

Палеонтологический журнал, 2023, № 2, стр. 88-99
О находке диадектоморфных тетрапод в нижней перми Восточной Европы
В. В. Буланов *
Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
* E-mail: bulanov@paleo.ru
Поступила в редакцию 19.09.2022
После доработки 07.11.2022
Принята к публикации 07.11.2022
- EDN: LDEMJJ
- DOI: 10.31857/S0031031X2302006X
Аннотация
Stephanospondylus (Bolosaurus) traati (Tatarinov, 1974), ранее рассматривавшийся в составе Bolosauridae, перенесен в семейство Diadectidae отряда Diadectomorpha. Данный таксон является первым диадектоморфом, описанным с территории Европейской России. Форма и характер дифференциации челюстных зубов свидетельствуют о большей примитивности S. traati по сравнению с типовым видом рода Stephanospondylus (S. pugnax) и его базальном положении внутри Diadectidae. Род Ambedus Kissel et Reisz, 2004 исключен из состава Diadectomorpha по причине несоответствия организации зубной системы морфотипу, характеризующему данный отряд. Сходство в строении зубной системы Ambedus с Diadectidae трактуется как конвергентное; положение этого рода в филогении тетрапод остается неопределенным. Обсуждается вопрос ранних этапов специализации зубной системы диадектид.
ВВЕДЕНИЕ
Bolosauridae (Bolosaurida) – одна из немногих и наиболее специализированных групп тетрапод, освоивших нишу мелкоразмерного фитофага в экосистемах ранней–средней перми Лавразии. Представители семейства известны из Северной Америки, Европы и Восточной Азии (Cope, 1878; Ивахненко, 1973; Li et al., 1995; Berman et al., 2000). Несмотря на обширный ареал, находки болозаврид редки, причем почти все известные виды описаны по фрагментарному челюстному материалу. Как следствие, основой таксономической идентификации болозаврид выступают детали строения зубной системы, глубокая специализация которой сильно выделяет группу на фоне остального разнообразия палеозойских амниот (Ивахненко, Твердохлебова, 1987; Reisz et al., 2002, 2007; Snyder et al., 2020; Bulanov et al., 2022).
Значительное число отнесенных к Bolosauridae форм происходит из перми Восточной Европы (Татаринов, 1968, 1974; Ивахненко, 1973, 1990; Ивахненко, Твердохлебова, 1987; Ивахненко и др., 1997; Bulanov et al., 2022), и также исходно установлено по фрагментарным челюстным остаткам. В настоящее время очевидно, что вследствие неполноты материала и дефицита данных по морфологии болозаврид в целом, в состав семейства был включен ряд проблематичных таксонов, систематическое положение которых в настоящее время требует пересмотра. Одной из таких форм является Bolosaurus traati, описанный Л.П. Татариновым (1974) по неполной верхнечелюстной кости из ассельских карбонатно-морских отложений Республики Коми (местонахождение Мылва).
До этой находки род Bolosaurus, объединяющий наиболее архаичных представителей Bolosauridae, был известен исключительно из нижнепермских отложений США (Cope, 1878; Case, 1907; Watson, 1954), и его определение с территории Восточно-Европейской платформы являлось одним из немногочисленных свидетельств общности тетраподных ассоциаций ранней перми Лавразии (Ивахненко, 1990). Позднее, однако, принадлежность B. traati к болозавридам была поставлена под сомнение (Reisz et al., 2002), поскольку голотип не несет признаков, характеризующих данное семейство; при этом вопрос систематической принадлежности образца не обсуждался. В 2008 г. М.Ф. Ивахненко указал на сходство голотипа B. traati (экз. ПИН, № 3318/1) с челюстными остатками Stephanospondylus pugnax Stappenbeck, 1905 – проблематичного диадектида, описанного по разрозненным элементам скелета из нижнего ротлегенда Германии (Stappenbeck, 1905; Romer, 1925; Kissel, 2010), и рассматривал восточно-европейскую форму как вид того же рода (S. traati) в составе Bolosauridae на основании деталей строения maxillare и челюстных зубов (Ивахненко, 2008).
Полученные в последнее время данные по организации зубной системы ранне- и среднепермских болозаврид (Snyder et al., 2020; Bulanov et al., 2022), однако, свидетельствуют против принадлежности голотипа B. traati к этой группе, что требует пересмотра таксономической принадлежности как этого образца, так и рода Stephanospondylus. Кроме того, челюсть, судя по положению площадок стирания на сохранившихся зубах, принадлежала одной из примитивных растительноядных форм, что заслуживает отдельного рассмотрения, принимая во внимание ограниченный набор раннепермских групп тетрапод, адаптированных к облигатной фитофагии.
МОРФОЛОГИЧЕСКОЕ ОПИСАНИЕ ГОЛОТИПА STEPHANOSPONDYLUS TRAATI (TATARINOV, 1974)
Голотип B. traati (экз. ПИН, № 3318/1; рис. 1, 2) представляет собой неполное левое maxillare. Передний край кости утерян. Длина сохранившегося фрагмента составляет 29 мм. Линия зубного ряда изогнута ближе к его окончанию в соответствии с выраженным латеральным изгибом максиллы. Maxillare широкое на всем протяжении, поскольку его дорсальный край по мере смещения в каудальном направлении отклоняется латерально, нависая ближе к концу челюсти над окончанием зубного ряда (рис. 1, б–г). Дорсальный фланг несет поверхность причленения, предположительно, скуловой кости. Контактная площадка имеет форму пологой депрессии с отверстиями для входа ветвей инфраорбитальной артерии; ее передний край маркируется резко очерченным гребнем, расположенным напротив второй из представленных на образце альвеол (рис. 1, г). Лицевая поверхность maxillare слабо скульптирована и несет несколько отверстий, образующих ряд вдоль вентрального края челюсти. Лабиальный парапет не выражен.
Рис. 1.
Stephanospondylus traati (Tatarinov, 1974), голотип ПИН, № 3318/1, левое maxillare: а – латерально, б – антеро-латерально, в – вентрально, г – дорсально, д – угол наклона максиллярных зубов, вид спереди; Республика Коми, местонахождение Мылва; нижняя пермь, ассельский ярус. Обозначения: pd – плицидентин, rt – не сформированная до конца коронка сменного зуба. Длина масштабной линейки для фиг. а–г – 0.5 см, для фиг. д – 1 мм.
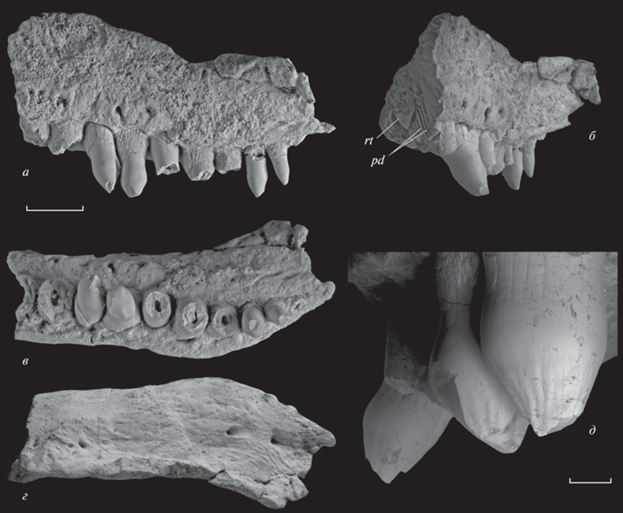
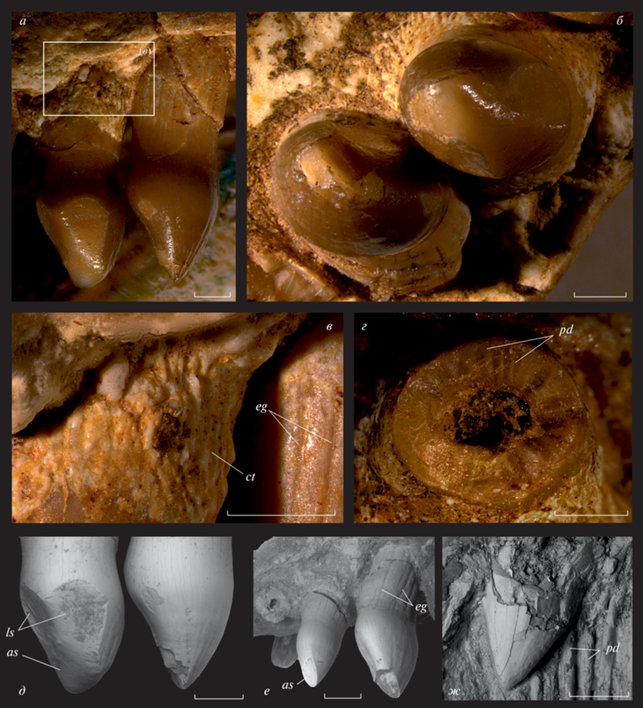
Рис. 2.
Stephanospondylus traati (Tatarinov, 1974), голотип ПИН, № 3318/1, детали строения maxillare: а – сохранившиеся передние буккальные зубы, лингвально; б – то же, вентрально; в – увеличенный участок прикорневой части зуба в третьей альвеоле, лингвально; г – плицидентиновая структура стенки сломанного зуба в седьмой альвеоле (в 1.5 мм от альвеолярного края); д – фасетки прижизненного стирания на зубе в четвертой альвеоле (слева) и микротрассы, предшествующие их формированию на зубе в третьей альвеоле (справа), лингвально; е – сохранившиеся задние буккальные зубы, лингвально; ж – коронка сменного зуба в первой альвеоле, вид спереди; Республика Коми, местонахождение Мылва; нижняя пермь, ассельский ярус. Обозначения: as – апикальная фасетка стирания; ct – неваскуляризованная костная ткань в прикорневой части зуба; eg – борозды, разделяющие складки дентина в прикорневой части зуба; ls – лингвальные фасетки стирания; pd – плицидентин. Длина масштабных линеек – 1 мм.
Образец содержит часть зубного ряда из 10 последних альвеол, которые расположены очень плотно на всем протяжении челюсти и занимают всю ширину зубной площадки. Судя по их размеру, зубы уменьшались спереди назад равномерно. Только четыре зуба в челюсти сохранили в различной степени поврежденные коронки, которые позволяют заключить, что зубы были дифференцированы не только по размеру, но и по форме. Кроме того, при виде спереди передние и задние сохранившиеся зубы направлены вниз под существенно разным углом, величина которого между первым и последним сохранившимися зубами составляет 35° (рис. 1, а, б, д).
Коронки зубов (рис. 1, 2) имеют неправильно-коническую, близкую к каплевидной форму. Их базальная часть заметно утолщена, что более выражено у зубов передней части зубного ряда. Выпуклая сторона коронок обращена вперед и слегка медиально; она орнаментирована сходящимися к вершине пологими гребнями, в совокупности образующими слабоволнистую поверхность (рис. 1, д; 2, е). Противоположная сторона коронок, напротив, слегка уплощена (что характерно, в том числе и для задних зубов maxillare: рис. 2, а, б, д–ж) и гладкая. У передних зубов вершина немного смещена к их лабиальной стороне.
К сожалению, вершинки всех представленных на образце зубов повреждены, но узость апикального отдела позволяет достаточно уверенно реконструировать уплощенно-коническую форму коронок и говорить об отсутствии здесь дополнительных конусов или плечевидных уступов. На это же указывает закладка сменного зуба в самой передней из представленных на образце альвеол, вскрытой в результате поперечного слома кости. Сменный зуб сформирован не полностью и представляет собой только привершинную часть коронки, которая имеет форму уплощенного конического колпачка с ровными краями и слабо выраженной режущей кромкой (рис. 1, б). Прижизненная ориентация сменного зуба в альвеоле не нарушена: он занимает базальное положение и при виде с торца челюсти экспонируется своей передней (расширенной) стороной, т.е. расположен в соответствии с ориентацией коронок сохранившихся близлежащих рабочих зубов (рис. 2, ж).
Форма поперечных сечений оснований зубов на уровне альвеолярного края меняется от слабо овальной в переднем отделе максиллы до все более округлой в направлении конца зубного ряда. Передние зубы вытянуты поперек челюсти не строго перпендикулярно, а расположены с едва заметным смещением лабиальной стороны вперед относительно лингвальной. Зубы в заднем отделе челюсти развернуты слабее, но, в связи с изгибом максиллы и зубного ряда, их коронки сохраняют ту же ориентацию, что и таковые передних максиллярных зубов. Предпоследний зуб на уровне альвеолярного края имеет округлую форму сечения, однако поперечный слом последнего, не сохранившегося, зуба имеет форму узкого овала, развернутого длинной осью вдоль внешнего края кости, что резко отличает его от остальных зубов maxillare (рис. 1, в).
Зубы имеют текодонтное крепление: их основания погружены в кость до дна альвеолы и прочно срощены с ее боковыми стенками, что хорошо видно на сломе первой из представленных на образце альвеол. Стенки зубов образуют хорошо выраженные складки (рис. 1, б; 2, ж). Вследствие их плотного смыкания в прикоронарной части стенка зубов сильно утолщена, сломы зубов на этом уровне демонстрируют петлевидную структуру дентина (рис. 2, г). Ближе к альвеолярному краю складки становятся обособленными и в таком виде протягиваются до конца корневой части зуба, составляющей примерно треть его полной высоты (рис. 1, б).
На внешней стороне борозды, разделяющие складки, хорошо различимы и доходят до базальной части коронки. У некоторых зубов челюсти они отчасти закрыты снаружи слоем визуально неваскуляризованной костной ткани, которая отличается значительно более осветленным тоном как от дентина зубов, так и несущей их кости (рис. 2, а, в). Ее поверхность осложнена резко выраженными гребнями и бороздами неправильной формы, ориентированными преимущественно вертикально. Поскольку материал не допускает возможности гистологического анализа данного образования, его природа остается дискуссионной. При этом следует отметить, что наличие осветленной “рубашки” на основаниях зубов хорошо коррелирует с их износом. В частности, она отсутствует на не стертых зубах, занимающих четвертую и восьмую из представленных на голотипе альвеол (а также на основании обломанного зуба в пятой альвеоле), и хорошо развита на стертых зубах в позициях три и девять. Из этого наблюдения следует, что данная ткань формировалась внутри десны уже после анкилозной фиксации зуба в рабочем положении. Мозаичное распределение в челюсти зубов, имеющих или лишенных этого образования, а также следов прижизненного износа указывает, что их смена у рассматриваемой формы не носила последовательный характер, что резко отличает ее, в частности, от всех Bolosauridae (Snyder et al., 2020). Отверстия резорбции на сохранившемся фрагменте челюсти отсутствуют, однако сменные зубы формировались внутриальвеолярно, что доказывается наличием сменной коронки in situ внутри поврежденной первой альвеолы.
Сохранившиеся рабочие зубы челюсти имеют хорошо выраженные площадки прижизненного износа. Наименьший из зубов – предпоследний в челюсти – несет только апикальную, резко наклоненную фасетку стирания на постеро-лингвальной стороне (рис. 2, е). Зуб, занимающий четвертую из представленных на образце альвеол, имеет аналогичную площадку, ориентированную более горизонтально, однако дистально к ней примыкают еще две большие фасетки стирания, оставленные, несомненно, двумя разными зубами-антагонистами (рис. 2, а, б, д). Они располагаются в существенно разных плоскостях с антеро- и каудо-лингвальной сторон зуба; вследствие их смыкания на лингвальной стороне коронки образуется гребень, отсутствующий на менее стертых зубах. Привершинная и антеро-лингвальная площадки стирания несут многочисленные царапины, ориентированные продольно по отношению к зубному ряду (рис. 2, д). У сменившегося позднее впередилежащего зуба имеется лишь небольшая фасетка стирания с лингвальной стороны базальной части коронки, но четкие горизонтальные микротрассы уже прослеживаются и в ее апикальной части (рис. 2, д), что в совокупности демонстрирует состояние челюстных зубов у рассматриваемой формы на начальной стадии их износа. Наличие по-разному ориентированных площадок стирания на зубах, а также горизонтальных микротрасс, указывает на способность нижней челюсти осуществлять достаточно сложные, в том числе прополинарные, движения в процессе питания.
ОБСУЖДЕНИЕ
Таксономическая идентификация голотипа Stephanospondylus (Bolosaurus) traati (Tatarinov, 1974). Строение зубов у голотипа ПИН, № 3318/1, их ориентация в челюсти, характер дифференцировки и стирания, их “разбалансированная” смена и складчатая структура оснований, а также форма самого maxillare, свидетельствующая, в частности, о выраженном расширении черепа в постдентальной части, однозначно не позволяют относить этот образец к Bolosauridae, и роду Bolosaurus в частности, как это предлагалось ранее (Татаринов, 1974; Ивахненко и др., 1997; Ивахненко, 2001, 2008). Тем не менее, голотип ПИН, № 3318/1 несет значительное число морфологических маркеров для уверенного определения его таксономической принадлежности.
Единственные специализированные растительноядные тетраподы, сочетающие текодонтный тип имплантации зубов с их анкилозной фиксацией и плицидентиновой структурой корневой части, описаны среди представителей отряда Diadectomorpha, внутри которого сложная форма зубов характеризует семейство Diadectidae (Cope, 1896, 1911; Vaughn, 1969; Berman, Sumida, 1995; Berman et al., 1998, 2004; Kissel, 2010; LeBlanс, Reisz, 2013). Все остальные черты голотипа ПИН, № 3318/1, такие как форма самого maxillare, ориентировка зубов в челюсти и их дифференцировка по длине зубного ряда, порядок замещения зубов и характер их прижизненного стирания, также аналогичны или находят много общего с таковыми описанного разнообразия диадектид, в первую очередь – с примитивными формами семейства.
Так, для всех видов Diadectidae, известных по полным черепам (Orobates pabsti, Diadectes absites, D. sideropelicus, D. zenos, D. lentus, D. carinates, D. sanmiguelensis), свойственно резкое расширение черепа в орбитальной области, что отражается в латеральном изгибе maxillare и его зубного ряда (Case, Williston, 1912, 1913; Lewis, Vaughn, 1965; Berman et al., 1998, 2004; Kissel, 2010). Типичным при этом является “нависание” дорсального края кости над задними зубами челюсти, а также латеральное отклонение передних зубов максиллы по сравнению с таковыми задней части зубного ряда, что отмечено выше и для голотипа ПИН, № 3318/1. Указанные признаки характеризуют всех представителей семейства, за исключением Ambedus pusillus Kissel et Reisz, 2004, – сильно дистанцированной от остальных диадектид формы, которая описана по фрагментарному челюстному материалу из нижней перми США и исключена из дальнейшего сравнения по причинам, обсуждаемым ниже.
Наблюдаемое на голотипе ПИН, № 3318/1 сильное прижизненное стирание зубов является характерной особенностью всех диадектид, зубная система которых функционировала по принципу контактной окклюзии (“tooth-on-tooth occlusion”) (Reisz, Sues, 2000; Reisz, 2006). Судя по опубликованным данным, характер износа зубов аналогичен таковому диадектида Desmatodon hesperis (Vaughn, 1969). Мозаичное распределение на образце стертых и не стертых зубов, а также коррелирующее с этим наличие/отсутствие неваскуляризованной костной ткани, покрывающей стенку зубов на участке между корнем и коронкой, указывают на “разбалансированное” обновление зубного ряда, что свойственно диадектидам (Vaughn, 1972; LeBlanс, Reisz, 2013), но не характерно для болозаврид, замещение зубов у которых носило последовательный характер (Snyder et al., 2020; Bulanov et al., 2022). Типичным для Diadectidae является и внутриальвеолярное формирование сменных зубов (LeBlanс, Reisz, 2013), что устанавливается для голотипа ПИН, № 3318/1 благодаря инситному положению коронки сменного зуба внутри первой из сохранившихся альвеол.
Таким образом, по совокупности указанных признаков можно уверенно относить вид, исходно описанный как Bolosaurus traati, к семейству Diadectidae, и полагать его первой формой диадектоморф, установленной с территории Восточной Европы. Вслед за Ивахненко (2008) мы относим этот вид к роду Stephanospondylus, однако здесь, вслед за А. Ромером (Romer, 1925), принимаем принадлежность последнего к примитивным диадектидам, а не к болозавридам. В пользу этого однозначно свидетельствует наличие у типового вида Stephanospondylus – S. pugnax – складчатой структуры оснований зубов (отчетливо фиксируется на всех зубах maxillaria и dentale: Stappenbeck, 1905, фиг. 2; Kissel, 2010, фиг. 8 ), характер дифференциации зубов буккальной серии, наличие у них характерных субвертикальных площадок стирания, аналогичных таковым Desmatodon hesperis (Vaughn, 1969), а также отсутствие выраженного наклона у увеличенных передних максиллярных зубов.
Несмотря на неполноту голотипа ПИН, № 3318/1, можно заключить, что число максиллярных зубов у S. traati, с большой вероятностью, соответствовало таковому S. pugnax. У описанного разнообразия диадектид их число невелико и во взрослом состоянии варьирует в узком диапазоне от 11 до 13, проявляя при этом коррелятивную зависимость от степени специализации зубной системы (Case, Williston, 1912; Vaughn, 1972; Berman, Sumida, 1995; Berman et al., 1998, 2004; Kissel, 2010). При этом, как правило, два зуба в переднем отделе максиллы сильно увеличены и отклоняются латерально по сравнению с другими максиллярными зубами. По форме и размеру они имитируют крупные долотовидные премаксиллярные зубы, в связи с чем получили название “резцевидных”. В большинстве случаев “резцевидные” зубы занимают первые две альвеолы в maxillare (Stappenbeck, 1905; Vaughn, 1969; Berman, Sumida, 1995; Berman et al., 1998; Kissel, 2010), однако у сравнительно примитивных форм по этому признаку отмечается бóльшая вариабельность: в частности, у Orobates они могут занимать позиции до четвертой альвеолы включительно (Berman et al., 2004).
Несмотря на то, что у голотипа ПИН, № 3318/1 два первых сохранившихся зуба, занимающих третью и четвертую сохранившиеся альвеолы, заметно отклоняются латерально по сравнению с задними зубами челюсти (рис. 1, д), они не могут трактоваться как “резцевидные”, поскольку: 1) не имеют сходства с премаксиллярными зубами диадектид и, за вычетом зауженной вершинки, своей “лопатовидной” формой и разворотом коронки больше соответствуют зубам буккальной серии; 2) не отличались по размеру среди соседних зубов (что следует из размера альвеол); 3) имеют характерную для буккальных зубов форму износа в виде фасеток стирания с лингвальной стороны (рис. 2, д), описанных, в частности, для буккальных зубов взрослой особи Desmatodon hesperis (Vaughn, 1969; Kissel, 2010, фиг. 18 ); 4) форма закладки сменного зуба в первой альвеоле не имеет сходства ни с премаксиллярными (Reisz, Sutherland, 2001), ни с “резцевидными” (если ориентироваться на существующие описания) максиллярными зубами других диадектид.
Кроме того, в отличие от наблюдаемого у голотипа ПИН, № 3318/1, у всех диадектид, для которых строение верхнечелюстных костей известно, “резцевидные” зубы приурочены к более низкой передней части кости, т.е. расположены впереди от максимального подъема lamina ascendens maxillare. Так, у Diadectes absitus в этой части максиллы размещаются не менее пяти зубов (Berman et al., 1998), у D. sanmiguelensis – четыре зуба (Lewis, Vaughn, 1965), у Orobates pabsti – четыре–пять зубов (Berman et al., 2004), у Stephanospondylus pugnax – четыре зуба, у голотипа Desmatodon hesperis – всего три зуба, разделенных, однако, увеличенным интервалом (Vaughn, 1969; Kissel, 2010)11.
У Stephanospondylus traati максимальный подъем lamina ascendens maxillare находится напротив границы первой (десятой с конца) и второй представленных на образце альвеол (рис. 1, а), из чего следует, что “резцевидные” максиллярные зубы у голотипа ПИН, № 3318/1, как и предполагалось выше, отсутствуют, а число максиллярных зубов составляло 12 или 13.
Таким образом, буккальная серия зубов представлена на голотипе ПИН, № 3318/1 полностью или почти полностью, причем по размеру альвеол можно полагать, что на всем протяжении сохранившегося отдела maxillare зубы градационно уменьшались в размере до конца зубного ряда. Данная особенность отличает S. traati от всех известных диадектид (у которых самые крупные буккальные зубы максиллы расположены в средней или задней частях зубного ряда) за исключением S. pugnax, у которого, однако, размерная градация зубов буккальной серии выражена значительно слабее (Stappenbeck, 1905; Vaughn, 1969, 1972; Berman, Sumida, 1995; Berman et al., 2004; Kissel, 2010). В настоящее время различный характер размерной дифференциации зубов в задней части maxillare является, по сути, единственным очевидным отличием видов рода Stephanospondylus.
В строении самих зубов S. traati принципиально отметить отсутствие следов их “моляризации”, которая проявляется у диадектид в наличии у коронок дополнительных вершинок и/или выраженных плечевидных уступов (Cope, 1896; Vaughn, 1969, 1972). Хотя значительная часть зубов у голотипа ПИН, № 3318/1 утеряна, а вершинки всех сохранившихся зубов повреждены вследствие стирания и во время отбора образца, узкий профиль коронок в их апикальной части позволяет предполагать отсутствие здесь подобных образований, что подтверждается и их отсутствием на коронке сменного зуба в передней альвеоле (рис. 2, ж). Другие признаки зубной системы, коррелятивно связанные с прогрессивной “моляризацией” и наблюдаемые у специализированных диадектид, также не характерны для S. traati. Так, исходя из опубликованных данных, можно заключить, что усложнение формы зубов в эволюции диадектид сопровождается: 1) удлинением их оснований в лабио-лингвальном направлении и приобретением их строго перпендикулярной ориентации по отношению к краям челюсти в местах крепления, 2) увеличением дистанции между зубами в дефинитивном состоянии, 3) увеличением относительного размера зубов в задней половине maxillare, 4) увеличением латеральной флексии размещенной здесь части зубного ряда, а также 5) увеличением длины корневой части зубов по отношению к их полной высоте, как это наблюдается у Diadectes spp. и Alveusdectes, у которых зубы заглублены в челюсть до основания коронки (LeBlanc, Reisz, 2013, фиг. 4 ; Liu, Bever, 2015). Можно предполагать, что последняя особенность складывалась в эволюции диадектид по мере специализации заднечелюстных зубов, что привело к увеличению нагрузок на зубную систему в связи с изменением принципов ее функционирования, увеличению глубины имплантации зубов за счет подъема альвеолярного края в направлении коронки и, как следствие, увеличению массивности элементов, несущих маргинальное озубление.
Отсутствие у S. traati всех перечисленных характеристик хорошо согласуется с предположением о простой форме буккальных зубов у данного вида, а также указывает и на его базальное положение внутри семейства Diadectidae, заставляя сближать с S. pugnax и сравнивать в первую очередь с другими примитивными представителями семейства, такими как Orobates pabsti и Desmatodon hesperis, у которых “моляризация” выражена слабо или проявляется локально по длине зубного ряда (Vaughn, 1969, 1972; Berman, Sumida, 1995; Berman et al., 2004; Kissel, 2010). Другими примитивными особенностями S. traati, общими с перечисленными таксонами, являются плотная посадка зубов в челюстях, смещение их лабиальной стороны вперед по отношению к лингвальной (сопоставимая с Desmatodon, но выраженная слабее, чем у Orobates), и слабое развитие корневой части зубов, высота которой составляет только треть их полной высоты.
Онтогенетический фактор. Как указывалось выше, длина голотипа ПИН, № 3318/1 составляет всего 29 мм, что, учитывая недостающие зубы, позволяет оценить длину полного maxillare примерно в 35 мм. Это несколько меньше правого maxillare голотипа S. pugnax (длина около 45 мм) и существенно меньше размера верхнечелюстных костей большинства описанных экземпляров других видов семейства, что требует учета при сравнении возрастных (размерных) характеристик.
Среди Diadectidae наиболее полные данные по онтогенезу зубной системы получены для Desmatodon hesperis (Vaughn, 1972; Berman, Sumida, 1995). Для этой формы минимальный известный размер верхнечелюстной кости (UCLAVP 1748) составляет 42 мм. На основе сравнения этого образца с более крупным по размеру голотипом D. hesperis показано (Vaughn, 1972), что для ювенильных особей этого вида в строении верхнечелюстной кости характерны: 1) меньшее число зубов (девять, включая “резцевидные”), 2) увеличенная дистанция между зубами, 3) увеличенная относительная высота “резцевидных” зубов и их более заостренная клыкоподобная форма, 4) более узкая, удлиненная лабио-лингвально форма оснований буккальных зубов, и 5) отсутствие на них площадок стирания.
По всем перечисленным параметрам, за исключением недоступного для наблюдения признака (3), голотип S. traati не соответствует ювенильным кондициям примитивных форм семейства, хотя, с большой вероятностью, принадлежал не вполне взрослой особи, что следует из типичных размеров большинства диадектид во взрослом состоянии и более крупного размера типового вида рода – S. pugnax.
В частности, не может рассматриваться как ювенильный признак голотипа ПИН, № 3318/1 отсутствие проявлений “моляризации” буккальных зубов. Организация зубного ряда, при которой наблюдается градационное уменьшение (и упрощение формы) зубов в каудальном направлении, наблюдаемое у S. traati, по-видимому, в принципе не подразумевает возможности их “моляризации”, которая в эволюции и онтогенезе диадектид приоритетно проявляется у самых крупных буккальных зубов, приуроченных к задней половине maxillare (и dentale). Так, наиболее крупные и сложно устроенные буккальные зубы у D. hesperis (в том числе у ювенильного экземпляра UCLA VP 1748) расположены в задней половине maxillare и уже несут дополнительные бугры и вершинки с лингвальной и лабиальной сторон от основного конуса (Vaughn, 1969, 1972). Максиллярные зубы Orobates pabsti имеют только вздутие с лабиальной стороны от вершины и, таким образом, “моляризованы” слабее таковых большинства диадектид, но при этом также приурочены к задней части зубного ряда (Berman et al., 2004).
На раннюю “моляризацию” зубов в онтогенезе более прогрессивных представителей семейства (род Diadectes) указывает состояние зубной системы у D. sanmiguelensis. Наиболее специализированные зубы у относящегося к этому виду ювенильного экземпляра MCZ 2989 (голотип; длина maxillare около 42 мм, если ориентироваться на масштаб приводимых изображений: Levis, Vaughn, 1964, фиг. 7 ) также располагаются несколько позади середины maxillare, и имеют при этом покатый выступ лабиально от основной вершины зуба, а также выраженную дополнительную вершинку лингвально от нее. Сложное строение зубов наблюдается также и у молодой особи Diadectes absitus при длине maxillare около 50 мм (экз. MNG 8747; Berman et al., 1998). Значительное сходство в строении передних максиллярных зубов с голотипом ПИН, № 3318/1 обнаруживает соразмерный фрагмент челюсти Diadectes sp. из нижней перми Техаса в США (Berman, Sumida, 1995, фиг. 5 ), однако образец не сохранил заднюю часть зубного ряда, в связи с чем не может выступать в качестве объекта для сравнения степени “моляризации” зубов у базальных и специализированных диадектид на ранних стадиях онтогенеза.
Таким образом, тезис о приоритетном усложнении именно задних буккальных зубов в эволюции прогрессивных Diadectidae подтверждается всеми имеющимися (в т.ч. онтогенетическими) данными, однако “моляризация” задних буккальных зубов не была характерна для базальных форм семейства (род Stephanospondylus). Можно заключить, что сравнительно простая форма зубов у голотипа S. traati и равномерное уменьшение их размера в каудальном направлении не являются ювенильным состоянием, но качественно совпадают с наблюдаемым у S. pugnax, и могут рассматриваться как диагностические признаки рода Stephanospondylus. Следует также отметить, что примитивность S. traati хорошо согласуется с ранним – ассельским – возрастом находки. Это, а также архаичность зубной системы, позволяют рассматривать данный вид как наиболее примитивную форму Diadectidae, если не считать описанного на основании челюстного материала Ambedus pusillus, происходящего, однако, из существенно более высокого стратиграфического уровня (формация Грини, Леонардиан; США: Kissel, Reisz, 2004; Kissel, 2010).
О положении рода Ambedus Kissel et Reisz, 2004. Несмотря на то, что род Ambedus демонстрирует отличное от остальных диадектид строение верхнечелюстной кости и очень примитивное состояние зубной системы, он включается в данное семейство как его базальный представитель (Kissel, Reisz, 2004; Kissel, 2010). Действительно, для Ambedus характерна едва выраженная “моляризация” маргинальных зубов, которая проявляется лишь в наличии покатых уступов на лингвальной и лабиальной сторонах коронки, разделенных узким режущим кантом, ориентированным продольно к зубному ряду (Kissel, Reisz, 2004, фиг. 2; рис. 3, б). Высота корневой части зубов у Ambedus составляла не более трети их полной высоты, что следует из соответствующей высоты зубной площадки. Для рода предполагается наличие у зубов корневой части, что подразумевает их текодонтное крепление (Kissel, Reisz, 2004, с. 199), однако ни один из экземпляров выборки не демонстрирует эту особенность.
Рис. 3.
Различия в ориентации структур коронки средних максиллярных зубов: а – у примитивных диадектид на примере Stephanospondylus traati; б – у Ambedus pussilus (по: Kissel, Reisz, 2004, с изменениями).
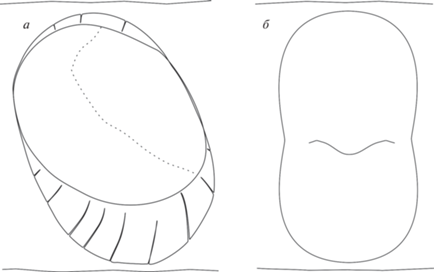
Кроме того, для Ambedus отмечается сравнительно большое число зубов: не менее 13 в maxillare и 22 в dentale, при полном отсутствии увеличенных “резцевидных” зубов как в верхней, так и в нижней челюстях. Дифференциация челюстных зубов выражается преимущественно в равномерном увеличении их размера в направлении заднего края челюстей; при этом самые крупные максиллярные и нижнечелюстные зубы, как у специализированных диадектид, расположены в задней половине зубного ряда и несут следы прижизненного износа (Kissel, Reisz, 2004; Reisz, 2006).
По совокупности перечисленных параметров род Ambedus дистанцирован от остальных родов Diadectidae настолько, что это могло бы служить основанием для его выделения в отдельное семейство, однако, по нашему мнению, правомерность его помещения в состав отряда Diadectomorpha более чем сомнительна. Так, род Ambedus характеризуется прямым в дорсальной проекции maxillare, что указывает на отсутствие выраженного расширения черепа на границе преорбитального и орбитального отделов, которое наблюдается у всех представителей Diadectomorpha, включая примитивные клады – Limnoscelis и Tseajaia. Верхнечелюстная кость A. pusillus высокая почти на всем своем протяжении и не образует характерного для диадектид понижения в своей передней части. Особое внимание обращает на себя отсутствие увеличенных зубов в переднем отделе черепа, что необычно, поскольку этот признак является одной из базовых характеристик организации челюстного аппарата всех, в том числе более базальных, диадектоморф (Williston, 1911; Romer, 1946; Vaughn, 1964). Стоит отдельно отметить, что для растительноядных форм отряда (Diadectidae) увеличенные зубы ростральной части черепа являются функционально значимой составляющей зубной системы, обеспечивавшей, совместно с премаксиллярными зубами, захват, срезание и/или фиксацию растительных объектов, на что указывает их интенсивное стирание (Reisz, Sutherland, 2001; Reisz, 2006). Кроме того, важная роль увеличенных максиллярных “резцевидных” зубов в ранней филогении отряда подтверждается онтогенетическими данными, а именно – их более выраженным развитием у ювенильной особи Desmatodon hesperis (Vaughn, 1972). Таким образом, увеличенные зубы переднего отдела черепа у Diadectomorpha унаследованы от предкового таксона и были дополнительно модифицированы в связи с переходом к фитофагии, в связи с чем их отсутствие у Ambedus при той же трофической специализации не имеет удовлетворительного объяснения.
Кроме того, для Ambedus pusillus с большой вероятностью можно исключить наличие плицидентиновой структуры зубов. В первоописании (Kissel, Reisz, 2004, с. 199) указано, что присутствие складчатости дентина у экземпляров типовой серии (пять челюстных костей) не может быть однозначно установлено, однако на ее отсутствие указывают как нескладчатый характер сломов поврежденных зубов, так и отсутствие продольных борозд на внешней стороне тех зубов, которые сохранились полностью.
Вне зависимости от степени специализации зубной системы характер складчатости оснований зубов у диадектид остается, по-видимому, почти неизменным: складки протягиваются от начала корня, где они обособлены друг от друга, до основания коронок, где они плотно смыкаются боковыми сторонами (LeBlanc, Reisz, 2013), что приводит к утолщению стенки зубов и увеличению прочности последних в прикоронарной части. У специализированных диадектид внешние борозды могут быть скрыты для наблюдения вследствие подъема альвеолярного края к основанию коронки, однако складчатая структура зубов при этом отчетливо фиксируется на поперечных срезах (LeBlanc, Reisz, 2013, фиг. 5 ). У примитивных диадектид с неглубокой имплантацией зубов наличие плицидентиновой структуры уверенно определяется по продольным бороздам на их наружной поверхности на участке между корнем и краем эмали, как это отмечено выше у Stephanospondylus traati (рис. 2, в, е), наблюдается у S. pugnax (Stappenbeck, 1905; Kissel, 2010), а также на изолированных зубах формально не описанных примитивных диадектид из карстового местонахождения Форт Силл в Оклахоме, США (Reisz, Sutherland, 2001). Отсутствие у Ambedus таких борозд на зубах является прямым указанием на отсутствие складчатости, что плохо согласуется с положением этого рода в составе Diadectomorpha, принимая во внимание, что для данного отряда плицидентиновая структура корневой части зубов, безусловно, является плезиоморфной характеристикой, и отмечается не только у Diadectidae, но также у Limnoscelis (Williston, 1911, цит. по: Romer, 1946, с. 163; Berman et al., 2010; LeBlanc, Reisz, 2013) и Tseajaia (Vaughn, 1964).
Характер дифференциации буккальных зубов Ambedus с учетом данных по морфологии зубной системы наиболее примитивных Diadectidae – Stephanospondylus traati, Orobates pabsti и Desmatodon hesperis – также не позволяет рассматривать конструкцию Ambedus как исходную для данного семейства. Увеличение размеров зубов в направлении заднего края челюстей складывается в рассматриваемом семействе на поздних стадиях его эволюции вследствие усиления адаптации к фитофагии, что подтверждается как филогенетическими, так и онтогенетическими данными (Romer, 1952; Vaughn, 1972; Kissel, 2010). Наблюдаемая гиперспециализация зубной системы при этом выражается в реализации широкого набора коррелятивно связанных модификаций, большинство из которых не проявляются у Ambedus (например, усложнение формы зубов, увеличение глубины их имплантации, увеличение дистанции между зубами, резкое увеличение латерального изгиба задней части зубного ряда). У примитивных диадектид зубы расположены более плотно, их основания имеют выраженный (иногда очень сильно, как у Orobates) разворот в челюсти, наблюдается градационное уменьшение размера зубов с начала (Stephanospondylus) или середины (Orobates, Desmatodon hesperis) зубного ряда в каудальном направлении, которое сопровождается упрощением их формы и уменьшением размера, что также не соответствует наблюдаемому у Ambedus. В этой связи представляется оправданным трактовать увеличение размера зубов к окончанию зубного ряда у Ambedus как конвергентное состояние по отношению к Diadectidae; аналогичный характер дифференциации проявляется независимо в зубной системе многих групп тетрапод палеозоя–раннего мезозоя (Procolophonidae, Bolosaurida, Cynodontia и др.), у которых наиболее специализированные зубы локализованы сразу впереди от места крепления основных челюстных аддукторов. Характер стирания челюстных зубов A. pusillus, безусловно, является аргументом в пользу растительноядной трофической специализации вида, но не может рассматриваться как показатель его принадлежности к рассматриваемому отряду, поскольку прямой контакт зубов при питании не является уникальной особенностью диадектид и наблюдается в различных линиях палеозойских тетрапод, где формировался независимо (Reisz, 2006). Кроме того, для челюстных зубов A. pusillus свойственно продольное положение режущего канта к линии зубного ряда, в то время как у наиболее примитивных форм семейства (Stephanospondylus, Orobates) наблюдается разворот фациальной (исходно лабиальной) стороны коронок буккальных зубов в ростральном направлении, что приводит и к развороту канта, который приобретает поперечную – лабио-лингвальную – ориентировку (рис. 3, a). Это позволяет предполагать, что дополнительные структуры, появляющиеся на зубах диадектид в процессе прогрессирующей “моляризации” (лингвально и лабиально от основной вершины зуба), топографически связаны с исходным режущим кантом (как и у подавляющего большинства других тетрапод), и не гомологичны покатым уступам челюстных зубов Ambedus, которые этим кантом разделены (рис. 3, б) (Kissel, Reisz, 2004).
Исходя из перечисленного, можно заключить, что род Ambedus по строению зубной системы не соответствует ее генеральному морфотипу у Diadectomorpha по целому ряду качественных параметров, в связи с чем его помещение в состав отряда представляется неоправданным. Обоснование филогенетического положения этого рода в системе тетрапод требует более развернутого изучения таксона с привлечением дополнительных данных по морфологии зубов уже описанных диадектоморф. В настоящий момент можно отметить, что некоторые признаки Ambedus, такие как отсутствие складчатости оснований зубов, отсутствие изгиба верхнечелюстной кости и несомого ею зубного ряда, отсутствие орнаментации фациальной стороны максиллы, а также небольшой размер обнаруживают большее сходство данного рода с Bolosauridae, чем c примитивными Diadectidae, что находит также больше соответствий и в стратиграфическом распространении указанных семейств (Kissel, 2010).
В заключение можно отметить, что находка базального диадектида в ассельских отложениях Восточной Европы является указанием на обширную географию распространения данного семейства уже на ранних этапах его радиации в пределах Лавразии. Последнее могло стимулировать провинциализм эволюционного развития отдельных филогенетических направлений, и может являться одним из вероятных объяснений недавнего обнаружения специализированных диадектид (Liu, Bever, 2015) в верхней перми Китая.
Ниже приводятся ревизованные диагнозы рода Stephanospondylus и вида S. traati.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ОТРЯД DIADECTOMORPHA
СЕМЕЙСТВО DIADECTIDAE COPE, 1880
Род Stephanospondylus Stappenbeck, 1905
Типовой вид – Phanerosaurus pugnax Geinitz et Deichmuller, 1882; нижний ротлегенд Германии.
Диагноз. Буккальные зубы maxillare субконической формы, без очевидных проявлений “моляризации” в виде бугров и вершин лингвально и лабиально от центрального конуса. Размер буккальных максиллярных зубов не увеличивается в каудальном направлении.
Видовой состав. S. pugnax (Geinitz et Deichmuller, 1882) и S. traati (Tatarinov, 1974).
Сравнение. От всех родов Diadectidae отличается более простой формой буккальных максиллярных зубов и отсутствием увеличения их размера в направлении конца зубного ряда.
Stephanospondylus traati (Tatarinov, 1974)
Голотип – ПИН, № 3318/1, левое maxillare; Республика Коми, местонахождение Мылва; нижняя пермь, ассельский ярус.
Диагноз. Передние буккальные зубы maxillare значительно крупнее задних максиллярных зубов.
Сравнение. От типового вида рода Stephanospondylus – S. pugnax (Geinitz et Deichmuller, 1882) – отличается выраженным градационным уменьшением размера буккальных зубов maxillare в каудальном направлении.
Замечание. Одним из критериев отличия от S. pugnax, требующим подтверждения, является сильно отличающийся угол наклона передних и задних буккальных зубов верхнечелюстной кости.
Материал. Голотип.
Список литературы
Ивахненко М.Ф. Новые котилозавры Приуралья // Палеонтол. журн. 1973. № 2. С. 131–134.
Ивахненко М.Ф. Раннепермские элементы фаунистических комплексов тетрапод Восточной Европы // Палеонтол. журн. 1987. № 2. С. 102–111.
Ивахненко М.Ф. Ископаемые позвоночные России и сопредельных стран. Ископаемые птицы и рептилии. Часть 1 / Ред. М.Ф. Ивахненко, Е.Н. Курочкин. М.: ГЕОС, 2008. 348 с.
Ивахненко М.Ф., Голубев В.К., Губин Ю.М. и др. Пермские и триасовые тетраподы Восточной Европы. М.: ГЕОС, 1997. 216 с. (Тр. Палеонтол. ин-та РАН. Т. 268).
Ивахненко М.Ф., Твердохлебова Г.И. Ревизия пермских болозавроморфов Восточной Европы // Палеонтол. журн. 1987. № 2. С. 98–106.
Татаринов Л.П. Находка примитивного хвостатого земноводного в верхней перми Поволжья // Верхнепалеозойские и мезозойские земноводные и пресмыкающиеся. М.: Наука, 1968. С. 7–10.
Татаринов Л.П. Находка болозавра в нижней перми СССР // Палеонтол. журн. 1974. № 2. С. 144–146.
Berman D.S., Henrici A.C., Kissel R.A. et al. A new diadectid (Diadectomorpha), Orobates pabsti, from the Early Permian of Central Germany // Bull. Carneg. Mus. Natur. Hist. 2004. V. 35. P. 1–36.
Berman D.S., Reisz R.R., Scott D. et al. Early Permian bipedal reptile // Science. 2000. V. 290. P. 969–972.
Berman D.S., Reisz R.R., Scott D. Redescription of the skull of Limnoscelis paludis Williston (Diadectomorpha, Limnoscelidae) from the Pennsylvanian of Canon del Cobre, Northern New Mexico // Bull. New Mexico Natur. Hist. Sci. Mus. 2010. V. 49. P. 185–210.
Berman D.S., Sumida S.S. New cranial material of the rare diadectid Desmatodon hesperis (Diadectomorpha) from the Late Pennsylvanian of Central Colorado // Ann. Carneg. Mus. 1995. V. 64. P. 315–336.
Berman D.S., Sumida S.S., Martens T. Diadectes (Diadectomorpha: Diadectidae) from the Early Permian of central Germany, with description of a new species // Ann. Carneg. Mus. 1998. V. 67. P. 53–93.
Bulanov V.V., Kovalenko E.S., MacDougall M.J. et al. Tooth replacement and reparative dentine formation in the middle Permian bolosaurids of European Russia // Hist. Biol. 2022. https://doi.org/10.1080/08912963.2022.2067752
Case E.C. Description of the skull of Bolosaurus striatus Cope // Bull. Amer. Mus. Natur. Hist. 1907. V. 23. P. 653–658.
Case E.C., Williston S.W. A description of the skulls of Diadectes lentus and Animasaurus carinatus // Amer. J. Sci. 1912. V. 33. P. 339–348.
Case E.C., Williston S.W. Description of a nearly complete skeleton of Diasparactus zenos Case // Publ. Carneg. Inst. Wash. 1913. V. 181. P. 17–35.
Cope E.D. Descriptions of the extinct Batrachia and Reptilia from the Permian formation of Texas // Proc. Amer. Phil. Soc. 1878. V. 17. № 101. P. 505–530.
Cope E.D. Second contribution to the history of the Cotylosauria // Proc. Amer. Phyl. Soc. 1896. V. 35. P. 122–138.
Cope E.D. A revision of the Cotylosauria of North America. Washington: Carnegie Inst., 1911. 122 p.
Kissel R.A. Morphology, phylogeny, and evolution of Diadectidae (Cotylosauria: Diadectomorpha). PhD. diss. (Graduate Department of Ecology and Evolutionary Biology, University of Toronto). 2010. 185 p.
Kissel R.A., Reisz R.R. Ambedus pusillus, new genus, new species, a small diadectid (Tetrapoda: Diadectomorpha) from the Lower Permian of Ohio, with a consideration of diadectomorph phylogeny // Ann. Carneg. Mus. 2004. V. 73. P. 197–212.
LeBlanc A.R.H., Reisz R.R. Periodontal ligament, cementum, and alveolar bone in the oldest herbivorous tetrapods, and their evolutionary significance // PLOS One. 2013. V. 8. № 6. e74697. https://doi.org/10.1371/journal.pone.0074697
Lewis G.E., Vaughn P.P. Early Permian vertebrates from the Cutler Formation of the Placerville Area, Colorado // U.S. Geol. Surv. Prof. Pap. 1965. № 503-C. P. 1–50.
Li J.L., Cheng Z.W. The first discovery of bolosaurs from Upper Permian of China // Vertebr. PalAsiat. 1995. V. 33. P. 17–23.
Liu J., Bever G.S. The last diadectomorph sheds light on Late Palaeozoic tetrapod biogeography // Biol. Lett. 2015. V. 11: 20150100.
Reisz R.R. Origin of dental occlusion in tetrapods: signal for terrestrial vertebrate evolution? // J. Exp. Zool. 2006. V. 306. P. 261–277.
Reisz R.R., Barkas V., Scott D. A new Early Permian bolosaurid reptile from the Richards Spur Dolese Brothers Quarry, near Fort Sill, Oklahoma // J. Vertebr. Paleontol. 2002. V. 22. № 1. P. 23–28.
Reisz R.R., Müller J., Tsuji L., Scott D. The cranial osteology of Belebey vegrandis (Parareptilia: Bolosauridae), from the Middle Permian of Russia, and its bearing on reptilian evolution // Zool. J. Linn. Soc. 2007. V. 151. P. 191–214.
Reisz R.R., Sues H.-D. Herbivory in late Paleozoic and Triassic terrestrial vertebrates // Evolution of herbivory in terrestrial vertebrates / Ed. H.-D. Sues. Cambridge: Cambr. Univ. Press, 2000. P. 9–42.
Reisz R.R., Sutherland T.E. A diadectid (Tetrapoda, Diadectomorpha) from the Lower Permian fissure fills of the Dolese Quarry, near Richards Spur, Oklahoma // Ann. Carneg. Mus. 2001. V. 70. № 2. P. 133–142.
Romer A.S. Permian amphibian and reptilian remains described as Stephanospondylus // J. Geol. 1925. V. 33. P. 447–467.
Romer A.S. The primitive reptile Limnoscelis restudies // Amer. J. Sci. 1946. V. 244. № 3. P. 149–188.
Romer A.S. Late Pennsylvanian and Early Permian vertebrates of the Pittsburg-West Virginia Region // Ann. Carneg. Mus. 1952. V. 33. P. 47–110.
Snyder A.J., LeBlanc A.R.H., Jun C. et al. Thecodont tooth attachment and replacement in bolosaurid parareptiles // PeerJ. 2020. 8:e9168. https://doi.org/10.7717/peerj.9168
Stappenbeck R. Uber Stephanospondylus n. g. und Phanerosaurus H. v. Meyer // Z. Dtsch. Geol. Ges. 1905. Bd 57. S. 380–437.
Vaughn P.P. Vertebrates from the Organ Rock Shale of the Cutler Group, Permian, Monument Valley and vicinity, Utah and Arizona // J. Paleontol. 1964. V. 38. № 3. P. 567–583.
Vaughn P.P. Upper Pennsylvanian vertebrates from the Sangre de Cristo Formation of central Colorado // Contrib. Sci. Los Angeles Cty Mus. Natur. Hist. 1969. V. 164. P. 1–28.
Vaughn P.P. More vertebrates, including a new microsaur, from the Upper Pennsylvanian of Central Colorado // Contrib. Sci. 1972. № 223. P. 1–30.
Watson D.M.S. On Bolosaurus and the origin and classification of reptiles // Bull. Mus. Comp. Zool. 1954. V. 111. P. 297–449.
Williston S.W. A new family of reptiles from the Permian of New Mexico // Amer. J. Sci. 1911. V. 4. № 31. P. 378–398.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал