

Палеонтологический журнал, 2023, № 2, стр. 100-114
Морская черепаха Tasbacka aldabergeni Nessov, 1987 из нижнепалеогеновых отложений местонахождения Кудиновка (Ростовская область, Россия)
Е. А. Звонок a, *, К. С. Беницкий b, И. Г. Данилов c
a Луганский государственный педагогический университет
ЛНР 91006 Луганск, Россия
b Независимый исследователь
346132 Ростовская обл., Миллерово, Россия
c Зоологический институт РАН
199034 Санкт-Петербург, Россия
* E-mail: evgenij-zvonok@yandex.ru
Поступила в редакцию 14.09.2022
После доработки 10.11.2022
Принята к публикации 10.11.2022
- EDN: LEUBEW
- DOI: 10.31857/S0031031X23020149
Аннотация
Описаны ископаемые остатки черепах из местонахождения Кудиновка (палеоген, палеоцен или ипр; Милеровский р-н, Ростовская обл., Россия). Эти остатки, представленные 16 экз., отнесены к морской черепахе Tasbacka aldabergeni Nessov, 1987 (Cheloniidae), ранее достоверно известной только из типового местонахождения Джилга 1 (палеоген, поздний танет–ранний ипр; Южный Казахстан). Один из описываемых экземпляров представляет собой большую часть посткраниального скелета в фосфоритовой конкреции и является наиболее полным экземпляром посткрания Tasbacka aldabergeni, который дает информацию о ранее неизвестных деталях строения этого вида. Новые материалы расширяют наши знания о географическом распространении Tasbacka aldabergeni и рода Tasbacka в целом.
ВВЕДЕНИЕ
Находки палеогеновых черепах на территории Восточной Европы плохо известны и представлены в основном фрагментарными остатками Cheloniidae, Dermochelyidae, Testudinidae, Trionychidae и не определенных черепах (см. Zvonok, Danilov, 2018; Звонок, Данилов, 2019, 2020; Звонок и др., 2019).
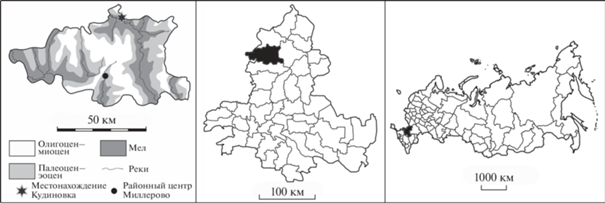
Новый материал по палеогеновым черепахам происходит из местонахождения Кудиновка, расположенного в карьере по добыче песка примерно в 2 км к востоку от одноименного хутора, в Миллеровском р-не Ростовской обл. России (49°12′37.3′′ N, 40°31′48.1′′ E; рис. 1). При разработке карьера в 2015–2016 гг. бульдозером была произведена расчистка примерно по продуктивному слою. В результате на дневной поверхности оказались многочисленные фосфоритовые конкреции с фоссилиями. Сборы остатков ископаемых животных велись в 2018 (К.Б.) и 2019 (Е.З. и К.Б.) гг. В их числе оказалось 16 экз., относящихся к морским черепахам (Cheloniidae) Tasbacka aldabergeni Nessov, 1987, описание которых является основной целью настоящей статьи. Данные о других ископаемых организмах, встречающихся в этом местонахождении, см. в описании геологического разреза ниже. Некоторые общие сведения о местонахождении также изложены в статье К.С. Беницкого и Е.А. Звонка (2020).
Рис. 1.
Слева – местонахождение Кудиновка на геологической картосхеме Миллеровского р-на; посередине – Миллеровский район на административной картосхеме Ростовской обл.; справа – Ростовская область на административной картосхеме России.
Остатки Tasbacka aldabergeni (типовой вид рода Tasbacka Nessov, 1987) впервые были описаны по массовым материалам (более 1700 костей) Л.А. Несовым (1987) из танет–ипрского местонахождения Джилга 1а Южного Казахстана. Позже А.О. Аверьянов (Averianov, 2002) упоминал фрагменты черепа и панциря T. aldabergeni из вышележащих уровней ипрского возраста этого же местонахождения (Джилга 1б, Джилга 2) и фрагменты панциря из отложений танетского возраста местонахождений Дарбаза 1 и 2 в 7–9 км юго-восточнее Джилги. Из них была изображена только нухальная пластинка из Джилги 1б (Averianov, 2002, рис. 5), не отличающаяся от таковой T. aldabergeni из типового местонахождения. Систематическая принадлежность остатков T. aldabergeni из остальных упомянутых местонахождений не подтверждена (Данилов и др., 2017). Помимо этого, остатки “морских черепах, похожих на Tasbacka aldabergeni Ness., но вдвое более крупных”, или cf. Tasbacka sp., указаны без описаний и изображений из местонахождения Малая Ивановка, Волгоградская обл., Россия; верхний палеоцен, палеоген (Несов, Ярков, 1989, с. 80; Averianov, 2002, с. 142; Данилов и др., 2017, с. 182).
Другие представители рода Tasbacka известны из палеоцена Марокко, США и, возможно, Дании (см. ниже “Обсуждение”). Таким образом, материалы данной статьи описывают первую достоверную находку представителя этого рода в Восточной Европе.
Геологический разрез местонахождения Кудиновка изображен на рис. 2. В нем снизу вверх залегают следующие осадочные горные породы:
Рис. 2.
Слева – геологический разрез местонахождения Кудиновка; справа – фотография слоя с фоссилиями и близко расположенных слоев. Описание разреза см. в тексте.

1. Слой белых мелоподобных известняков меловой системы видимой мощностью свыше 30 м. В верхней части слоя многочисленные остатки головоногих моллюсков Belemnella, стратиграфическое распространение которых ограничено маастрихтским ярусом (Christensen, 1996).
2. Слой серо-зеленой глины мощностью около 15 см.
3. Слой разнозернистого кварцевого песка, преимущественно бурого цвета, мощностью около 15 см; содержит окатанные конкреции фосфорита и гальку. В большей части конкреций содержатся остатки древесных растений, губок, моллюсков, крупных десятиногих ракообразных, рыб и черепах. В слое также попадаются зубы и кости рыб, зубы крокодилов и фрагменты костей черепах вне конкреций.
4. Слой желтого кварцевого песка видимой мощностью свыше 7 м. В верхней части содержит зубы акул.
5. Несогласно залегающий слой красновато-бурой глины плейстоценового отдела видимой мощностью до 4 м. Содержит фоссилизированные кости и зубы лошадей, носорогов, бизонов.
Галечниково-фосфоритовые слои, подобные продуктивному слою местонахождения Кудиновка, в Миллеровском р-не известны в основании палеоцена, а также каневской и бучакской свит, обычно относимых к ипрскому ярусу и низам лютетского яруса, поэтому литологические особенности продуктивного слоя не позволяют отнести его к какому-либо конкретному стратону в пределах палеоцена–нижнего лютета (Геологическая карта…, 1963).
Для уточнения геологического возраста продуктивного слоя использовались зубы акул Otodus Agassiz, 1843 sensu Zhelezko et Kozlov, 1999, определенные по крупному размеру, толстым и выпуклым с двух сторон коронкам и зазубренности на коронке и боковых зубцах у некоторых экземпляров (Железко, Козлов, 1999). Зубы этих акул найдены как в продуктивном (3), так и в вышележащем (4) слоях в карьере местонахождения Кудиновка и в еще одном карьере в 1.5 км на северо-запад. Зазубренность на коронках и боковых зубцах Otodus из Кудиновки варьирует от почти полного отсутствия до хорошо развитой. Такая изменчивость характерна для средне-позднеипрских представителей рода (Железко, Козлов, 1999; Беницкий, Звонок, 2020). В то же время, тип сохранности всех найденных зубов Otodus, а также двух не находившихся в конкрециях фрагментов костей черепах, отличается от сохранности остатков позвоночных в конкрециях. Почти все остатки позвоночных вне конкреций имеют более светлый окрас и хорошо сохранившуюся структуру губчатой ткани, тогда как все остатки позвоночных в конкрециях в среднем имеют более темный окрас и сильнее разрушенную губчатую ткань кости. Кроме того, конкреции окатаны и явно переотложены.
Таким образом, большинство остатков позвоночных продуктивного слоя вне конкреций имеет средне-позднеипрский возраст, а остатки позвоночных в конкрециях, в т.ч. остатки черепах, скорее всего, переотложены из более древних ипрских или палеоценовых отложений.
МАТЕРИАЛ И МЕТОДЫ
Агломераты костей черепах из местонахождения Кудиновка представляют собой скопления близких по анатомическому положению и анатомически соразмерных костей в конкрециях, каждое из которых, по-видимому, принадлежит одной особи черепахи. Элементы скелета на поверхности конкреций видны в виде отпечатков, вскрытых полостей от раскрошившихся костей и участков поверхности или слома костей в различных сечениях. Большая часть костей экз. ZIN PH, №№ 1–2/283 скрыта в толще конкреции. Для этих экземпляров было выполнено CT-сканирование с использованием медицинского томографа Toshiba Aquilon 64 с толщиной среза 0.5 мм. В результате получено 1464 изображений (слайдов) в DICOM формате для экз. ZIN PH, № 1/283 и 953 изображения для экз. ZIN PH, № 2/283. Измерения изображений на слайдах были выполнены с помощью RadiAnt DICOM Viewer 2020.2. Для получения максимально полных сведений об анатомии экз. ZIN PH №№ 1–2/283 фотографии их наружной поверхности совмещены с результатами CT-сканирования (рис. 3; 4, а–д).
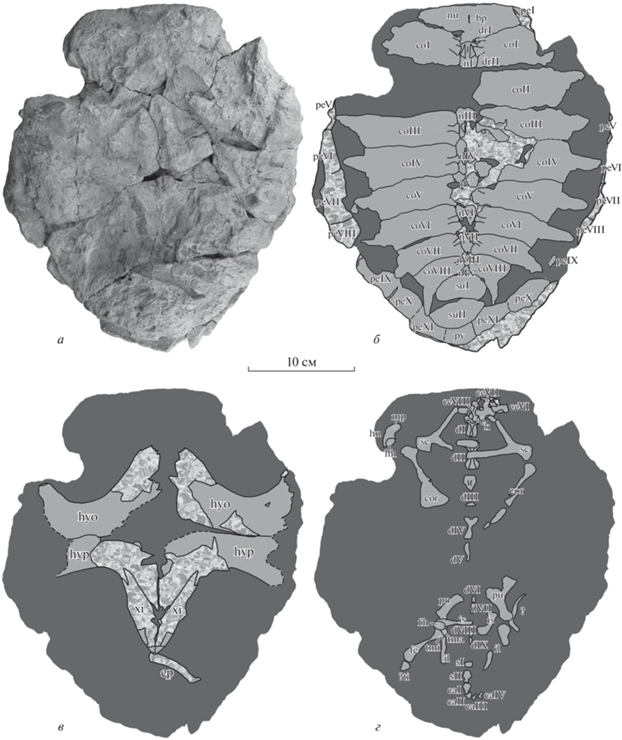
Рис. 3.
Tasbacka aldabergeni Nessov, 1987, экз. ZIN PH, № 1/283, бόльшая часть посткраниального скелета одной особи в фосфоритовой конкреции с вентральной стороны: а – общий вид конкреции; б – карапакс; в – пластрон; г – непанцирный посткраний [заливка на фиг. б–г: темно-серым цветом – горная порода; светло-серым цветом – кость внутри конкреции (видна на томограммах); узором – кость на поверхности конкреции]; Россия, Ростовская обл., местонахождение Кудиновка; палеоцен–ипр. Обозначения: bp – основание выступа для VIII шейного позвонка; caI–caIV – I–IV хвостовые позвонки; ceVI–ceVIII – VI–VIII шейные позвонки; coI–coVIII – I–VIII костальные пластинки; cor – коракоид; dI–dIX – I–IX спинные позвонки; ep – эпипластрон; fe – бедренная кость; fh – головка бедренной кости; hh – головка плечевой кости; hu – плечевая кость; hyo – гиопластрон; hyp – гипопластрон; il – подвздошная кость; is – седалищная кость; k – вентральный киль шейного позвонка; mp – медиальный отросток; nI, nIII–nIX – I, III–IX невральные пластинки; nu – нухальная пластинка; peI, peV–peXI – I, V–XI периферальные пластинки; pu – лобковая кость; sc – лопатка; tma – большой трохантер; tmi – малый трохантер; py – пигальная пластинка; drI, drII – I, II спинные ребра; sI, sII – I, II крестцовые позвонки; suI, suII – I, II супрапигальные пластинки; ?ti – (?)большая берцовая кость; xi – ксифипластрон.
Материалы, описываемые и упоминаемые в настоящей статье, хранятся в следующих учреждениях: ЦНИГР музей – Центральный научно-исследовательский геологоразведочный музей им. Ф.Н. Чернышева, С.-Петербург, Россия; IRSNB – Королевский бельгийский ин-т естественных наук, Брюссель, Бельгия; MAE – Музей Эльбёфа, Эльбёф, Франция; MHM – Музей Морс, Нюкебинг-Морс, Дания; NHMUK – Музей естественной истории, Лондон, Великобритания; ZIN PH – Зоологический ин-т РАН, палеогерпетологическая коллекция, С.-Петербург, Россия.
Для сравнения использовались данные по следующим таксонам Cheloniidae: Argillochelys antiqua (Konig, 1825) из ипрской формации “Лондонские глины” (London Clay, Англия; Owen, Bell, 1849; Lydekker, 1889), а также верхнего лютета – нижнего бартона киевской свиты местонахождения Вышгород (Украина; Zvonok, Danilov, 2017); Cabindachelys landanensis Myers et al., 2017 из датского яруса формации Ландана (Landana, Ангола; Myers et al., 2017); Eochelone spp.: E. brabantica Dollo, 1903 из нижнего лютета Брюссельской (Brussel) формации (Бельгия) и E. voltregana Lapparent de Broin et al., 2018 из приабона формации “мергели Вик-Манльеу” (Vic-Manlleu marls, Испания; Lapparent de Broin et al., 2018); Eretmochelys imbricata (Linnaeus, 1766) (Zangerl, 1953: рис. 63c ); Erquelinnesia gosseleti (Dollo, 1886) из танета нижней части Ланденской серии (Landen Group, Бельгия; Zangerl, 1971), а также его вероятные синонимы Glossochelys planimentum (Owen, 1842) и отчасти Puppigerus crassicostatus (Owen, 1849) из ипрских отложений местонахождения Харидж (Harwich, Англия; Owen, Bell, 1849); Euclastes spp.: Euclastes acutirostris Jalil et al., 2009 из палеоцена бассейна Уле Абдун (Oulad Abdoun Basin, Марокко; Jalil et al., 2009) и E. wielandi (Hay, 1908) из типовой маастрихт-датской формации Хорнерстаун (Hornerstown, США; Ullmann, Carr, 2021); “Euclastes” douvillei de Stefano, 1902 из ипра бассейна Гафса-Митлави (Gafsa-Metlaoui, Тунис; Bergounioux, 1952); Osonachelus decorata Lapparent de Broin et al., 2014 из приабона формации “мергели Вик-Манльеу” (Испания; Lapparent de Broin et al., 2014); Puppigerus camperi (Gray, 1831) из ипрской формации “Лондонские глины” (Англия), а также лютетской серии Браклшем (Bracklesham Group, Англия; Owen, Bell, 1849; Lydekker, 1889; Moody et al., 2015), нижнелютетской Брюссельской формации (Бельгия; Moody, 1974), ипра бассейна Уле Абдун (Марокко; Tong et al., 2012), ипрской формации Нейнджемой (Nanjemoy Formation, США; Weems, 1999, 2014), нижнелютетского местонахождения Иково (Россия; Zvonok et al., 2013), пограничного лютет–бартонского местонахождения Джерой 2 (Узбекистан; Аверьянов, 2005); кроме того, для сравнения использовались фотографии и 3D модели P. camperi экз. IRSNB, №№ R 0072-0076, 0080 (Брюссельские Пески, Бельгия), загруженные с сайта IRSNB (virtualcollections.naturalsciences.be), и личные наблюдения по P. nessovi Averianov, 2005 из колл. ZIN PH, № 46 (местонахождение Джерой 2, Узбекистан) и ZIN PH, № 145 (местонахождение Иково, Россия); Tasbacka spp.: T. aldabergeni Nessov, 1987 из верхнего танета или нижнего ипра местонахождения Джилга 1 (Казахстан; Несов, 1987; личные наблюдения по типовым материалам из колл. ЦНИГР музея, № 12175); T. danica Karl et Madsen, 2012 из нижнего ипра местонахождения Эйерслев Моклэйпит формации Фур (Ejerslev Moclaypit locality of Fur Formation, Дания; Karl, Madsen, 2012); T. ouledabdounensis Tong et Hirayama, 2003 из дания бассейна Оулед Абдун (Марокко; Tong, Hirayama, 2002); T. ruhoffi (Weems, 1988) из танетского местонахождения Пискатауэй Крик формации Акия (Piscataway Creek locality of Aquia Formation, Мэриленд, США; Weems, 1988, 2014).
При описании элементов посткраниального скелета черепах названия морфологических структур панциря даются по И.Г. Данилову с соавт. (2017), а непанцирного посткрания – по В.Б. Суханову (1964). Таксономия черепах приводится по Данилову с соавт. (2017).
ОТРЯД TESTUDINES
СЕМЕЙСТВО CHELONIIDAE OPPEL, 1811
Род Tasbacka Nessov, 1987
Tasbacka aldabergeni Nessov, 1987
Tasbacka aldabergeni: Несов, 1987, с. 82, рис. 1–3, табл. VII–VIII ; Averianov, 2002, c. 142, рис. 5; Данилов и др., 2017, с. 163, рис. 37 .
Голотип – ЦНИГР музей, № 1/12175, череп; Казахстан, Туркестанская обл., Приташкентские Чули, в 7 км к северу от ж/д ст. Джилга, местонахождение Джилга 1а; палеоген, верхний палеоцен или ?нижний эоцен, танет или ?ипр, белые кварцевые пески (Несов, 1987; геологический возраст см.: Данилов и др., 2017).
Описание (рис. 3, 4). Описывается только новый материал. Экз. ZIN PH, № 1/283 (рис. 3) – большая часть посткраниального скелета одной особи в фосфоритовой конкреции. В конкреции длиной 29 см (рис. 3, а) сохранились следующие части посткрания: панцирь, кроме II невральной пластинки и правой II костальной пластинки, левых II–IV периферальных пластинок и правых I–IV периферальных пластинок, одного из эпипластронов и энтопластрона; часть позвоночного столба от VI шейного позвонка до IV хвостового позвонка, возможно, за исключением X спинного позвонка; пояса конечностей; правая плечевая кость, правая бедренная кость и, вероятно, правая большая берцовая кость. На поверхности конкреции полностью или частично видны все сохранившиеся краевые элементы карапакса, участок в центральной части карапакса, большая часть пластрона; остальные элементы полностью или почти полностью находятся в толще конкреции и видны на томограммах. Часть костей смещены относительно нормального положения, в т.ч. сохранившаяся часть скелета до окончательной седиментации была подвергнута компрессии с дорсальной стороны с центром в области отсутствующей правой II костальной пластинки 2. Длина карапакса (рис. 3, б) была примерно на 1 см больше длины конкреции, т.е. около 30 см.
Рис. 4.
Tasbacka aldabergeni Nessov, 1987: а–д – экз. ZIN PH, № 2/283: а – общий вид конкреции; б – все кости внутри и на поверхности конкреции; в – сохранившаяся часть симфиза зубных костей, вид снизу; г – левая часть зубной кости в сечении, вид сзади; д – правая плечевая кость, вид снизу; е–з – экз. ZIN PH, № 3/283: е – средняя часть костальной пластинки, вид сверху; ж – дистальная часть костальной пластинки, вид сверху; з – периферальные пластинки в сечении [заливка на фиг. б: темно-серым цветом – горная порода, светло-серым цветом – кость внутри конкреции (видна на томограммах), узором – кость на поверхности конкреции]; Россия, Ростовская обл., местонахождение Кудиновка; палеоцен–ипр. Обозначения: cai – canalis alveolaris inferior; hh – головка плечевой кости; cr – радиальный мыщелок; co – костальная пластинка; cu – ульнарный мыщелок; de – симфиз зубных костей; fr – свободное ребро; hu – плечевая кость; ips – интерплевральная борозда; ne – невральная пластинка; peIII, peIV, peVI – III и, вероятно, IV и VI периферальные пластинки; pl – латеральный отросток; pm – медиальный отросток; rh – головка ребра; scm – sulcus cartilaginis meckelii; ts – альвеолярная поверхность.

Нухальная пластинка сохранилась почти полностью, кроме ее передней кромки. Ширина пластинки превышала ее длину не менее чем в три раза. Постеромедиальная часть кости слегка вытянута назад до шва с I невральной пластинкой. Постнухальные фонтанели отсутствуют, в отличие от Erquelinnesia gosseleti. На вентральной поверхности сохранилось основание выступа для контакта с VIII шейным позвонком.
Невральные пластинки представлены I и III–IX элементами, из которых III–VI в разной степени повреждены разломами в конкреции. Все невральные пластинки вытянуты в передне-заднем направлении. I невральная пластинка имеет форму овала. II невральная пластинка отсутствует, но по форме примыкающих пластинок ясно, что она была шестиугольной с короткими переднебоковыми сторонами и вогнутым передним краем, как и последующие III–VIII невральные пластинки. IX невральная пластинка мелкая, овальной формы. Все невральные пластинки с плоской наружной поверхностью, без дорсального киля, в отличие от Argillochelys antiqua, Cabindachelys landanensis и Tasbacka ruhoffi.
Супрапигальных пластинок две, в отличие от Eochelone brabantica, E. voltregana и Osonachelus decorata с тремя супрапигальными пластинками. I супрапигальная пластинка при виде сверху имеет форму, близкую к трапециевидной; ее задний край немного вогнут. II супрапигальная пластинка немного шире, чем I, линзовидной формы. Она контактировала с пигальной пластинкой и задней частью XI периферальных пластинок, что из раннепалеогеновых хелониид характерно только для Puppigerus camperi и Tasbacka aldabergeni (Moody, 1974, рис. 15 ; Несов, 1987, рис. 2а).
Пигальная пластинка сохранилась почти полностью, кроме задней кромки. Пигальная пластинка длинная в передне-заднем направлении. О наличии или отсутствии вырезки заднего края неизвестно из-за его повреждения. Пигальная пластинка по всей ширине контактировала со II супрапигальной пластинкой.
Костальные пластинки представлены всеми левыми и I, III–VIII правыми элементами; латеральные части обеих I костальных пластинок повреждены; медиальные части левых III–VI костальных пластинок и правой V костальной пластинки повреждены разломами в конкреции. Все костальные пластинки имеют свободные ребра и, соответственно, принимают участие в формировании костопериферальных фонтанелей. Головка I спинного ребра мелкая, имеет большой отступ от головки II ребра, как у Eretmochelys imbricata (Zangerl, 1953, рис. 63c ). Свободное ребро VIII костальной пластинки направлено назад, где оно контактирует с XI периферальной пластинкой.
Периферальные пластинки представлены антеромедиальной частью левой I периферальной пластинки и медиальными частями V–XI периферальных пластинок обеих сторон. V периферальная пластинка самая высокая (толстая) дорсовентрально в сохранившейся части ряда; VI‒VIII периферальные пластинки последовательно уплощаются в заднем направлении; IX–XI периферальные пластинки дорсовентрально тонкие, примерно в одинаковой степени. Углубления для свободных ребер находятся в задней части V–IX периферальных пластинок и в передней части XI периферальной пластинки; X периферальная пластинка со свободным ребром не контактировала.
Большая часть пластрона (гио-, гипо- и ксифипластроны) сохранилась в прижизненном положении (рис. 3, в); один из эпипластронов (вероятно, левый) сохранился в виде отпечатка на поверхности конкреции в области заднего края ксифипластронов.
Эпипластрон узкий в передней части, его задняя часть отломана. Поверхность для контакта со вторым эпипластроном короткая, как у Tasbacka aldabergeni (Несов, 1987, табл. VIII , фиг. 21, 22 ) и в отличие от Puppigerus camperi, у которого эпипластрон более широкий в передней части, а контакт со вторым эпипластроном более протяженный (Moody, 1974, табл. I ).
Медиальные и латеральные части гио- и гипопластронов плохо различимы на томограммах. Центральная и латеральные фонтанели, если присутствовали, были маленькими. Медиальный контакт между гио-гипопластронами был, по крайней мере, редуцированным, судя по наличию зубцов медиального края гиопластронов. Часть гипопластрона медиальнее контакта с ксифипластроном примерно такой же длины, как остальная (передняя) часть гипопластрона. Расстояние между аксилярной и ингвинальной вырезками – около 56 мм; ширина гиопластронов – около 90 мм; пластральный индекс А (Zangerl, 1953) – около 62, что меньше, чем у Puppigerus camperi (70–88.5; Moody, 1974), но больше, чем у Tasbacka aldabergeni (48–53; Несов, 1987).
Ксифипластроны сохранились в виде отпечатков и небольших фрагментов кости. Они узкие, их медиальные края были параллельны средней линии и расположены близко друг к другу, как у Argillochelys antiqua, Erquelinnesia gosseleti, Euclastes wielandi и Tasbacka aldabergeni. Ксифипластроны имеют вытянутый антеролатеральный отросток, контактирующий с постеролатеральным краем гипопластрона. Длина ксифипластрона (максимальная) – 76 мм, ширина (максимальная) – 23 мм. Отношение ширины ксифипластрона к его длине – 0.3, что соответствует таковому у Tasbacka aldabergeni (около 0.3) и значительно ниже, чем у Puppigerus camperi (0.44–0.47).
Из костей осевого скелета сохранились VI–VIII шейные позвонки, I–IX cпинные позвонки, I и II крестцовые позвонки, I–IV хвостовые позвонки (рис. 3, г).
Шейные позвонки сохранились в прижизненном сочленении, их ряд повернут передней частью влево и немного вентрально; антеровентральная часть VI шейного позвонка обнажается на поверхности конкреции. Передняя сочленовная поверхность тела VI шейного позвонка округлой формы, вогнутая; задняя сочленовная поверхность шире, слегка выпуклая; постзигапофизы вытянуты назад, заостренные. Передняя сочленовная поверхность тела VII шейного позвонка слабовогнутая, а задняя – слегка выпуклая, вероятно, двойная; постзигапофизы вытянуты назад, заостренные. Тело VIII шейного позвонка короткое, передняя сочленовная поверхность слегка вогнутая, задняя – выпуклая; постеродорсальная часть невральной дуги с постзигапофизами, сильно вытянутыми назад, образующими площадку для контакта с выступом нухальной пластинки. Ширина сочленовных поверхностей тел шейных позвонков VII и VIII значительно превышает их высоту. Сохранившиеся шейные позвонки имеют широко расставленные презигапофизы и высокий вентральный киль.
Из сохранившихся спинных позвонков III–VII повреждены разломами в конкреции, VIII и IX – плохо различимы на томограммах; X – неразличим. Передняя сочленовная поверхность тела I спинного позвонка шире задней, ориентирована антеровентрально. Головка I спинного ребра сочленяется с фасеткой, расположенной с небольшим отступом от переднего края I спинного позвонка. Самый длинный спинной позвонок – III, от него в переднем и заднем направлении спинные позвонки уменьшаются в длину.
Из двух крестцовых позвонков I поврежден разломом в конкреции (его крестцовые ребра не различимы), а II меньше и сочленяется с широкими крестцовыми ребрами.
Хвостовые позвонки (I–IV) сохранились в прижизненном положении, их ряд повернут задней частью влево. На I хвостовом позвонке различимы близко посаженные вытянутые назад постзигапофизы, а на I и II – крупные поперечные отростки.
В разной степени сохранились все элементы переднего и заднего поясов конечностей (рис. 3, г): лопатки, коракоиды, лобковые, седалищные и подвздошные кости.
У правой лопатки отломаны и немного смещены медиальные части акромиона и лопаточного отростка, у левой лопатки – только лопаточного отростка. Лопаточный отросток и акромион тонкие. Лопаточный угол составляет 100°, что соответствует Tasbacka aldabergeni, у которого он варьирует от 95° до 110° (наши данные), равно как и Puppigerus camperi, у которого он составляет 100°–110° (Bardet et al., 2013).
Левый коракоид поврежден разломом в конкреции, его тело в значительной части длины узкое, постепенно расширяется дистально, как у Puppigerus camperi и Tasbacka aldabergeni (Moody, 1974, табл. V , фиг. E ; Несов, 1987, табл. VIII , фиг. 36, 37 ).
Контур передней части обеих лобковых костей на томограммах виден нечетко; левая кость, вероятно, перевернута на 180° вокруг своей оси. Лобковая кость медиолатерально непосредственно спереди от ацетабулярной части узкая, затем становится широкой и дорсовентрально уплощенной в передней части.
Левая седалищная кость смещена латеральной частью вперед. На правой седалищной кости сохранился длинный задний отросток, как у Puppigerus camperi и Tasbacka aldabergeni (Moody, 1974, рис. 12C ; Несов, 1987, табл. VIII , фиг. 28 ).
У левой подвздошной кости дорсальная пластина отломана и слегка смещена медиально. Дорсальная пластина подвздошной кости длинная, отграничена от передней части кости перегибом.
Из костей конечностей сохранились проксимальная часть правой плечевой кости, правая бедренная кость и проксимальная часть правой большой берцовой кости (рис. 3, г). Проксимальная часть правой плечевой кости сохранилась в виде полости, кость перевернута на 180° вокруг своей оси относительно нормального положения. Головка плечевой кости широкая дорсовентрально. Медиальный отросток вытянутый, как у экз. ZIN PH, № 2/283.
Правая бедренная кость имеет узкий и слабоизогнутый диафиз. Гребень между трохантерами развит слабо. Присутствует ли вырезка на гребне большого трохантера, не ясно. Мыщелки дистального эпифиза сильновыпуклые. Сравнение бедренной кости с таковой Puppigerus camperi затруднено, т.к. на ее изображении у этого вида (Moody, 1974, табл. 5 , фиг. H ) плохо различимы структуры проксимальной части кости. На бедренной кости Tasbacka aldabergeni из типовой коллекции видно, что гребень между трохантерами развит слабо (Несов, 1987, табл. VII , фиг. 23 б; наши данные).
Проксимальная часть правой большой берцовой кости в виде округлого элемента различима на томограмме у дистальной части правой бедренной кости.
Экз. ZIN PH, № 2/283 (рис. 4, а–д) представлен агломератом костей в фосфоритовой конкреции (длиной 19 см), включающем в себя хаотично расположенные симфиз зубной кости, две невральные, шесть костальных и две периферальные пластинки, а также плечевую кость. Из них на поверхности конкреции видны часть симфиза зубных костей и плечевая кость, остальные элементы полностью или почти полностью находятся в толще конкреции и видны на томограммах.
Симфиз зубных костей представлен фрагментом, включающим в себя переднюю и левую дорсолатеральную части (рис. 4, в, г). Симфиз треугольный, узкий и вытянут в длину, с заостренным, ориентированным вперед передним окончанием, что характерно для Puppigerus spp. и Tasbacka spp. Лабиальные гребни не развиты, хотя в задней части зубной кости имеется переходящее в венечный отросток поднятие, как у Puppigerus spp. и Tasbacka spp. (наши данные). Симфизный гребень отсутствует. Левый canalis alveolaris inferior виден близко к дорсальной поверхности на сломе кости сзади. На слайдах видно, что он проходит от этого слома почти до переднего окончания зубной кости. Также виден глубокий sulcus cartilaginis meckelii, что указывает на то, что левая часть кости сломана немного спереди от заднего края симфиза.
Две невральные пластинки неопределенного положения (рис. 4, б) имеют поврежденные края и плоскую наружную поверхность. Из шести костальных пластинок у трех сохранилось короткое свободное ребро (рис. 4, б). Одна из костальных пластинок выпуклая спереди, вероятно, правая I. Две правые периферальные пластинки – III и VI или VII (рис. 4, б). Правая III периферальная пластинка закруглена с латеральной стороны, наращивает высоту спереди назад. Правая VI или VII периферальная пластинка заострена с латеральной стороны, умеренно уплощена дорсовентрально. Обе пластинки имеют медиолатерально широкие дорсальные части.
Правая плечевая кость (рис. 4, д) экспонирована на поверхности конкреции со своей вентральной стороны. Латеральный отросток расположен близко к головке, округленный. Медиальный отросток вытянут. Мыщелки дистального эпифиза (радиальный и ульнарный) выпуклые. Эти элементы плечевой кости не отличаются от Puppigerus camperi и Tasbacka aldabergeni (личные наблюдения колл. ЦНИГР; фотографии IRSNB). Диафиз выглядит более узким, чем у P. camperi и T. aldabergeni, но это, по-видимому, связано с тем, что он сохранился не по всей ширине. Длина кости, с учетом отломанной части головки, составляла около 7 см. Это близко к значению для P. camperi (экз. IRSNB, № R 15; 74 мм; Moody, 1974) и среднему значению для T. aldabergeni (76 мм; Несов, 1987).
Экз. ZIN PH, № 3/283 (рис. 4, е–з) представлен агломератом из одной костальной и двух периферальных пластинок карапакса, видимых на поверхности фосфоритовой конкреции. II или IV костальная пластинка (рис. 4, е, ж) шириной 32 мм; медиальный и латеральный края отломаны; на дорсальной поверхности видна интерплевральная борозда; свободное ребро длинное. Две мостовые периферальные пластинки представлены передними или задними частями (рис. 4, з). Одна из них, вероятно, IV или V, высокая дорсовентрально, с закругленным латеральным краем. Вторая, видимо, VI, субтреугольная в сечении, с острым латеральным краем. Наружная поверхность костальной и периферальных пластинок гладкая, со слабовыраженными короткими желобками, разбросанными по поверхности.
В материалах (экземпляры в колл. ZIN PH, № 283) также имеются агломерат костей в фосфоритовой конкреции (экз. ZIN PH, № 4/283) и 12 отдельных костей и их фрагментов. В составе агломерата содержатся не менее двух костальных пластинок. Из других костей, которые удалось определить, три также являются костальными пластинками. Сохранность этих материалов не позволяет определить их точнее, чем Testudines indet., и поэтому их отнесение к T. aldabergeni следует считать условным.
Размеры. См. табл. 1.
Таблица 1.
Сравнение отдельных промеров у Tasbacka aldebergeni (экз. ZIN PH, №№ 1/283, 2/283 и 3/283). Обозначения: d – правый элемент; s – левый элемент
| Промеры (в мм) | экз. ZIN PH, № 1/283 |
экз. ZIN PH, № 2/283 |
экз. ZIN PH, № 3/283 |
|---|---|---|---|
| Медиолатеральная ширина костальных пластинок | 105 (III, d) | 109 (II–VI, s или d) | – |
| Длина латерального края тех же пластинок | 33 | 32 | 32 (II или IV, s или d) |
| Длина свободного ребра тех же пластинок | 26 (25%) | 17 (15%) | – |
| Медиальная длина периферальных пластинок | 35 (VII, s) | 35 (VI или VII, d) | – |
Распространение. Танет или ?ипр Южного Казахстана; палеоцен–ипр южной части Европейской России.
Материал. Ранее описанный: см. Несов (1987); Averianov (2002); Данилов и др. (2017). Новый: экз. ZIN PH, № 1/283, бόльшая часть посткрания одной особи в фосфоритовой конкреции; экз. ZIN PH, №№ 2–4/283, три агломерата костей в фосфоритовых конкрециях; другие экземпляры в колл. ZIN PH, № 283: три плохо сохранившиеся костальные пластинки и семь неопределенных плоских костей в фосфоритовых конкрециях, а также два фрагмента неопределенных плоских костей, найденных в продуктивном слое вне конкреций. Местонахождение Кудиновка, Миллеровский р-н, Ростовская обл., Россия; ипрский ярус, возможно, переотложены из палеоцена.
ОБСУЖДЕНИЕ
Определение черепах из Кудиновки. Экз. ZIN PH, №№ 1–3/283 отнесены к Chelonioidea на следующем основании: (1) у экз. ZIN PH, № 1/283 шовный контакт между костальными и периферальными пластинками отсутствует (по крайней мере, у части косто-периферальной серии), контакт между гио-гипопластронами редуцирован, ксифипластроны имеют вытянутый антеролатеральный отросток, контактирующий с постеролатеральным краем гипопластрона; (2) у экз. ZIN PH, № 2/283 плечевая кость с латеральным отростком, расположенным дистально по отношению к головке плечевой кости, но у проксимального окончания диафиза; (3) у экз. ZIN PH, № 3/283 шовный контакт между костальными и периферальными пластинками отсутствует, по крайней мере у части косто-периферальной серии (судя по длинному свободному ребру) (Evers et al., 2019).
Кайнозойские Chelonioidea представлены только Dermochelyidae и Cheloniidae. У плечевой кости экз. ZIN PH, № 2/283 отсутствуют синапоморфии Dermochelyidae: латеральный отросток плечевой кости, расположенный в средней части диафиза; выраженный передний выступ латерального отростка плечевой кости (Evers et al., 2019). Поэтому мы отнесли этот и другие материалы из местонахождения Кудиновка к Cheloniidae.
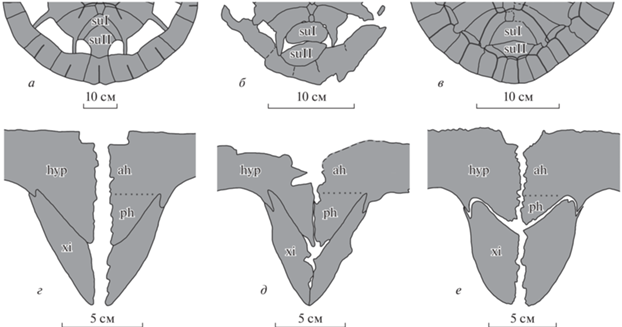
Экз. ZIN PH, № 1/283 имеет сочетание признаков, характерное среди раннепалеогеновых хелониид только для черепах рода Tasbacka: (1) II супрапигальная пластинка значительно шире пигальной пластинки (рис. 5, б); (2) ксифипластроны узкие и длинные, не образуют ксифипластральную фонтанель (рис. 5, д); (3) длина задней части гипопластрона медиальнее контакта с ксифипластроном примерно равна длине передней части гипопластрона (рис. 5, д). Первый из этих признаков не указан в диагнозе Tasbacka (Несов, 1987; Данилов и др., 2017), однако очень широкая II супрапигальная пластинка известна только у раннепалеогеновых хелониид с высокой степенью оссификации карапакса (Tasbacka и Puppigerus; рис. 5, а, в). Узкие и длинные ксифипластроны указаны в диагнозе рода Tasbacka (рис. 5, г; Несов, 1987). Похожие ксифипластроны наблюдаются у Argillochelys antiqua (sensu Lapparent de Broin et al., 2018), Erquelinnesia gosseleti и Euclastes wielandi. У Puppigerus camperi ксифипластроны более широкие (рис. 5, е), а у Eochelone они образуют ксифипластральную фонтанель. Последний признак (удлиненная задняя часть гипопластрона медиальнее контакта с ксифипластроном, примерно равная по длине с передней частью гипопластрона) также не указан в диагнозе Tasbacka (Несов, 1987; Данилов и др., 2017). Помимо Tasbacka (рис. 5, г), вытянутая постеромедиальная часть гипопластрона наблюдается у Argillochelys antiqua (sensu Lapparent de Broin et al., 2018), Erquelinnesia gosseleti и Euclastes wielandi. У Eochelone brabantica и Puppigerus camperi (рис. 5, е) часть гипопластрона медиальнее контакта с ксифипластроном значительно короче и составляет около трети длины гипопластрона.
Рис. 5.
Сравнение элементов панциря у раннепалеогеновых хелониид: а–в – задние части карапакса: а – Erquelinnesia gosseleti, экз. IRSNB, № 1563 (по: Zangerl, 1971, рис. 11 ); б, в – Tasbacka aldabergeni: б – экз. ZIN PH, № 1/283; в – экз. ЦНИГР, № 34/12175 (по: Несов, 1987, рис. 2а); г–е – задние части пластрона: г, д – Tasbacka aldabergeni: г – реконструкция по экз. ЦНИГР, №№ 19/12175 и 24/12175 (по: Несов, 1987, рис. 3, табл. VIII : фиг. 19, 24 ); д – ZIN PH, № 1/283; е – Puppigerus camperi, экз. IRSNB, № R 0073 (по: Evers et al., 2018, рис. S1.26B ). Обозначения: ah – передняя часть гипопластрона, ph – задняя часть гипопластрона медиальнее контакта с ксифипластроном, остальные как на рис. 3.
У экз. ZIN PH, № 2/283 симфиз зубных костей треугольный, вытянутый, заострен спереди. Этот морфотип известен у Puppigerus spp. и Tasbacka spp. У Euclastes acutirostris, судя по форме верхней челюсти, морфология зубных костей могла быть такой же. У Erquelinnesia (вероятный синоним Glossochelys и, отчасти, Puppigerus crassicostatus) и “Euclastes” douvillei похожие зубные кости, но у Erquelinnesia симфизная часть шире, а у “Euclastes” douvillei – короче. Присутствие в местонахождении Кудиновка представителя рода Tasbacka (экз. ZIN PH, № 1/283) делает слишком маловероятным одновременное присутствие сходных по морфологии и, видимо, занимавших ту же экологическую нишу Euclastes acutirostris или Puppigerus spp. Поэтому экз. ZIN PH, № 2/283 также отнесен к Tasbacka.
Морфология Tasbacka из Кудиновки отличается от таковой T. ouledabdounensis и T. ruhoffi. У экз. ZIN PH, № 1/283 все невральные пластинки плоские, что исключает принадлежность к T. ruhoffi, у которого невральные пластинки имеют киль. Экз. ZIN PH, № 2/283 имеет плоскую альвеолярную поверхность симфиза зубных костей, что исключает принадлежность к T. ouledabdounensis с хорошо выраженным симфизным гребнем. С другой стороны, по указанным признакам Tasbacka из Кудиновки cоответствует по морфологии T. aldabergeni и, таким образом, по-видимому, относится к этому виду. Сравнение с T. danica не проводилось, т.к. этот вид нуждается в переописании (см. ниже) и не содержит общих с материалом из Кудиновки, специфичных на уровне вида, анатомических элементов. Сравнение с T. salisburgensis не проводилось, т.к. отнесение данного вида к роду Tasbacka (Karl, Madsen, 2012) недостаточно обосновано из-за отсутствия диагностичных черт этого рода. Вместе с тем, имеются также некоторые отличия экз. ZIN PH, № 1/283 от T. aldabergeni. (1) У экз. ZIN PH, № 1/283 I супрапигальная пластинка уже II, тогда как у Tasbacka aldabergeni I супрапигальная пластинка, наоборот, шире II. Однако, во-первых, усеченные латеральные части I супрапигальной пластинки экз. ZIN PH, № 1/283 указывают на ее незавершенную оссификацию, при завершении которой I супрапигальная пластинка должна быть примерно равна по ширине II. Во-вторых, мы допускаем, что у Tasbacka может быть некоторое варьирование взаимных пропорций супрапигальных пластинок, как, например, у Chelonia mydas и Toxochelys moorevillensis (Zangerl, 1953, рис. 75 , табл. 23 ; 1958, рис. 22 ). Поэтому для разграничения Puppigerus и Tasbacka относительная ширина супрапигальных пластинок в настоящей работе не применяется. (2) Более длинные, чем у T. aldabergeni, свободные ребра экз. ZIN PH, № 1/283, очевидно, связаны с более молодым индивидуальным возрастом черепахи, т.к. у экз. ZIN PH, № 2/283 свободные ребра более короткие. (3) Пластральный индекс морских черепах (отношение расстояния между аксилярной и ингвинальной вырезками к половине ширины пластрона; Zangerl, 1953) подвержен возрастной изменчивости – у взрослых особей он ниже (Zangerl, 1971, с. 20–21). Поэтому пластральный индекс экз. ZIN PH, № 1/283 (62), слишком высокий для классических T. aldabergeni (48–53), может объясняться его онтогенетической незрелостью. В то же время, значение пластрального индекса у экз. ZIN PH, № 1/283 ниже, чем у Puppigerus camperi (70–88.5), что свидетельствует о невозможности принадлежности экз. ZIN PH, № 1/283 к этому виду.
Экз. ZIN PH, №№ 1/283, 2/283 и 3/283 принадлежали особям почти одинакового размера, но у экз. ZIN PH, № 1/283 и 3/283 длиннее свободные ребра, что указывает на незавершенный индивидуальный рост (табл. 1). Рост панциря T. aldabergeni из Кудиновки останавливался примерно при длине 30–35 см, что соответствует размеру зрелых особей T. aldabergeni из типового местонахождения (30–39 см).
Остальные материалы черепах из Кудиновки вполне могут относиться к T. aldabergeni, но слишком фрагментарны для точного определения.
Проблема раннеипрских хелониид с мозаикой черт Puppigerus и Tasbacka. Один из видов Tasbacka – T. danica – описан по ювенильной особи из нижнеипрских отложений формации Фур (местонахождение Эйерслев, Дания; верхи NP 10; Karl, Madsen, 2012). Череп крупной особи (экз. MHM, № K1), отнесенной к этому виду, обладает почти прямым нижним краем верхнечелюстных костей при виде сбоку (Karl, Madsen, 2012). Черепа хелониид с такой же морфологией верхнечелюстных костей описаны из ипра (вероятно, нижнего ипра) бассейна Уле Абдун (Марокко; экз. MAE, № 2005.1.1; как Puppigerus camperi; Tong et al., 2012) и нижнего ипра формации “Лондонские глины” (местонахождение Уолтон-на-Назе, Англия; верхи NP 10 (?) – низы NP 11; экз. NHMUK, № R14375; как ?Puppigerus camperi; Moody et al., 2015). Все эти материалы, вероятно, немного древнее древнейших P. camperi из верхов формации “Лондонские глины” (местонахождение о. Шеппи, Англия; NP 12; Owen, Bell, 1849).
Почти прямой нижний край верхнечелюстных костей, характерный для указанных раннеипрских экземпляров (экз. MHM, № K1; экз. MAE, № 2005.1.1; экз. NHMUK, № R14375), является одним из признаков отличий Tasbacka от Puppigerus, указанных в диагнозе первого рода (Несов, 1987). В то же время, передний край хоан у этих экземпляров находится спереди от fossa temporalis inferior, как у Puppigerus, тогда как у Tasbacka передний край хоан находится позади переднего края fossa temporalis inferior (см. исправленный краниальный диагноз Tasbacka; Tong, Hirayama, 2002). Мы согласны с тем, что эти экземпляры относятся к самостоятельному виду (pro Karl, Madsen, 2012) с мозаикой черт обоих родов, хотя этот вид требует переописания. К сожалению, у взрослых особей данного вида не описан пластрон, по которому его можно было бы сравнить с Tasbacka, в т. ч. из Кудиновки.
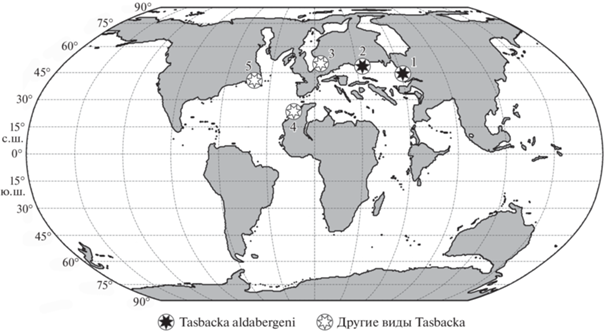
Таким образом, новые материалы по Tasbacka aldabergeni из местонахождения Кудиновка (Ростовская обл., Россия) расширяют представления о географическом распространении этого вида, известного ранее только из танета–ипра местонахождения Джилга 1 Южного Казахстана, и рода в целом, известного ранее, помимо танета–ипра Казахстана, также из палеоцена местонахождений Марокко, США и, возможно, ипра Дании (рис. 6). Материал по T. aldabergeni из Кудиновки несколько отличается от типового материала по этому виду, но эти отличия вполне укладываются в рамки внутривидовой изменчивости. Экз. ZIN PH, № 1/283 из Кудиновки представляет собой наиболее полный посткраний одной особи рода Tasbacka, который в дальнейшем может использоваться для сравнения. Материалы по T. aldabergeni из типового местонахождения Джилга 1а в сумме анатомически более полные, но представлены меньшими частями скелета или отдельными костями и принадлежат разным особям (Несов, 1987). Некоторые элементы позвоночника (VI шейный, спинные и крестцовые позвонки) T. aldabergeni описаны впервые по материалу из местонахождения Кудиновка. Сам род Tasbacka требует ревизии, в т.ч. с дополнительным исследованием T. danica из нижнего ипра Дании и похожих материалов из нижнего ипра Англии и, предположительно, нижнего ипра Марокко (Karl, Madsen, 2012; Tong et al., 2012; Moody et al., 2015).
Рис. 6.
Местонахождения черепах рода Tasbacka на палеогеографической картосхеме мира рубежа танетского и ипрского веков палеогена (по: Carmichael et al., 2017, рис. 1). Обозначения: 1 – Джилга 1, Туркестанская обл., Казахстан, танетский или ипрский ярус (T. aldabergeni); 2 – Кудиновка, Ростовская обл., Россия, палеоценовый отдел или ипрский ярус (T. aldabergeni); 3 – Эйерслев Моклэйпит, формация Фур, Дания, ипрский ярус (T. danica); 4 – бассейн Уле Абдун, Марокко, датский ярус (T. ouledabdounensis); 5 – Пискатауэй Крик, формация Акия, Мэрилэнд, США, танетский ярус (T. ruhoffi).
Список литературы
Аверьянов A.O. Новая морская черепаха (Testudines, Cheloniidae) из среднего эоцена Узбекистана // Палеонтол. журн. 2005. № 6. P. 73–80.
Беницкий К.С., Звонок Е.А. Местонахождения палеогеновых рептилий Миллеровского района (Ростовская область Российской Федерации) // География – от теории к практике: матер. ІІІ Междунар. науч.-практ. конф., приуроченной к 100-летию основания Луганского гос. пед. ун-та. Луганск: Книта, 2020. С. 98–104.
Геологическая карта СССР. Серия Донбасская. Лист М-37-XXIX. Объяснительная записка / Ред. А.М. Розентулер. М.: Гос. науч.-тех. изд-во лит-ры по геол. и охране недр СССР, 1963. 64 с.
Данилов И.Г., Сыромятникова Е.В., Суханов В.Б. Подкласс Testudinata // Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Часть 4 / Ред. А.В. Лопатин, Н.В. Зеленков. М.: ГЕОС, 2017. С. 27–395.
Железко В.И., Козлов В.А. Эласмобранхии и биостратиграфия палеогена Зауралья и Средней Азии // Материалы по стратиграфии и палеонтологии Урала. Вып. 3. Екатеринбург: УрО РАН, 1999. 324 с.
Звонок Е.А., Данилов И.Г. Палеогеновые черепахи Крыма // Палеонтол. журн. 2019. № 1. С. 61–72.
Звонок Е.А., Данилов И.Г. Среднеэоценовые сухопутные черепахи (Testudinidae) местонахождения Иково (Луганская область, Украина) // Биогеография и эволюционные процессы. Матер. LXVI сессии Палеонтол. об-ва. СПб., 2020. С. 241–243.
Звонок Е.А., Данилов И.Г., Сыромятникова Е.В., Удовиченко Н.И. Остатки морских черепах из местонахождения Иково (Луганская обл., Украина; средний эоцен) // Палеонтол. журн. 2013. № 6. С. 47–58.
Звонок Е.А., Сыромятникова Е.В., Данилов И.Г., Банников А.Ф. Морская черепаха (Cheloniidae) из среднего эоцена Северного Кавказа // Палеонтол. журн. 2019. № 5. С. 90–99.
Несов Л.А. О морских черепахах палеогена Южного Казахстана и филогенетических соотношениях Toxochelyidae и Cheloniidae // Палеонтол. журн. 1987. № 4. С. 76–87.
Несов Л.А., Ярков А.А. Новые птицы мела–палеогена СССР и некоторые замечания по истории возникновения и эволюции класса // Тр. Зоол. ин-та АН СССР. 1989. Т. 197. С. 78–97.
Суханов В.Б. Подкласс Testudinata // Основы палеонтологии. Земноводные, пресмыкающиеся и птицы / Ред. А.К. Рождественский, Л.П. Татаринов. М.: Наука, 1964. С. 354–438.
Averianov A.O. Review of Mesozoic and Cenozoic sea turtles from the former USSR // Russ. J. Herpetol. 2002. № 9(2). P. 137–154.
Bardet N., Jalil N.E., de Lapparent de Broin F. et al. Giant chelonioid turtle from the Late Cretaceous of Morocco with a suction feeding apparatus unique among tetrapods // PLOS One. 2013. V. 8(7). e63586. https://doi.org/10.1371/journal.pone.0063586
Bergounioux F.-M. Les Cheloniens fossiles de Gafsa // Notes Mém. Serv. Géol. Maroc. 1952. № 92. P. 377–396.
Christensen W.K. A Review of the Upper Campanian and Maastrichtian belemnite biostratigraphy of Europe // Cret. Res. 1996. V. 17. P. 751–766.
Carmichael M.J., Inglis G.N., Badger M.P.S. et al. Hydrological and associated biogeochemical consequences of rapid global warming during the Paleocene-Eocene Thermal Maximum // Glob. Planet. Change. 2017. V. 157. P. 114–138.
Evers S.W., Benson R.B.J. A new phylogenetic hypothesis of turtles with implications for the timing and number of evolutionary transitions to marine lifestyles in the group // Palaeontology. 2019. V. 62. № 1. P. 93–134.
Jalil N.-E., de Lapparent de Broin F., Bardet N. et al. Euclastes acutirostris, a new species of littoral turtle (Cryptodira, Cheloniidae) from the Palaeocene phosphates of Morocco (Oulad Abdoun Basin, Danian-Thanetian) // C. R. Palevol. 2009. V. 8. № 5. P. 447–459.
Karl H.-V., Madsen H. Tasbacka danica n. sp., a new Eocene marine turtle of Denmark (Testudines: Chelonioidea) // Studia Palaeocheloniologica. 2012. № 4. P. 193–204.
Lapparent de Broin F. de, Murelaga X., Farrés F., Altimiras J. An exceptional cheloniid turtle, Osonachelus decorata nov. gen., nov. sp., from the Eocene (Bartonian) of Catalonia (Spain) // Geobios. 2014. V.47. № 3. P. 111–132.
Lapparent de Broin F. de, Murelaga X., Pérez-García A. et al. The turtles from the Upper Eocene, Osona County (Ebro Basin, Catalonia, Spain): new material and its faunistic and environmental context // Fossil Record. 2018. V. 21. P. 237–284.
Lydekker R. Part III. The Order Chelonia // Catalogue of the fossil Reptilia and Amphibia in the British Museum (Natural History). L.: Brit. Mus. (Natur. Hist.), 1889. 239 p.
Moody R.T.J. The taxonomy and morphology of Puppigerus camperi (Gray), an Eocene sea turtle from northern Europe // Bull. Brit. Mus. (Natur. Hist.) Geol. 1974. V. 25. P. 153–186.
Moody R.T.J., Chapman S.D., King C. A new specimen of Puppigerus from the London Clay of Walton on the Naze, Essex, United Kingdom and the speciation and palaeo-distribution of marine turtles during the Eocene // PeerJ Preprints. 2015. № 3. e892v1. https://doi.org/10.7287/peerj.preprints.892v1
Myers T.S., Polcyn M.J., Mateus O. et al. A new durophagous stem cheloniid turtle from the Lower Paleocene of Cabinda, Angola // Pap. Palaeontol. 2018. V. 4. Pt. 2. P. 161–176.
Owen R., Bell A. Reptilia of the Tertiary beds. Part 1. Chelonia // Monography of the Fossil Reptilia of the London Clay. L.: Palaeontogr. Soc., 1849. P. 1–76.
Tong H., Hirayama R. A new species of Tasbacka (Testudines: Cryptodira: Cheloniidae) from the Paleocene of the Ouled Abdoun phosphate basin, Morocco // N. Jb. Geol. Paläontol. Mh. 2002. № 5. P. 277–294.
Tong H., Hirayama R., Tabouelle J. Puppigerus camperi (Testudines: Cryptodira: Cheloniidae) from the Ypresian (Early Eocene) of Ouled Abdoun basin, Morocco // Bull. Soc. Géol. Fr. 2012. V. 183. № 6. P. 635–640.
Ullmann P., Carr E. Catapleura Cope, 1870 is Euclastes Cope, 1867 (Testudines: Pan-Cheloniidae): synonymy revealed by a new specimen from New Jersey // J. Syst. Palaeontol. 2021. V. 19. № 7. P. 491–517.
Weems R.E. Paleocene turtles from the Aquia and Brightseat Formations, with a discussion of their bearing on sea turtle evolution and phylogeny // Proc. Biol. Soc. Wash. 1988. V. 101. № 1. P. 109–145.
Weems R.E. Reptile remains from the Fisher/Sullivan Site // Early Eocene Vertebrates and Plants from the Fisher/ Sullivan Site (Nanjemoy Formation) Stafford County, Virginia / Eds. R.E. Weems, G.J. Grimsley. Rep. Va. Div. Min. Res. Pub. 1999. № 152. P. 101–121.
Weems R.E. Paleogene chelonians from Maryland and Virginia // PaleoBios. 2014. V. 31. № 1. https://doi.org/10.5070/P9311022744
Zangerl R. The vertebrate fauna of the Selma Formation of Alabama: Pt IV. The turtles of the family Toxochelyidae // Fieldiana Geol. 1953. V. 3. № 4. P. 138–276.
Zangerl R. Die oligocänen Meerschildkröten von Glarus // Schweiz. Paläontol. Abh. 1958. Bd 73. S. 1–56.
Zangerl R. Two toxochelyid sea turtles from the Landenian sands of Erquelinnes (Hainaut) of Belgium // Bull. Inst. Roy. Sci. Natur. Belg. 1971. V. 169. P. 1–32.
Zvonok E.A., Danilov I.G. A revision of fossil turtles from the Kiev Clays (Ukraine, Middle Eocene) with comments on the history of the collection of fossil vertebrates of A.S. Rogovich // Proc. Zool. Inst. Russ. Acad. Sci. 2017. V. 321. № 4. P. 485–516.
Zvonok E.A., Danilov I.G. Paleogene turtles of Eastern Europe: new findings and reinterpretation of the previous materials // Turtle Evolution Symposium / Eds. R. Hirayama et al. Tübingen: Scidinge Hall Verlag, 2018. P. 93–97.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал