

Палеонтологический журнал, 2022, № 4, стр. 87-94
Применение зигапофизной скелетохронологии для определения индивидуального возраста базального мозазаврида (Squamata, Mosasauridae) из кампана Саратовской области
Д. В. Григорьев a, b, *, М. С. Архангельский c, d, В. В. Колчанов a, b, В. В. Буланов e, А. Г. Сенников e, В. К. Голубев e, П. П. Скучас a, b
a Санкт-Петербургский государственный университет
199034 Санкт-Петербург, Россия
b Зоологический институт РАН
199034 Санкт-Петербург, Россия
c Саратовский государственный технический университет
410054 Саратов, Россия
d Саратовский государственный университет
410012 Саратов, Россия
e Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
* E-mail: grigoriev_dmitry@mail.ru
Поступила в редакцию 10.01.2022
После доработки 03.02.2022
Принята к публикации 03.02.2022
- EDN: JGCBMV
- DOI: 10.31857/S0031031X2204002X
Аннотация
Для определения индивидуального возраста базального мозазаврида (представителя подсемейства Halisaurinae) из позднемелового местонахождения Белое Озеро (Саратовская область) по шейному позвонку был использован метод зигапофизной скелетохронологии. В результате подсчета зигапофизных ростовых гребней на правом переднем сочленовном отростке было установлено, что возраст животного, которому принадлежал этот позвонок, составлял не менее 11 лет. Отсутствие резкого сокращения расстояния между зигапофизными гребнями указывает на то, что первые 11 лет жизни, до момента гибели, животное росло быстро и равномерно, и достигло длины около 6 метров.
ВВЕДЕНИЕ
Мозазавриды – группа крупных вторично-водных морских варанообразных ящериц, существовавшая в позднем мелу с туронского времени (возможно, сеноманского) по конец маастрихтского века. Мозазавриды обитали в эпиконтинентальных морях и на шельфах, прилегающих к глубоководным впадинам. Их остатки были найдены на всех континентах, в т.ч. в Антарктиде (Polcyn et al., 2014). В Европейской части России и на смежных территориях разрозненные остатки и неполные скелеты мозазаврид обычны (Первушов и др., 1999).
Подавляющее число костных остатков мозазаврид из России были найдены на территории Поволжья – в Пензенской, Саратовской и Волгоградской областях. Материал с исключительной сохранностью происходит из “Белого Озера” – кампанского местонахождения, расположенного у одноименного села в Лысогорском р-не Саратовской обл. Местонахождение представляет собой серию оврагов, вскрывающих отложения рыбушкинской свиты, по стратиграфическому объему соответствующей большей части нижнего и низам верхнего кампана (Олферьев, Алексеев, 2005). В верхней части отложений свиты находится фосфоритовый горизонт – прослой, состоящий из фосфоритовых желваков, имеющих различную концентрацию. Позвонок мозазаврида, описываемый в настоящей работе, был найден в фосфоритовом слое. Из местонахождения ранее были описаны разрозненные остатки рыб, птерозавров (Аверьянов, Попов, 2014; Аверьянов и др., 2016; Averianov, Arkhangelsky, 2020), лобная кость мозазаврида Clidastes propython Cope, 1869 (Григорьев и др., 2015), остатки черепах и плезиозавров (Очев, 1976; Архангельский и др., 2007; Danilov et al., 2018; Zverkov et al., 2018).
В 2017 г. в ходе раскопочных работ на местонахождении студент Саратовского государственного технического ун-та А.А. Щетинкин обнаружил шейный позвонок мозазаврида с необычной морфологией, отнесенный к представителю подсемейства хализаврин (Halisaurinae). Остатки хализаврин ранее не описывались с территории России.
На передних и задних сочленовных отростках (пре- и постзигапофизах соответственно) изученного позвонка мозазаврида невооруженным глазом видны концентрические структуры. Подобные структуры (зигапофизные ростовые зоны, или зигапофизные циклиты) ранее отмечались на сочленовной поверхности зигапофизов у разных тетрапод, в т.ч. у чешуйчатых рептилий (Venczel et al., 2015; Petermann, Gauthier, 2018; Skutschas et al., 2020), и использовались для определения индивидуального возраста (в рамках зигапофизного скелетохронологического анализа).
В настоящей статье приводится морфологическое описание шейного позвонка мозазаврида из местонахождения “Белое Озеро” и данные о его индивидуальном возрасте, полученные методом зигапофизной скелетохронологии. Для представителей семейства Mosasauridae подобные сведения публикуются впервые.
Описываемый экземпляр (ZIN PH № 29/90) хранится в палеогерпетологической коллекции Зоологического ин-та РАН (ЗИН) в С.-Петербурге (ZIN PH). Для сравнения использовались материалы из коллекции Музея естественной истории Маастрихта (NHMM, Нидерланды) и Музея естественной истории Марракеша (MHNM.KH, Марокко).
МЕТОД ЗИГАПОФИЗНОЙ СКЕЛЕТОХРОНОЛОГИИ
В скелетохронологии для определения индивидуального возраста применяется прямой подсчет циклических годичных ростовых меток (Woodward et al., 2013; Buffrenil, Quilhac, 2021). Такие циклические годичные ростовые метки могут наблюдаться внутри скелетных структур (в костях, в цементе), и их подсчет осуществляется при применении классической гистологической скелетохронологии, когда количество ростовых меток определяется на тонких срезах. При этом сами ростовые метки, которые учитываются при анализе, представлены либо линиями остановки роста (lines of arrested growth, LAGs), соответствующими полной остановке остеогенеза, либо линиями замедления роста (annuli), представленными слоями плотной аваскулярной костной ткани, которые соответствуют существенному замедлению остеогенеза (Woodward et al., 2013; Buffrenil, Quilhac, 2021).
Кроме внутренних ростовых меток существуют годичные ростовые метки, формирующиеся на поверхности костей, в т.ч. на поверхностях сочленовных отростков позвонков. Как было показано в предыдущих исследованиях, зигапофизные ростовые метки формируются в онтогенезе и отражают сезонные циклы (обычно годовые) нарастания костной ткани, хорошо коррелируя при этом с внутрикостными ростовыми метками (LAGs) (Petermann, Gauthier, 2018; Skutschas et al., 2020). Прямой подсчет их числа может использоваться как альтернатива (или как дополнение) классическому деструктивному скелетохронологическому подходу, который требует подготовки тонких срезов костей и последующего подсчета линий остановок роста или линий замедления роста (Petermann, Gauthier, 2018; Skutschas et al., 2020).
Поскольку устоявшейся терминологии для зигапофизной скелетохронологии не существует, в данной работе мы вводим основные термины и понятия (по аналогии с гистологической скелетохронологией) (рис. 1).
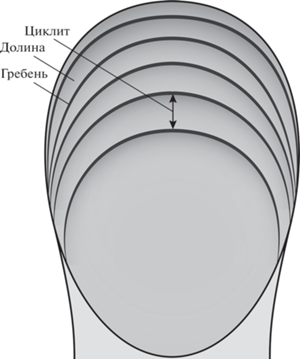
Собственно, годичная ростовая метка в зигапофизной скелетохронологии – это зигапофизный гребень, который формируется в конце каждого цикла роста (при его замедлении/остановке). Зигапофизный гребень соответствует линиям остановок роста и/или линиям замедления роста в гистологии. Обозначать зигапофизный гребень как линию остановки роста (LAG), как это было сделано у Х. Питерманна и Ж. Готье (Petermann, Gauthier, 2018), нам представляется не совсем корректным, поскольку эти гребни могут быть достаточно широкими и формироваться при замедлении роста, а не при полной остановке (т.е., больше соответствовать линиям замедления роста в гистологии), а также потому, что использование одних и тех же терминов для обозначения внутренних и внешних ростовых меток вызовет путаницу при сравнении результатов разных скелетохронологических подходов.
Между зигапофизными гребнями расположены более низкие участки, соответствующие активному росту кости – данные структуры мы обозначаем как зигапофизные долины (valles). Весь годовой ростовой цикл включает в себя зигапофизную долину и гребень и обозначается как зигапофизный циклит [соответствует годичному ростовому циклу (annual growth cycle) в работе Питерманна и Готье (Petermann, Gauthier, 2018)].
Подсчет числа зигапофизных гребней и циклитов позволяет реконструировать индивидуальный возраст, а оценка расстояния между гребнями (=оценка ширины циклитов) позволяет делать реконструкции характера роста (при быстром росте расстояние между гребнями будет большим, при замедлении роста оно будет уменьшаться).
Зигапофизные гребни состоят из минерализованного (кальцифицированного) хряща) (Skutschas et al., 2020). На ископаемом материале кальцифицированный хрящ сохраняется хуже, чем кость, и часть зигапофизных гребней может утрачиваться при захоронении или препарировании. Соответственно, если сохранность зигапофизов на одном позвонке разная, то для анализа необходимо выбирать зигапофиз (или зигапофизы) с максимальным числом сохранившихся зигапофизных гребней. В нашем исследовании это наиболее хорошо сохранившийся правый презигапофиз с 11 зигапофизными гребнями.
МОРФОЛОГИЧЕСКОЕ ОПИСАНИЕ ПОЗВОНКА
Высокие вертикальные фасетки для причленения ребер позволяют предположить, что позвонок мог иметь положение с четвертого по седьмой в шейном отделе (Holmes, Sues, 2000).
Тело позвонка (рис. 2) сильно вытянуто и дорсовентрально сжато (соотношение длины к ширине – 1.95, ширины к высоте – 1.9). Его длина составляет 74 мм. Форма заднего мыщелка эллипсоидная, его ширина – 38 мм, высота – 20 мм. Боковые отростки короткие, незначительно выступающие за презигапофизы. Фасетки для причленения ребер высокие и узкие, вентральные края практически вплотную примыкают к переднему мыщелку. Ножка для причленения гипапофиза имеет каплевидную форму без ярко выраженного углубления. Презигапофизы массивные и сильно вытянутые вперед. Их длина – 27 мм от основания у спинномозгового канала.
Рис. 2.
Шейный позвонок Halisaurinae gen. indet., экз. ZIN PH № 29/90: а – вид спереди, б – вид сзади, в – вентральный вид, г – дорсальный вид, д, е – виды сбоку; Саратовская обл., Белое Озеро; верхний мел, нижний кампан, рыбушкинская свита. Полупрозрачными четырехугольниками показаны области на зигапофизах с ростовыми гребнями, изображенными крупно на рис. 3. Обозначения: cdl – задняя сочленовная поверхность позвонка, ctl – передняя сочленовная поверхность позвонка, hyp – ножка гипапофиза, mpc – задний среднесагиттальный гребень, poz – постзигапофиз, или задний сочленовный отросток позвонка, prz – презигапофиз, или передний сочленовный отросток позвонка, syn – боковой отросток позвонка.
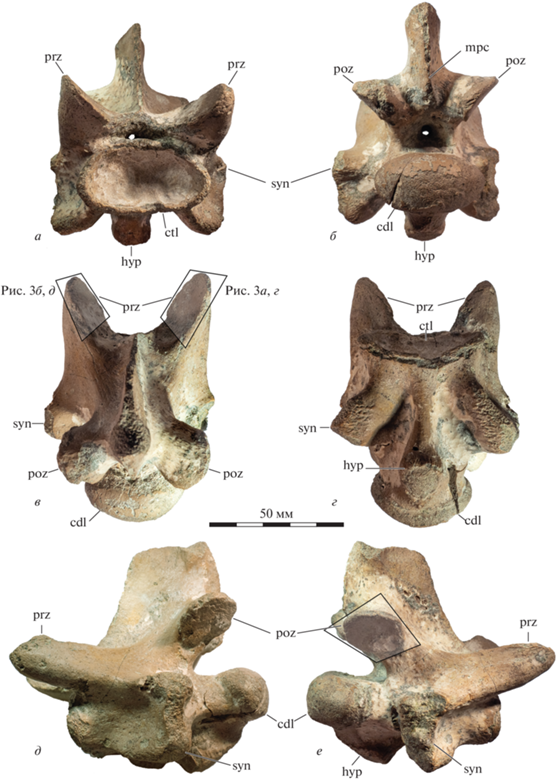
Сочленовные поверхности имеют сильно вытянутую эллипсоидную форму. Зигосфены и зигантры отсутствуют. Остистый отросток имеет ярко выраженный задний среднесагиттальный гребень, отсутствующий, например, у Mosasaurus hoffmannii Mantell, 1829 (шейный позвонок, экз. NHMM 06696-4, личные наблюдения Д.В. Григорьева), но хорошо выраженный, например, у Phosphorosaurus ponpetelegans Konishi et al., 2015.
СРАВНЕНИЕ
Сильно уплощенная форма заднего мыщелка характерна для представителей подсемейств Plioplatecarpinae и Halisaurinae, тогда как у большинства представителей Mosasaurinae и Tylosaurinae они практически круглые (Russell, 1967; Caldwell, Bell, 1995). Согласно М. Колдуэллу и Дж. Бэллу (Caldwell, Bell, 1995), соотношение длины тела позвонка/высоты заднего мыщелка к длине тела позвонка/ширине заднего мыщелка у Plioplatecarpinae находится в диапазоне от 1.18 до 1.47, а у Halisaurinae – от 1.46 до 2.16. У экз. ZIN PH № 29/90 это значение равно 1.94, что позволяет с большой уверенностью отнести его к Halisaurinae. В подавляющем большинстве представители хализаврин были некрупными мозазавридами длиной два–три метра (Polcyn et al., 2013). Исключение составляют представители рода Pluridens Lingham-Soliar, 1998, которые могли достигать 10 м и более (Longrich et al., 2021). В частности, для экз. MHNM.KH.262, отнесенного к Pluridens serpentis Longrich, 2021, и имеющего расчетную длину приблизительно 6.5 м, длина четвертого шейного позвонка составляет примерно 81 мм (измерено по фотографии). У экз. ZIN PH № 29/90 длина тела позвонка составляет 74 мм, соответственно, длина тела могла быть приблизительно шесть метров. Такие крупные размеры косвенно указывают на то, что экз. ZIN PH № 29/90 мог принадлежать представителю рода Pluridens, но имеющихся данных недостаточно для обоснования этого предположения.
ЗИГАПОФИЗНЫЙ СКЕЛЕТОХРОНОЛОГИЧЕСКИЙ АНАЛИЗ
На сочленовной поверхности зигапофизов различимы концентрические ростовые метки, зигапофизные гребни, отражающие этапы ежегодного замедления/остановки роста. Подобные структуры различимы под косым освещением, и их максимальное число отмечено на правом презигапофизе (рис. 3, а, г). Левый презигапофиз и правый постзигапофиз были незначительно повреждены у основания при препарировании, поэтому зигапофизные ростовые гребни на них прослеживаются не в полной мере. На наиболее хорошо сохранившемся правом презигапофизе можно насчитать до 11 ростовых зигапофизных гребней. Гребни прослеживаются не по всей сочленовной поверхности и имеют дугообразную форму.
Рис. 3.
Шейный позвонок Halisaurinae gen. indet., экз. ZIN PH № 29/90, сочленовные поверхности правого презигапофиза (а, г), левого презигапофиза (б, д) и правого постзигапофиза (в, е) с ростовыми гребнями, где г, д, е – пояснительные прорисовки ростовых ребер. На прорисовках темным серым цветом показаны сочленовные поверхности. Нумерация ростовых ребер на разных зигапофизах не соотносится друг с другом из-за различной степени сохранности.

Ширина зигапофизных циклитов между первым и вторым и между вторым и третьим зигапофизными гребнями примерно одинаковая, далее ширина циклита между третьим и четвертым гребнями несколько увеличивается. С четвертого и по 11-й гребень ширина циклитов незначительно различается (визуально циклиты между седьмым и восьмым гребнями, а также между десятым и 11-м гребнями немного шире соседних и по ширине соответствуют циклитам между первым и вторым и между вторым и третьим зигапофизными гребнями). В целом, резкого сокращения расстояния между гребнями (т.е. резкого уменьшения ширины циклитов) не наблюдается.
ОБСУЖДЕНИЕ
Гистологическая скелетохронология – метод, широко используемый для определения индивидуального возраста позвоночных животных путем подсчета линий остановки роста (LAGs) – концентрических линий, образующихся ежегодно в связи с остановкой роста. Этот метод используется для определения возраста современных (Matsuki, Matsui, 2009; Эпова и др., 2016; Fornasiero et al., 2016; Guarino et al., 2016) и вымерших (Buffrenil, Buffetaut, 1981; Horner et al., 1999; Erickson, Tumanova, 2000; Skutschas et al., 2020) позвоночных. Для вымерших животных этот метод является основным источником получения информации об индивидуальном возрасте особи и темпах роста. Однако метод имеет ряд ограничений.
Например, с возрастом вместе с ростом кости происходит расширение медуллярной полости за счет резорбции внутренней части кортекса, “стирающее” информацию о раннем росте кости (соответственно, ранние ростовые метки не сохраняются). Именно поэтому получить достоверную информацию об индивидуальном возрасте животного, основываясь на данных классической скелетохронологии, можно лишь при использовании онтогенетических выборок с использованием сравнительного материала из разных частей скелета (Chinsamy, 1990; Horner et al., 1999; Erickson, Tumanova, 2000). Также на скелетах современных животных было показано (Schucht et al., 2021), что различные методики гистологической подготовки образцов дают разное число линий остановки роста при их подсчете. Более того, число этих линий не соответствует реальному возрасту животных – практически во всех случаях их число несколько меньше возраста. Тем не менее, это исследование подтверждает, что число линий остановки роста не больше возраста животного.
Еще одно ограничение связано с тем, что этот метод деструктивный и предполагает повреждение (подготовка гистологических срезов) исследуемого материала.
Сравнительно недавно на практике был применен метод зигапофизной скелетохронологии, лишенный ограничений, описанных выше. Данный метод заключается в подсчете количества зигапофизных гребней и позволяет реконструировать индивидуальный возраст, а также характер роста. Питерманн и Готье (Petermann, Gauthier, 2018) на примере современных и ископаемых змей показали, что зигапофизные гребни соответствуют линиям остановки роста (LAGs) на гистологических срезах, что позволяет определять индивидуальный возраст для некоторых групп позвоночных по внешним признакам на костных структурах (на основе изолированных позвонков). Зигапофизная скелетохронология может быть использована для всех тетрапод, у которых сохраняются зигапофизные гребни, например, саламандр, лягушек, антракозавров, сеймуриаморфов, парейазавров, лепидозавров и архозавров (Petermann, Gauthier, 2018; Skutschas et al., 2020).
Исходя из максимального числа видимых зигапофизных ростовых гребней (11), можно предположить, что возраст животного, которому принадлежал шейный позвонок ZIN PH № 29/90, равнялся 11 годам. Это было достаточно крупное животное, достигавшее около 6 м в длину. Учитывая, что последний (11-й) зигапофизный гребень находится не у самого внешнего края зигапофиза (внешний край зигапофиза соответствует зигапофизной долине), то можно предположить, что возраст животного был немногим более 11 лет на момент смерти.
Поскольку по всей длине зигапофиза не наблюдается резкого сокращения расстояния между зигапофизными гребнями, то можно сделать предположение, что первые 11 лет жизни животное росло достаточно быстро и равномерно до момента гибели. Отсутствие резкого сокращения расстояния между зигапофизными гребнями во внешней части зигапофиза свидетельствует, что рост животного на момент гибели не замедлился, что оно не вышло на ростовое плато и не достигло максимальных возможных размеров.
Сходное равномерное расположение зигапофизных гребней и, соответственно, сходный равномерный характер роста на протяжении первых нескольких лет жизни отмечался ранее для современных чешуйчатых рептилий – змей и ящериц (Petermann, Gauthier, 2018; Skutschas et al., 2020).
* * *
Авторы благодарят А.А. Щетинкина (Саратовский государственный технический ун-т, Россия) за помощь в проведении раскопок, М. Полцина (Южный методистский ун-т, Даллас, Техас, США) за ценные замечания по таксономической принадлежности ZIN PH № 29/90, Дж. Яхта (Музей естественной истории Маастрихта, Нидерланды) за доступ к коллекции NHMM, а также рецензентов А.О. Аверьянова (Зоологический ин-т РАН, С.-Петербург, Россия) и Н.Г. Зверькова (Геологический ин-т РАН, Москва, Россия) за конструктивные комментарии к тексту рукописи. Работа поддержана Российским фондом фундаментальных исследований (проект 20-04-00545a).
Список литературы
Аверьянов А.О., Архангельский М.С., Меркулов С.М. Плечевая кость аждархида (Pterosauria, Azhdarchidae) из верхнего мела Саратовской области // Палеонтол. журн. 2016. № 4. С. 93–97.
Аверьянов А.О., Попов Е.В. Позвонок птерозавра из верхнего мела Саратовской области // Палеонтол. журн. 2014. № 3. С. 101–104.
Архангельский М.С., Аверьянов А.О., Первушов Е.М. Короткошеие плезиозавры семейства Polycotylidae из кампана Саратовской области // Палеонтол. журн. 2007. № 6. С. 62–66.
Григорьев Д.В., Архангельский М.С., Меркулов С.М. О находке Clidastes propython (Squamata, Mosasauridae) в верхнем мелу Саратовской области // Палеонтол. журн. 2015. № 5. С. 60–68.
Олферьев А.Г., Алексеев А.С. Стратиграфическая схема верхнемеловых отложений Восточно-Европейской платформы. Объяснительная записка. М.: ПИН РАН, 2005. 203 с.
Очев В.Г. Новый плезиозавр из верхнего мела Пензенской области // Палеонтол. журн. 1976. № 2. С. 135–138.
Первушов Е.М., Архангельский М.С., Иванов А.В. Каталог местонахождений остатков морских рептилий в юрских и меловых отложениях Нижнего Поволжья. Саратов: Изд-во “Колледж”, 1999. 232 с.
Эпова Л.А., Куранова В.Н., Ярцев В.В., Абсалямова Е.Н. Возраст, размеры тела и рост в горных популяциях живородящей ящерицы, Zootoca vivipara (Sauria: Lacertidae) Кузнецкого Алатау (юго-восток Западной Сибири) // Совр. герпетол. 2016. Т. 18. № 1/2. С. 51–60.
Averianov A.O., Arkhangelsky M.S. A large pteranodontid from the Late Cretaceous of Eastern Europe // Geol. Mag. 2020. P. 1143–1155. https://doi.org/10.1017/S0016756820001119
Buffrenil V., Buffetaut E. Skeletal growth lines in an Eocene crocodilian skull from Wyoming as an indicator of ontogenic age and paleoclimatic conditions // J. Vertebr. Paleontol. 1981. V. 1. № 1. P. 57–66.
Buffrenil V., Quilhac A. Bone tissue types: a brief account of currently used categories // Vertebrate Skeletal Histology and Paleohistology Flyer / Eds. Buffrenil V., Ricqles A.J., Zylberberg L., Padian K. P.: CRC Press, 2021. P. 147–182. https://doi.org/10.1201/9781351189590
Caldwell M.W., Bell G.L. Halisaurus sp. (Mosasauridae) from the Upper Cretaceous (?Santonian) of east-central Peru, and the taxonomic utility of mosasaur cervical vertebrae // J. Vertebr. Paleontol. 1995. V. 15. P. 532–544.
Chinsamy A. Physiological implications of the bone histology of Syntarsus rhodesiensis (Saurischia: Theropoda) // Palaeontol. Afr. 1990. V. 27. P. 77–82.
Cope E.D. Remarks on Clidastes propython, Polycotylus latipinnus, Ornithotarsus immanis // Proc. Amer. Phil. Soc. 1869. V. 11. 117 p.
Danilov I.G., Obraztsova E.M., Arkhangelsky M.S. et al. Cretaceous chelonioid turtles of Northern Eurasia: previous records and new findings // Turtle Evolution Symposium. 2018. Waseda University, Japan, May, 26–27, 2018. P. 30–33.
Erickson G.M., Tumanova T.A. Growth curve of Psittacosaurus mongoliensis Osborn (Ceratopsia: Psittacosauridae) inferred from long bone histology // Zool. J. Linn. Soc. 2000. V. 130. P. 551–566.
Fornasiero S., Bonnet X., Dendi F., Zuffi M.A.I. Growth, longevity and age at maturity in the European whip snakes, Hierophis viridiflavus and H. carbonarius // Acta Herpetol. 2016. V. 11. № 2. P. 135–149.
Guarino F.M., Mezzasalma M., Odierna G. Usefulness of postpygal caudal vertebrae and osteoderms for skeletochronology in the limbless lizard Anguis veronensis Pollini, 1818 (Squamata: Sauria: Anguidae) // Herpetozoa. 2016. V. 29. № 1/2. P. 69–75.
Holmes R., Sues H. A partial skeleton of the basal mosasaur Halisaurus platyspondylus from the Severn Formation (Upper Cretaceous: Maastrichtian) of Maryland // J. Paleontol. 2000. V. 74. P. 309–316.
Horner J.R., Ricqlès A., Padian K. Variation in dinosaur skeletochronology indicators: implications for age assessment and physiology // Paleobiol. 1999. V. 25. № 3. P. 295–304.
Konishi T., Caldwell M.W., Nishimura T. et al. A new halisaurine mosasaur (Squamata: Halisaurinae) from Japan: the first record in the western Pacific realm and the first documented insights into binocular vision in mosasaurs // J. Syst. Palaeontol. 2016. V. 14. № 10. P. 809–839. https://doi.org/10.1080/14772019.2015.1113447
Lingham-Soliar T. A new mosasaur Pluridens walkeri from the Upper Cretaceous, Maastrichtian of the Iullemmeden Basin, southwest Niger // J. Vertebr. Paleontol. 1998. V. 18. № 4. P. 709–707.
Longrich N.R., Bardet N., Khaldoune F. et al. Pluridens serpentis, a new mosasaurid (Mosasauridae: Halisaurinae) from the late Maastrichtian of Morocco and implications for mosasaur diversity // Cret. Res. 2021. https://doi.org/10.1016/j.cretres.2021.104882
Mantell G.A. A tabular arrangement of the organic remains of the county of Sussex // Trans. Geol. Soc. London, 2 ser. 1829. V. 3. P. 201–216.
Matsuki T., Matsui M. The validity of skeletochronology in estimating ages of Japanese clouded salamander, Hynobius nebulosus (Amphibia, Caudata) // Curr. Herpetol. 2009. V. 28. № 2. P. 41–48.
Petermann H., Gauthier J.A. Fingerprinting snakes: paleontological and paleoecological implications of zygantral growth rings in serpentes // PeerJ. 2018. 6:e4819. https://doi.org/10.7717/peerj.4819
Polcyn M.J., Jacobs L.L., Araújo R. et al. Physical drivers of mosasaur evolution // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2014. V. 400. P. 17–27.
Russell D.A. Systematics and Morphology of American Mosasaurs // Bull. Peabody Mus. Natur. Hist. Yale Univ. 1967. V. 23. 242 p.
Schucht P.J., Klein N., Lambertz M. What’s my age again? On the ambiguity of histology-based skeletochronology // Proc. R. Soc. B. 2021. V. 288. 20211166. https://doi.org/10.1098/rspb.2021.1166
Skutschas P.P., Kolchanov V.V., Bulanov V.V. et al. Reconstruction of the life history traits in the giant salamander Aviturus exsecratus (Caudata, Cryptobranchidae) from the Paleocene of Mongolia using zygapophyseal skeletochronology // Hist. Biol. 2020. V. 32. № 5. P. 645–648.
Venczel M., Vasile S., Csiki-Sava Z. A Late Cretaceous madtsoiid snake from Romania associated with a megaloolithid egg nest – paleoecological inferences // Cret. Res. 2015. V. 55. P. 152–163.
Woodward H.N., Padian K., Lee A.H. Skeletochronology // Bone Histology of Fossil Tetrapods – Advancing Methods, Analysis and Interpretation / Eds. Padian K., Lamm E.-T. Berkeley: Univ. California Press, 2013. P. 195–216.
Zverkov N.G., Averianov A.O., Popov E.V. Basicranium of an elasmosaurid plesiosaur from the Campanian of European Russia // Alcheringa: Austral. J. Palaeontol. 2018. V. 42. № 4. P. 528–542. https://doi.org/10.1080/03115518.2017.1302508
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал