

Палеонтологический журнал, 2022, № 4, стр. 77-86
Новый вид ризодонтиформной кистеперой рыбы (Sarcopterygii: Rhizodontiformes) из нижнего карбона Московской области
А. Ю. Смирнова *
Московский государственный университет им. М.В. Ломоносова
119991 Москва, Россия
* E-mail: smirula220900@mail.ru
Поступила в редакцию 04.08.2021
После доработки 15.02.2022
Принята к публикации 05.03.2022
- EDN: ZDUWAM
- DOI: 10.31857/S0031031X22040122
Аннотация
Из серпуховского яруса (нижний карбон) Московской области (местонахождение Заборье) описан новый вид рода Rhizodus – R. serpukhovensis, представленный фрагментом нижней челюсти. Ранее черепные остатки этого рода были известны только из визейских отложений Западной Европы и США. Хорошая сохранность позволяет описать такие ранее неизвестные для этого рода признаки, как наличие интеркороноидной ямы и ветви мандибулярного сейсмосенсорного канала на зубной кости. Проведен морфофункциональный анализ зубов ризодонтид, который позволяет предположить, что R. serpukhovensis sp. nov. занимал уровень наивысшего консумента в трофической структуре стешевского сообщества позвоночных, на что указывают также гигантские размеры и мощные челюсти рыбы. Предположительно, объектами ее питания были крупные хрящевые и лучеперые рыбы, а также четвероногие.
ВВЕДЕНИЕ
Ризодонтиформные рыбы широко распространены в каменноугольных отложениях Британских островов и Северной Америки и описаны различными исследователями в XIX в. (Hibbert, 1836; Buckland, 1837; Egerton, Cole, 1837; Owen, 1840). Отсутствие находок целых скелетов или черепов, а также фрагментарность и плохая сохранность материала до сих пор не позволяли хорошо изучить эту группу.
Остатки ризодонтиформных рыб в основном представлены фрагментами нижних челюстей. К Rhizodus hibberti (Agassiz et Hibbert in Hibbert, 1836) (единственному описанному к настоящему времени виду рода Rhizodus) и Rhizodus sp. также относят изолированные элементы посткраниального скелета (Andrews, Westoll, 1970; Johanson et al., 2020). В настоящее время известны находки рода Rhizodus из нижнего карбона Шотландии (асбийский, бригантский, пендлейский и холкерский подъярусы) (Johanson et al., 2020) и миссисипия США (мерамекский подъярус) (Jeffery, 2006).
В 2000 г. впервые в России в Московской обл. И.С. Барсковым и С.А. Дорофеевым был найден уникальный образец – фрагмент нижней челюсти ризодонтиформной рыбы хорошей сохранности. Экземпляр (ПИН, № 2878/581) происходит из слоя 26 разреза местонахождения Заборье (Московская обл.) (рис. 1) по нумерации и описанию И.С. Барскова и Н.В. Горевой (Gibshman et al., 2009). Этот слой относится к нижней (глазеченской) подсвите стешевской свиты и представлен криноидным известняком, глинистым, серовато-коричневым, с пятнами охры, в верхней части вишневого цвета, очень твердым, непористым, с обильными целыми брахиоподами (Eomarginifera lobata Sowerby, Antiquatonia khimenkovi Sarytcheva). Слой относится к зоне Eostaffellina decurta по фораминиферам и к зоне Lochriea cruciformis по конодонтам (Gibshman et al., 2009).
Рис. 1.
Географическое положение местонахождения Заборье на территории Московской области (показано звездочкой).
Стешевский горизонт серпуховского яруса соответствует верхнепендлейскому–нижнеарнсбергскому подъярусам региональной стратиграфической шкалы Западной Европы, и верхней части честерского яруса Северной Америки (Menning et al., 2006; Aretz et al., 2020). Однако некоторые авторы опускают нижнюю границу серпуховского яруса в верхнее визе Западно-Европейской последовательности (Nikolaeva et al., 2020).
Препарирование образца выполнялось с помощью 10%-го раствора уксусной кислоты, что дало возможность сохранить тонкие структуры и отметить некоторые неизвестные ранее детали строения. Образец сфотографирован с напылением хлористым аммонием методом послойной съемки; полученные изображения обработаны с использованием программы Helicon Focus 6 и графического редактора Adobe Photoshop.
В работе используется классификация по Э.И. Воробьевой (2004) и терминология по Э. Ярвику (Jarvik, 1944).
Автор выражает благодарность О.А. Лебедеву (Палеонтологический ин-т им. А.А. Борисяка РАН; ПИН) за предоставленный материал и помощь в работе с ним, Дж. Джеффри (Бристольский ун-т) за ценные комментарии, С.В. Багирову (ПИН РАН) за выполненные им фотографии, а также рецензентам статьи А.О. Аверьянову (ЗИН РАН), А.С. Алексееву (МГУ) и Е.В. Попову (СГУ).
СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
КЛАСС SARCOPTERYGII
ОТРЯД RHIZODONTIFORMES
СЕМЕЙСТВО RHIZODONTIDAE TRAQUAIR, 1881
Род Rhizodus Owen, 1840
Megalichthys: Hibbert, 1836, с. 202 (part.).
Rhizodus: Owen, 1840, с. 12; Young, 1865, с. 38; Miall, 1875, с. 624; Traquair, 1878, с. 657; White, 1927, с. 258; Воробьева, Обручев, 1964, с. 297; Andrews, 1985, с. 67; Long, 1989, с. 1; Jeffery, 2003, с. 256; 2006, с. 113.
Типовой вид – Megalichthys hibberti Agassiz et Hibbert in Hibbert, 1836; нижний карбон Шотландии.
Диагноз. Гигантские ризодонтидные рыбы, длина нижней челюсти около 45 см. Прекороноидная яма отсутствует. Симфизный клык вдвое крупнее короноидных клыков, которые, в свою очередь, в два раза крупнее маргинальных зубов. Поверхность дентина короноидных клыков без скульптуры, клыки с гладкими режущими кромками на передней и задней сторонах, овальные в сечении. Краевые зубы на зубной кости округлые в сечении, слегка изогнуты; некоторые почти прямые. Мандибулярная ветвь сейсмосенсорного канала дает ветвь на зубной кости. Сейсмосенсорный канал открывается на поверхности костей многочисленными порами ветвящихся каналов. Скульптура покровных костей – бугорки и ребрышки, образующие сеть.
Видовой состав. Типовой вид и R. serpukhovensis sp. nov.
Сравнение. У представителей рода Rhizodus, в отличие от рыб рода Strepsodus Young, эмалеподобная ткань на зубах не скульптирована тонкой вертикальной струйчатостью, и клыки не изогнуты сигмовидно. От рода Barameda Long Rhizodus отличается расширением зубной коcти в передней части нижней челюсти. Представители рода Rhizodus отличаются от таковых рода Screbinodus Andrews покровной скульптурой, не распадающейся на отдельные изгибающиеся гребешки и бугорки. От рода Pycnoctenion Vorobyeva et Obrucheva род Rhizodus отличается покровной скульптурой, не состоящей из гребешков и бугорков, а также срастанием зубной кости и инфраденталий без участия симфизных костей, и отсутствием прекороноидной ямы. От рыб рода Gooloogongia Johanson et Ahlberg представители рода Rhizodus отличаются более широкими и короткими клыками, не загнутыми внутрь челюстями и наличием ветки сейсмосенсорного канала на зубной кости. Клыки рыб рода Rhizodus, в отличие от таковых у представителей рода Archichthys Hancock et Atthey, имеют режущие канты. От рода Letognathus Brazeau род Rhizodus отличается отсутствием дентальной части симфиза и краевых зубов спереди от симфизного клыка; адсимфизная пластинка у рыб рода Rhizodus, в отличие от таковой у представителей рода Letognathus, не контактирует с прекороноидом.
Rhizodus serpukhovensis Smirnova, sp. nov.
Название вида – от города Серпухов, рядом с которым находится типовое местонахождение.
Голотип – ПИН, № 2878/581, часть среднего участка нижней челюсти с короноидными клыками и краевыми зубами; Московская область, Серпуховский р-н, местонахождение Заборье; нижний карбон, серпуховский ярус, стешевская свита, глазеченская подсвита.
Описание (рис. 2–5). Зубная кость состоит из вертикальной пластины, несущей 19 маргинальных (краевых) зубов, и утолщенного дорсального фланга. На заднем крае, между угловой и зубной костями, располагается треугольная вырезка, в которую заходил передний отросток надугловой кости (supraangulare). Толщина дорсального фланга зубной кости увеличивается напротив переднего края всех короноидов.
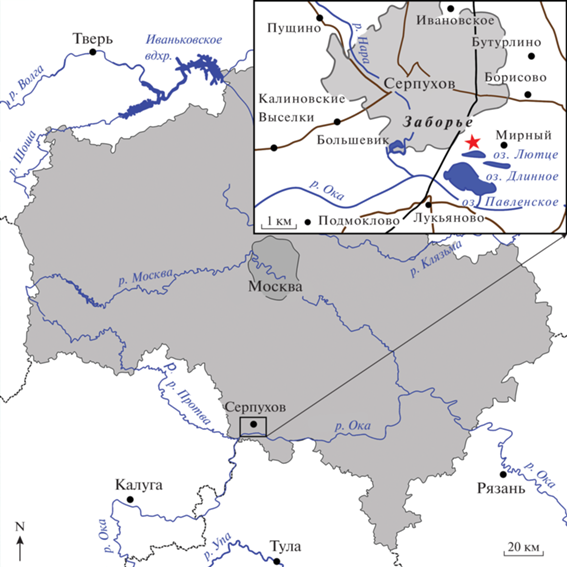
Рис. 2.
Rhizodus serpukhovensis sp. nov., голотип ПИН, № 2878/581, фрагмент нижней челюсти, с лингвальной стороны; внешний вид и прорисовка. Обозначения: Ang – угловая кость; cf – короноидный клык; Co1 – прекороноид, или первый короноид; Co2 – интеркороноид, или второй короноид; De – зубная кость; fs –лунка замещающего клыка; if – интеркороноидная яма; ll – внешняя кромка ротового края зубной кости; mt – маргинальные зубы; plic – складки основания.
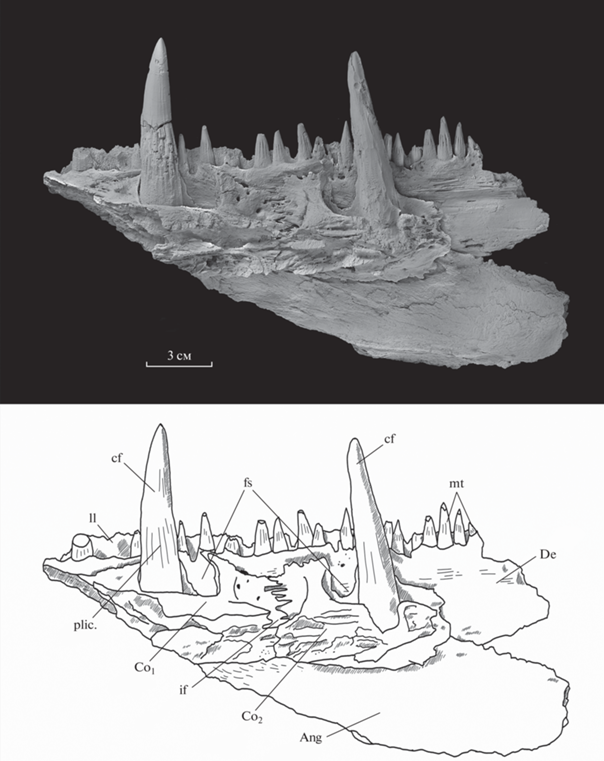
Рис. 3.
Rhizodus serpukhovensis sp. nov., голотип ПИН, № 2878/581, фрагмент нижней челюсти, с лабиальной стороны; внешний вид и прорисовка. Обозначения: lp – поры боковой линии, pl – ямочная линия; остальные как на рис. 2.
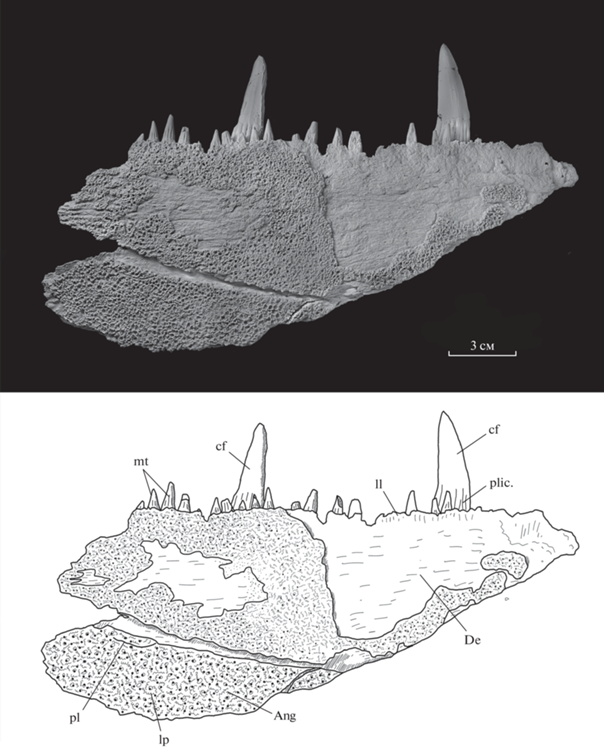
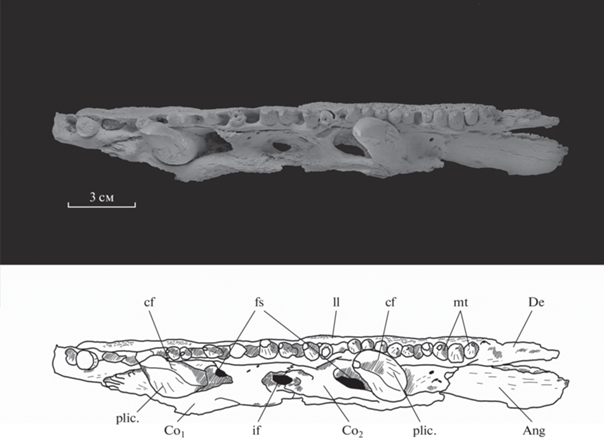
Рис. 4.
Rhizodus serpukhovensis sp. nov., голотип ПИН, № 2878/581, фрагмент нижней челюсти, с лабиальной стороны; покровная скульптура угловой кости (а) и увеличенное изображение ямочной линии (б). Обозначение: pl – ямочная линия.

Рис. 5.
Rhizodus serpukhovensis sp. nov., голотип ПИН, № 2878/581, фрагмент нижней челюсти, с дорсальной стороны; внешний вид и прорисовка. Обозначения как на рис. 2.
Внутренняя поверхность зубной кости образует площадки прикрепления короноидов, которые скульптированы прямыми продольными гребнями. Отсутствие третьего короноида позволяет наблюдать площадку его прикрепления к зубной кости. Вырезки, образующиеся в прилегающих к внутренней поверхности зубной кости вертикальных пластинах обоих короноидов в лунках замещающих зубов, открывают внутреннюю поверхность зубной кости, которая в этих местах более гладкая, чем в других. Внешняя кромка ротового края зубной кости прямая, располагается выше, чем ундулирующая внутренняя, ограничивающая зубной ряд лингвально (рис. 2).
Снаружи угловая кость отделена от зубной широкой шовной поверхностью. С лингвальной стороны к угловой кости прикрепляются вентральные части вертикальных пластин короноидов. На внутренней поверхности кости пучки костных волокон создают рисунок из продольных разветвляющихся гребешков. Вентральный край угловой кости с внутренней стороны образует тонкий, загнутый лингвально гребень, ограничивающий желоб с несколькими отверстиями, отмечающими прохождение веточек сейсмосенсорного мандибулярного канала. Не сохранившаяся заднепластинчатая кость (postspleniale) скреплялась с угловой костью с образованием плоской площадки.
Покровная скульптура на зубной и угловой костях состоит из ямок и гребешков, сливающихся в сеть. Дно и стенки ямок пронизаны мелкими отверстиями многочисленных сосудов; вероятнее всего, в этих ямках располагались сосудистые петли, питавшие кожный покров. Ячейки скульптуры на угловой кости несколько крупнее, чем на зубной (рис. 3). С лабиальной стороны на угловой кости в постеродорсальной части, вблизи ее шва с зубной костью, располагается короткая изогнутая ямочная линия, передним концом упирающаяся в шов между костями (рис. 4).
Из короноидного ряда сохранились прекороноид (или первый короноид) и интеркороноид (или второй короноид), между ними расположена короноидная яма с отверстием в дне (рис. 5). Вертикальная пластина на месте соприкосновения короноидных клыков с зубной костью истончена, вытесненная основаниями клыков, т.е., последние контактируют одной стороной с зубной костью. Зубной ряд вертикальных пластин не развит. Горизонтальная пластина короноидов развита в большей степени у второго, и в меньшей – у первого короноида. Днища лунок замещающих зубов на горизонтальной пластине перфорированы крупными отверстиями, предположительно возникшими в результате деятельности одонтокластов в процессе замены клыков. Предсочленовная кость (praearticulare) не сохранилась, но площадка ее прикрепления к короноидам отмечена грубой шовной поверхностью.
Зубная система состоит из краевых зубов, расположенных в один ряд на зубной кости, и клыков на короноидах. На имеющемся фрагменте сохранились 19 краевых зубов и 10 лунок замещающих зубов, которые по отношению к краевым зубам расположены неравномерно. Краевые зубы достигают наибольшего размера в местах лингво-лабиальных утолщений ротового края зубной кости, связанных с примыканием передних и задних пластин короноидов, а наименьшего – напротив пар короноидных клыков. Краевые зубы высококонические, их вершинки слегка загнуты лингвально. Самый крупный краевой зуб высотой 20.1 мм расположен перед прекороноидом. Диаметр основания краевого зуба приблизительно в 2.5–3 раза меньше его высоты. Складки основания зуба занимают около 2/3 высоты. В поперечном сечении краевые зубы круглые, без режущего канта. Эмалеподобная ткань, выстилающая зубы, прослеживается до орального края зубной кости. Лабиальнее ряда краевых зубов на внешнем ротовом крае зубной кости на уровне интеркороноидного и, предположительно, короноидного клыков расположены два маленьких зуба “первой генерации” (Chen et al., 2020).
На каждом из двух имеющихся короноидов располагаются по рабочему высококоническому клыку и по лунке замещающего клыка. Высота короноидного клыка составляет около 10 см, приблизительно в четыре раза больше его диаметра и в пять раз больше высоты краевых зубов. Складки основания занимают более половины высоты клыка. Средняя часть прекороноидного клыка деформирована в процессе седиментогенеза.
Апикальная половина клыка интеркороноида слегка наклонена вперед и лингвально. Форма сечения обоих клыков изменяется по высоте от округлой у основания до уплощенной лингво-лабиально у вершины. В отличие от краевых зубов, на короноидных клыках с передней и задней сторон располагаются острые гладкие режущие канты, протягивающиеся до уровня верхней кромки зубной кости. Базальная граница распространения эмалеподобной ткани на клыках, отмечающая границу прилегания мягких тканей, приблизительно совпадает с уровнем оснований краевых зубов.
Размеры в мм. Длина образца (экз. ПИН, № 2878/581) – 250, ширина в средней части – 80, расстояние между короноидными клыками – 80, высота прекороноида с клыком – 103.2, высота интеркороноида с клыком – 95.4. Диаметр прекороноидного клыка – 18.6, диаметр интеркороноидного клыка – 17.3. Целые краевые зубы: высота самого крупного – 20.1, диаметр – 10.3; высота самого мелкого – 4.5, диаметр – 2.3; высота среднего – 13.5, диаметр – 5.9. Толщина зубной кости в среднем – 3.2.
Сравнение. Новый вид отличается от R. hibberti наличием интеркороноидной ямы и наличием на зубной кости пор, через которые на внешнюю поверхность открываются веточки сейсмосенсорного канала. Кроме того, в отличие от R. hibberti, у нового вида заднепластинчатая (postspleniale) и угловая кости скреплялись с образованием лишь плоской площадки, а не с помощью зубчатого шва. Угловая кость у R. serpukhovensis sp. nov. длиннее, доходит до середины прекороноида, а у R. hibberti – до уровня интеркороноидной ямы, а надугловая кость (supraangulare) также длиннее, доходит до середины третьего короноида, в то время как у R. hibberti – до передней половины аддукторной ямы.
Материал. Голотип.
МОРФОФУНКЦИОНАЛЬНЫЙ АНАЛИЗ СТРОЕНИЯ ЗУБОВ РИЗОДОНТИФОРМОВ
Зубы ризодонтиформов, в отличие от остеолепиформов и поролепиформов, очень сильно укреплены в челюсти (рис. 6). Основания симфизных клыков представителей рода Rhizodus равны по высоте или даже превышают высоту коронки и могут достигать высоты зубной кости в самой широкой ее части. Высота оснований короноидных клыков уменьшается по мере сужения зубной кости в каудальном направлении (Jeffery, 2003). Скорее всего, основная зона захвата добычи, связанная с максимальными нагрузками, приходилась на передние клыки. Откусывание части тела добычи ризодонтами могло совершаться в этих условиях при вращательном движении туловища, как это делают современные крокодилы и акулы (Andrews, 1985).
Рис. 6.
Rhizodus hibberti (Agassiz et Hibbert in Hibbert) (слева) и Eusthenopteron foordi Whiteaves (справа); схематическое изображение нижней челюсти на уровне прекороноидного клыка в поперечном сечении (из: Jeffery, 2003). У Rhizodus hibberti короноид сильно зажат между зубной и предсочленовной костями, благодаря чему короноидный клык глубоко входит в кость и за счет этого укрепляется. Обозначения: ic – меккелева полость, Mb – Меккелева кость, Pra – предсочленовная кость; остальные как на рис. 2.
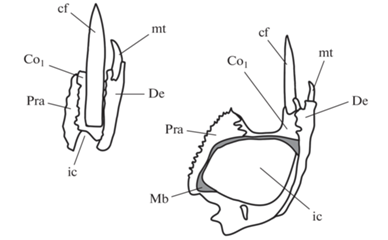
Краевые зубы ризодонтиформов несколько загнуты лингвально, и принцип их действия можно сравнить с принципом действия сигмовидно изогнутых зубов удавообразных змей. Во-первых, такая форма позволяет увеличить расстояние между верхушками зубов, не изменяя расстояния между основаниями, что увеличивает устойчивость каждого из зубов под давлением и дает больше шансов удержать жертву. Во-вторых, поверхность раны, наносимой изогнутым клыком, на теле жертвы будет больше, так как удар будет наноситься под углом. Тот же эффект помогает при попытках жертвы вырваться – загнутые зубы удерживают ее лучше (Deufel, Cundall, 1999).
Рабочие канты на короноидных клыках представителей родов Rhizodus и Screbinodus были важны для увеличения диаметра раны на теле добычи, а также имели значение для укрепления зуба при давлении в ростро-каудальном направлении. Края такого типа могли компенсировать негативный эффект, производимый изогнутостью зубов, которая выражена несколько слабее у рыб, относимых к родам Rhizodus и Screbinodus в сравнении с родами Gooloogongia и Strepsodus (Jeffery, 2003). Кроме того, режущие канты могли служить для отрывания куска от тела жертвы во время резких движений передней части туловища из стороны в сторону (Andrews, 1985).
Схожими с представителями рода Rhizodus формой и размерами зубов обладали некоторые юрские мозазавры (напр., Metiorhynchus cultridens, Temnodontosaurus platyodon). Их тонкие, гладкие, прямые зубы с острыми режущими кантами с обеих сторон и заостренной верхушкой не просто пронизывали жертву, а рассекали ее. Мозазавры охотились в основном на некрупную рыбу (0.6–0.9 м), гигантских кальмаров, белемнитов и аммонитов, и даже черепах (Massare, 1987). Это говорит о большом разнообразии возможной добычи для хищников с таким типом зубов.
Симфизные клыки представителей родов Rhizodus, Screbinodus и Strepsodus крупнее короноидных клыков. Расстояние между симфизными и короноидными клыками позволяет предположить, что жертва умертвлялась симфизными клыками, затем переводилась в продольное положение для глотания (скорее всего, головой вперед по направлению движения в пасть хищника) (Jeffery, 2003). По-видимому, размер добычи по ширине (высоте) тела был не меньше расстояния между клыками верхней и нижней челюстей при их смыкании, то есть, добыча была достаточно крупной, чтобы быть удержанной хищником. Так как на изученном фрагменте Rhizodus serpukhovensis sp. nov. можно измерить расстояние между каждым из двух короноидных клыков и интеркороноидной ямой между ними (куда входил клык верхней челюсти), то можно сказать, что расстояние между клыками верхней и нижней челюстей изученного экземпляра в сомкнутом положении достигало примерно 5 см.
Крупнейшие представители рода Rhizodus достигали 7 м в длину, и этого было достаточно, чтобы проглатывать целиком даже относительно крупную добычу (Andrews, 1985). По мнению Джеффри (Jeffery, 2012), длина челюсти составляет 15.8% длины тела рыбы. Размер изученного в работе фрагмента (25 см) составляет примерно треть всей челюсти (если опираться на реконструкции челюстей ризодонтид, сделанные Джеффри), следовательно, оценить длину рыбы при жизни можно приблизительно в 4.5–5 м.
Из того же слоя, из которого происходит описанный в настоящей статье вид R. serpukhovensis sp. nov., неизвестны остатки крупных рыб, которые могли бы считаться его добычей. Однако, из других слоев стешевского горизонта известны находки гигантских хрящевых рыб (колл. ПИН, № 2878, неопубликованный материал): Stethacanthus altonensis St. John et Worthen, Metaxyacanthus rossicus Inostrantzeff и Erismacanthus sp. Кроме того, из британских местонахождений Ворди и Бёрдихаус, в которых был найден Rhizodus hibberti, известны четвероногие Lethiscus stocki Wellstead, Dolichopareias disjectus Watson и Pholidogaster pisciformis Huxley (Smithson, 1985), которые также могли быть потенциальной добычей для рыб рода Rhizodus. Учитывая гигантские размеры рыбы, размеры и форму ее зубов, можно предположить, что Rhizodus serpukhovensis sp. nov. был крупнейшим хищником в стешевском сообществе позвоночных.
ЗАКЛЮЧЕНИЕ
Из нижнего карбона (серпуховский ярус) местонахождения Заборье (Московская обл., Серпуховский р-н) описан новый вид ризодонтиформной рыбы – Rhizodus serpukhovensis sp. nov. Изученный экземпляр найден в криноидных известняках стешевского горизонта и является самой молодой находкой черепного материала, относимого к роду Rhizodus, среди известных ранее, а также первой каменноугольной ризодонтиформной рыбой, найденной в России.
Описанный в настоящей статье материал отличается от известного ранее британского и североамериканского рядом морфологических признаков: наличием интеркороноидной ямы, пор мандибулярного сейсмосенсорного канала на зубной кости, ямочной линии на угловой кости, а также несколько другой формой и расположением костей.
Морфофункциональный анализ строения зубов ризодонтид показал, что загнутые внутрь симфизные клыки служили для захвата добычи, прямые короноидные клыки с режущими кантами способствовали нанесению глубокой режущей раны на теле жертвы и переведением ее в пригодное положение для заглатывания, загнутые лингвально краевые зубы удерживали добычу.
Гигантские размеры, сильные челюсти, острые клыки R. serpukhovensis sp. nov. позволяют предположить, что он был доминантным хищником в стешевском сообществе позвоночных.
Список литературы
Воробьева Э.И. Подкласс Crossopterygii. Кистеперые рыбы // Ископаемые позвоночные России и сопредельных стран. Бесчелюстные и древние рыбы / Ред. Новицкая Л.И., Афанасьева О.Б. М.: ГЕОС, 2004. С. 272–372.
Воробьева Э.И., Обручев Д.В. Подкласс Sarcopterygii // Основы палеонтологии. Бесчелюстные, рыбы / Ред. Обручев Д.В. М.: Наука, 1964. С. 268–322.
Andrews S.M. Rhizodont crossopterygian fish from the Dinantian of Foulden, Berwickshire, Scotland, with a re-evaluation of this group // Trans. Roy. Soc. Edinburgh. Earth and Environ. Sci. 1985. V. 76. № 1. P. 67–95.
Andrews S.M., Westoll T.S. The postcranial skeleton of rhipidistian fishes excluding Eusthenopteron // Trans. Roy. Soc. Edinburgh. Earth and Environ. Sci. 1970. V. 68. № 12. P. 391–489.
Aretz M., Herbi H.G., Wang X.D. The Carboniferous Period // Geologic Time Scale 2020. V. 2 / Eds Gradstein F.M., Ogg J.G., Schmitz M.D., Ogg G.M. Amsterdam: Elsevier, 2020. P. 811–874.
Buckland W. Geology and Mineralogy Considered with Reference to Natural Theology. V. 1. Philadelfia: Carey, Lea and Blanchard, 1837. 468 p.
Chen D., Blom H., Sanchez S. et al. The developmental relationship between teeth and dermal odontodes in the most primitive bony fish Lophosteus // Elife. 2020. V. 9. P. 1–20.
Deufel A., Cundall D. Do booids stab prey? // Copeia. 1999. P. 1102–1107.
Egerton P.G., Cole E.W.W. A Systematic and Stratigraphical Catalogue of the Fossil Fish in the Cabinets of Lord Cole and Sir Philip Grey Egerton: Together with an Alphabetical and Stratigraphical Catalogue of the Same Species, with References to Their Published Figures and Descriptions. L.: R. and J.E. Taylor, 1837. 29 p.
Gibshman N.B., Kabanov P.B., Alekseev A.S. et al. Novogurovsky Quarry–Upper Visean and Serpukhovian // Type and reference Carboniferous Sections in the South Part of the Moscow Basin. M.: Borissiak Paleontol. Inst. Russ. Acad. Sci., 2009. P. 13–44.
Hibbert S. On the fresh-water limestone of Burdiehouse in the neighbourhood of Edinburgh, belonging to the Carboniferous Group of Rocks. With supplementary notes on other freshwater limestones // Trans. Roy. Soc. Edinburgh. Earth and Environ. Sci. 1836. V. 13. № 1. P. 169–282.
Jarvik E. On the Dermal Bones, Sensory Canals and Pit-lines of the Skull in Eusthenopteron foordi Whiteaves: With Some Remarks on E. säve-söderberghi Jarvik. Stockholm: Almqvist & Wiksells, 1944. 115 p.
Jeffery J.E. Mandibles of rhizodontids: anatomy, function and evolution within the tetrapod stem group // Trans. Roy. Soc. Edinburgh. Earth and Environ. Sci. 2003. V. 93. № 3. P. 255–276.
Jeffery J.E. The Carboniferous fish genera Strepsodus and Archichthys (Sarcopterygii: Rhizodontida): clarifying 150 years of confusion // Palaeontology. 2006. V. 49. Pt 1. P. 113–132.
Jeffery J.E. Cranial morphology of the Carboniferous rhizodontid Screbinodus ornatus (Osteichthyes: Sarcopterygii) // J. Syst. Palaeontol. 2012. V. 10. № 3. P. 475–519.
Johanson Z., Jeffery J.E., Challands T. et al. A new look at Carboniferous rhizodontid humeri (Sarcopterygii; Tetrapodomorpha) // J. Vertebr. Paleontol. 2020. V. 40. № 3: e1813150. https://doi.org/10.1080/02724634.2020.1813150
Long J.A. A new rhizodontiform fish from the Early Carboniferous of Victoria, Australia, with remarks on the phylogenetic position of the group // J. Vertebr. Paleontol. 1989. V. 9. № 1. P. 1–17.
Massare J. Tooth morphology and prey preference of Mesozoic marine reptiles // J. Vertebr. Paleontol. 1987. T. 7. № 2. P. 121–137.
Menning M., Schneider J.W., Alekseev A.S. et al. Global time scale and regional stratigraphic reference scales of central and west Europe, east Europe, Tethys, south China, and North America as used in the Devonian–Carboniferous–Permian Correlation Chart 2003 (DCP 2003) // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2006. V. 240. P. 318–372.
Miall L.C. On the structure of the skull of Rhizodus // Quart. J. Geol. Soc. 1875. V. 31. № 1–4. P. 624–627.
Nikolaeva S.V., Alekseev A.S., Kulagina E.I. et al. An evaluation of biostratigraphic markers across multiple geological sections in the search for the GSSP of the base of the Serpukhovian Stage (Mississippian) // Palaeoworld. 2020. V. 29. № 2. P. 270–302.
Owen R. Odontography, or, a treatise on the comparative anatomy of the teeth; their physiological relations, mode of development, and microscopic structure in the vertebrate animals. L.: Hippolyte Bailliere, 1840. 655 p.
Smithson T.R. Scottish Carboniferous amphibian localities // Scottish J. Geol. 1985. V. 21. № 2. P. 123–142.
Traquair R.H. On the genus Rhizodus // Proc. Roy. Soc. Edinburgh. 1878. V. 9. P. 657–660.
White E. The fish-fauna of the Cementstones of Foulden, Berwickshire // Trans. Roy. Soc. Edinburgh. Earth and Environ. Sci. 1927. V. 55. № 1. P. 255–287.
Young J. Notice of the occurrence of Rhizodus hibberti in Carboniferous Strata in the neighbourhood of Glasgow // Trans. Geol. Soc. Glasgow. 1865. V. 2. № 1. P. 38–39.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал