

Палеонтологический журнал, 2023, № 1, стр. 93-101
Акантоморфные акритархи из отложений венда внутренних районов Сибирской платформы
Е. Ю. Голубкова *
Институт геологии и геохронологии докембрия РАН
199034 Санкт-Петербург, Россия
* E-mail: golubkovaeyu@mail.ru
Поступила в редакцию 18.08.2022
После доработки 31.08.2022
Принята к публикации 31.08.2022
- EDN: FIZBVF
- DOI: 10.31857/S0031031X23010051
Аннотация
Изучен таксономический состав акантоморфных акритарх из терригенных отложений венда Предпатомского, Непско-Ботуобинского, Сюгджерского и Анабарского структурно-фациальных районов Сибирской платформы. Разнообразные орнаментированные микрофоссилии обнаружены на трех стратиграфических уровнях – в средних частях нижнего и верхнего подгоризонтов непского горизонта и в нижней подсвите бюкской свиты тирского горизонта. Проведено монографическое описание новых акантоморфных акритарх Tanarium longidigitatum sp. nov., Tuberculum proteum sp. nov., Uniexcrescentia tubulosa gen. et sp. nov. и двухслойных гладкостенных микрофоссилий Pterospermopsimorpha mutabilis sp. nov.
ВВЕДЕНИЕ
К докембрийским органикостенным микрофоссилиям относят большую группу разнообразных по морфологии и гетерогенных по своей природе ископаемых микроорганизмов и их фрагментов, средние размеры которых не превышают, как правило, 500 мкм. Микрофоссилии сохраняются в сероцветных аргиллитах и алевролитах в виде сплющенных до тончайших пленок органических остатков. Ископаемые организмы извлекают из породы в результате воздействия на нее 45%-й плавиковой (HF) и 10%-й соляной (HCl) кислот и в дальнейшем изучают в постоянных препаратах (Герман, 1974; Голубкова и др., 2021). При разработке классификации докембрийских микрофоссилий использовался формальный подход, что определялось дефицитом биологической информации, связанной с неполной сохранностью ископаемого материала (Микрофоссилии…, 1989 и др.).
Несмотря на длительную историю изучения докембрийских микрофоссилий, большинство выявленных таксонов до сих пор не находят своего места в систематике современных организмов. Особенно остро стоит проблема с вендскими (эдиакарскими) акантоморфными акритархами. Эти ископаемые микроорганизмы сопоставляются с современными динофлагеллятами или с зигоспорами зеленых водорослей (Moczydłowska et al., 2011 и др.), отдельные таксоны или эдиакарские биоты в целом интерпретируются как цисты яиц многоклеточных морских беспозвоночных животных (Yin et al., 2007; Cohen et al., 2009; Sergeev et al., 2011; Воробьева, Сергеев, 2014 и др.) или как гигантские серные бактерии, близкие к современному роду Thiomargarita Schulz, Brinkhoff, Ferdelman, Mariné, Teske et Jorgensen (Bailey et al., 2007). Некоторые из предлагаемых интерпретаций были позднее поставлены под сомнение. Так, изучение морфологии и процессов постмортального разложения современных бактерий Thiomargarita sp. не подтвердило вывод о сопоставлении микрофоссилий вида Tianzhushania ornata (Xiao et Knoll), emend. Yin, Bengtson et Yue с серными бактериями (Cunningham et al., 2011).
Вендские (эдиакарские) акантоморфные биоты распространены в интервале от 630 до 540 млн лет назад (Liu et al., 2013; Голубкова, Кузнецов, 2014; Голубкова и др., 2015; Zhou et al., 2019; Grazhdankin et al., 2020; Xiao et al., 2022 и др.). Они отличаются высоким морфологическим разнообразием, а их размеры колеблются в интервале от 30 до 1000 мкм. Такой разброс данных скорее указывает на гетерогенную природу микрофоссилий, а это предполагает, что выводы авторов могут быть вполне корректны применительно к отдельным таксонам, но не к группе эдиакарских акантоморфит в целом.
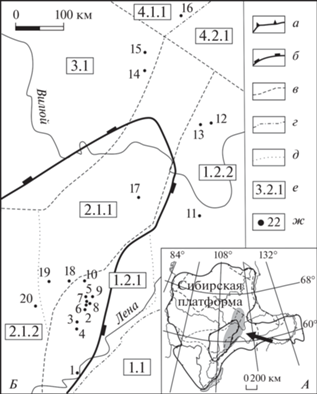
В настоящей статье представлены результаты изучения новых акантоморфных акритарх, обнаруженных в терригенных отложениях венда внутренних районов Сибирской платформы (рис. 1).
Рис. 1.
Район исследований (А) и местоположение изученных скважин (Б) на схеме структурно-фациального районирования Сибирской платформы (Стратиграфия…, 2005): а – граница Сибирской платформы; б – граница Непско-Ботуобинской антеклизы; в – граница фациальных регионов; г – граница фациальных районов; д – граница фациальных зон; е – индексы фациальных регионов, районов, зон: 1 – Байкало-Патомского регион, 1.1 – Патомский район, 1.2 – Предпатомский район, 1.2.1 – Нюйско-Пелейдуйская зона, 1.2.2 – Вилючанская зона, 2.1 – Непско-Ботуобинский район Катангско-Ботуобинского региона, 2.1.2 – Ботуобинская зона, 2.1.2 – Приленско-Непская зона, 3.1 – Сюгджерский район Турухано-Сюгджерского региона, 4 – Анабаро-Алданский регион, 4.1.1 – Куонамская зона Анабарского района, 4.2.1 – Синская зона Алданского района; ж – положение скважин: 1 – Чайкинская-367; Талаканская площадь: 2 – 806, 3 – 823, 4 – 826; Нижнехамакинская плошадь: 5 – 841, 6 – 843, 7 – 845, 8 – 848, 9 – 849, 10 – Озерная-761, 11 – Вилюйско-Джербинская-642, 12 – Меикская-2231, 13 – Средне-Ыгыаттинская-2630, 14 – Дюданская-2910, 15 – Накынская-2950, 16 – Эйикская-3430, 17 – Северо-Юрегинская-1591, 18 – Пелейдуйская-750, 19 – Западная-741, 20 – Верхнечонская-96.
Автор выражает искреннюю признательность Б.Б. Кочневу за предоставленные образцы керна по скважинам Предпатомского и Сюгджерского районов. Исследования проведены в рамках темы НИР ИГГД РАН № FMUW-2021-0003 и при использовании оборудования Центра коллективного пользования аналитических исследований ранней истории земли (ЦКП АИРИЗ) ИГГД РАН, С.-Петербург.
ГЕОЛОГИЧЕСКИЙ ОБЗОР И ПАЛЕОНТОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА ОТЛОЖЕНИЙ ВЕНДА
Разнообразные по таксономическому составу ассоциации акантоморфных микрофоссилий обнаружены в 20 скважинах, пробуренных в Предпатомском, Непско-Ботуобинском, Сюгджерском и Анабарском структурно-фациальных районах (Волкова и др., 1980; Рудавская, Васильева, 1989; Колосова, 1991; Moczydłowska et al., 1993; Голубкова и др., 2010; Наговицин, Кочнев, 2015; Голубкова, Кочнев, 2018 и др.) (рис. 1).
Осадочная последовательность венда внутренних районов Сибирской платформы сложена ледниковыми образованиями конгломератовой и доломитовой (cap-dolomites) толщ, которые выше по разрезу перекрываются терригенными отложениями талаканской свиты, вилючанского и непского горизонтов, а также карбонатными породами тирского и даниловского горизонтов (Решения…, 1989; Стратиграфия…, 2005; Кочнев и др., 2015; Мельников, 2018) (рис. 2). Вилючанский горизонт с перерывом залегает на талаканской свите или породах фундамента и также с несогласием перекрывается непским горизонтом. К вилючанскому горизонту отнесены бетичинская и хоронохская свиты. Непский горизонт расчленяется на два подгоризонта, которые имеют ритмичное строение. Нижняя часть каждого ритма сложена песчаниками, а верхняя – аргиллитами и алевролитами. В Вилючанской зоне Предпатомского р-на к нижнему подгоризонту отнесены талахская, бесюряхская и ынахская свиты, а к верхнему – харыстанская свита. Этим стратиграфическим подразделениям в Нюйско-Пеледуйской зоне отвечают талахская и паршинская свиты, на востоке Сюгджерского и юге Анабарского р-нов – талахская и харыстанская свиты, в Ботуобинской зоне – талахская и курсовская свиты, а на востоке Приленско-Непской зоны – талахская и паршинская свиты. Тирский горизонт согласно залегает на породах непского горизонта и расчленяется на терригенную нижнюю подсвиту бюкской свиты и карбонатную верхнюю подсвиту бюкской свиты. Граница венда–кембрия проведена внутри даниловского горизонта по появлению мелкораковинной фауны зоны Nochoroicyathus sunnaginicus (Хоментовский и др., 1998; Стратиграфия…, 2005). Наиболее мощные и стратиграфически полные разрезы вскрыты на восточном склоне Непско-Ботуобинской антеклизы и в Предпатомском прогибе (рис. 2).
Рис. 2.
Положение микрофоссилий в Региональной стратиграфической схеме венда внутренних районов Сибирской платформы (Стратиграфия…, 2005). Обозначения: 1 – конгломераты и гравелиты, 2 – песчаники, 3 – алевролиты, 4 – аргиллиты, 5 – ангидрито-доломиты, 6–18 – микрофоссилии: 6 – Leiosphaeridia sp., 7 – Appendisphaera sp., 8 – ?Appendisphaera tabifica, 9 – Cavaspina acuminata, 10 – Tanarium sp., 11 – Tuberculum proteum, 12 – Talakania obscura, 13 – Nepia calycina, 14 – Hamakinia sp., 15 – Siphonophycus sp, 16 – Oscillatoriopsis sp., 17 – Obruchevella sp., 18 – Vanavarataenia insolita, 19 – интервал распространения акантоморфных акритарх. Сокращения: ОСШ – Общая стратиграфическая шкала, РСШ – Региональная стратиграфическая шкала, kt – конгломератовая толща, dt – доломитовая толща, tlk – талаканская свита, bt – бетичинская свита, hrn – хоронохская свита, tl – талахская свита, kr1 – нижняя подсвита курсковской свиты, kr2 – верхняя подсвита курсковской свиты, pr1 – нижняя подсвита паршинской свиты, pr2 – верхняя подсвита паршинской свиты, bs – бесюряхская свита, in – ынахская свита, hrs – харыстанская свита, bk1 – нижняя подсвита бюкской свиты, bk2 – верхняя подсвита бюкской свиты.
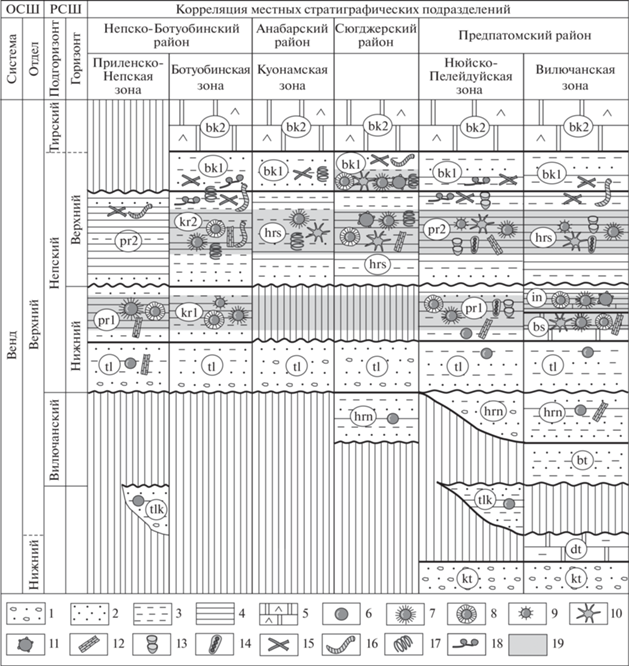
В талаканской свите, вилючанском горизонте и в нижней части нижнего подгоризонта непского горизонта распространены морфологически просто устроенные, транзитные микрофоссилии Leiosphaeridia crassa (Naumova) emend. Jankauskas, L. minutissima (Naumova) emend. Jankauskas, L. tenuissima Eisenack и Siphonophycus Schopf emend. Knoll et Swett. На отдельных стратиграфических уровнях в глинистых прослоях обнаружены нитчатые цианобактерии Talakania obscura Kolosov emend. Golubkova.
Средняя часть непского горизонта охарактеризована таксономически разнообразными ассоциациями, содержащими эдиакарские акантоморфные акритархи. Наиболее богатые биоты установлены в непском горизонте Предпатомского района и на востоке Сюгджерского района (рис. 2). Отсюда определены: акантоморфные акритархи Appendisphaera grandis (Moczydłowska, Vidal et Rudavskaya) emend. Moczydłowska, A. minima Nagovitsin et M.Faizullin (табл. VII , фиг. 3; см. вклейку), A. tenuis (Moczydłowska, Vidal et Rudavskaya) emend. Moczydłowska (табл. VII , фиг. 2), ?Appendisphaera tabifica (Moczydłowska, Vidal et Rudavskaya) emend. Moczydłowska (табл. VII , фиг. 4 ), ?Asseserium fusulentum Moczydłowska et Nagovitsin, Cavaspina acuminata (Kolosova) emend. Moczydłowska, Vidal et Rudavskaya, emend. Moczydłowska (табл. VII , фиг. 5 ), Ceratosphaeridium glaberosum Grey, Densisphaera arista Moczydłowska et Nagovitsin, Tanarium araithekum (Zang in Zang et Walter) emend. Grey, T. conoideum (Kolosova) emend. Moczydłowska, Vidal et Rudavskaya, emend. Moczydłowska (табл. VII , фиг. 6 ), T. longidigitatum sp. nov. (табл. VII , фиг. 9–11 ), T. paucispinosum Grey, T. tuberosum (Moczydłowska, Vidal et Rudavskaya) emend. Moczydłowska, Vidal et Rudavskaya (табл. VII , фиг. 8 ), Knollisphaeridium maximum (Yin) emend. Knoll, emend. Willman (табл. VII , фиг. 7 ), Mengeosphaera triangularis Liu, Xiao, Yin, Chen, Zhou et Li, Uniexcrescentia tubulosa gen. et sp. nov. (табл. VIII , фиг. 5–7 ; см. вклейку), гладкостенные оболочки Leiosphaeridia crassa, L. jacutica (Timofeev) emend. Mikhailova et Jankauskas, L. minutissima, L. tenuissima, Cucumiforma vanavaria Mikhailova, Pterospermopsimorpha insolita (B.V. Timofeev) emend. Mikhailova (табл. VIII , фиг. 8 ), P. mutabilis sp. nov. (табл. VIII , фиг. 9–12 ), овальные формы Navifusa Combaz, Lange et Pansart., цианобактерии Nepia calycina Golubkova, Oscillatoriopsis Schopf emend. Knoll et Golubic, emend. Knoll, Swett et Mark, emend. Butterfield, Siphonophycus sp., Talakania obscura, грибоподобные организмы Caudosphaera expansa Hermann et Timofeev и проблематики Hamakinia Rudavskaya. В верхнем подгоризоте непского горизонта Сюгджерского района дополнительно обнаружены новые акантоморфные акритархи Tuberculum proteum sp. nov. (табл. VIII , фиг. 1–4 ) и спирально свернутые цианобатерии Obruchevella Reitlinger emend. Yakschin et Luchinina. В Анабарском и Непско-Ботуобинском районах наблюдается постепенное сокращение таксономического разнообразия микрофоссилий (рис. 2). Из состава ассоциаций исчезают представители родов Densisphaera, Knollisphaeridium, Mengeosphaera, Tanarium, Nepia, Hamakinia, а широкое распространение на юго-западе и севере изученной площади получают виды Appendisphaera grandis (табл. VII , фиг. 1), A. tenuis, ?A. tabifica и Talakania obscura.
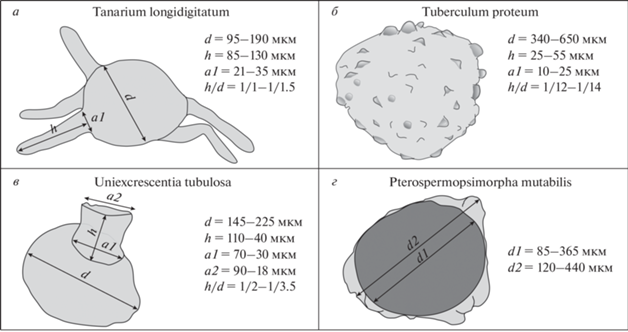
Рис. 3.
Морфологическое строение и основные параметры размеров новых акритарх: а – Tanarium longidigitatum, б – Tuberculum proteum, в – Uniexcrescentia tubulosa, г –Pterospermopsimorpha mutabilis. Обозначения: d – диаметр оболочки, h – длина выроста, a1 – ширина основания выроста, a2 – ширина окончания выроста, d1 – диаметр внутреннего тела, d2 – диаметр внешней оболочки.
Выше по разрезу, в переходных отложениях непского – нижней части тирского горизонтов отмечается заметное сокращение видового разнообразия микрофоссилий. Из состава биот постепенно исчезают акантоморфные и некоторые другие характерные таксоны, а широкое, но кратковременное распространение получают цианобактерии Obruchevella sp., Oscillatoriopsis sp., Siphonophycus и грибоподобные организмы Vanavarataenia insolita Pjatiletov (рис. 2). Исключение составляет разрез скв. Дюданская-2910, вскрытый бурением на западе Сюгджерского района (рис. 1, 2). В алевролитах нижней подсвиты бюкской свиты скв. Дюданская-2910 обнаружены акритархи Appendisphaera grandis, A. minima, A. tenuis, ?A. tabifica, Ceratosphaeridium glaberosum, Knollisphaeridium maximum, Tanarium conoideum, Tuberculum proteum sp. nov., Leiosphaeridia jacutica, L. crassa, L. tenuissima, L. minutissima, Pterospermopsimorpha insolita, P. mutabilis sp. nov., Navifusa sp. и нитчатые цианобактерии Talakania obscura, Obruchevella sp., Siphonophycus sp. В верхней подсвите бюкской свиты тирского горизонта микрофоссилии выявлены не были.
Далее в работе приведено монографическое описание новых акантоморфных акритарх, отнесенных к четырем формальным родам: Tanarium (Kolosova) emend. Moczydłowska, Vidal et Rudavskaya, Tuberculum Golubkova, Bobrovskiy, Kushim et Plotkina, Uniexcrescentia gen. nov. и Pterospermopsimorpha B.V. Timofeev emend. Mikhailova et Jankauskas (рис. 3).
INCERTAE SEDIS
ГРУППА МИКРОФОССИЛИИ
ПОДГРУППА ACRITARCHA EVITT, 1963
Род Tanarium (Kolosova, 1991) emend. Moczydłowska, Vidal et Rudavskaya, 1993
Tanarium longidigitatum Golubkova, sp. nov.
Табл. VII , фиг. 9–11
Название вида от longus лат. – длинный и digitatus лат. – пальцевидный.
Голотип – ИГГД РАН, № II-2; кол. № ВС-2010-НХ848; Россия, Восточная Сибирь, Нюйско-Пеледуйская зона Предпатомского р-на, скв. Нижне-хамакинская-848, гл. 1571.5 м, обр. 22, преп. 3766/1; верхний венд, непский горизонт, верхняя подсвита паршинской свиты; обозначен здесь, табл. VII , фиг. 10 .
Diagnosis. Large spherical smooth-walled vesicle, bearing irregularly spaced homogeneous processes. Processes are finger-shaped, long, smoothly rounded at the ends, hollow and communicate openly with vesicle interior. The observed number of processes on the vesicle is from 4 to 12. Vesicles 95–190 μm in diameter. Processes 85–130 μm in length, 21–35 μm in basal width. The ratio of the length of the processes to the diameter of the vesicle is 1/1–1/1.5.
Описание (рис. 3, а). Большие, сферические гладкостенные оболочки, орнаментированные нерегулярно расположенными, гомогенными, полыми выростами, открытыми во внутреннюю полость. Выросты пальцевидной формы, длинные, плавно закругленные на окончаниях. У наименее деформированных экземпляров ширина выроста постоянная на всем его протяжении. Наблюдаемое количество выростов на оболочке – от 4 до 12. Диаметр оболочек у разных экземпляров составляет от 95 до 190 мкм, длина выростов изменяется от 85 до 130 мкм, ширина основания выростов – от 21 до 35 мкм. Те же параметры у голотипа составляют соответственно 120–150 мкм, 100–120 мкм и 25 мкм. Отношение длины выростов к диаметру оболочки – 1/1–1/1.5.
Сравнение. Новый таксон отличается от других видов рода Tanarium длинными, одинаковыми по ширине и плавно закругленными на окончаниях выростами.
Распространение. Сибирская платформа, Нюйско-Пеледуйская и Вилючанская зоны Предпатомского р-на; верхний венд, непский горизонт, паршинская и харыстанская свиты.
Материал. 39 целых и частично разрушенных экз. хорошей и удовлетворительной сохранности из скв. Нижне-хамакинская-848 (гл. 1571.5 м) и Вилюйско-Джербинская-642 (инт. 2173.1–2169.9 м).
Род Tuberculum Golubkova, Bobrovskiy, Kushim et Plotkina, 2021
Tuberculum proteum Golubkova, sp. nov.
Табл. VIII , фиг. 1–4
Название вида от proteus лат. – переменчивый, непостоянный.
Голотип – ИГГД РАН, № II-3; кол. № ВС-2010-Н2950; Россия, Восточная Сибирь, Сюгджерский р-н, скв. Накынская-2950, гл. 3062.5 м, обр. 2950-2, преп. 2/1; верхний венд, непский горизонт, харыстанская свита; обозначен здесь, табл. VIII , фиг. 1.
Diagnosis. Large spherical vesicle with irregularly arranged pimples–tubercles. Pimples are communicate openly with vesicle interior. The ends of the processes are wide, smoothly rounded. Vesicles 340–650 μm in diameter. Processes 25–55 μm in length, 10–25 μm in basal width.
Описание (рис. 3, б). Крупные гладкостенные сферические оболочки, орнаментированные нерегулярно расположенными выростами–бугорками, свободно сообщающимися с полостью оболочки. Окончания выростов широкие, плавно закругленные. Внутри выростов наблюдаются темные, бесструктурные уплотнения сферической формы, повторяющие их контуры. Диаметр оболочки 340–650 мкм (у голотипа – 480 мкм). Длина выростов в среднем составляет 25–55 мкм, ширина их основания – 10–25 мкм. Для данного таксона характерен высокий полиморфизм, проявляющийся в степени выраженности, размерах и количестве выростов на поверхности оболочек. В изученном материале присутствуют экземпляры от почти гладких, неявно орнаментированных форм, выросты которых лишь намечаются за счет приподнятых участков стенки оболочки, до экземпляров, выросты которых сформированы в отчетливо выраженные бугорки.
Сравнение. От выделенного ранее вида T. mamillatum Golubkova, Bobrovskiy, Kushim et Plotkina новый таксон отличается отсутствием характерного сосочковидного окончания на вершине выроста.
Распространение. Сибирская платформа, Сюгджерский р-н, Вилючанская зона Предпатомского р-на, Приленско-Непская зона Непско-Ботуобинского р-на; верхний венд, ынахская, харыстанская и паршинская свиты непского горизонта, нижняя подсвита бюкской свиты тирского горизонта.
Материал. 64 целых и частично разрушенных экз. хорошей и удовлетворительной сохранности из скв. Накынская-2950 (гл. 3062.5 м), Дюданская-2910 (инт. 3415.3–3414.8 м), Средне-Ыгыаттинская-2230 (гл. 3421.4) и Верхнечонская-96 (гл. 1646.5).
Род Uniexcrescentia Golubkova, gen. nov.
Название рода от uni- лат. – один, одиночный, и excrescentia лат. – вырост.
Типовой вид – Uniexcrescentia tubulosa sp. nov.
Диагноз. Сферические оболочки с одиночным полым крупным выростом, открытым во внутреннюю полость. Вырост цилиндрической формы.
Diagnosis. Spherical vesicle bearing a single large process. Process are cylindrical shape, hollow inside and freely communicate with vesicle cavity.
Видовой состав. Типовой вид.
Сравнение. От рода Ceratosphaeridium Grey описываемые новые формы отличаются характерной цилиндрической формой и значительно более крупным размером выроста. В отличие от австралийского рода Sticcasphaeridium Zang et Walter новый таксон имеет одиночный крупный вырост в форме трубки.
Uniexcrescentia tubulosa Golubkova, sp. nov.
Табл. VIII , фиг. 5–7
Название вида от tubulosus лат. – трубчатый.
Голотип – ИГГД РАН, № II-4; кол. № ВС-2010-НХ849; Россия, Восточная Сибирь, Нюйско-Пелейдуйская зона Предпатомского р-на, скв. Нижнехамакинская-849, гл. 1605 м, обр. 20, преп. 3761/1; верхний венд, непский горизонт, верхняя подсвита паршинской свиты; обозначен здесь, табл. VIII , фиг. 7 .
Diagnosis. Spherical smooth-walled vesicle with a single process freely communicating with the vesicle cavity. Process are large, cylindrical shape. Vesicles 145–225 μm in diameter. Process 40–110 μm in length, 30–70 μm wide at the base of the process, 18–90 µm wide at the end of the process. The ratio of the length of the processes to the diameter of the vesicle is 1/2–1/3.5.
Описание (рис. 3, в). Сферические гладкостенные оболочки с одиночным выростом, свободно сообщающимся с внутренней полостью. Вырост крупный, цилиндрической формы. Окончание выроста без сужения, предположительно, открыто во внешнюю среду. Параметры размеров у голотипа составляют: диаметр оболочки (d) 225 мкм, длина выроста (h) – 110 мкм, ширина основания выроста (а1) – 70 мкм, ширина окончания выроста (а2) – 90 мкм, отношение длины выроста к длине оболочки (h/d) – 1/2. Эти же параметры для второй и третьей оболочек соответственно: d – 155 мкм, h – 60 мкм, а1 – 35 мкм, а2 – 25 мкм, h/d – 1/2.5 и d – 145 мкм, h – 40 мкм, а1 – 30 мкм, а2 – 18 мкм, h/d – 1/3.5.
Распространение. Сибирская платформа, Нюйско-Пелейдуйская зона Предпатомского р-на, Ботуобинская зона Непско-Ботуобинского р-на; верхний венд, паршинская свита непского горизонта.
Материал. 3 целых экз. хорошей сохранности из скв. Нижнехамакинская-849 (гл. 1605 м), Пелейдуйская-750 (гл. 1836 м) и Чайкинская-367 (гл. 1419.7 м).
Род Pterospermopsimorpha B.V. Timofeev, 1966, emend. Mikhailova et Jankauskas, 1989
Pterospermopsimorpha mutabilis Golubkova, sp. nov.
Табл. VIII , фиг. 9–12
Название вида mutabilis лат. – изменчивый, непостоянный.
Голотип – ИГГД РАН, № II-5; кол. № ВС-2010-О-761; Россия, Восточная Сибирь, Нюйско-Пелейдуйская зона Предпатомского р-на, скв. Озерная-761, гл. 1879 м, обр. 210, преп. 1953/4; верхний венд, непский горизонт, нижняя подсвита паршинской свиты; обозначен здесь, табл. VIII , фиг. 9 .
Diagnosis. Vesicles are mooth-walled and consist of an inner body and an outer shell. The inner body 85–365 μm in diameter, spherical, smooth, thick, the outer contour is even. The outer shell 120–440 μm in diameter, smooth, thin, elastic. The outer shell arbitrarily adjoins the inner body, forming smoothly rounded hemispheres.
Описание (рис. 3, г). Гладкостенные формы, состоящие из внутреннего тела и внешней оболочки. Внутреннее тело сферической формы, гладкое, толстое, внешний контур четкий, ровный. Диаметр внутреннего тела у разных экземпляров изменяется от 85 до 365 мкм, этот же параметр у голотипа составляет 260 мкм. Внешняя оболочка гладкая, тонкая, эластичная, произвольно прилегает к внутреннему телу, образуя разнообразные по очертаниям плавно закругленные полусферы. Диаметр внешней оболочки составляет 120–440 мкм, у голотипа – 325 мкм.
Сравнение. От других видов рода Pterospermopsimorpha (табл. VIII , фиг. 8 ) описываемые новые формы отличаются невыдержанным внешним краем оболочки, свободно прилегающим к внутреннему телу.
Распространение. Восточная Сибирь, Сюгджерский р-н, Вилючанская зона Предпатомского р-на, Приленско-Непская и Ботуобинская зоны Непско-Ботуобинского р-на; верхний венд, нижняя подсвита бюкской свиты тирского горизонта, бесюряхская, ынахская, харыстанская, курсовская и паршинская свиты непского горизонта.
Материал. Десятки целых и частично разрушенных экземпляров хорошей сохранности из скважин: Эйикская-3430 (инт. 2241.3–2235.3 м), Накынская-2950 (гл. 3062.5 м), Дюданская-2910 (гл. 3414.8 м), Меикская-2231 (инт. 4669.8–4666.35 м), Средне-Ыгыаттинская-2230 (инт. 3426.4–3420.7 м), Северо-Юрегинская-1591 (инт. 1962.3–1958.1 м), Пелейдуйская-750 (гл. 1836 м), Западная-741 (гл. 1820 м), Талаканская-806 (инт. 1584.2–1467 м), Талаканская-823 (гл. 1536 м), Талаканская-826 (гл. 1560.8 м), Нижнехамакинская-841 (инт. 1572.9–1570.5 м), Нижнехамакинская-843 (инт. 1508.7–1499.4 м), Нижнехамакинская-845 (инт. 1571.7–1567.8 м), Озерная-761 (инт. 1881–1867 м).
ЗАКЛЮЧЕНИЕ
Анализ палеонтологических данных позволил расширить наше представление о морфологическом разнообразии вендских (эдиакарских) акантоморфных акритарх. Близкие по таксономическому составу ассоциации установлены на трех стратиграфических уровнях – в нижнем и верхнем подгоризонтах непского горизонта и в нижней части тирского горизонта. Находка акантоморфной биоты непского типа в нижней подсвите бюкской свиты Сюгджерского горизонта указывает на возможность отнесения терригенной части тирского горизонта к непскому горизонту.
В работе проведено монографическое описание нового рода и четырех новых видов акантоморфных и гладкостенных акритарх: Tanarium longidigitatum sp. nov., Tuberculum proteum sp. nov., Uniexcrescentia tubulosa gen. et sp. nov., Pterospermopsimorpha mutabilis sp. nov. Выявленные таксоны имеют характерный облик и относительно широкое латеральное распространения в разрезах венда внутренних районов Сибирской платформы, что может быть использовано в стратиграфических, палеоэкологических и палеофациальных построениях.
Список литературы
Волкова Н.А., Кирьянов В.В., Пятилетов В.Г. и др. Микрофоссилии верхнего докембрия Сибирской платформы // Изв. АН СССР. Сер. геол. 1980. № 1. С. 23–29.
Воробьева Н.Г., Сергеев В.Н. Радикальные изменения в биосфере при переходе от позднего рифея к венду // Водоросли в эволюции биосферы / Ред. Рожнов С.В. М.: ПИН РАН, 2014. С. 6–36 (Сер. “Гео-биологические процессы в прошлом”).
Герман Т.Н. Из опыта извлечения крупных растительных остатков и микрофоссилий с помощью химического растворения пород // Микрофоссилии СССР. Новосибирск: Наука, 1974. С. 97–99 (Тр. ИГиГ СО АН СССР. Вып. 81).
Голубкова Е.Ю., Зайцева Т.С., Кузнецов А.Б. и др. Микрофоссилии и Rb-Sr возраст глауконитов в опорном разрезе верхнего протерозоя северо-востока Русской плиты (скв. Кельтменская-1) // Докл. Акад. наук. 2015. Т. 462. № 4. С. 444–448.
Голубкова Е.Ю., Кочнев Б.Б. Органостенные микрофоссилии в верхнедокембрийских отложениях внутренних районов Сибирской платформы // Эволюция вещественного и изотопного состава докембрийской литосферы / Ред. Глебовицкий В.А., Балтыбаев Ш.К. СПб.: Издательско-полиграфическая ассоциация высших учебных заведений, 2018. С. 129–144.
Голубкова Е.Ю., Кузнецов А.Б. Таксономическое разнообразие и корреляция эдиакарских комплексов акантоморфной палинофлоры мира // Водоросли в эволюции биосферы / Ред. Рожнов С.В. М.: ПИН РАН, 2014. С. 119–140 (Сер. “Гео-биологические процессы в прошлом”).
Голубкова Е.Ю., Кузьменкова О.Ф., Кушим Е.А. и др. Распространение микрофоссилий в отложениях венда Оршанской впадины Восточно-Европейской платформы (Беларусь) // Стратигр. Геол. корреляция. 2021. Т. 29. № 6. С. 24–38.
Голубкова Е.Ю., Раевская Е.Г., Кузнецов А.Б. Нижневендские комплексы микрофоссилиий Восточной Сибири в решении стратиграфических проблем региона // Стратигр. Геол. корреляция. 2010. Т. 18. № 4. С. 3–27.
Колосова С.П. Позднедокембрийские шиповатые микрофоссилии востока Сибирской платформы // Альгология. 1991. Т. 1. № 2. С. 53–59.
Кочнев Б.Б., Покровский Б.Г., Прошенкин А.И. Верхненеопротерозойский гляциокомплекс центральных областей Сибирской платформы // Докл. Акад. наук. 2015. Т. 464. № 4. С. 448–451.
Мельников Н.В. Венд-кембрийский соленосный бассейн Сибирской платформы (Стратиграфия, история развития). Изд. 2-е. Новосибирск: СНИИГГиМС, 2018. 177 с.
Микрофоссилии докембрия СССР / Ред. Янкаускас Т.В. Л.: Наука, 1989. 190 с.
Наговицин К.Е., Кочнев Б.Б. Микрофоссилии и биофации вендской ископаемой биоты юга Сибирской платформы // Геол. и геофизика. 2015. Т. 56. № 4. С. 748–760.
Решения четвертого межведомственного регионального совещания по уточнению и дополнению стратиграфических схем венда и кембрия внутренних районов Сибирской платформы. Новосибирск: СНИИГГиМС, 1989. 40 с.
Рудавская В.А., Васильева Н.И. Талсинский комплекс акритарх Непско-Ботуобинской антеклизы // Фитостратиграфия и морфология спор древних растений нефтегазоносных провинций СССР. Л.: ВНИГРИ, 1989. С. 5–11.
Стратиграфия нефтегазоносных бассейнов Сибири. Рифей и венд Сибирской платформы и ее складчатого обрамления / Ред. Конторович А.Е. Новосибирск: “Гео”, 2005. 432 с.
Хоментовский В.В., Федоров А.Б., Карлова Г.А. Нижняя граница кембрия во внутренних районах севера Сибирской платформы // Стратигр. Геол. корреляция. 1998. Т. 6. № 1. С. 3–11.
Bailey J.V., Joye S.B., Kalanetra K.M. et al. Evidence of giant sulfur bacteria in Neoproterozoic phosphorites // Nature. 2007. V. 445. P. 198–201.
Cohen P.A., Knoll A.H., Kodner R. Large spinose microfossils in Ediacaran rocks as resting stages of early animals // Proc. Nat. Acad. Sci. USA. 2009. V. 106. P. 6519–6524.
Cunningham J.A., Thomas C.-W., Bengtson S. et al. Experimental taphonomy of giant sulphur bacteria: implications for the interpretation of the embryo-like Ediacaran Doushantuo fossils // Proc. R. Soc. B. 2011. V. 279. P. 1857–1864.
Grazhdankin D., Nagovitsin K., Golubkova E. et al. Doushantuo-Pertatataka-type acanthomorphs and Ediacaran ecosystem stability // Geology. 2020. V. 48. P. 708–712.
Liu P., Yin C., Chen S. et al. The biostratigraphic succession of acanthomorhic acritarchs of the Ediacaran Doushantuo Formation in Yangtze Gorges, South China and its biostratigraphic correlation with Australia // Precambrian Res. 2013. V. 225. P. 29–43.
Moczydłowska M., Landing E., Zang W., Palacios T. Proterozoic phytoplankton and timing of Chlorophyta algae original // Palaeontology. 2011. V. 54. P. 721–733.
Moczydłowska M., Vidal G., Rudavskaya V.A. Neoproterozoic (Vendian) phytoplankton from the Siberian Platform, Yakutia // Palaeontology. 1993. V. 36. P. 495–521.
Sergeev V.N., Knoll A.N., Vorob’eva N.G. Ediacaran microfossils from the Ura Formation, Baikal-Patom Uplift, Siberian: taxonomy and biostratigraphic significance // J. Paleontol. 2011. V. 85. № 5. P. 987–1011.
Xiao S., Jiang G., Ye Q. et al. Systematic paleontology, acritarch biostratigraphy, and δ13C chemostratigraphy of the early Ediacaran Krol A Formation, Lesser Himalaya, northern India // J. Paleontol. 2022. P. 1–62. https://doi.org/10.1017/jpa.2022.7
Yin L., Zhu M., Knoll A.H. et al. Doushantuo embryos preserved inside dipause egg cyst // Nature. 2007. V. 446. P. 661–663.
Zhou C., Yuan X., Xiao S. et al. Ediacaran integrative stratigraphy and timescale of China // Sci. China: Earth Sci. 2019. V. 62. № 1. P. 7–24.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал