

Палеонтологический журнал, 2023, № 2, стр. 80-87
Новый вид ставридовых рыб рода Archaeus (Carangidae, Percomorpha) из нижнего олигоцена Северного Кавказа
А. Ф. Банников a, *, И. Г. Еребакан a, b, **
a Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
b Московский государственный университет им. М.В. Ломоносова
119991 Москва, Россия
* E-mail: aban@paleo.ru
** E-mail: erebakan.ivan@mail.ru
Поступила в редакцию 03.11.2022
После доработки 18.11.2022
Принята к публикации 18.11.2022
- EDN: LBPZWJ
- DOI: 10.31857/S0031031X23020046
Аннотация
Из нижнего олигоцена (пшехская свита) Северного Кавказа (местонахождение Горный Луч) по отпечатку целого скелета описан новый вид ставридовых рыб (Carangidae) Archaeus solus sp. nov., первая находка Carangidae в олигоцене Северного Кавказа. Это третий известный вид эоцен–олигоценового рода Archaeus Agassiz, 1844. Новый вид отличается от A. glarisianus Agassiz, 1844 из рюпеля Швейцарии и A. oblongus Daniltshenko, 1968 из базального эоцена Туркменистана большим числом лучей в спинном плавнике – как колючих, так и мягких.
ВВЕДЕНИЕ
Ископаемые ставридовые рыбы (семейство Carangidae) известны с территории Тетиса, начиная с базального эоцена [глобальное аноксическое событие, случившееся в ответ на парниковые условия, связанные с палеоцен-эоценовым термальным максимумом (PETM) (Gavrilov et al., 2003; Банников, 2017)]. Из туркменского местонахождения близ с. Уйля-Кушлюк в предгорьях Копетдага, образованного во время PETM, известен комплекс морских рыб (Данильченко, 1968), включающий в себя три вида ставридовых – Archaeus oblongus Daniltshenko, 1968, Seriola paleocenica (Sytchevskaya et Prokofiev, 2005) и Trachicaranx tersus Daniltshenko, 1968. Последний таксон А.Ф. Банников (1990, 2010) относит к Apolectidae, однако самостоятельность этого семейства в последнее время отрицается (Smith-Vaniz, 1984; Springer, Smith-Vaniz, 2008; Santini, Carnevale, 2015; Jose et al., 2022 и др.). Также с РЕТМ связано местонахождение в Дании, откуда известны не описанные пока ставридовые (Bonde, 1987, рис. на с. 42; Heingård et al., 2021, рис. 4 ). Уже в позднем ипре (Монте Болька, Италия) разнообразие Carangidae составляет шесть родов (Bannikov, 2014) (или даже семь, если относить Trachicaranx к ставридовым). Впоследствии Carangidae являются непременным компонентом большинства морских фаун рыб Тетиса и Паратетиса, хотя их обилие и разнообразие там обычно невелико (см. Банников, 1990, 2010). Так, в раннем олигоцене Восточного Паратетиса отмечено лишь два вида ставридовых – Selar quassus Bannikov, 1990 и Caranx daniltshenkoi Bannikov, 1990, причем первый из них известен по единственной находке, а второй – по четырем экземплярам. Все эти находки сделаны в Абхазии, а с территории Северного Кавказа раннеолигоценовых Carangidae до недавнего времени не было известно, несмотря на продолжительные сборы ископаемых рыб на нескольких олигоценовых северокавказских местонахождениях.
В 2014 г. в ходе полевых работ Палеонтологического ин-та им. А.А. Борисяка РАН (ПИН РАН) на местонахождении раннеолигоценовых рыб на северо-западе Кавказа (Краснодарский край, р. Пшеха) был обнаружен отпечаток скелета с противоотпечатком небольшой ставридовой рыбы. Ниже он описан как принадлежащий новому виду Archaeus solus sp. nov.
Семейство Carangidae традиционно помещалось в подотряд Percoidei отряда Perciformes (Берг, 1940; Johnson, 1984; Банников, 1990, 2010; Nelson, 2006 и др.). В последнее время, однако, на смену традиционной классификации организмов зачастую приходит формальная кладистическая систематика, построенная на компьютерной обработке молекулярных данных и часто противоречащая морфологическим свидетельствам. В кладистической систематике ставридовым и нескольким другим семействам придается ранг отряда Carangiformes11 (Wiley, Johnson, 2010; Near et al., 2012, 2013; Betancur-R. et al., 2013, 2017; Nelson et al., 2016 и др.) в “отделе (division) Percomorphacea” (Wiley, Johnson, 2010), “серии Carangaria” (Betancur-R. et al., 2017) или “подсерии Carangaria” (Grande, Wilson, 2021). Высокие таксоны кладистической иерархии морфологически гетерогенны, что признают и сами кладисты (Miya et al., 2013); это побуждает нас использовать традиционную, естественную систематику, как это сделали недавно Ф. Химстра и др. (Coastal Fishes…, 2022).
В семействе Carangidae принято выделять четыре современных подсемейства [Seriolinae (=Naucratinae), Trachinotinae, Scomberoidinae и Caranginae: Банников, 1986, 1990, 2010], либо четыре трибы (Naucratini, Trachinotini, Scomberoidini и Carangini: Smith-Vaniz, 1984; Santini, Carnevale, 2015; Reed et al., 2002), а также эоценовые подсемейство Vomeropsinae (Bannikov, 1984; Банников, 1986) и трибу Paratrachinotini (Springer, Smith-Vaniz, 2008). Монофилия трех современных подсемейств поддержана одними молекулярными данными (Reed et al., 2002; Jose et al., 2022), они группируются как (Trachinotinae + (Caranginae + Seriolinae)) (Jose et al., 2022); монофилия Scomberoidinae остается под вопросом, но возможно, эта клада является сестринской Trachinotinae (Reed et al., 2002). По другим данным, Scomberoidinae является монофилетичной кладой, а Trachinotinae, по-видимому, разделены на две отдельные линии (Lichia и Trachinotus) (Santini, Carnevale, 2015).
МАТЕРИАЛ И МЕТОДЫ
Материалом для работы послужил отпечаток скелета рыбы с противоотпечатком из колл. ПИН РАН стандартной длиной тела (SL) 44 мм из слоистых слабоизвестковистых глин пшехской свиты (см. Попов и др., 2019) нижнего олигоцена на правом берегу р. Пшеха (Краснодарский край) против хут. Горный Луч.
Образец отпрепарирован штыковым зондом и изучен с помощью бинокулярного микроскопа Leica M165C в ПИН РАН. В качестве сравнительного материала использовались образцы ранее описанных ископаемых видов ставридовых рыб из колл. ПИН РАН, а также остеологические коллекции и рентгеноснимки скелетов современных ставридовых. Фотографии выполнены С.В. Багировым (ПИН РАН).
Исследование проводилось за счет гранта Российского научного фонда № 22-27-00162, https://rscf.ru/project/22-27-00162/.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
СЕМЕЙСТВО CARANGIDAE RAFINESQUE, 1815
Род Archaeus Agassiz, 1844
Archaeus solus Bannikov et Erebakan, sp. nov.
Название вида solus лат. – одинокий, единственный.
Голотип – ПИН, № 5419/15, отпечаток полного скелета с противоотпечатком; Краснодарский край, Апшеронский р-н, р. Пшеха напротив хут. Горный Луч; нижний олигоцен, пшехская свита, планорбелловые слои.
Описание (рис. 1–3). Тело относительно удлиненное, веретеновидное, по-видимому, несколько сжатое с боков, с довольно коротким и умеренно тонким хвостовым стеблем; его наибольшая высота находится у начала первого спинного плавника и лишь немного уступает длине головы. Голова относительно высокая, ее длина до 3.2 раза укладывается в SL. Высота хвостового стебля в 3.1 раза меньше высоты тела. Глаза довольно крупные, диаметр орбиты лишь немного уступает длине рыла и составляет 29% длины головы. Рот довольно крупный, конечный, по-видимому, не выдвижной; нижняя челюсть выступает вперед.
Рис. 1.
Archaeus solus sp. nov., голотип ПИН, № 5419/15, полный скелет: а – общий вид, б – противоотпечаток; Краснодарский край, Апшеронский р-н, р. Пшеха напротив хут. Горный Луч; нижний олигоцен, пшехская свита, планорбелловые слои. Масштабная линейка – 0.5 см.

Рис. 2.
Archaeus solus sp. nov., голотип ПИН, № 5419/15, хвостовой скелет: а – общий вид, отражено зеркально; б – прорисовка (крайние главные лучи хвостового плавника заштрихованы); Краснодарский край, Апшеронский р-н, р. Пшеха напротив хут. Горный Луч; нижний олигоцен, пшехская свита, планорбелловые слои. Обозначения – ep – epurale, h – hypurale, hpu3 – гемальный отросток третьего преурального позвонка, ph – parhypurale, pu2 – второй преуральный позвонок, st – stegurale. Масштабная линейка – 2 мм.
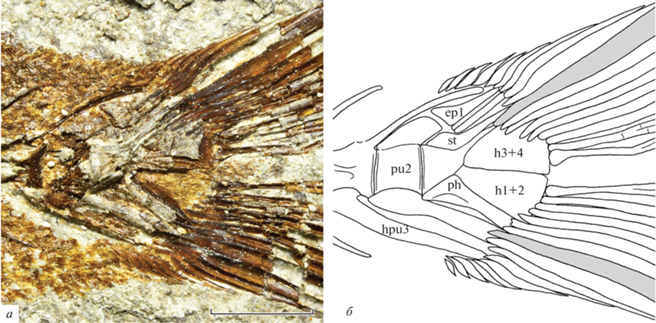
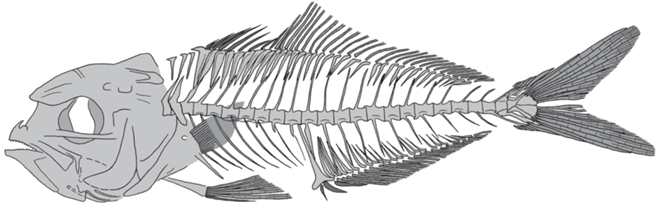
Рис. 3.
Archaeus solus sp. nov., реконструкция скелета по голотипу ПИН, № 5419/15, чешуя не показана.
Нейрокраний умеренно высокий, почти прямой парасфеноид проецируется в нижней части орбиты. Лобно-затылочный гребень относительно невысокий. Этмоидный район черепа довольно короткий; передняя часть frontalia нависает над ним. В орбите имеются окостенения склеротики. Сохранность не позволяет различить подглазничные кости. Нижнечелюстное сочленение расположено примерно под серединой орбиты. Рraemaxillare сильно разрушено; его восходящий отросток тонкий, умеренной длины, явно не слит с сочленовным. Передний край восходящего отростка составляет с альвеолярной ветвью praemaxillare угол, близкий к прямому. Зубы в верхней челюсти небольшие, конические. Maxillare сохранилось фрагментарно, supramaxillare не различимо. Нижняя челюсть невысокая, ее длина составляет около 51% длины головы и соответствует длине шести последних туловищных позвонков. Симфиз низкий, без подбородочного выступа, отогнут вниз. Неподалеку от симфиза на внутренней части dentale имеется отверстие сосуда. Dentale V-образное, в вырезку на его заднем крае входит аngulo-articulare. Озубление нижней челюсти сходно с таковым верхней челюсти. Сочленовный отросток аngulo-articulare довольно короткий, но крепкий; retroarticulare не различимо. Ось hyomandibulare очень слабо наклонена вперед от вертикали. Метаптеригоид соединяет hyomandibulare с quadratum и ограничивает орбиту постеро-вентрально. Quadratum довольно широкое, округло-треугольное, с утолщенным постеровентральным краем и умеренным сочленовным мыщелком. Еctopterygoideum образует передний стержень и расположенный под углом к нему постеровентральный отросток для сочленения с передним краем quadratum. Еctopterygoideum несет мелкие ямки на медиальной поверхности. Относительно крупное плоское entopterygoideum образует дно орбиты. Жаберная крышка умеренно широкая. Praeoperculum относительно крупное, плоское, довольно слабо вогнуто вдоль утолщенного переднего края. Свободный край предкрышки ровный. Operculum довольно крупное, плоское, широкое; его нижний край приострен, а передний утолщен. Граница между suboperculum и interoperculum не различима. Кости гиоидного комплекса сохранились плохо. Точное число саблевидных, тонких, заостренных radii branchiostegi неизвестно. На противоотпечатке голотипа, обращенном головой налево (рис. 1, а), имеются минерализованные жаберные лепестки, и различимы несколько конических глоточных зубов.
Позвонков 24, из них 10 туловищные и 14 каудальные. Длина туловищной части позвоночника в 1.7 раза короче хвостовой его части. Линия позвоночника очень слабо вогнута вниз у границы туловищной и хвостовой частей. Тела позвонков от почти квадратных в латеральном аспекте до слабо удлиненных, слегка пережатые посередине. Позвонки несут с каждой стороны по продольному гребню, разделяющему две латеральные впадины. Большинство остистых отростков относительно тонкие и длинные, почти прямые или слегка изогнутые. Четыре передних невральных отростка и передний гемальный слегка расширены медиально. Передние гемальные отростки отходят от передней части позвонков. Три задних туловищных позвонка несут небольшие парапофизы, удлиняющиеся каудально. Ребра довольно длинные, тонкие, наклонены назад (сначала умеренно, затем слабо); передние из них прикрепляются к телам позвонков (начиная со второго), а задние – к дистальной части парапофизов. Ребра последней пары особенно тонкие и короткие. Epineuralia плохо различимы под туловищными позвонками.
Хвостовой скелет частично скрыт гипуростегией (раздвоенными основаниями лучей хвостового плавника): не различимы uroneuralia второй пары и пятое hypurale (рис. 2). Второй преуральный позвонок слабо укорочен, его невральный отросток редуцирован до невысокого гребня, а гемальный отросток сильный и автогенный. Остистые отростки третьего преурального позвонка длиннее, чем у предшествующего позвонка. Epuralia три, переднее расширено проксимально. Uroneuralia первой пары образуют stegurale. Hypuralia слиты в две пластинки без гипуральной диастемы. Parhypurale без видимого parhypurapophysis. Хвостовой плавник относительно крупный, сильно выемчатый, состоит из 17 главных лучей (I8-7I). Верхние и нижние дополнительные лучи относительно многочисленные, различимо около девяти лучей сверху и снизу. “Procurrent spur” (Johnson, 1975, 1984) отсутствует.
Posttemporale – плоская эллиптическая кость умеренного размера с двумя сильными передними отростками, связывающими грудной пояс с нейрокранием. От удлиненного supracleithrum различима только нижняя часть. Cleithrum крупное, почти прямое, загнуто верхним концом вперед. Границы coracoideum, scapula и пекторальных radialia различимы плохо. Postcleithrum заостренное, узкое, ребровидное, направлено назад и вниз. Грудные плавники небольшие, их основания расположены под третьим–четвертым позвонком над серединой расстояния от линии позвоночника до вентрального края тела. Точное число лучей в грудном плавнике неизвестно, но их не менее 15.
Тазовые кости довольно длинные, узкие, клиновидные. Брюшные плавники крупные, длиннее грудных, прикрепляются непосредственно позади основания грудных; в брюшном плавнике тонкая колючка и пять мягких ветвящихся лучей.
Между затылком и первым спинным плавником расположено три supraneuralia, предорсальная формула (Ahlstrom et al., 1976; Johnson, 1984) – 0/0/0+2+1/1/. Supraneuralia небольшие, каждое с направленным вперед заостренным выступом в верхней части. Спинные плавники соприкасаются; первый из них короткий в основании, начинается на вертикали четвертого позвонка; в нем восемь крепких колючек, удлиняющихся до четвертой (которая в 2.8 раза длиннее первой), затем постепенно укорачивающихся назад. Высота плавника короче длины его основания. Две первых колючки сверхштатные. Первый птеригиофор довольно мощный, пластинчато расширен медиально и несет продольный гребень. В верхней его части имеется мощный заостренный направленный вперед отросток, нависающий над третьим supraneurale. Последующие птеригиофоры первого спинного плавника постепенно сужаются.
Второй спинной плавник начинается на вертикали восьмого–девятого позвонка, в нем один жесткий и 28 мягких членистых лучей. Шип второго спинного плавника ближе прилегает к первому мягкому лучу, чем к последнему шипу первого спинного плавника (которого он немного длиннее). Мягкие лучи расположены довольно тесно. Второй луч самый длинный, каудально лучи постепенно укорачиваются. Высота второго спинного плавника превышает высоту первого спинного плавника. Птеригиофоры второго спинного плавника очень узкие; в каудальном направлении птеригиофоры постепенно укорачиваются и становятся более наклоненными. Межневральные промежутки под вторым спинным плавником вмещают проксимальные концы одного–трех птеригиофоров каждый. Паттерн вставки супраневралий и дорсальных птеригиофоров (supraneural and pterygiophore insertion pattern, по: Springer, Smith-Vaniz, 2008): 0/0/0-2/ 1/2/2/3/1/2/2/2/2/2/1/2/2/2/3/3/1/-/-/-/c. Возможно, этот паттерн слегка искажен посмертной деформацией тела рыбы.
Анальный плавник сходен со вторым спинным по размерам, но сдвинут назад относительно начала второго спинного (начинается под первым–вторым хвостовым позвонком). Длина основания анального плавника в 1.5 раза превышает расстояние между брюшным и анальным плавниками. В анальном плавнике три крепкие, но недлинные удлиняющиеся назад колючки (третья из них ближе прилегает к первому мягкому лучу, чем ко второму шипу) и 23 мягких, членистых лучей. Передний мягкий луч самый длинный, каудально лучи укорачиваются. Первый анальный птеригиофор очень мощный, умеренно наклонен назад; проксимальным концом доходит до позвоночника, но не образует крепкого соединения с гемальным отростком первого хвостового позвонка. Дистально птеригиофор Т-образно расширен и несет все три анальных шипа. Птеригиофоры мягких лучей анального плавника очень узкие, несколько длиннее противолежащих птеригиофоров второго спинного плавника и также укорачиваются спереди назад. Проксимальные концы двух–трех птеригиофоров входят в межгемальные промежутки над анальным плавником. Паттерн вставки анальных птеригиофоров (по: Springer, Smith-Vaniz, 2008): 1/3/3/3/2/3/3/3/3/-/-/-/-/c.
Чешуя очень мелкая, тонкая, циклоидная, образует сплошной покров на теле. Боковая линия не прослеживается, щитков на ней явно нет.
Размеры. SL голотипа – 44 мм.
В процентах к SL голотипа: длина тела с хвостовым плавником – 122, длина головы – 31, наибольшая высота тела – 28, высота хвостового стебля – 9, первое предорсальное расстояние – 41, второе предорсальное расстояние – 52, преанальное расстояние – 59, вентроанальное расстояние – 21, длина наибольших шипов спинного плавника – 9.5, длина наибольших мягких лучей спинного плавника – 11, длина наибольшего (третьего) шипа анального плавника – 6, длина наибольших мягких лучей анального плавника – 10, длина основания жесткой части спинного плавника – 12.5, длина основания мягкой части спинного плавника – 39, длина основания целого спинного плавника – 52, длина основания анального плавника – 32, длина брюшного плавника – 17, длина наибольших лучей хвостового плавника – 26, длина рыла – 9.5, горизонтальный диаметр орбиты – 9, длина нижней челюсти – 18.
Сравнение. Новый вид отличается от A. glarisianus Agassiz, 1844 из рюпеля Швейцарии (Woodward, 1901) и A. oblongus Daniltshenko, 1968 из базального эоцена Туркменистана (Данильченко, 1968; Банников, 1990, 2010) большим числом лучей во втором спинном плавнике (28 против не более 25 лучей у этих видов) и дорсальных шипов (9 против 8 у A. glarisianus и 7 у A. oblongus). Также, у нового вида относительно более низкий первый спинной плавник (у A. glarisianus он явно выше второго спинного, а у A. oblongus равен ему).
Материал. Голотип.
ОБСУЖДЕНИЕ
На принадлежность нового таксона к семейству Carangidae указывают форма и пропорции тела, меристические признаки (в частности, наличие 24 позвонков и протяженный, многолучевой анальный плавник), единый лобно-затылочный гребень, мелкая циклоидная чешуя, отсутствие “procurrent spur”, а также наличие характерного для ставридовых промежутка между второй и третьей колючкой анального плавника. Новый таксон лишен апоморфий подсемейств Scomberoidinae (овальная, ланцетовидная или игловидная чешуя, увеличенное число хвостовых позвонков, часто редуцированное число шипов в спинном плавнике), Trachinotinae (укороченная вентральная ветвь posttemporale, редукция второй сверхштатной колючки в начале жесткого спинного плавника), Vomeropsinae (высокое тело, редукция числа колючек в спинном плавнике, округлый сзади хвостовой плавник) и Caranginae (наличие щитков на боковой линии, тонкий хвостовой стебель). Первый анальный птеригиофор нового таксона, хотя и мощный, неплотно связан с первым гемальным отростком. Этот признак характерен для рыб подсемейства Naucratinae, но у них первый анальный птеригиофор существенно слабее, а анальный плавник короче (Банников, 1986, 1990), не превышает вентроанальное расстояние.
Такие признаки нового вида как удлиненное тело с относительно высоким хвостовым стеблем, наличие 24 позвонков, неплотно связанный с первым гемальным отростком первый анальный птеригиофор, отсутствие щитков на боковой линии, относительно протяженный анальный плавник – не противоречат отнесению его к ископаемому роду Archaeus Agassiz, 1844. Типовой вид этого рода, A. glarisianus Agassiz, 1844, описан из рюпеля Швейцарии (кантон Глярус) (Agassiz, 1833-1844) по неполному, частично расчлененному экземпляру. Позже еще несколько видов родов Archaeus и Archaeoides описано из рюпеля Гляруса (Rath, 1859; Wettstein, 1886) по преимущественно фрагментарным и сильно искаженным растяжением породы в различных направлениях образцам; все они сведены в синонимию к A. glarisianus А. Вудвардом (Woodward, 1901), который привел краткий диагноз этого вида. П.Г. Данильченко (1960, 1980) описал ставридовую рыбу из нижнего олигоцена Абхазии как A. glarisianus, не обнаружив щитков на боковой линии рыб. Последующее изучение абхазского материала выявило наличие таких щитков, что послужило основанием для выделения нового вида подсемейства Caranginae Caranx daniltshenkoi (Банников, 1990).
Банников (1986) включил род Archaeus в подсемейство Seriolinae (=Naucratinae) условно, поскольку не все его признаки укладываются в диагноз Seriolinae. Вскоре (Банников, 1990) этот род был выделен в особое подсемейство Archaeinae. Однако по причине омонимии это название группы семейства является невалидным – еще в XIX в. для пауков-убийц рода Archaea Koch et Berendt, 1854 выделено семейство Archaeidae (Koch, Berendt, 1854). Таким образом, род Archaeus Agassiz, 1844 должен считаться Carangidae inc. subfam. Помимо типового вида, в составе рода Archaeus описан А. oblongus из базального эоцена Туркменистана (Данильченко, 1968; Банников, 1990, 2010). Соответственно, находка A. solus sp. nov. расширяет состав эоцен–олигоценового рода Archaeus до трех видов.
Список литературы
Банников А.Ф. К систематическому положению, составу и происхождению семейства ставридовых (Carangidae) // Вопр. ихтиол. 1986. Т. 25. Вып. 6. С. 883–889.
Банников А.Ф. Ископаемые ставридовые и ворон-рыбы СССР // Тр. Палеонтол. ин-та АН СССР. 1990. Т. 244. 108 с.
Банников А.Ф. Ископаемые позвоночные России и сопредельных стран. Ископаемые колючеперые рыбы (Teleostei, Acanthopterygii). М.: ГЕОС, 2010. LXI + 244 с.
Банников А.Ф. Развитие и смена фаун костистых рыб в эоцене Тетиса // Геология и биоразнообразие Тетиса и Восточного Паратетиса. Матер. Всеросс. научно-практич. конф. 23-26 июля 2017 г., Горячий Ключ / Ред. С.В.Попов. Горячий Ключ, 2017. С. 7–10.
Берг Л.С. Система рыбообразных и рыб, ныне живущих и ископаемых // Ежегодн. Зоол. музея АН СССР. 1940. Т. 5. С. 87–517.
Данильченко П.Г. Костистые рыбы майкопских отложений Кавказа // Тр. Палеонтол. ин-та АН СССР. 1960. Т. 78. 208 с.
Данильченко П.Г. Рыбы верхнего палеоцена Туркмении // Очерки по филогении и систематике ископаемых рыб и бесчелюстных / Ред. Д.В.Обручев. М.: Наука, 1968. С. 113–156.
Данильченко П.Г. Отряд Perciformes // Ископаемые костистые рыбы СССР / Ред. Новицкая Л.И. М.: Наука, 1980. С. 115–121, 125–169 (Тр. Палеонтол. ин-та АН СССР. Т. 178).
Попов С.В., Табачникова И.П., Банников А.Ф. и др. Лектостратотип майкопской серии по р. Белая выше г. Майкопа (Западное Предкавказье) в его олигоценовой части // Стратигр. Геол. корреляция. 2019. Т. 27. № 3. С. 70–92.
Третьяков Д.К. Очерки по филогении рыб. Киев: Изд‑во АН УССР, 1944. 176 с.
Agassiz L. Recherches sur les Poissons Fossiles. Neuchâtel: Petitpierre, 1833–1844. T. 5. Pt 1. 122 p.
Ahlstrom E.H., Butler J.L., Sumida B.Y. Pelagic stromateoid fishes (Pisces, Perciformes) of the eastern Pacific: kinds, distributions and early life histories and observations on five of these from the northwest Atlantic // Bull. Mar. Sci. 1976. V. 26. P. 285–402.
Bannikov A.F. The new subfamily of carangid fishes // Studi ric. giacim. terz. Bolca. Verona, 1984. V. 4. P. 319–321.
Bannikov A.F. The systematic composition of the Eocene actinopterygian fish fauna from Monte Bolca, northern Italy, as known to date // Studi ric. giacim. terz. Bolca. Verona, 2014. V. 15. P. 23–33.
Betancur-R. R., Broughton R.E., Wiley E.O. et al. The tree of life and a new classification of bony fishes // PLOS Currents Tree of Life. 2013. P. 1–41.
Betancur-R. R., Wiley E.O., Arratia G. et al. Phylogenetic classification of bony fishes // BMC Evol. Biol. 2017. 17:162. P. 1–40. https://doi.org/10.1186/s12862-017-0958-3
Bonde N. Moler—its origin and its fossils especially fishes. Nykøbing Mors: SKAMOL, 1987. 53 p.
Coastal Fishes of the Western Indian Ocean / Eds. Heemstra P.C., Heemstra E., Ebert D.A. et al. Makhanda, South Africa: S. Afr. Inst. for Aquatic Biodiversity, 2022. 5 vols.
Gavrilov Yu.O., Shcherbinina E.A., Oberhänsli H. Paleocene-Eocene boundary events in the northeastern Peri-Tethys // Causes and Consequences of Globally Warm Climates in the Early Paleogene / Eds. Wing S.L., Gingerich P.D., Schmitz B., Thomas E. Boulder, Colorado, 2003. P. 147–168 (Geol. Soc. Amer. Spec. Pap. № 369).
Grande T., Wilson M.W.H. Chapter 1. Evolution and phylogeny // The Physiology of Fishes, 5th ed. / Eds. Currie S., Evans D.H. Boca Raton: CRC Press, 2021. P. 1–17.
Heingård M., Sjövall P., Sylvestersen R.L. et al. Crypsis in the pelagic realm: evidence from exceptionally preserved fossil fish larvae from the Eocene Stolleklint Clay of Denmark // Palaeontology. 2021. V. 64. Pt 6. P. 805–815. https://doi.org/10.1111/pala.12574
Johnson G.D. The procurrent spur, an undescribed perciform character and its phylogenetic implications // Occas. Pap. Calif. Acad. Sci. 1975. V. 121. P. 1–23.
Johnson G.D. Percoidei: development and relationships // Ontogeny and Systematics of Fishes / Eds. Moser H.G., Richards W.J., Cohen D.M. Lawrence: Allen Press, 1984. P. 464–498 (Amer. Soc. Ichthyol. Herpetol. Spec. Publ. № 1).
Jose A., Sukumaran S., Mukundan L.P. et al. Comparative mitogenomics and phylogenetics of the family Carangidae with special emphasis on the mitogenome of the Indian Scad Decapterus russelli // Sci. Reports. 2022. 12:5642. https://doi.org/10.1038/s41598-022-09636-5
Koch C.L., Berendt G.C. Die im Bernstein befindlichen Crustaceen, Myriapoden, Arachniden und Apteren der Vorwelt // Die im Bernstein befindlichen organischen Reste der Vorwelt, Erster Band, II. Abtheilung / Ed. Berendt G.C. Berlin, 1854. S. 1–124.
Miya M., Friedman M., Satoh T.P. et al. Evolutionary origin of the Scombridae (tunas and mackerels): Members of a Paleogene adaptive radiation with 14 other pelagic fish families // PLOS One. 2013. V. 8. № 9. P. e73535:1–19.
Near T.J., Dornburg A., Eytan R.I. et al. Phylogeny and tempo of diversification in the superradiation of spiny-rayed fishes // Proc. Nat. Acad. Sci. USA. 2013. V. 110. P. 12738–12743.
Near T.J., Eytan R.I., Dornburg A. et al. Resolution of ray-finned fish phylogeny and timing of diversification // Proc. Nat. Acad. Sci. USA. 2012. V. 109. P. 13698–13703.
Nelson J.S. Fishes of the World. 4th ed. Hoboken, NJ: Wiley, 2006. 601 p.
Nelson J.S., Grande T.C., Wilson M.V.H. Fishes of the World. 5th ed. Hoboken, NJ: Wiley, 2016. 707 p.
Rath J.J.G. vom. Beitrag zur Kenntniss der fossilen Fische des Plattenberges im Canton Glarus // Z. Dtsch. Geol. Ges. 1859. Bd 11. H. 1. S. 108–132.
Reed D.L., Carpenter K.E., deGravelle M.J. Molecular systematics of the Jacks (Perciformes: Carangidae) based on mitochondrial cytochrome b sequences using parsimony, likelihood, and Bayesian approaches // Mol. Phylogen. Evol. 2002. V. 23. P. 513–524.
Santini F., Carnevale G. First multilocus and densely sampled timetree of trevallies, pompanos and allies (Carangoidei, Percomorpha) suggests a Cretaceous origin and Eocene radiation of a major clade of piscivores // Mol. Phylogen. Evol. 2015. V. 83. P. 33–39. https://doi.org/10.1016/j.ympev.2014.10.018
Smith-Vaniz W.F. Carangidae: relationships // Ontogeny and Systematics of Fishes / Eds. Moser H.G., Richards W.J., Cohen D.M. Lawrence: Allen Press, 1984. P. 522–530 (Amer. Soc. Ichthyol. Herpetol. Spec. Publ. № 1).
Springer V.G., Smith-Vaniz W.F. Supraneural and pterygiophore insertion patterns in carangid fishes, with description of a new Eocene carangid tribe, †Paratrachinotini, and a survey of anterior anal-fin pterygiophore insertion patterns in Acanthomorpha // Bull. Biol. Soc. Wash. 2008. № 16. P. 1–73.
Wettstein A. Über die Fische des tertiären Glarnerschiefers // Abh. Schweiz. Paläontol. Ges. 1886. Bd 13. S. 5–103.
Wiley E.O., Johnson G.D. A teleost classification based on monophyletic groups // Origin and Phylogenetic Interrelationships of Teleosts / Eds. Nelson J.S., Schultze H.-P., Wilson M.V.H. Munich: Verl. Dr. F. Pfeil, 2010. P. 123–182.
Woodward A.S. Catalogue of the fossil fishes in the British Museum (Natural History). Vol. IV. L.: Brit. Mus. (Natur. Hist.), 1901. 638 p.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал