурочены к мелководным участкам бассейна (рис. 5). Некоторые местонахождения связаны с мшанково-биогермовыми рифами в пределах лагун: например, Майкоп, Полевое и Мелек-Чесме (Tarasenko, 2020; Тарасенко, Пахневич, 2020; Тарасенко и др., 2020). Другие связаны с шельфовой зоной и характеризуются значительной окатанностью материала: Хомутово, Желтокаменка, Золотая балка (Koretsky, Rahmat, 2013). Биогеографическая важность данного региона связана с разнообразием условий в мелководных водоемах – от заливов и лагун до полностью опресненных изолированных участков (Palcu et al., 2019).
Рис. 5.
Основные местонахождения морских млекопитающих в миоцене Восточного Паратетиса: 1– Юркино; 2 – Приморское; 3 – Узунлар; 4 – Тарханкут; 5 – Фортепьянка 2; 6 – Полевое; 7 – Беспутка; 8 – Пеклы; 9 – Эльдар; 10 – Первомайский карьер; 11– Желтокаменка; 12 – Грицев; 13 – Майкоп-2.

В позднемиоценовое время Восточный Паратетис стал местом интенсивной диверсификации цетотериевых китов, особенно на границе бессарабия и херсония (вторая половина валезия), что подтверждается находками представителей различных родов цетотериин на территории Кавказа, Крыма и сопредельных территорий (Мчедлидзе, 1964; Тарасенко, Лопатин, 2012; Тарасенко, 2014; Gol’din, Startsev, 2014; Подвинцев и др., 2016; Тарасенко и др., 2020). Очевидно, развитие в позднем миоцене на севере Восточного Паратетиса мелководного Скифского шельфа с образованием многочисленных биогенных рифов обусловило разнообразие условий и возможную изоляцию для отдельных групп цетотериевых китов (Popov et al., 2004). Широкое распространение в Восточном Паратетисе получили цетотериины, в то время как герпетоцетины более характерны для Западного Паратетиса. Систематическое положение цетотериид из Восточного Паратетиса, ранее отнесенных к герпетоцетинам (Тарасенко, Лопатин, 2012), в настоящий момент пересматривается (Тарасенко и др., 2018). Эндемизм данного региона, скорее всего, связан с наметившейся еще в раннем олигоцене тенденцией – образованием перешейка в районе Фракийского бассейна и отделением восточного Средиземноморского бассейна на юго-западе (Popov et al., 2004). Для Средиземного моря отмечаются довольно редкие находки представителей Cetotheriidae (верхний миоцен; Италия), что подчеркивает очевидную редкость семейства в этом бассейне и может отражать местную конкуренцию с экологически схожими серыми китами – это, в свою очередь, может объяснить географическую изоляцию и разрозненность находок, казалось бы, эндемичных цетотериев в Центральном Паратетисе (Collareta et al., 2021). Для Восточного Паратетиса находки позднемиоценовых ископаемых серых китов пока неизвестны.
Считается, что полуморской замкнутый Сарматcкий бассейн лишь эпизодически вступал в связь со Средиземноморьем; Эвксино-Каспийский бассейн сообщался с Дакийским водоемом, который через узкий Транскарпатский пролив был связан с Паннонским бассейном Западного Паратетиса. На северо-западе Западно-Черноморская депрессия была отделена от Карпатского бассейна зоной шельфов и участками суши (Popov et al., 2004). Изолированность Сарматского бассейна от Средиземноморья и Западного Паратетиса, его опреснение во многом способствовали диверсификации цетотериевых китов в Восточном Паратетисе.
Представители Phocidae были широко распространены в позднем миоцене в пределах Паратетиса (Koretsky, 2001). Считается, что тюлени Паратетиса были приспособлены к богатой кислородом проточной воде мелководной зоны литорали; они были хорошо адаптированы к температурам речных систем (Koretsky, 2001). Адаптации позднемиоценовых тюленей позволили им широко распространиться на территории мелководных заливов и рифовых островов в пределах восточной части Восточного Паратетиса.
ОБСУЖДЕНИЕ
Приведенный выше обзор показывает, что по мере прогрессирующего иссушения климата и распространения открытых ландшафтов в позднемиоценовое время регион современного Северного и Западного Причерноморья не только заселялся мигрантами из более древних евроазиатских очагов аридности (таких, как Анатолия или Центральная Азия), но для ряда позвоночных также служил и областью автохтонной эволюции и диверсификации. Биогеографическая значимость данного региона может в значительной степени объясняться его пограничным положением по отношению к основным ареалам распространения биомов в миоценовое время. Так, в конце среднего миоцена и в самом начале позднего миоцена (ранний–средний сармат Восточного Паратетиса) области современного Северо-Западного Причерноморья (Молдова и Юго-Западная Украина) в основном еще были покрыты лесами, включающими значительное число субтропических элементов, однако на востоке Украины в это время уже были широко распространены открытые злаково-разнотравно-маревые травянистые ассоциации (Velichko et al., 2005; Syabryaj et al., 2007). Роль открытых стаций возрастает в херсонии (верхний сармат Восточного Паратетиса), когда на юге и востоке Украины уже появляются первые травянистые ковыльные протостепи и семиаридные полупустынные ассоциации (Syabryaj et al., 2007). В конце миоцена на территории, прилегающей с севера к Восточному Паратетису, зона преимущественно открытых ландшафтов простиралась на восток от низовьев Днепра и включала в себя Северное Приазовье, низовья Дона, районы Закаспия и южные районы р. Урал. Восточные и юго-восточные границы этой зоны находились в Центральной Азии (Семененко, Тесленко, 1994). Во время эпизода возросшей аридности климата в конце херсония–начале мэотиса отдельные ксерофитные степные элементы травянистой флоры регистрируются заметно юго-западнее – на территории Нижнедунайской низменности (юг Румынии), хотя в данном регионе широколиственные лесные ассоциации оставались доминирующим типом растительности вплоть до самого конца миоцена, а открытые ландшафты получили широкое распространение на равнинах к югу от Карпат только в понтическое время (Casas-Gallego et al., 2021). В Центральных Балканах (запад Болгарии) смешанные мезофитные леса, произраставшие в условиях теплого и достаточно гумидного климата, доминировали даже в самом конце миоцена (понт; Ivanov et al., 2021). В Карпатах на западе Украины лесные биомы (бореальных хвойных и широколиственных лесов) также сохранялись вплоть до самого конца миоцена (Syabryaj et al., 2007).
Таким образом, на территории обсуждаемого в данной работе региона Юго-Восточной Европы на протяжении всего позднего миоцена проходила граница между лесными биомами и областью распространения открытых ландшафтов степного и/или саванного типа. Положение этой границы менялось со временем в зависимости от климата, при этом палеоботанические данные свидетельствуют о том, что проникновение открытых ландшафтов на запад сопровождалось пространственной дифференциацией растительных ассоциаций (Syabryaj et al., 2007; Casas-Gallego et al., 2021). В результате этой дифференциации возникали мозаичные полуоткрытые местообитания, существование которых неоднократно реконструировалось предыдущими исследователями (Короткевич, 1988; Лунгу, 1990; Несин, 2013). Мозаичность распространенных в данной области биоценозов позволяет считать регион Северного и Западного Причерноморья переходной (экотонной) зоной между лесной центрально-европейской и саванной греко-иранской биогеографическими провинциями. Прикарпатские (Грицев) и северокавказские местонахождения (Гавердовский, Волчья Балка) показывают большую роль мезофитных закрытых местообитаний.
Преобразования ландшафтно-климатических обстановок, обусловленные изменениями климата, могли играть определяющую роль в формировании условий для исторической эволюции фаун в обсуждаемой области. В связи с этим, важно отметить, что для позднего миоцена, в отличие от предшествующей эпохи, характерны выраженные климатические флуктуации, явно проявляющиеся в локальных ландшафтно-климатических обстановках Юго-Восточной Европы и определяющие динамику локальных растительных ассоциаций (Ivanov et al., 2011; Palcu et al., 2019). Роль климатических колебаний и связанных с ними изменений растительности в преобразовании фаун была убедительно проиллюстрирована на примере четвертичных копытных млекопитающих Капской области в Южной Африке (Faith, Behrensmeyer, 2013). Эта работа показала, как динамика фаун связана с вымиранием и миграцией видов в соответствии с географическим распространением отдельных растительных ассоциаций.
Диверсификация и морфологическая эволюция отдельных представителей фауны наземных позвоночных в обсуждаемом регионе в значительной степени может быть связана с викариацией популяций предковых форм, обусловленной фрагментированностью биотопов. Именно фрагментация популяций рассматривается как основной фактор формообразования, объясняющий пульсирующий характер динамики фаун млекопитающих в позднем кайнозое (Vrba, 2005). В области Северного и Северо-Западного Причерноморья фрагментация популяций наземных позвоночных (за исключением летающих птиц) могла дополнительно обеспечиваться флуктуациями береговой линии Восточного Паратетиса, испытавшего во время позднего миоцена ряд значительных трансгрессий и регрессий, создававших условия для формирования крупных островов (Попов и др., 2010; Palcu et al., 2019). Эта географическая нестабильность морских бассейнов дополнительно создавала условия для диверсификации морских млекопитающих и водно-болотных птиц (именно к данному региону приурочено первое появление в палеонтологической летописи ряда вымерших и современных родов).
Биогеографическая значимость обсуждаемого региона определяется не только диверсификацией как таковой, но и эволюционным и экологическим успехом форм, сформировавшихся в экотонной зоне. Яркой иллюстрацией последнего представляются обсуждаемые выше страусы и ряд представителей пикермийской фауны крупных млекопитающих. Как было показано Д. Костопулосом (Kostopoulos, 2009), сформировавшиеся в континентальной Греции в раннемэотическое время таксоны крупных млекопитающих затем, при объединении фаун в конце мэотиса, распространили свои ареалы, по меньшей мере, в анатолийскую область – несмотря на то, что на территории последней существовал более древний очаг распространения открытых ксерофитных ландшафтов. Формирование общей греко-иранской пикермийской биоты, таким образом, осуществилось преимущественно за счет обогащения древних фаун аридных ландшафтов вселенцами, сформировавшимися позднее на периферии распространения открытого биома. Важная роль периферийных регионов в становлении фаунистических сообществ климаксных биоценозов подтверждается и изучением истории формирования современных фаун: она недавно была показана на примере богатой (около 1000 современным видов) неотропической радиации кричащих воробьиных птиц (Tyranni; Harvey et al., 2020). Эта группа наиболее разнообразна в тропических регионах Южной Aмерики с теплым климатом и со стабильными ландшафтно-климатическими условиями – однако в этих областях идет лишь накопление таксонов, в то время как их интенсивная диверсификация приурочена к маргинальным регионам с более холодным и сухим климатом и нестабильными ландшафтно-климатическими условиями (Harvey et al., 2020).
Эволюционный успех представителей периферийных сообществ в общем виде может иметь следующее объяснение. Маргинальные фауны, ввиду неоднородности ландшафтно-климатических условий, несомненно, включают меньшее число специализированных видов, по сравнению с ядрами фаунистических ассоциаций. Это определяет иной характер экологических взаимоотношений, ослабляет конкуренцию между специалистами и может способствовать диверсификации. В то же время, обедненный состав маргинальных фаун позволяет рекрутизировать формы из географически смежных, но экологически отличных местообитаний. Эти факторы сами по себе обуславливают ускоренную эволюцию и способствуют появлению новых форм. В случае с открытыми аридными ландшафтами можно говорить о том, что периферийные области их распространения, такие как Северо-Западное Причерноморье, представляют собой несомненные вторичные очаги морфологической дифференциации и эволюции. Первоначально происходило заселение этих областей древними обитателями открытых ландшафтов, сформировавшихся еще в олигоцене – среднем миоцене. Дальнейшая морфологическая эволюция, по крайней мере, некоторых таксонов происходила уже в позднем миоцене в условиях усиливающейся аридизации климата и фрагментации местообитаний на периферии основной области распространения биома. Формы, эволюционировавшие в данных условиях, таким образом, могут рассматриваться как “эвадаптивные” (Расницын, 1986) – являющиеся результатом более длительной эволюции, и поэтому более приспособленные, по сравнению с “инадаптивными” таксонами, чье эволюционное становление произошло в условиях быстрого приспосабливания к изменяющимся условиям на основе неспециализированных предковых морфотипов.
Список литературы
Бурчак-Абрамович Н.И. Ископаемые страусы Кавказа и юга Украины. Баку: Изд-во АН Азерб. ССР, 1953. 204 p.
Вангенгейм Э.А., Лунгу А.Н., Тесаков А.С. Возраст нижней границы валлезия (континентальный миоцен Европы) // Стратигр. Геол. корреляция. 2006. Т. 14. № 6. С. 81–91.
Вислобокова И.А. Ассоциации жвачных восточноальпийских экосистем в миоцене // Палеонтол. журн. 2006. № 4. С. 85–95.
Вислобокова И.А. История развития и географического распространения большерогих оленей (Cervidae, Megacerini) // Палеонтол. журн. 2011. № 6. С. 75–90.
Вислобокова И.А., Тарасенко К.К. Находки жвачных (Artiodactyla, Ruminantia) и новые данные по стратиграфии позднемиоценового местонахождения Фортепьянка 2 (Северный Кавказ, Республика Адыгея) // Палеонтол. журн. 2019. № 6. С. 105–110.
Волокитин В.В., Швырева А.К., Титов В.В. Бугорчатозубые мастодонты (Gomphotheriidae) из позднего сармата Ставрополья // Теоретические и прикладные аспекты палеонтологии. Матер. LXVII сесс. Палеонтол. об-ва при РАН. СПб.: ВСЕГЕИ, 2021. С. 102–103.
Зеленков Н.В. Эволюция утиных (Aves: Anatidae s.l.) Евразии в кайнозое // Журн. общ. биол. 2019. Т. 80. № 5. С. 323–333.
Зеленков Н.В., Горобец Л.В. Ревизия Plioperdix (Aves: Phasianidae) из плио-плейстоцена Украины // Палеонтол. журн. 2020. № 5. С. 90–101.
Зеленков Н.В., Курочкин Е.Н. Класс Aves // Ископаемые рептилии и птицы. Часть 3 / Ред. Курочкин Е.Н., Лопатин А.В., Зеленков Н.В. М.: ГЕОС, 2015. С. 86–290.
Зерова Г.А. Местонахождения позднемиоценовых–раннеплейстоценовых ящериц и змей Украины // Тр. Ин-та зоол. АН УССР. 1987. № 12. С. 12–18.
Ковальчук А.Н., Горобец Л.В., Зеленков Н.В. и др. Позвоночные из понтических отложений Шкодовой горы (Северо-Западное Причерноморье, верхний миоцен) // Палеонтол. журн. 2017. № 4. С. 78–92.
Крахмальная Т.В. Гиппарионовая фауна древнего мэотиса Северного Причерноморья. Киев: Наук. думка, 1996. 225 с.
Короткевич Е.Л. История формирования гиппарионовой фауны Восточной Европы. Киев: Наук. думка, 1988. 164 с.
Курочкин Е.Н., Лунгу А.Н. Новый страус из среднего сармата Молдавии // Палеонтол. журн. 1970. № 1. С. 118–126.
Лунгу А.Н. Гиппарионовая фауна среднего сармата Молдавии (копытные млекопитающие). Кишинев: Штиинца, 1984. 157 с.
Лунгу А.Н. Ранние этапы развития гиппарионовой фауны континентального Паратетиса. Дис. … докт. геол.-мин. наук. Тирасполь, 1990. 444 с.
Михайлов К.Е., Курочкин Е.Н. Скорлупа яиц ископаемых Struthioniformes из Палеарктики и ее место в системе представлений об эволюции Ratitae // Ископаемые рептилии и птицы Монголии / Ред. Е.Н. Курочкин. М.: Наука, 1988. С. 43–64.
Мчедлидзе Г.А. Ископаемые китообразные Кавказа. Тбилиси: Мецниереба, 1964. 144 с.
Несин В.А. Неогеновые Murinae (Rodentia, Muridae) Украины. Сумы: Универ. книга, 2013. 174 с.
Подвинцев В.Г., Куршаков С.В., Малышкина Т.П. и др. Новое гетерогенное местонахождение морских и наземных позвоночных позднего миоцена Гладковское (Краснодарский край) // Матер. LXII Юбил. сесс. Палеонтол. об-ва при РАН. СПб., 2016. С. 267–268.
Расницын А.П. Инадаптация и эвадаптация // Палеонтол. журн. 1986. №1. С. 4–7.
Попов С.В., Антипов М.П., Застрожнов А.С. и др. Колебания уровня моря на северном шельфе Восточного Паратетиса в олигоцене–неогене // Стратигр. Геол. корреляция. 2010. Т. 18. № 2. С. 99–124.
Семененко В.Н., Тесленко Ю.В. Геологические события в Восточном Паратетисе на рубеже миоцена и плиоцена // Геол. журн. 1994. № 1. С. 58–68.
Тарасенко К.К. Новые роды усатых китов (Cetacea, Mammalia) из миоцена Северного Кавказа и Предкавказья: 3. Zygiocetus gen. nov. (средний сармат, Адыгея) // Палеонтол. журн. 2014. № 5. С. 99–109.
Тарасенко К.К., Коваленко Е.С., Подурец К.М. Новые данные о морфологии костного лабиринта каменистой кости миоценового усатого кита из Чечни – Vampalus sayasanicus // Матер. LXIV сесс. Палеонтол. об-ва при РАН. СПб., 2018. С. 234–235.
Тарасенко К.К., Лопатин А.В. Новые роды усатых китов (Cetacea, Mammalia) из миоцена Северного Кавказа и Предкавказья: 1. Kurdalagonus gen. nov. из среднего–позднего сармата Адыгеи // Палеонтол. журн. 2012. № 5. С. 86–98.
Тарасенко К.К., Лопатин А.В., Старцев Д.Б. Первая находка позднемиоценовых усатых китов Zygiocetus (Cetotheriidae, Mysticeti) в Крыму (местонахождение Мелек-Чесме, Керченский полуостров) // Докл. Акад. наук. 2020. Т. 491. № 1. С. 176–179.
Тарасенко К.К., Пахневич А.В. Формирование строматолитовых биогермов на костях ископаемых китообразных // Биогеография и эволюционные процессы. Матер. LXVI сесс. Палеонтол. об-ва при РАН. СПб.: ВСЕГЕИ, 2020. С. 159–160.
Титов В.В., Тесаков А.С., Данилов И.Г. и др. Первая представительная фауна позвоночных позднего миоцена на юге европейской России // Докл. Акад. наук. 2006. Т. 411. № 5. С. 715–717.
Титов В.В., Тесаков А.С., Сотникова М.В. и др. Позднемиоценовые наземные сообщества позвоночных и моллюсков Предкавказья // Геология и биоразнообразие Тетиса и Восточного Паратетиса. Матер. Всеросс. науч.-практ. конф. / Ред. Попов С.В. Горячий Ключ, 2017. С. 89–93.
Швырева А.К., Титов В.В. Позднесарматская фауна из отложений Бешпагирского карьера (Ставропольский край) // Биогеография и эволюционные процессы. Матер. LXVI сесс. Палеонтол. об-ва при РАН. СПб.: ВСЕГЕИ, 2020. С. 281–282.
Agustí J., Cabrera L., Gracés M. et al. A calibrated mammal scale for the Neogene of Western Europe. State of the art // Earth-Sci. Rev. 2001. V. 52. P. 247–260.
Averianov A. The Neogene rabbit Hypolagus igromovi Gureev, 1964 (Lagomorpha, Leporidae) from southern European Russia // Acta Zool. Cracov. 1996. V. 39. P. 61–66.
Bernor R.L., Göhlich U.B., Harzhauser M., Semprebon G.M. The Pannonian C hipparions from the Vienna Basin // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2017. V. 476. P. 28–41.
Bernor R.L., Kaya F., Kaakinen A. et al. Old world hipparion evolution, biogeography, climatology and ecology // Earth-Sci. Rev. 2021. V. 221: 103784. https://doi.org/10.1016/j.earscirev.2021.103784
Bibi F. Mio-Pliocene faunal exchanges and African biogeography: the record of fossil bovids // PLoS One. 2011. V. 6: e16688. https://doi.org/10.1371/journal.pone.0016688
Bochenski Z., Wertz K., Tomek T., Gorobets L. A new species of the late Miocene charadriiform bird (Aves: Charadriiformes), with a summary of all Paleogene and Miocene Charadrii remains // Zootaxa. 2019. V. 4624. P. 41–58.
Boev Z.N., Kovachev D. Euroceros bulgaricus gen. nov., sp. nov. from Hadzhidimovo (SW Bulgaria) (Late Miocene) – the first European record of Hornbills (Aves: Coraciiformes) // Geobios. 2007. V. 40. P. 39–49.
Böhme M., Spassov N., Ebner M. et al. Messinian age and savannah environment of the possible hominin Graecopithecus from Europe // PLoS One. 2017. V. 12: e0177347. https://doi.org/10.1371/journal.pone.0177347
Böhme M., Van Baak C.G.C., Prieto J. et al. Late Miocene stratigraphy, palaeoclimate and evolution of the Sandanski Basin (Bulgaria) and the chronology of the Pikermian faunal changes // Glob. Planet. Change. 2018. V. 170. P. 1–19.
Borgh M. ter, Vasiliev I., Stoica M. et al. The isolation of the Pannonian basin (Central Paratethys): New constraints from magnetostratigraphy and biostratigraphy // Glob. Planet. Change. 2013. V. 103. P. 99–118.
Caballero Y., Montoya P., Crespo V.D. et al. The autopodial skeleton of Paracamelus aguirrei (Morales 1984) (Tylopoda, Mammalia) from the late Miocene site of Venta del Moro (Valencia, Spain) // J. Iber. Geol. 2021. V. 47. P. 483–500.
Casanovas-Vilar I., Moyà-Solà S., Agustí J., Köhler M. The geography of a faunal turnover: tracking the Vallesian Crisis // Migrations of Organisms. Climate. Geography. Ecology / Ed. Elewa A.M.T. Berlin, Heidelberg: Springer, 2005. P. 247–300.
Casas-Gallego M., Marza A., Tudor E. Palaeovegetation and palaeoclimate evolution during the Late Miocene to Early Pliocene of SE Romania // Geol. J. 2021. V. 56. P. 821–838.
Čermák S. The Plio-Pleistocene record of Hypolagus (Lagomorpha, Leporidae) from the Czech and Slovak Republics with comments on systematics and classification of the genus // Bull. Geosci. 2009. V. 84. P. 497–524.
Čermák S., Angelone Ch., Sinitsa M.V. New late Miocene Alilepus (Lagomorpha, Mammalia) from Eastern Europe – a new light on the evolution of the earliest Old World Leporinae // Bull. Geosci. 2015. V. 90. P. 431–451.
Čerňanský A., Syromyatnikova E.V. The first Miocene fossils of Lacerta cf. trilineata (Squamata, Lacertidae) with a comparative study of the main cranial osteological differences in green lizards and their relatives // PLoS ONE. 2019. V. 14: e0216191. https://doi.org/10.1371/journal.pone.0216191
Čerňanský A., Syromyatnikova E.V. The first pre-Quaternary fossil record of the clade Mabuyidae with a comment on the enclosure of the Meckelian canal in skinks // Pap. Palaeontol. 2021. V. 7. P. 195–215.
Čerňanský A., Syromyatnikova E.V., Jablonski D. The first record of amphisbaenian and anguimorph lizards (Reptilia, Squamata) from the upper Miocene Solnechnodolsk locality in Russia // Histor. Biol. 2020. V. 32. P. 869–879.
Collareta A., Marx F.G., Casati S., et al. A cetotheriid whale from the upper Miocene of the Mediterranean // N. Jb. Geol. Paläontol. Abh. 2021. Bd 301. S. 9– 16.
Colombero S., Bonelli E., Pavia M. et al. Paracamelus (Mammalia, Camelidae) remains from the late Messinian of Italy: insights into the last camels of Western Europe // Histor. Biol. 2017. V. 29. P. 509–518.
Faith J.T., Behrensmeyer A.K. Climate change and faunal turnover: testing the mechanics of the turnover-pulse hypothesis with South African fossil data // Paleobiology. 2013. V. 39. P. 609–627.
Fang X., Wang J., Zhang W. et al. Tectonosedimentary evolution model of an intracontinental flexural (foreland) basin for paleoclimatic research // Glob. Planet. Change. 2016. V. 145. P. 78–97.
Flower B.P., Kennett J.P. The middle Miocene climatic transition: East Antarctic ice sheet development, deep ocean circulation and global carbon cycling // Palaeogeogr., Palaeoclimatol., Palaeoecol. 1994. V. 108. P. 537–555.
Fortelius M., Eronen J., Liu L. et al. Late Miocene and Pliocene large land mammals and climatic changes in Eurasia // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2006. V. 238. P. 219–227.
Fortelius M., Werdelin L., Andrews P. et al. Provinciality, diversity, turnover, and paleoecology in land mammal faunas of the Later Miocene of Western Eurasia // The Evolution of Western Eurasian Neogene Mammal Faunas / Eds. Bernor R.L., Fahlbusch V., Mittmann H.-W. N.Y.: Columbia Univ. Press, 1996. P. 414–448.
Fuss J., Spassov N., Begun D.R., Böhme M. Potential hominin affinities of Graecopithecus from the late Miocene of Europe // PLoS One. 2017. V. 12: e0177127. https://doi.org/10.1371/journal.pone.0177127
Gol’din P.E., Startsev D.B. Brandtocetus, a new genus of Baleen Whales (Cetacea, Cetotheriidae) from the Late Miocene of Crimea, Ukraine // J. Vertebr. Paleontol. 2014. V. 34. P. 419–433.
Harvey M.G., Bravo G.A., Claramunt S. et al. The evolution of a tropical biodiversity hotspot // Science. 2020. V. 370. P. 1343–1348.
Hilgen F.J., Lourens L.J., van Dam J.A. The Neogene Period // The Geologic Time Scale 2012 / Eds. Gradstein F. M., Ogg J.G., Schmitz M., Ogg G. Oxford: Elsevier, 2012. P. 924–956.
Ivanov D., Utescher T., Djorgova N. et al. The late Miocene Beli Breg Basin (Bulgaria): palaeoecology and climate reconstructions based on pollen data // Palaeobiodiv. Palaeoenvir. 2021. V. 101. P. 79–102.
Ivanov D., Utescher T., Mosbrugger V. et al. Miocene vegetation and climate dynamics in Eastern and Central Paratethys (Southeastern Europe) // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2011. V. 304. P. 262–275.
Kaya F., Bibi F., Žliobaitė I. et al. The rise and fall of the Old World savannah fauna and the origins of the African savannah biome // Nature Ecol. Evol. 2018. V. 2. P. 241–246.
Koretsky I.A. Morphology and systematics of Miocene Phocinae (Mammalia: Carnivora) from Paratethys and the North Atlantic Region // Geol. Hung. Ser. Paleontol. 2001. V. 54. P. 1–109.
Koretsky I.A., Rahmat S.J. First record of fossil Cystophorinae (Carnivora, Phocidae): Middle Miocene seals from the northern Paratethys // Riv. Ital. Paleontol. Stratigr. 2013. V. 119. P. 325–350
Kostopoulos D.S. The Pikermian Event: temporal and spatial resolution of the Turolian large mammal fauna in SE Europe // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2009. V. 274. P. 82–95.
Lirer F., Harzhauser M., Pelosi N. et al. Astronomically forced teleconnection between Paratethyan and Mediterranean sediments during the Middle and Late Miocene // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2009. V. 275. P. 1–13.
Mikhailov K.E., Zelenkov N. The late Cenozoic history of the ostriches (Aves: Struthionidae), as revealed by fossil eggshell and bone remains // Earth-Sci. Rev. 2020. V. 208: 103270. https://doi.org/10.1016/j.earscirev.2020.103270
Mirzaie Ataabadi M., Liu L., Eronen J. et al. Continental-scale patterns in Neogene mammal community evolution and biogeography: a Europe-Asia perspective // Fossil Mammals of Asia. Neogene Biostratigraphy and Chronology / Eds. Wang X., Flynn J.J., Fortelius M. N.Y.: Columbia Univ. Press, 2013. P. 629–655.
Mlíkovský J. Cenozoic Birds of the World. Part 1: Europe. Praha: Ninox press, 2002. 406 p.
Palcu D.V. The Dire Straits of Paratethys. Dating, matching and modeling connectivity between the Miocene seas of Eurasia // Utrecht Stud. Earth Sci. 2018. V. 155. P. 1–200.
Palcu D.V., Vasiliev I., Stoica M., Krijgsman W. The end of the Great Khersonian Drying of Eurasia: Magnetostratigraphic dating of the Maeotian transgression in the Eastern Paratethys // Basin Res. 2019. V. 31. P. 33–58.
Petronio C., Krakhmalnaya T., Bellucci L., Di Stefano G. Remarks on some Eurasian pliocervines: characteristics, evolution, and relationships with the tribe Cervini // Geobios. 2007. V. 40. P. 113–130.
Popov S.V., Rӧgl F., Rozanov A.Y. (Eds.) Lithological-Paleogeographic maps of Paratethys: 10 Maps Late Eocene to Pliocene // Cour. Forschungsinst. Senckenb. 2004. V. 250. P. 1–46.
Qui Zh.-X., Qui Zd.-D., Deng T. et al. Neogene land mammal stages/ages of China: toward the goal to establish an Asian land mammal stage/age scheme // Fossil Mammals of Asia. Neogene Biostratigraphy and Chronology / Eds. Wang X., Flynn L.J., Fortelius M. N.Y.: Columbia Univ. Press, 2013. P. 29–90.
Roček Z. A contribution to the herpetofauna from the late Miocene of Gritsev (Ukraine) // C. R. Palevol. 2019. V. 18. P. 817–847.
Rӧgl F. Palaeogeographic considerations for Mediterranean and Paratethys seaways (Oligocene to Miocene) // Ann. Naturhist. Mus. Wien. 1998. Bd 99. P. 279–310.
Sinitsa M.V., Delinschi A. The earliest member of Neocricetodon (Rodentia: Cricetidae): a redescription of N. moldavicus from Eastern Europe, and its bearing on the evolution of the genus // J. Paleontol. 2016. V. 90. P. 771–784.
Sinitsa M.V., Nesin V.A. Systematics and phylogeny of Vasseuromys (Mammalia, Rodentia, Gliridae) with a description of a new species from the late Miocene of Eastern Europe // Palaeontology. 2018. V. 61. P. 679–701.
Sotnikova M., Rook L. Dispersal of the Canini (Mammalia, Canidae: Caninae) across Eurasia during the Late Miocene to Early Pleistocene // Quart. Intern. 2010. V. 212. P. 86–97.
Spassov N., Geraads D., Hristova L. et al. A hominid tooth from Bulgaria: the last prehuman hominid of continental Europe // J. Hum. Evol. 2012. V. 62. P. 138–145.
Syabryaj S., Utescher T., Molchanoff S., Bruch A.A. Vegetation and palaeoclimate in the Miocene of Ukraine // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2007. V. 253. P. 153–168.
Syromyatnikova E.V. Redescription of Pelobates praefuscus Khosatzky, 1985 and new records of Pelobates from the late Miocene–Pleistocene of Eastern Europe // Histor. Biol. 2019. V. 31. P. 888–897.
Tarasenko K.K. New data on sedimentology and taphonomy of the Late Miocene locality Polevoe 1 (Northwest Caucasus, Republic of Adygea) // XXI Int. conf. young geol. Herlany. Kraków: MINERALPRESS, 2020. P. 79–82
Tesakov A.S., Titov V.V., Simakova A.N. et al. Late Miocene (early Turolian) vertebrate faunas and associated biotic record of the Northern Caucasus: geology, taxonomy, paleoenvironment, biochronology // Fossil imprint. 2017. V. 73. P. 383–444.
Titov V.V., Logvinenko V.V. Early Paracamelus (Mammalia, Tylopoda) in the Eastern Europe // Acta Zool. Cracov. 2006. V. 49. P. 163–178.
Titov V.V., Tesakov A.S. Late Miocene (Turolian) vertebrate faunas of the southern European Russia // Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology / Eds. Wang X., Flynn L.J., Fortelius M. N.Y.: Columbia Univ. Press, 2013. P. 536–543.
Tobien H. Subdivision of Pontian mammal faunas // Giorn. Geol. 1967. V. 35. P. 1–5.
Vasiliev I., Iosifidi A.G., Khramov A.N. et al. Magnetostratigraphy and radio-isotope dating of upper Miocene–lower Pliocene sedimentary successions of the Black Sea Basin (Taman Peninsula, Russia) // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2011. V. 310. P. 163–175.
Velichko A.A., Akhlestina E.F., Borisova O.K. et al. East European Plain // Cenozoic climatic and environmental changes in Russia. Geol. Soc. Amer. Spec. Pap. 2005. V. 382. P. 31–66.
Vrba E.S. Mass turnover and heterochrony events in response to physical change // Paleobiology. 2005. V. 31. Suppl. 2. P. 157–174.
Wang X., Flynn L.J., Fortelius M. (Eds.). Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology. N.Y.: Columbia Univ. Press, 2013.
Woodburne M.O., Bernor R.L., Swisher C.C. An appraisal of the stratigraphic and phylogenetic bases for the “Hipparion” datum in the Old World // The Evolution of Western Eurasian Neogene Mammal Faunas / Eds. Bernor R.L., Fahlbusch V., Mittmann H.-W. N.Y.: Columbia Univ. Press, 1996. P. 124–136.
Zachos J., Pagani M., Sloan L. et al. Trends, rhythms, and aberrations in global climate 65 Ma to present // Science. 2001. V. 292. P. 686–693.
Zelenkov N.V. Evolution of bird communities in the Neogene of Central Asia, with a review of the Neogene fossil record of Asian birds // Paleontol. J. 2016. V. 50. № 12. P. 1421–1433.
Zelenkov N.V. The revised avian fauna of Rudabànya (Hungary, Late Miocene) // Paleontologia y Evolucion de las Aves / Eds. Acosta Hospitaleche C., Agnolin F., Haidr N. et al. Buenos-Aires: Mus. Argent. Cienc. Natur., 2017. P. 253–266.
Zelenkov N., Boev Z., Lazaridis G. A large ergilornithine (Aves, Gruiformes) from the Late Miocene of the Balkan Peninsula // Paläontol. Z. 2016. Bd 90. P. 145–151.
Zelenkov N.V., Panteleyev A.V. Three bird taxa (Aves: Anatidae, Phasianidae, Scolopacidae) from the Late Miocene of the Sea of Azov (Southwestern Russia) // Paläontol. Z. 2015. Bd 89. P. 515–527.
Zelenkov N.V., Panteleyev A.V., De Pietri V.L. Late Miocene rails (Aves: Rallidae) from southwestern Russia // Palaeobiodiv. Palaeoenvir. 2017. V. 97. P. 791–805.
Zelenkov N.V., Stidham T.A., Martynovich N.V. et al. The middle Miocene duck Chenoanas (Aves, Anatidae): new species, phylogeny and geographical range // Pap. Palaeontol. 2018. V. 4. P. 309–326.
Zelenkov N.V., Volkova N.V., Gorobets L.V. Late Miocene buttonquails (Charadriiformes, Turnicidae) from the temperate zone of Eurasia // J. Ornithol. 2016. V. 157. P. 85–92.
Zhang W., Appel E., Wang J. et al. New paleomagnetic constraints for Platybelodon and Hipparion faunas in the Linxia Basin and their ecological environmental implications // Glob. Planet. Change. 2019. V. 176. P. 71–83.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал