

Палеонтологический журнал, 2023, № 1, стр. 74-83
Первая находка овцебыка рода Megalovis (Artiodactyla, Bovidae) в нижнем плейстоцене пещеры Таврида в Крыму
И. А. Вислобокова *
Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
* E-mail: ivisl@paleo.ru
Поступила в редакцию 15.07.2022
После доработки 30.08.2022
Принята к публикации 30.08.2022
- EDN: FKXZSO
- DOI: 10.31857/S0031031X23010142
Аннотация
На основе новых материалов установлено присутствие Megalovis latifrons Schaub, 1923 в раннеплейстоценовой фауне пещеры Таврида в Крыму, возраст которой, по данным биохронологии, составляет 1.8–1.5 млн лет. Дано описание фрагмента лобной кости с частью рогового стержня, неполных верхнечелюстной и нижнечелюстной костей, а также пястной и плюсневой костей. Редкие находки этого крупного ископаемого представителя Ovibovini были известны лишь из нижнего плейстоцена юга Западной и Центральной Европы. Мегаловисы обитали в Евразии во второй половине раннего и самом начале среднего плейстоцена. В России они найдены впервые. Находка в Крыму расширяет представления об эволюции, а также географическом и стратиграфическом распространении этого вида.
ВВЕДЕНИЕ
Пещера Таврида у пос. Зуя к востоку от Симферополя – крупное местонахождение плейстоценовых позвоночных, открытое в 2018 г. при прокладке федеральной автотрассы “Таврида” (Лопатин и др., 2019). Среди материалов, полученных в 2021 г., присутствуют остатки ископаемого рода Megalovis. Они найдены в основном слое с фауной, возраст которой, по биостратиграфическим данным, – 1.8–1.5 млн лет. На территории России мегаловисы найдены впервые.
Род Megalovis Schaub, 1923 относится к подсемейству Caprinae Gill, 1872 семейства Bovidae Gray, 1821 и обычно включается в состав трибы Ovibovini Gray (овцебыков). Он отличается от всех других родов этой трибы очень крупными размерами, особенностями строения черепа, роговых стержней, зубов и костей конечностей. Этот род известен по очень редким находкам из плейстоцена Евразии в интервале примерно от 2.2 до 0.7 млн л. н.
В Европе род Megalovis представлен двумя видами: M. latifrons Schaub, 1923 и M. balcanicus Crégut-Bonnoure et Dimitrijević, 2006. Остатки M. latifrons найдены в трех местонахождениях: Сенез во Франции (Schaub, 1923, 1937, 1943) (типовое местонахождение), Эрпфингер Хёле в Германии (Lehmann, 1957) и Фынтына луи Митилан в Румынии (Radulesco, Samson, 1962). M. balcanicus известен лишь из Трлицы в Черногории (Crégut-Bonnoure, Dimitrijević, 2006). Кроме того, остатки мегаловисов присутствуют в местонахождениях Болгарии (Сливница, Варшец и пещера Козарника) (Spassov, Crégut-Bonnoure, 1999; Spassov, 2005; Crégut-Bonnoure, 2007; Fernandez, Crégut, 2007) и Италии (Пирро Норд) (Ovibovini: De Giuli et al., 1986, 1990; Crégut-Bonnoure, 2002; Masini, Sala, 2007).
В Азии род Megalovis представлен тремя видами, найденными на территории Китая: M. piveteaui Schaub, 1937, Нихэвань, Хэбэй (Schaub 1937; Tong et al., 2021, 2022), M. wimani Schaub, 1937, Ян-Шао-Цун (Loc. A), Сангкан-хо, Хэнань (Schaub, 1937), и M. guangxiensis Han, 1987, Лючен, Гуанси (Han, 1987). Остатки последнего присутствуют в ряде местонахождений Южного Китая (в основном, южнее р. Янцзы), часто вместе с Gigantopithecus (e.g., Dong et al., 2020; Dong, Bai, 2021).
Диагностика видов M. latifrons и M. balcanicus основывается на особенностях строения лобной кости и положения роговых стержней, а также на размерах и некоторых особенностях щечных зубов. Китайские виды различаются размерами и особенностями щечных зубов и пястных костей.
Сведения о морфологии Megalovis довольно скудны, и родственные связи не очень ясны. Находки остатков этих “гигантских овец” в Тавриде и результаты их изучения представляют большой интерес, так как позволяют получить новые данные для уточнения эволюции и распространения этого рода, а также расширяют представления о биоразнообразии раннеплейстоценовой фауны Тавриды и региона в целом.
Автор выражает благодарность А.В. Лаврову (Палеонтологический ин-т им. А.А. Борисяка РАН, ПИН), Д.О. Гимранову (Ин-т экологии растений и животных УрО РАН) и всем участникам раскопок за сбор материалов в 2021 г.; д-рам Л. Костеру (Музей естественной истории, г. Базель), и Вей Донгу (Ин-т палеонтологии позвоночных и палеоантропологии АН Китая, г. Пекин) за присланные фотографии образцов и статьи; а также проф. Н. Спассову (Национальный музей естественной истории, г. София) за полезную информацию о находках в Болгарии и проф. В. Димитриевич (Ун-т Белграда) о находках в Трлице в Черногории.
ОПИСАНИЕ И СРАВНЕНИЕ МАТЕРИАЛА
СЕМЕЙСТВО BOVIDAE GRAY, 1821
ПОДСЕМЕЙСТВО CAPRINAE GILL, 1872
Триба Ovibovini Gray, 1872
Род Megalovis Schaub, 1923
Megalovis latifrons Schaub, 1923
Megalovis latifrons: Schaub, 1923, с. 292–295, рис. 5; 1937, c. 30; 1943, с. 281, 282, рис. 5–7 ; 1962, c. 262; Crégut-Bonnoure, Dimitrijević, 2006, с. 725; Crégut-Bonnoure, 2007, с. 80; Fernandez, Crégut, 2007, с. 432.
Antilope sp.: Schaub, 1923, c. 291; 1937, с. 28.
Голотип – Музей естественной истории, г. Базель, № NMB, Se 556, почти полный череп с основаниями роговых стержней, изображен в работе С. Шауба (Schaub, 1923, рис. 5); Франция, местонахождение Сенез; нижний плейстоцен (поздний виллафранк).
Описание (рис. 1–3). Фрагмент лобной кости с частью основания рогового стержня экз. ПИН, № 5644/301 спереди обломан чуть позади орбиты, а сзади примерно на уровне середины основания рогового стержня (рис. 1). Лобная кость сильно пневматизирована. Лобная часть лобной кости плоская, четко отделена от наклоненной и вогнутой латеральной части. Основания рогов сравнительно слабо сдвинуты назад. Минимальное расстояние между небольшим сосудистым отверстием (2.5 мм) у аборального конца надглазничного желоба и основанием рога – около 34 мм. Сохранившая часть основания рогового стержня направлена в сторону и немного назад и вверх. Контакт рогового стержня с пеньком хорошо выражен. Он скошен относительно продольной оси черепа и, следовательно, расстояние между передними краями роговых стержней было больше, чем между их задними краями. Костная ткань основания рогового стержня с каналами для крупных сосудов. Довольно широкий лобный синус заходит в основание рогового стержня примерно на высоту 8 мм (рис. 1, в).
Рис. 1.
Megalovis latifrons Schaub, 1923, экз. ПИН, № 5644/301, фрагмент лобной кости с частью основания рогового стержня: а – спереди, б – сверху, в – снизу; Крым, пещера Таврида; нижний плейстоцен.

Рис. 2.
Megalovis latifrons Schaub, 1923: а–в – экз. ПИН, № 5644/302, фрагмент правой верхнечелюстной кости с P3‒М1; г–е – экз. ПИН, № 5644/303, фрагмент правой нижнечелюстной кости с dP3–M2; а, г – с буккальной, б, д – с окклюзиальной, в, е – с лингвальной сторон; Крым, пещера Таврида; нижний плейстоцен.
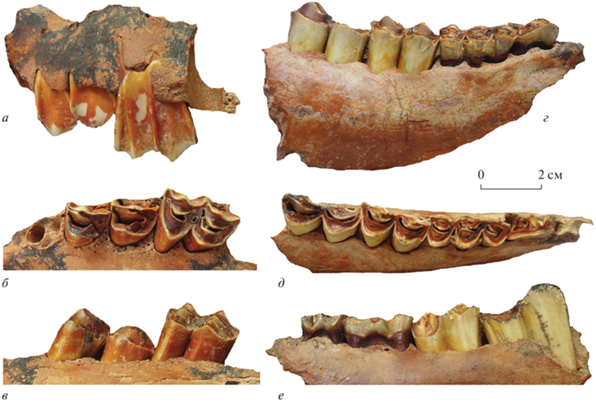
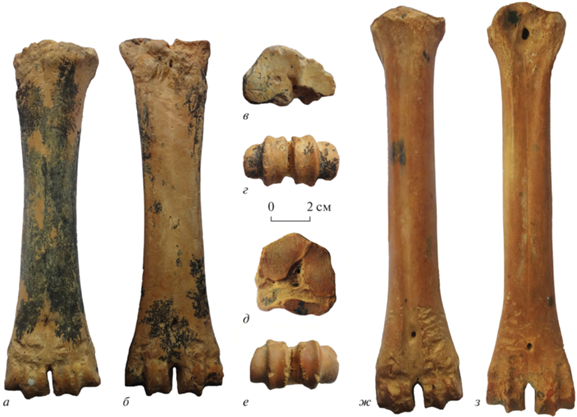
Рис. 3.
Megalovis latifrons Schaub, 1923: а–г – экз. ПИН, № 5644/304, правая пястная кость; д–з – экз. ПИН, № 5644/305, правая плюсневая кость; а, ж – спереди, б, з – сзади, в, д – сверху, г, е – снизу; Крым, пещера Таврида; нижний плейстоцен.
Фрагмент левой стороны лицевого отдела черепа экз. ПИН, № 5644/302 состоит из части верхнечелюстной кости с Р3–М1 и альвеолами Р2 и небольшой части предчелюстной кости (рис. 2). В верхней части образца сохранился широкий желоб (шириной около 6 мм) подглазничного канала, ведущий в подглазничное отверстие, foramen infraorbitale, расположенное на уровне контакта P3 и P4, как у голотипа M. latifrons. Ряд премоляров относительно длинный, его длина (от альвеолы Р2) – ~58.2 мм.
Премоляры P3 и Р4 экз. ПИН, № 5644/302 лишь слегка затронуты стиранием. Коронки довольно высокие (высота P3 – 24.75 мм), немного отклонены назад. Они сужаются к основанию на буккальной стороне, и на поверхности окклюзии спереди немного шире, чем сзади. Передний край коронки P3 слабовыпуклый, а на Р4 он более прямой. Коронка P4 сильнее уплощена лингвально. На буккальной стороне коронок столбик паракона широкий (6–7 мм), на Р3 он слегка сдвинут кпереди, а на Р4 занимает срединное положение. Хорошо развитый парастиль (шириной до 4 мм) и более слабый метастиль (~1.6 мм) немного расширяются к основанию коронок. Нижние концы парастиля и столбика паракона слегка вздуты, сливаются в основании, выемка между ними узкая, V-образная. На лингвальной стороне протокон шире гипокона (сильнее выступает лингвально), их разделяет слабая вертикальная бороздка. В долинке присутствует медиальный гребень (crista medialis), отходящий от протокона и сильно отклоненный назад в направлении к метастилю. Размеры зубов даны в табл. 1.
Таблица 1.
Сравнение промеров щечных зубов видов рода Megalovis1
| Промеры (мм) | M. latifrons | M. latifrons | M. balcanicus | M. piveteaui | M. wimani | M. guangxiensis | |
|---|---|---|---|---|---|---|---|
| Таврида | Сенез | Трлица | Нихэвань | Хэнань | Лючен | Куекуе | |
| P3 L | 17.16 | 16.6–17.2 | 16.6–17.2 | 20* | 17* | ||
| P3 W | 16.41 | 19.3 | |||||
| P4 L | 17.31 | 15.8–16.7 | 16.0–17.8 | 21* | 18* | ||
| P4 W | 15.6–15.8 | 17.4–20.6 | |||||
| M1 L | 25.3 | 22.4–23.1 | 21.7–27.9 | 28* | 27.5* | 23.9–26.9 | 20.6+ |
| M1 W | 21.93 | 19.4 | 20.7–24.5 | 21.3–25.3 | |||
| dP3 L | 14.97 | 20.6–24.3 | 22.3–24.8 | ||||
| dP3 W | 8.92 | 11.4–12.6 | 9.9–11.4 | ||||
| dP4 L | 25.98 | 33.5–35.0 | 34.1–35.8 | ||||
| dP4 W | 11.03 | 14.6–14.8 | 14.8–15.9 | ||||
| M1 L | 24.14 | 21.6–24.1 | 23.6–26.4 | 25 | 24.0–28.4 24* |
21.1–31.1 | |
| M1 W | 13.68 | 15.7–16.28 | 14.2 | 14.0–15.4 15.8* |
13.9–20.4 | ||
| M2 L | 27.36 | 26.4–27.3 | 24.6–29.1 | 30 | 30.0–35.5 28* |
24.9–35.8 | |
| M2 W | 14.88 | 16.9 | 16.1–16.8 | 13.2–18.5 16.0* |
16.1–20.2 | ||
Верхний моляр M1 экз. ПИН, № 5644/302 слабо стерт. Его коронка сравнительно высокая (32.75 мм), индекс гипсодонтности равен 67%. Коронка сужается к основанию на буккальной стороне и расширяется к основанию букколингвально. На поверхности окклюзии задняя половина коронки немного длиннее и заметно уже передней. Переднее лингвальное полулуние более угловатое по сравнению с задним. На буккальной стороне коронки хорошо развиты парастиль, мезостиль и метастиль; из них парастиль наиболее широкий (3.3 мм), а метастиль – наиболее слабый. Парастиль и мезостиль округлены, они чуть вздуты в основании. Столбик паракона прямой, развит сильнее, чем столбик метакона. На поверхности окклюзии парастиль и мезостиль выступающие, столбики паракона и метакона слегка угловатые. Парастиль, столбик паракона и мезостиль немного отогнуты вперед, как и у других представителей Ovibovini.
Тело нижней челюсти экз. ПИН, № 5644/304 заметно утолщено под молярами. Ряд премоляров длинный, его длина (от альвеолы dP2) ~ 52.49 мм. Нижние моляры M1 и M2 с частично разрушенными стенками лингвальных полулуний. Коронки этих зубов сравнительно высокие, удлиненные, без эктостилида. Лингвальные стенки сравнительно плоские, с хорошо развитым парастилидом, более слабым энтостилидом, слабо выступающим метастилидом и слабо вздутыми столбиками метаконида и энтоконида. Между метастилидом и столбиком энтоконида лингвальная стенка коронки заментно углублена, имеет вид выемки с гранями. На поверхности окклюзии передние половины коронок немного короче и шире задних, лингвальные полулуния очень слабо угловатые.
Нижние молочные зубы dP3 и dP4 сильно стерты. Коронка dP3 узкая спереди, сильно расширена кзади. Парастилид и параконид хорошо развиты. Метаконид скошен назад, отделен от параконида открытой, широкой, V-образной долинкой и очень слабо выступает лингвально. На поверхности окклюзии задний край коронки вогнут. На dP4 передняя доля зуба немного уже двух задних и чуть скошена по отношению к ним. Две задние доли зуба примерно равны по ширине и длине. На буккальной стороне коронки присутствуют два базальных столбика.
Пястная и плюсневая кости умеренно массивные (рис. 3). Медиальные части их диафизов значительно шире и толще латеральных частей. Ширина дистальных эпифизов превышает ширину проксимальных эпифизов. На пястной кости экз. ПИН, № 5644/304 проксимальный эпифиз немного поврежден сзади у латерального края. Проксимальная суставная поверхность округленно-треугольного очертания. Ее более крупная медиальная фасетка для os trapezoideocapitatum с сильновыпуклым передним краем. Сзади между этой фасеткой и латеральной фасеткой для os hamatum (unciforme), лежит серповидный узкий желоб (шириной около 4 мм), с небольшим отверстием в средней части. На передней (дорсальной) поверхности пястной кости у проксимального конца хорошо развита шероховатость для лучевого разгибателя запястья, m. extensor carpi radialis, а на задней поверхности присутствуют медиальный и латеральный бугорки для сухожилий мышц-флексоров (e.g., m. flexor carpi radialis et m. flexor carpi ulnaris). Диафиз постепенно расширяется к дистальному эпифизу примерно с середины длины кости. Передняя поверхность диафиза округленная. Отверстие проксимального канала пястной кости, canalis metacarpi proximalis, крошечное. Дорсальный продольный желоб, sulcus longitudinalis dorsalis, на месте слияния третьей и четвертой пястных костей (mc III и mc IV) очень слабый. Он немного углубляется и расширяется (до 3–3.5 мм) около очень маленького отверстия дистального канала пястной кости, canalis metacarpi distalis, расположенного на значительной высоте от суставных блоков для сочленения с первыми фалангами III и IV пальцев. Задняя поверхность диафиза уплощенная, немного вогнутая в проксимальной части, где с боков хорошо выражены шероховатости для связок II и V пястных костей.
Переднезадний диаметр проксимального эпифиза плюсневой кости экз. ПИН, № 5644/305 превышает его ширину. Проксимальная суставная поверхность с передним краем, выступающим округленным углом вперед; медиальная и латеральная стороны суставной поверхности почти прямые, слабо расходящиеся, а задний край S-образно изогнут. Между передними (дорсальными) фасетками, медиальной для os cuneiforme II + III и латеральной для os naviculocuboideum, лежит маленькое сосудистое отверстие, а позади него более крупное отверстие, которое ведет в canalis metatarsi proximalis, открывающийся на плантарной поверхности у проксимального конца. Задняя (плантарная) латеральная фасетка для os naviculocuboideum длинная, немного смещена латерально и слабо наклонена. На передней поверхности кости sulcus longitudinalis dorsalis расширяется на уровне небольшого canalis metatarsi distalis, расположенного на значительной высоте от суставных блоков. Задняя поверхность диафиза в нижней части уплощена. На середине длины кости, переднезадний диаметр диафиза больше его ширины.
Размеры в мм, индексы в %. Экз. ПИН, № 5644/304, пястная кость, длина (L), поперечный диаметр (DT), переднезадний диаметр (DAP), проксимальный (prox), дистальный (dist), диафиз (diaph) в середине: L – 220.5; DTprox × DAPprox – 56.5 × 38.2; DTdist × DAPdist – 62 × 36; DTdiaph × DAPdiaph – 34.83 × 27.27; индекс уплощенности поксимального эпифиза (DAPprox/ DTprox – 68; индекс массивности (DTdiaph/L) – 16; индекс ширины дистального эпифиза (DTdist/L) – 28.
Экз. ПИН, № 5644/305, плюсневая кость: L – 245.5; DTprox × DAPprox – 45.75 × 46; DTdist × × DAPdist – 55.75 × 31.8; DTdiaph × DAPdiaph – 31.25 × 32.3; индекс массивности (DTdiaph/L) – 13; индекс ширины дистального эпифиза (DTdist/L) – 23.
Сравнение и замечания. По морфологии и размерам образцы из Тавриды соответствуют M. latifrons Schaub, 1923 из нижнего плейстоцена Сенез, Франция (2.2–1.5 млн л. н., ca. 2 млн л. н.; Delson et al., 2006), реперного местонахождения позневиллафранкской зоны MNQ18 (рис. 1–3, табл. 1). Особенности строения сохранившихся частей лобной и верхнечелюстной костей, а также положение основания рогового стержня идентичны таковым голотипа M. latifrons – Se 556, описанного Шаубом (1923).
По морфологии и размерам щечных зубов экз. ПИН, № 5644/302 также хорошо соответствует голотипу, и лишь длина верхнего ряда премоляров немного превосходит таковую голотипа (длина P2–P4 = 52 мм; длина P2–P4 = 51 мм; Schaub, 1923). Некоторые отличия Р3, Р4 и М1 экз. ПИН, № 5644/302 от аналогичных зубов голотипа (напр., присутствие crista medialis на Р3 и Р4) обусловлены разной степенью стирания щечных зубов у форм из Тавриды и Сенез (у последних они более стерты). Строение и размеры нижних моляров M1 и М2 экз. ПИН, № 5644/303 сходны с таковыми нижнечелюстной кости паратипа Se 401.
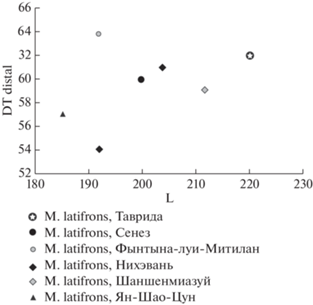
Пястная кость зкз. ПИН, № 5644/303 из Тавриды по морфологии, а также ширине проксимального и дистального эпифизов, соответствует паратипу Se 1731 M. latifrons из Сенез и из близкого ему по возрасту местонахождения Фынтына луи Митилан, Румыния (DTprox: 56.5 и 56 мм; DTdist: 60 и 64 мм соответственно), но она немного длиннее и стройнее (рис. 4). Индекс DTdist/L пястной кости M. latifrons из Сенез – 30% (по Schaub, 1937), а из Фынтына луи Митилан – 33.3% (Radulesco, Samson, 1962, рис. 1, табл. 1).
Рис. 4.
Соотношение параметров пястных костей у овибовин рода Megalovis (с использованием данных: Teilhard de Chardin, Piveteau, 1930; Schaub, 1937; Radulesco, Samson, 1962; Tong et al., 2022).
Oт M. balcanicus Crégut-Bonnoure et Dimitrijević, 2006 из Трлицы, Черногория (MNQ18–20, 1.8–0.7 млн л. н.), Megalovis из Тавриды отличается более плоским лбом, более резким наклоном латеральной вогнутости между основанием рогового стержня и орбитой, меньшим расстоянием переднего края основания рога от орбиты, а также менее косым положением контакта между пеньком и роговым стержнем.
Голотип M. balcanicus – неполный череп TRL 88/66/1 с роговыми стержнями, без носовых костей и базальной части – хранится в Университете Белграда (Crégut-Bonnoure, Dimitrijević, 2006, табл. I , фиг. 1a–d). В. Димитриевич сообщила мне, что он найден в слое 2 (Layer II) (конец раннего плейстоцена, MmQ3; Crégut-Bonnoure, Dimitrijević, 2006, табл. III , фиг. 1a-c; MNQ20, >0.7 млн л. н.; Fernandez, Crégut, 2007), который соответствует нашему слою 5 (начало среднего плейстоцена, MmQ3, MIS19) (Agadzhanyan et al., 2017; Vislobokova et al., 2020). Паратипы найдены и в нижележащих слоях. Возраст нижнего фаунистического горизонта TRL11–10 – 1.8–1.5 млн лет (Vislobokova et al., 2020). Диагноз M. balcanicus включает в себя особенности лобных костей между глазницами и роговыми стержнями, а также особенности положения основания рогового стержня, которые отличают его от голотипа M. latifrons. Эта часть черепа у обоих видов пневматизирована. Полное строение роговых стержней у M. latifrons остается неизвестным. Их более поздняя реставрация у голотипа (Schaub, 1932, 1943, рис. 5, 6 ), возможно, основанная на образце Se 1983 Pliotragus, не может использоваться для сравнительных анализов до новых находок, подтверждающих такое строение роговых стержней у этого вида (см. Spassov, Crégut-Bonnoure, 1999; Gentry, 2001; Crégut-Bonnoure, 2002).
У голотипа M. latifrons Se 556 лоб плоский, и верхние края орбит лежат в одной плоскости; основания роговых стержней, округлые в сечении, сильно наклонены в стороны и несколько назад (Schaub, 1923). А у голотипа M. balcanicus TRL 88/66/1 лоб выпуклый; основания роговых стержней, вытянутые в переднезаднем направлении и уплощенные, направлены в стороны и почти сразу отгибаются вперед. Они сильнее сдвинуты назад от заднего края орбит и больше скошены относительно сагиттальной плоскости черепа (передний край рога расположен гораздо латеральнее, чем его задняя сторона) (Crégut-Bonnoure, Dimitrijević, 2006). По этим признакам образец из Тавриды соответствует M. latifrons и отличается от M. balcanicus.
Megalovis из Тавриды отличается от M. balcanicus также по ряду признаков строения зубов, таких как: 1) присутствие вертикальной бороздки на лингвальной стенке Р3 и Р4; 2) отсутствие на M1 базального столбика (есть на M1/ TRL 88/23/5); 3) отсутствие на нижних молярах буккального цингулума между лингвальными полулуниями; 4) большая длина метастилида на нижних молярах (на зубах из Трлицы метастилид развит лишь в верхней половине коронки). Все эти признаки, по данным Э. Крегут-Боннур и В. Димитриевич, отличают M. latifrons от M. balcanicus (см. Crégut-Bonnoure, Dimitrijević, 2006).
Эти виды отличаются и по строению Р4, которое отражает их разные эволюционные уровни. В процессе эволюции происходило сокращение длины и ширины второй доли зуба. У M. latifrons Se 401 вторая доля сравнительно длинная и шире передней доли. У M. balcanicus вторая доля уже передней доли, и она короче, чем у M. latifrons (см. Crégut-Bonnoure, Dimitrijević, 2006, табл. II , фиг. 14, 15 ).
Следует отметить, что до находки в Тавриде строение нижних молочных зубов у M. latifrons было неизвестно. У европейских Megalovis единственный dP4 TRL 90/79/4 был описан из Трлицы (Crégut-Bonnoure, Dimitrijević, 2006, табл. II , фиг. 10b, c ). Строение dP4 экз. ПИН, № 5644/303 в целом сходно с таковым у M. balcanicus, но этот зуб стерт сильнее и его стили слабее. Особенности строения dP3 на этом образце в целом согласуются с эволюционным уровнем P3 и P4 Se 401. По строению P3 М. latifrons немного примитивнее, чем у M. balcanicus. Метаконид на P3 у Se 401 и у TRL 01/22/7 скошен назад, но у первого, в отличие от второго, переднее крыло метаконида не сливается с параконидом. Нижний молочный dP3 у M. guangxiensis из нижнего плейстоцена пещеры Куекуе (1.2–0.8 млн л. н.), Гуанси, имеет более прогрессивное строение, чем у формы из Тавриды; его коронка короче и спереди шире (см. Dong et al., 2020, рис. 2B, F).
Другие находки мегаловисов в Европе менее представительны. В Болгарии M. aff. latifrons идентифицирован в фауне Варшец по М3 и двум М2 (Spassov, 2005), M. aff. balcanicus – в фауне пещеры Козарника по единственному неполному M3 (Fernandez, Crégut, 2007) и Megalovis sp. – по неполным нижним челюстям и костям конечностей из Сливницы (Spassov, Crégut-Bonnoure, 1999; Megalovis aff. latifrons: Spassov, 2005; Crégut-Bonnoure, 2007). По данным исследователей, Megalovis из Сливницы отличается от M. latifrons некоторыми признаками строения нижних моляров (в частности, более слабым лингвальным рельефом), но отражают ли эти отличия географическую изменчивость вида или в Сливнице представлен другой вид, пока не ясно (Spassov, Crégut-Bonnoure, 1999; Spassov, 2005; Crégut-Bonnoure, 2007). Кроме того, Megalovis присутствует в фауне Пирро Норд в Италии, где представлен кончиком рогового стержня, двумя верхними молярами (M2 и M3) и астрагалом (Ovibovini: De Giuli et al., 1986, табл. I , фиг. 3, 4; Megalovis sp.: Crégut-Bonnoure, 2002; Crégut-Bonnoure, Dimitrijević, 2006; Masini, Sala, 2007).
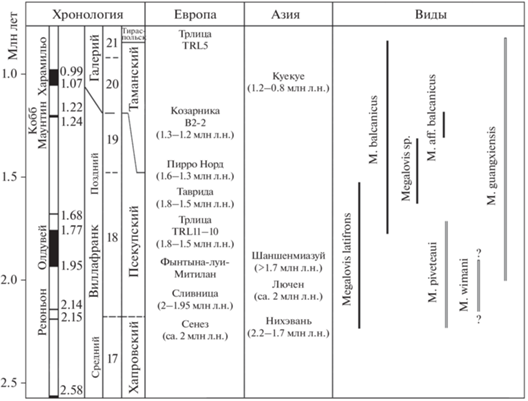
В конце раннего плейстоцена европейские и китайские виды Megalovis были значительно разобщены географически и палеозоогеографически и, скорее всего, представляли разные ветви эволюции рода. Многие особенности строения этих видов отражают общность происхождения и параллелизм в эволюции европейских и китайских ветвей. Хронологическое положение местонахождений и распространение видов рода Megalovis показаны на рис. 5.
Рис. 5.
Хронологическое положение местонахождений и распространение видов рода Megalovis (c использованием данных: Crégut-Bonnoure, 2005, 2007; Delson et al., 2006; Fernandez, Crégut, 2007; Spassov, 2016; Sardella et al., 2018; Dong et al., 2020; Tong et al., 2021, 2022).
Виды M. piveteaui Schaub, 1937 и M. wimani Schaub, 1937 из Северного Китая лишь немного отличаются от M. latifrons размерами зубов, а также длиной и массивностью пястных костей (Schaub, 1937).
Вид M. piveteaui выделен Шаубом по верхнечелюстной и пястной костям из местонахождения Санкан-хо классической фауны Нихэвань (2.2–1.7 млн л. н.; Tong et al., 2021). Они были описаны П. Тейяр де Шарденом и Ж. Пивто как “Oviboviné gen. indet.” и хранятся в Национальном музее естественной истории в Париже (Teilhard de Chardin, Piveteau, 1930, табл. XIII , фиг. 2, 4). К этой же форме был отнесен фрагмент основания правого рогового стержня, уплощенного (DAP = 82 mm, DT = 55 mm), направленного в сторону и загнутого вперед (Teilhard de Chardin, Piveteau, 1930, рис. 27 ), который также, возможно, принадлежал M. piveteaui. Лучевая и пястная кости этого вида недавно найдены в местонахождении Шаншенмиазуй бассейна Нихэвань (Tong et al., 2021, 2022).
Вид M. wimani представлен лишь двумя образцами в коллекции Университета Уппсалы: верхнечелюстной костью с P2–M3 (голотип) и пястной костью из местонахождения Ян-Шао-Цун (Loc. A), Хэнань (Schaub, 1937, рис. 1, 2). Вид M. guangxiensis Han, 1987 выделен по неполным верхне- и нижнечелюстным костям и отдельным зубам из раннего плейстоцена Лючена (~2 млн л. н.), Гуанси; его голотип – нижнечелюстная кость с P2–M3 V 58829.1 хранится в Ин-те палеонтологии позвоночных и палеоантропологии АН Китая (Han, 1987, табл. IX , фиг. 4). По присутствию хорошо развитых базальных столбиков на молярах этот вид отличается почти от всех представителей рода, в том числе и от Megalovis из Тавриды. Базальные столбики отмечены лишь на M3 M. aff. balcanicus из пещеры Козарника (Fernandez, Crégut, 2007).
Пястная кость из Тавриды длиннее, чем у китайских видов. Индекс ее массивности (DTdist/L) близок к таковым у M. piveteaui (28–30%, n = 2) из классической фауны Нихэвань и у M. piveteaui (28%) из фауны Шаншенмиазуй бассейна Нихэвань (Teilhard de Chardin, Piveteau, 1930; Radulesco, Samson, 1962; Tong et al., 2022), и меньше, чем у M. wimani (ca. 31%) из Ян-Шао-Цун (Loc. A), Хэнань (см. Schaub, 1937, рис. 2).
В фауне Тавриды Megalovis latifrons сосуществовал с другим представителем трибы Ovibovini – Soergelia minor Moya-Sola, 1987, который встречался в интевале 1.8–1.2 млн л. н. (Вислобокова, 2022). Эти виды имеют определенное сходство, на что обращали внимание и другие исследователи, и предполагали, что первый мог быть предком второгo (напр., Moyá-Solá, 1987; Crégut-Bonnoure, 2002). Новые данные о морфологии и распространении видов M. latifrons и S. minor позволяют считать, что это сходство говорит о близком родстве и, по-видимому, общности происхождения Megalovis и Soergelia. M. latifrons был крупнее и отличается от S. minor рядом морфологических признаков (как примитивных, так прогрессивных): 1) основания рогов у M. latifrons довольно сильно сдвинуты назад по сравнению с голотипом S. minor VM 85 SI M24-39 и экз. VM 84. P-5-23 из типового местонахождения Вента Мицена (MNQ20, 1.3 млн л. н., MmQ2; Moyá-Solá, 1987); 2) коронки верхних предкоренных P3 и P4 отклоняются назад (y S. minor не отклоняются); 3) задние половины коронок P3 и P4 сильнее удлинены, столбик паракона лежит ближе к переднему краю коронок; у S. minor столбик паракона на равном расстоянии от переднего и заднего краев коронки; 4) коронки P3 и P4 более уплощены лингвально, у S. minor их контур здесь более угловатый; 5) на буккальной стенке M1 основания столбика паракона и парастиля слиты и образуют узкую V‑образную “выемку”; у S. minor эта “выемка” U-образная (у паратипа, Moyá-Solá, 1987, табл. 3C ); 6) нижнечелюстная кость в области диастемы у паратипа Se 401 выше, и длина диастемы меньше длины M1–M3; у S. minor oна низкая и длинная, длина диастемы немного меньше длины P2–M3 (см. Вислобокова, 2022, рис. 3); 7) четвертая (задняя) долинка на P3 и P4 открывается на заднем краю коронки, у S. minor – у ее задневнутреннего угла; 8) столбик метастилида на M2 и M3 мощнее и длиннее (простирается до основания коронки), и между ним и энтоконидом стенка коронки сильнее углублена, с резкими гранями; у S. minor столбик метастилида короткий, углубление слабое и сглаженное, стенка коронки в целом более уплощена; 9) парастилид развит сильнее; 10) пястная кость длиннее (признаки 4, 8, 9 и10 по Moyá-Solá, 1987 и нашим данным).
В процессе эволюции у овибовин метаподии становились массивнее. Индексы ширины дистального эпифиза пястной кости и массивности диафиза у M. latifrons близки к таковым у S. minor из Вента Мицены, 27–30% (m = 28.54) и 15–17% (m = 15.75, n = 8) (вычислены по: Moyá-Solá, 1987). У S. elisabethae Schaub, 1951 из среднего плейстоцена Зюссенборна эти индексы равны 29 и 19% (по: Schaub, 1951). Сравнительно небольшие величины этих индексов у M. latifrons из Tавриды могут говорить о довольно низком эволюционном уровне этой популяции. По размерам пястные и плюсневые кости у M. latifrons (в том числе из Тавриды) крупнее, чем у S. minor и S. elisabethae. Так, длина пястной кости у этих видов Soergelia, соответственно, равна 160.2–177.0 мм (m = 168.46, n = 8) и 180 мм (по данным Schaub, 1951; Moyá-Solá, 1987).
Общие размеры у европейских Megalovis, как и у Soergelia, возможно, увеличивались от более ранних раннеплейстоценовых популяций к более поздним. Связано ли это увеличение с индивидуальной изменчивостью, эволюцией автохтонных форм или, как у Soergelia, с разными дисперсионными волнами, покажут новые находки и дальнейшие исследования.
Материал. Фрагмент лобной кости с частью основания рогового стержня, экз. ПИН, № 5644/301; фрагмент левой стороны лицевого отдела черепа с РЗ–М1 и альвеолами Р2, экз. ПИН, № 5644/302; часть левой нижнечелюстной кости с dP3–М2, экз. ПИН, № 5644/303; правая пястная кость, экз. ПИН, № 5644/304; правая плюсневая кость, экз. ПИН, № 5644/305; Крым, пещера Таврида; нижний плейстоцен.
ЗАКЛЮЧЕНИЕ
Проведенное исследование позволило установить присутствие еще одного вида Ovibovini, Megalovis latifrons Schaub, 1923, в раннеплейстоценовой фауне пещеры Таврида (1.8–1.5 млн л. н.), и получить дополнительные данные о морфологии этого вида (в частности, строении нижних молочных зубов и плюсневой кости). Крымские M. latifrons, по-видимому, были немного крупнее более ранних форм из Франции и Румынии. Находка M. latifrons в Крыму уточняет некоторые аспекты эволюции этого вида, а также его географическое и стратиграфическое распространение.
Исследование выполнено за счет гранта Российского научного фонда, № 22-14-00214, https://rscf.ru/project/22-14-00214/.
Список литературы
Вислобокова И.А. О первой находке Soergelia minor (Artiodactyla, Bovidae) в нижнем плейстоцене пещеры Таврида в Крыму и истории рода Soergelia // Палеонтол. журн. 2022. № 3. С. 61–70.
Лопатин А.В., Вислобокова И.А., Лавров А.В. и др. Пещера Таврида – новое местонахождение раннеплейстоценовых позвоночных в Крыму // Докл. Акад. наук. 2019. Т. 485. № 3. С. 381–385.
Agadzhanyan A.K., Vislobokova I.A., Shunkov M.V., Ulyanov V.A. Pleistocene mammal fauna of the Trlica locality, Montenegro // Fossil Imprint. 2017. V. 73. № 1–2. P. 93–114.
Crégut-Bonnoure E. Les Ovibovini et Caprini (Mammalia, Artiodactyla, Bovidae, Caprinae) du Plio-Pléistocène dʼEurope: systématique, évolution et biochronologie. Thèse Doct. dʼEtat ès Sciences, Université de Lyon I, 2002. 386 p.
Crégut-Bonnoure E. Nouvelles données paléogéographiques et chronologiques sur les Caprinae (Mammalia, Bovidae) du Pléistocène moyen et supérieur d’Europe // Munibe Antropol. Arkeol. 2005. V. 57. P. 205–219.
Crégut-Bonnoure E. Apport des Caprinae et Antilopinae (Mammalia, Bovidae) à la biostratigraphie du Pliocène terminal et du Pléistocène d’Europe // Quaternaire. 2007. V. 18. № 1. P. 73–97.
Crégut-Bonnoure E., Dimitrijević V. Megalovis balcanicus sp. nov. and Soergelia intermedia sp. nov. (Mammalia, Bovidae, Caprinae), new Ovibovini from the Early Pleistocene of Europe // Rev. Paléobiol. 2006. V. 25. № 2. P. 723–773.
De Giuli C., Masini F., Torre D. The latest Villafranchian faunas in Italy: the Pirro Nord Fauna (Apricena, Gargano) // Palaeontogr. Ital. 1986. V. 74. P. 56–62.
De Giuli C., Masini F., Torre D. The latest Villafranchian faunas in Italy: the Pirro Nord local fauna (Apricena, Gargano) // Quartärpaläontologie. 1990. Bd 8. P. 29–34.
Delson E., Faure M., Gueren C. Franco-American renewed research at the Late Villafranchain locality of Senèze (Haute Loire), France // Cour. Forsch.-Inst. Senckenb. 2006. Bd 256. P. 275–290.
Dong W., Bai W.P. Artiodactyla assemblages associated with Gigantopithecus blacki in China // Acta Anthropol. Sin. 2021. V. 40. № 3. P. 490–502.
Dong W., Wang Y., Bai W.P. et al. Late Early Pleistocene Artiodactyls associated with Gigantopithecus from Queque Cave, Chongzuo, Guangxi, South China // Acta Anthropol. Sin. 2020. V. 39. P. 71–83.
Fernandez P., Crégut E. Les Caprinae (Rupicaprini, Ovibovini, Ovini et Caprini) de la séquence Pléistocène de Kozarnika (Bulgarie du Nord): morphométrie, biochronologie et implications phylogéniques // Rev. Paléobiol. 2007. V. 26. № 2. P. 425–503.
Gentry A.W. An ovibovine (Mammalia, Bovidae) from the Neogene of Stratzing, Austria // Ann. Naturhist. Mus. Wien. 2001. Bd 102A. P. 189–199.
Han D. Artiodactyla fossils from the Giant Ape Cave in Liucheng, Guangxi // Mem. Inst. Vertebr. Palaeontol. Palaeoanthropol. Acad. Sin. 1987. № 18. P. 135–208.
Lehmann U. Weitere Fossilfunde aus dem ältesten Pleiztozän der Erpfinger Höhle (Schwäbische Alb) // Mitt. Geol. Staatinst. Hamburg. 1957. Bd 26. S. 60–99.
Masini F., Sala B. Large- and small-mammal distribution patterns and chronostratigraphic boundaries from the Late Pliocene to the Middle Pleistocene of the Italian peninsula // Quartern. Intern. 2007. V. 160. P. 43–56.
Moyá-Solá S. Los bovidos (Artiodactyla, Mammalia) del yacimiento del Pleistoceno inferior de Venta Micena (Orce, Granada, España) // Paleontol. Evol. 1987. № 1. P. 181–236.
Radulesco C., Samson P. Sur la présence de Megalovis latifrons dans le Villafranchien d’Olténie (Roumanie) // Vertebr. PalAsiat. 1962. V. 6. № 3. P. 262–269.
Radulesco C., Samson P. Biochronology and evolution of the Early Pliocene to the Early Pleistocene mammalian faunas of Roumania // Boll. Soc. Paleontol. Ital. 2001. V. 40. P. 285–291.
Sardella R., Bellucci L., Bona F. et al. Before and after the earliest Homo dispersal in Europe: Evidence from the Early Pleistocene sites of the Italian Peninsula // C. R. Palevol. 2018. V. 17. № 4–5. P. 287–295.
Schaub S. Neue und wenig bekannte Cavicornier von Senèze // Ecl. Geol. Helv. 1923. Bd 18. S. 281–295.
Schaub S. Die Ruminantier des ungarischen Praeglacialis // Ecl. Geol. Helv. 1932. Bd 25. S. 319–335.
Schaub S. Ein neuer Cavicornier aus dem Oberpliocaen von Honan // Bull. Geol. Inst. Univ. Uppsala. 1937. Bd 27. P. 25–31.
Schaub S. Die oberpliocaene Säugetierfauna von Senèze (Haute-Loire) und ihre verbreitungsgeschichtliche Stellung // Ecl. Geol. Helv. 1943. Bd 36. S. 270–289.
Schaub S. Soergelia n. gen., ein Caprine aus dem thuringischen Altpleistocaen // Ecl. Geol. Helv. 1951. Bd 44. S. 375–381.
Spassov N. The mammalian megafauna from the Late Villafranchian localities Varshets and Slivnitsa (Bulgaria) and the biochronology of the Villafranchian in S.-E. Europe. Ph. D. Thesis. Sofia: NMNH, 1999.
Spassov N. Brief review of the Pliocene ungulate fauna of Bulgaria // Quaternaire. 2005. V. 2. P. 201–212.
Spassov N. Southeastern Europe as a route for the earliest dispersal of Homo toward Europe: ecological conditions and the timing of the first human occupation of Europe // Paleoanthropology of the Balkans and Anatolia: Human Evolution and its Context / Eds. Harvati K., Roksandic M. Dordrecht: Springer, 2016. P. 281–290.
Spassov N., Crégut-Bonnoure E. Premières données sur les Bovidae Villafranchiens de Bulgarie // C. R. Acad. Sci. Paris. 1999. V. 328. P. 493–498.
Teilhard de Chardin P., Piveteau J. Les mammiféres fossils de Nihowan (Chine) // Ann. Paléontol. 1930. V. 19. P. 1–134.
Tong H.-W., Zhang B., Chen X. et al. Chronological significance of the mammalian fauna from the Early Pleistocene Shanshenmiaozui site in Nihewan Basin, northern China // Acta Anthropol. Sin. 2021. V. 40. № 3. P. 469–489.
Tong H.-W., Zhang B., Chen X. et al. New fossils of small and medium-sized bovids from the Early Pleistocene site of Shanshenmiaozui in Nihewan Basin, North China // Vertebr. PalAsiat. 2022. V. 60. № 2. P. 134–168.
Vislobokova I.A., Agadzhanyan A.K., Lopatin A.V. The case of Trlica TRL11–10 (Montenegro): Implications for possible early hominin dispersals into the Balkans in the middle of the Early Pleistocene // Quatern. Intern. 2020. V. 554. P. 15–35.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал