

Почвоведение, 2019, № 6, стр. 711-721
Характеристика сообществ микроартропод постагрогенных и естественных тундровых почв европейского северо-востока России
А. А. Таскаева 1, *, Е. А. Мандрик 2, Т. Н. Конакова 1, А. А. Кудрин 1
1 Институт биологии Коми НЦ УрО РАН
167928 Сыктывкар, ул. Коммунистическая, 28, Россия
2 Сыктыварский государственный университет им. П. Сорокина
167000 Сыктывкар, Октябрьский пр-т, 55, Россия
* E-mail: taskaeva@ib.komisc.ru
Поступила в редакцию 11.09.2017
После доработки 04.10.2018
Принята к публикации 28.11.2018
Аннотация
Дана характеристика сообществ микроартропод (панцирных, гамазовых клещей, коллембол) в постагрогенных и естественных тундровых почвах европейского северо-востока России. Показано, что в постагрогенных почвах, сформированных на месте возделывания многолетних и однолетних культур, доминирующей группой среди микроартропод являются коллемболы, а в тундровых – панцирные клещи. Плотность населения гамазовых клещей не претерпевает существенных изменений. В исследованных участках зарегистрировано 58 видов коллембол, 33 из которых отмечено в постагрогенных почвах и 46 – в тундровых. Сравнение видового состава коллембол рассмотренных участков свидетельствует о существенном отличии в структуре комплексов доминирующих видов, однако резко специализированной фауны в постагрогенных почвах не выявлено. Исследования не обнаружили связи между почвенными параметрами и численностью различных групп микроартропод, но отмечено их разделение в зависимости от типа исследованных почв.
ВВЕДЕНИЕ
Тундровые экосистемы формируются в суровых природно-климатических условиях. По сравнению с таежными биомами они характеризуются низким видовым разнообразием, продуктивностью и являются восприимчивыми к нарушениям, особенно антропогенного происхождения [33, 48]. С развитием угольной промышленности на Крайнем Севере (Республика Коми, Воркутинский район) и необходимостью обеспечения населения местной продукцией молочного животноводства в конце 50-х гг. прошлого столетия началось активное сельскохозяйственное освоение тундровых ландшафтов, не имеющее аналогов в России и за рубежом [23, 24]. С учетом специфики климатических условий и тундровых почв была разработана географически адаптированная система освоения тундровых земель. Многочисленные исследования показали, что оптимальным для тундровых условий является создание многолетних агроценозов из специально подобранных местных видов многолетних трав – лисохвоста лугового (Alopecurus pratensis) и мятлика лугового (Poa pratensis) [16, 24]. С 1958 г. Коми филиал АН СССР проводил опытную разработку приемов создания в тундре луговых травостоев длительного пользования на производственных площадях (1.5 га). Работами руководил И.С. Хантимер, который показал, что для сельскохозяйственного освоения пригодны 70–80% площади Воркутинской тундры. Это в первую очередь пространства преобладающие на водоразделах ерниковой тундры [24]. Длительные наблюдения за посевами местных многолетних трав выявили, что на 8–10 год формируется луговое сообщество с характерными чертами луговой почвы, не требующее применения сложных приемов агротехники – достаточно скашивания и умеренной подкормки. Результаты исследований первого десятилетия позволили разработать метод залужения, как оптимальный для Крайнего Севера способ эффективного использования малопродуктивных почв тундры [23]. После прекращения в конце 90-х гг. прошлого столетия их сельскохозяйственного использования участки пашни, где высевали однолетние и многолетние культуры, стали удобным объектом для изучения постагрогенной трансформации пахотных угодий в тундровой зоне [28].
В настоящее время на территории России большое количество пахотных земель выведено из сельскохозяйственного использования [21, 22]. На заброшенных землях возникают новые, постагрогенные экосистемы, характеризующиеся другим составом и структурой растительности. Изучению постагрогенных экосистем посвящены многочисленные исследования, но в большинстве работ особое внимание уделяется химическим свойствам почвы, оценке изменений запасов углерода, микробоценозам [6, 7, 17, 21, 42, 57]. Однако смена растительности играет ведущую роль не только в трансформации почв, но и в изменении сообществ почвенных беспозвоночных [55], являющихся важным функциональным компонентом как естественных, так и антропогенных экосистем. К настоящему времени достоверно установлено большое значение в почвообразовательном процессе микроартропод, среди которых наиболее заметную роль играют клещи и ногохвостки. Важность знания закономерностей их распределения в агроценозах была продемонстрирована рядом авторов [5, 11, 26, 39, 52]. Сравнивая особенности животного населения природных биогеоценозов и агроценозов, в качестве ведущего фактора отбора М.С. Гиляров выдвигал разнообразие растительного покрова. Основное отличие агроценоза от природного биогеоценоза он усматривал в замене исторически сложившегося многовидового растительного сообщества одной культурой, что ведет к упрощению базового ресурса цепей питания и энергопотоков [4]. Но при этом даже в монокультурных агроценозах формируется закономерная структура сообществ, повторяющаяся в сходных условиях [19]. Вместе с тем состояние почвенной микрофауны является важной характеристикой для оценки устойчивости наземных экосистем в тундровой зоне. В процессе хозяйственного использования территории в той или иной степени изменяется весь комплекс животных, населяющих данный район, что дает основания говорить о постепенной синантропизации фауны на обширных пространствах [9]. Почвенные микроартроподы, являющиеся наряду с микрофлорой обязательным звеном в цепи биологического круговорота веществ, изучены в этом плане недостаточно.
Цель работы – дать общую характеристику сообществ почвенных микроартропод постагрогенных экосистем, сформированных на месте возделывания многолетних и однолетних культур в тундровой зоне европейского северо-востока.
ОБЪЕКТЫ И МЕТОДЫ
Полевые методы. Исследования проведены в постагрогенных экосистемах, расположенных в подзоне южной кустарниковой тундры около промышленного центра г. Воркута. Пробы отобрали на двух парах участков: в постагрогенных экосистемах и в тундре (контроль). Участки характеризуются отсутствием многолетней мерзлоты до глубины 2 м от поверхности. Постагрогенные и контрольные участки находятся на удалении друг от друга, что обусловлено расположением первых на территории бывших совхозов (в 2 км от города), а вторых – на целинных участках тундры (в 10–15 км от города). Контрольные участки подобрали таким образом, чтобы они были аналогичны бывшим агроценозам как по исходному типу тундры, так и по положению в рельефе. Описания конкретных участков приводятся ниже.
Участок 1. Многолетний сеяный луг (67°31′ N, 64°03′ E), располагающийся на надпойменной террасе, в настоящее время представляет собой злаково-разнотравный луг с доминированием мятлика лугового (Poa pratensis), лисохвоста лугового (Alopecurus pratensis) и щучки дернистой (Deschampsia cespitosa). Краткое описание технологии создания и история функционирования многолетней агроэкосистемы даны в работе Ковалевой с соавт. [7]. Следует отметить, что помимо обработки почвы путем дискования, обязательным приемом освоения целины в тундре является известкование, проводимое с учетом кислотности почв, а также внесение минеральных удобрений [24]. В начале 1990-х гг. на сеяный луг стали сбрасывать в недозированных количествах навоз со скотных дворов, что вызвало резкое увеличение количества азота, калия и, особенно, фосфора, содержание которого в первое время возросло до 0.16 [14], а в 2013 г. составляло 0.8 моль/кг почвы [7]. Высокое содержание Cорг в освоенном слое сохраняется уже много лет. С 1998 г. внесение удобрений и уборка урожая прекратились. Почва – дерново-луговая (Mollic Stagnosol (Siltic, Gelic)), поверхность почвы покрыта уплотненным слоем слаборазложившегося растительного материала толщиной 2–3 см.
Участок 2. Разнотравно-злаковое сообщество (67°32′ N, 64°02′ E) расположено в пределах правобережной пойменной террасы р. Воркута. В травостое доминирующее положение занимают кострец безостый (Bromopus inermis) и виды разнотравья: вероника длиннолистная (Veronica longifolia), толстореберник альпийский (Pachipleurum alpinum), дудник лекарственный (Angelica archanlelica) и ряд других, приуроченных именно к пойме [6]. Данный участок был включен в сельскохозяйственное производство в 50-х годах прошлого столетия. На этом участке в течение 40 лет возделывали преимущественно однолетние культуры – овес (Avena sativa L.) и овсяно-гороховую смесь (Avena sativa + Pisum sativum L.) [23, 24]. Его хозяйственное использование в качестве пашни было прекращено в 2001 г. [14]. В настоящее время под этим сообществом формируется аллювиальная гумусовая глееватая постагрогенная почва (Gleyic Fluvisol (Arenic, Humic)).
Участок 3. Мелкоерниковая кустарничково-моховая тундра (67°25′ N, 63°50′ E) находится в долине р. Воркута в пределах второй надпойменной террасы, полого наклонной в сторону русла реки. Микрорельеф умеренно выражен, представлен, в основном, моховыми кочками вокруг кустов, местами встречаются пятна вымораживания. Кустарниковый ярус образуют Betula nana при участии Salix lanata и Salix phylicifolia, его сомкнутость 0.6–0.8. Кустарничковый ярус разреженный, состоит преимущественно из голубики (Vaccinium uliginosum), брусники (Vaccinium vitis-idaea), морошки (Rubus chamaemorus), вороники (Empetrum hermafroditum). Травянистые виды немногочисленны: Carex arctisibirica, Chamaenerion angustifoluim, Solidago virgaurea, Petasites frigidus, Pedicularis lapponica. Моховой покров хорошо развит, практически сплошной, состоит из Hylocomium proliferum, отмечены также политриховые мхи. Кустистые и листоватые лишайники встречаются преимущественно небольшими пятнами на бугорках. Почва – глеезем криометаморфический (Folic Stagnosol (Gelic)).
Участок 4. Кустарничковая мохово-лишайниковая тундра (67°26′03″ N, 63°49′35″ E) располагается на относительно выровненной первой надпойменной террасе р. Воркута. Микрорельеф представлен в виде небольших торфяных кочек и понижений. Проективное покрытие травянисто-кустарничкового яруса 40–50%. В сообществе обычно несколько содоминантов, в основном это кустарнички Vaccinium uliginosum, Arctous alpina, Vaccinium vitis-idaea, а также Betula nana, из трав обильны Carex arctisibirica, Eriophorum vaginatum, E. scheuchzeri, Calamagrostis neglecta. Мохово-лишайниковый покров сплошной. Лишайники располагаются рассеянно (занимают до 30%), в основном из родов Cetraria, Cladina и Cladonia. До 80% поверхности покрывают зеленые и сфагновые мхи. Почва – криометаморфическая грубогумусовая глееватая (Reductaquic Folic Cryosol).
На каждом участке отобрано по 10 проб из органогенного горизонта для анализа абиотических почвенных параметров 31 августа 2016 г. и по 20 проб площадью 100 см2 до глубины 7 см для учета почвенных микроартропод 7 июля и 31 августа 2016 г.
Абиотические параметры почвы. Аналитическую обработку образцов почв выполняли в экоаналитической лаборатории Института биологии Коми НЦ УрО РАН. Перед проведением анализов почву высушивали при температуре 50°С, затем измельчали при помощи мельницы Плаун – 40. Величину рН определили потенциометрически в соляной вытяжке (0.01 М СаСl2), массовую долю органического углерода (Собщ) и общего азота (Nобщ) – на CHNS-элементном анализаторе EA 1110 (Carlo E-rba), азот аммонийный – фотометрически на фотометре КФК-3 по методу ЦИНАО (ГОСТ 26489-85).
Почвенно-зоологические параметры. Для оценки численности микроартропод их экстракцию осуществляли по общепринятой методике с использованием эклекторов Берлезе–Тульгрена в 96%-ном спирте в течение 7–10 дней – времени, достаточного для достижения воздушно-сухого состояния почвы. Животных из проб отсортировывали под бинокулярным микроскопом и определяли до надсемейства в случае клещей и до вида в случае ногохвосток при помощи определителей [36, 37, 50] под микроскопом Leica DM 4000 B. Всего из 80 почвенных проб извлечено более 30 тыс. экземпляров микроартропод. Жизненные формы ногохвосток определяли по системе Стебаевой [18]. Классы доминирования коллембол выделяли по шкале Энгельмана [35].
Статистическая обработка данных. Для каждого параметра рассчитывали среднее на участок и стандартную ошибку. Для оценки значимости различий между выборками использовали непараметрический критерий Манна–Уитни при p < < 0.05. В качестве меры сходства сообществ коллембол выбран индекс Жаккара. Канонический анализ соответствий проводили на основе численности наиболее обильных видов коллембол (>2% от общей численности) и показателей почвенных параметров на исследованных участках. Статистическую обработку результатов приводили с помощью программ Statistica 6.0 и PAST 3.0.
РЕЗУЛЬТАТЫ
Абиотические параметры почвы. Значения абиотических характеристик в почвах варьируют незначительно (табл. 1). В тундрах отмечена кислая реакция среды, высокое содержание Собщ, Nобщ, а также широкое соотношение C : N. В постагрогенных экосистемах реакция среды практически нейтральная с низким значением C : N. Однако содержание азота и углерода на участках 1 и 2 отличалось практически в 3 раза. Самые низкие значения влажности отмечены на участке 1.
Таблица 1.
Абиотические параметры почвы и обилие почвенных микроартропод в постагрогенных и контрольных участках тундры (среднее и стандартная ошибка, n = 10 для почвенных параметров, n = 20 для микроартропод)
| Параметр | Постагрогенные экосистемы | Тундры | ||
|---|---|---|---|---|
| участок 1 | участок 2 | участок 3 | участок 4 | |
| Абиотические параметры почвы | ||||
| рН | 6.3 ± 0.08a | 6.3 ± 0.07a | 3.3 ± 0.02b | 3.4 ± 0.06b |
| Полевая влажность, % | 45.8 ± 2.3c | 55.4 ± 1.7b | 58.5 ± 3.1ab | 68.1 ± 2.8a |
| Содержание углерода, % | 22.0 ± 1.6b | 5.9 ± 0.3c | 32.1 ± 3.0a | 31.7 ± 1.8a |
| Содержание азота, % | 1.4 ± 0.09a | 0.4 ± 0.01c | 1.0 ± 0.08b | 0.8 ± 0.04b |
| NH4 из почвы, мг/кг | 56.0 ± 8.9a | 11.3 ± 0.6b | 29.0 ± 2.9a | 28.2 ± 2.69a |
| C : N | 19 ± 0.2c | 19 ± 0.2c | 38 ± 1.5b | 45 ± 1.6a |
| Микроартроподы | ||||
| Орибатиды, численность, экз./м2 | 5565 ± 1485b | 1140 ± 272c | 45530 ± 5544a | 41915 ± 6463a |
| Гамазовые клещи, численность, экз./м2 | 1735 ± 375b | 1960 ± 357b | 3605 ± 356a | 4635 ± 2187ab |
| Коллемболы, численность, экз./м2 | 20925 ± 2489b | 7335 ± 1972c | 32815 ± 3324a | 35940 ± 4772a |
| Число видов на пробу | 8.6 ± 0.5b | 6.8 ± 0.6c | 13.9 ± 0.5a | 14.9 ± 0.8a |
Микроартроподы. Общая численность микроартропод была в 4 раза ниже в постагрогенных экосистемах (в среднем 19.3 ± 3.5 тыс. экз./м2) по сравнению с тундровыми (82.2 ± 8.4 тыс. экз./м2). Если в первых преобладали коллемболы, то в целинной тундровой почве, напротив, клещи, особенно орибатиды. Биотоп (тундра или постагрогенная экосистема) оказывал влияние на численность панцирных клещей и разнообразие коллембол, а также на ряд почвенных параметров: кислотность, влажность, содержание углерода и азота (табл. 1).
Коллемболы. На исследованных участках зарегистрировано 58 видов коллембол, 33 из которых отмечено в постагрогенных экосистемах и 46 – в тундрах (табл. 2). Уровень сходства фаун коллембол постагрогенных и тундровых почв составил всего 34.5% − найдено только 20 общих видов. Среди всех биотопов наибольшими значениями индекса Шеннона (H = 2.54) характеризовался участок 4, где выявлены максимальные значения плотности населения, высокое видовое богатство, хорошая выравненность таксонов по численности. Доминантный комплекс ногохвосток постагрогенных экосистем и тундр различался (рис. 1). Только широко распространенный вид Folsomia quadrioculata преобладал во всех биотопах; на его долю приходилось от 20 до 25% от общей численности в биотопе. В постагрогенных почвах также доминантами являлись Desoria violacea (38.2%; участок 1), Desoria tshernovi (13.6%; участок 1), Desoria blufusata (23.3%; участок 2), причем на других участках он отсутствовал, Parisotoma notabilis (20.6%; участок 2), Protaphorura boedvarssoni (13%; участок 2). Уровня субдоминирования достигал Protaphorura subarctica (4.3%). В тундрах доминировали Tetracanthella wahlgreni (17.3%), и Protaphorura subarctica (13.3%). Четыре вида Pachyotoma miserabilis, Folsomia amplissima, Willemia anophthalma и Parisotoma notabilis оказались субдоминантами. В спектре жизненных форм на участках 1 и 3 преобладали поверхностно-обитающие виды, а на участках 2 и 4 – гемиэдафические (рис. 2).
Таблица 2.
Население коллембол исследуемых участков (средняя численность на м2 ± стандартная ошибка, n = 20)
| Вид | Постагрогенные экосистемы | Тундры | ||
|---|---|---|---|---|
| участок 1 | участок 2 | участок 3 | участок 4 | |
| Tullbergiidae | ||||
| Mesaphorura macrochaeta Rusek, 1976 | − | 15 ± 11 | − | − |
| Tullbergia simplex Gisin, 1958 | − | 5 ± 5 | − | − |
| Onychiuridae | ||||
| Hymenaphorura anatolii Pomorski, 2001 | 50 ± 41 | 5 ± 5 | − | − |
| Micraphorura absoloni (Börner, 1901) | 5 ± 5 | − | 45 ± 31 | − |
| Oligaphorura cf. groenlandica (Tullberg, 1876) | 40 ± 17 | − | − | − |
| Protaphorura bicampata (Gisin, 1956) | − | 260 ± 128 | − | − |
| Protaphorura boedvarssoni Pomorski, 1993 | 155 ± 55 | 950 ± 561 | 10 ± 7 | 430 ± 344 |
| Protaphorura borealis (Martynova, 1973) | 65 ± 35 | 5 ± 5 | − | − |
| Protaphorura jacutica (Martynova, 1976) | − | − | 5 ± 5 | 1840 ± 713 |
| Protaphorura subarctica (Martynova, 1976) | 1175 ± 375 | 35 ± 25 | 5185 ± 910 | 3980 ± 962 |
| Protaphorura subuliginata (Gisin, 1956) | 25 ± 25 | 15 ± 8 | − | 5 ± 5 |
| Protaphorura sp. 1 | − | 200 ± 190 | − | − |
| Protaphorura sp. 2 | − | 5 ± 5 | − | − |
| Supraphorura furcifera (Börner, 1901) | − | − | 115 ± 34 | 715 ± 311 |
| Uralophorura schilovi (Martynova, 1976) | − | − | 15 ± 15 | − |
| Hypogastruridae | ||||
| Ceratophysella denticulata (Bagnall, 1941) | 115 ± 34 | − | − | − |
| Schaefferia czernovi (Martynova, 1978) | − | − | − | 45 ± 28 |
| Willemia anophthalma Börner, 1901 | − | 5 ± 5 | 1765 ± 1348 | 2620 ± 623 |
| Willemia denisi Mills, 1932 | − | − | 5 ± 5 | − |
| Neanuridae | ||||
| Anurida beringii Fjellberg, 1985 | − | − | − | 35 ± 22 |
| Endonura reticulata (Axelson, 1905) | − | − | 105 ± 68 | 185 ± 79 |
| Friesea truncata Cassagnau, 1958 | 10 ± 7 | 15 ± 15 | 220 ± 55 | 10 ± 7 |
| Micranurida pygmaea Börner, 1901 | − | − | 175 ± 124 | 330 ± 85 |
| Neanura muscorum (Templeton, 1835) | − | − | 85 ± 60 | − |
| Pseudachorutes dubius Krausbauer, 1898 | − | − | − | 20 ± 14 |
| Pseudachorutes sibiricus Rusek, 1991 | − | − | 20 ± 15 | 5 ± 5 |
| Odontellidae | ||||
| Xenyllodes armatus Axelson, 1905 | − | − | 5 ± 5 | − |
| Isotomidae | ||||
| Desoria alaskensis (Fjellberg, 1978) | − | − | − | 80 ± 54 |
| Desoria blufusata (Fjellberg, 1978) | − | 1705 ± 1070 | − | − |
| Desoria hiemalis (Schött, 1893) | − | − | 50 ± 28 | − |
| Desoria neglecta (Schéffer, 1900) | 5 ± 5 | − | 35 ± 22 | 775 ± 335 |
| Desoria tshernovi (Martynova, 1974) | 2840 ± 1098 | 65 ± 33 | 200 ± 61 | 60 ± 41 |
| Desoria violacea (Tullberg, 1876) | 7980 ± 2436 | 20 ± 14 | 875 ± 471 | 635 ± 243 |
| Folsomia amplissima Potapov et Babenko, 2000 | 5 ± 5 | − | 2530 ± 832 | 1915 ± 442 |
| Folsomia bisetosa Gisin, 1953 | 140 ± 48 | 40 ± 17 | 515 ± 312 | − |
| Folsomia longidens Potapov et Babenko, 2000 | − | − | 30 ± 30 | 20 ± 15 |
| Folsomia manolachei Bagnall, 1939 | − | − | − | 65 ± 38 |
| Folsomia palaearctica Potapov et Babenko, 2000 | 460 ± 358 | 65 ± 25 | 5 ± 5 | − |
| Folsomia quadrioculata (Tullberg, 1871) | 4800 ± 1280 | 1685 ± 657 | 6490 ± 1591 | 8800 ± 2978 |
| Isotoma anglicana Lubbock, 1873 | − | − | − | 5 ± 5 |
| Isotoma gorodkovi (Martynova, 1970) | − | − | − | 175 ± 175 |
| Isotoma viridis Bourlet, 1839 | 10 ± 10 | 90 ± 26 | 310 ± 150 | 1085 ± 288 |
| Isotomiella minor (Schäffer, 1896) | − | 40 ± 22 | 1715 ± 638 | 510 ± 151 |
| Pachyotoma miserabilis Potapov, 2017 | − | − | 2610 ± 487 | 4025 ± 3330 |
| Parisotoma ekmani (Fjellberg, 1977) | − | − | 160 ± 86 | − |
| Parisotoma notabilis (Schäffer, 1896) | 1870 ± 459 | 1505 ± 252 | 1145 ± 188 | 1680 ± 542 |
| Parisotoma reducta (Rusek, 1984) | − | − | − | 1505 ± 379 |
| Tetracanthella wahlgreni Axelson, 1907 | − | − | 7975 ± 1532 | 3900 ± 541 |
| Tomoceridae | ||||
| Tomocerina minuta (Tullberg, 1876) | 10 ± 10 | 95 ± 60 | 240 ± 54 | 265 ± 121 |
| Entomobryidae | ||||
| Entomobrya nivalis (Linnaeus, 1758) | 135 ± 62 | 5 ± 5 | 10 ± 7 | 30 ± 30 |
| Lepidocyrtus lignorum (Fabricius, 1793) | − | 70 ± 28 | 35 ± 13 | 105 ± 35 |
| Lepidocyrtus violaceus (Geoffroy, 1762) | − | − | − | 30 ± 25 |
| Neelidae | ||||
| Megalothorax sp. 1 | − | − | 80 ± 44 | 45 ± 15 |
| Sminthurididae | ||||
| Sphaeridia pumilis (Krausbauer, 1898) | 35 ± 24 | 395 ± 224 | 10 ± 10 | − |
| Katiaanidae | ||||
| Sminthurinus aureus (Lubbock, 1862) | 960 ± 239 | 10 ± 10 | − | − |
| Sminthurinus niger (Lubbock, 1868) | 5 ± 5 | − | − | − |
| Arrhopalitidae | ||||
| Pygmarrhopalites principalis (Stach, 1945) | − | 5 ± 5 | 5 ± 5 | 10 ± 7 |
| Bourletiellidae | ||||
| Heterosminthurus claviger Gisin, 1958 | − | − | 10 ± 10 | − |
| Видовое богатство, S | 23 | 27 | 36 | 35 |
| Индекс разнообразия, Н | 1.79 | 2.07 | 2.28 | 2.54 |
| Выравненность, J | 0.57 | 0.62 | 0.63 | 0.71 |
Рис. 1.
Диаграмма канонического анализа (CCA) сообщества коллембол тундровых и постагрогенных экосистем. Стрелками указаны почвенные параметры: Вл – влажность почвы (%), N – общее содержание азота (%), С – общее содержание углерода (%), NH4 – содержание аммонийного азота (мг/кг), С : N – отношение углерода к азоту. Цифрами указаны номера исследованных участков (1 и 2 – постагрогенные, 3 и 4 – контрольная тундра). Черными точками указаны виды коллембол.
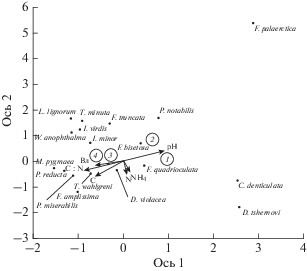
Рис. 2.
Соотношение жизненных форм коллембол в постагрогенной (участки 1 и 2) и контрольной тундровой почве (участки 3 и 4).

Связь параметров почвенной фауны с абиотическими факторами. Для оценки вклада почвенных параметров в объяснение особенностей распределения коллембол в исследованных тундровых и постагрогенных экосистемах был использован канонический анализ соответствий (CCA). На CCA диаграмме ось 1 определялась такими почвенными свойствами, как pH, влажность, C : N, общее содержание углерода; ось 2 – содержанием общего и аммонийного азота. Собственные значения (Eigenvalues) оси 1 и 2 равнялись 0.37 и 0.10 соответственно. Проведенный анализ указывает на четкое разделение сообществ коллембол в зависимости от типа исследованных участков (тундры или постагрогенные системы). Однако четкой связи между численностью отдельных видов микроартропод и исследованными почвенными параметрами не обнаружено.
ОБСУЖДЕНИЕ
Анализ, проведенный для четырех участков в пределах южной тундры, является пионерным исследованием, но он выявил ряд интересных закономерностей. Известно, что основной трофической группировкой луговых и тундровых почв являются сапрофаги – панцирные клещи и коллемболы, которые в исследованных постагрогенных экосистемах составили около 80–94% всех рассмотренных микроартропод. При этом соотношение их обилия на участках различалось. Следует отметить, что структура комплекса микроартропод участков 3 и 4 не совсем типична для тундровых экосистем. Плотность населения орибатид на этих участках не только не уступала численности населения коллембол, но и преобладала, что не соответствует коллембоидному облику тундровых сообществ [10]. Обилие панцирных клещей на участке 1 не превышало 20% всех микроартропод, а основная доля приходилась на ногохвосток (74%). Обилие хищных гамазовых клещей в комплексе микроартропод обычно мало колеблется и составляет около 7% [15]. В нашем исследовании обилие гамазид оказалось близким к этому значению, как в контрольных участках (4–6%), так и на участке 1 (6%). Участок 2 отличался по всем этим показателям: отмечена самая низкая численность микроартропод, при этом на долю орибатид приходилось около 11%, гамазид – 19% и доминирующей группой по-прежнему оставались коллемболы. На наш взгляд, такая перестройка структуры микроартропод на участке 2 связана с тем, что он располагается в пойме и регулярно затапливается паводковыми водами. Сообщества панцирных клещей реагируют на этот неблагоприятный фактор уменьшением общей численности и выпадением ряда видов, свойственных зональному сообществу [2].
В естественных биогеоценозах комплекс орибатид характеризуется более высокой видовой дифференцированностью и значительно меньшей динамичностью, чем коллемболы. Антропогенное воздействие на почву более резко и часто необратимо влияет именно на комплекс панцирных клещей. Например, они первыми исчезают при загрязнении почв продуктами нефтепромыслов и последними восстанавливаются [33, 46]. Уничтожение поверхностных горизонтов почвы, особенно отмерших растительных остатков, кардинально деформирует сообщество панцирных клещей, так как подавляющее большинство разнообразных видов, связанных с подстилкой, совершенно отсутствуют в агроценозах даже при близком соседстве целинных биотопов [27]. Согласно данным литературы, такая реакция орибатид обусловлена их низким уровнем метаболизма, медленным развитием и низкой плодовитостью [30]. Разбалансировка комплексов панцирных клещей отрицательно сказывается на его возможностях реагировать на изменения среды и делает трудно восстановимым после нарушения.
Плотность населения гамазовых клещей в агроценозах, как и в рассмотренных постагрогенных экосистемах, напротив, не претерпевала существенных изменений и оставалась относительно устойчивой в комплексе микроартропод. Это может быть обусловлено тем, что данная группа клещей включает семейства, которые по-разному реагируют на интенсивность обработки почвы, что имеет важное сельскохозяйственное значение и требует дальнейшего анализа влияющих факторов [40].
Наши исследования показали, что отличительной чертой населения комплекса микроартропод постагрогенных экосистем является доминирование коллембол, которое, по-видимому, связано с внесением органических и минеральных удобрений, что подтверждается и данными литературы [29, 32]. Несмотря на то, что численность коллембол по сравнению с таковой для тундровых экосистем была в 1.5–5 раз меньше, она соответствовала данным, полученных для агроэкосистем [1, 5, 43]. Сравнение видового состава ногохвосток рассмотренных участков обнаружило существенное отличие в структуре комплексов доминирующих видов, однако резко специализированной фауны на участках 1 и 2 не выявлено. Группировки, заселяющие постагрогенные экосистемы, в основном складывались из элементов фауны тундр, но на участке 1 присутствовали компостный (C. denticulata) и луговой (S. aureus) виды, а на участке 2 – виды, приуроченные к пойменным местообитаниям (D. blufusata, S. pumilis). Все рассмотренные участки сближает общий доминант F. quadrioculata, являющийся убиквистом. Определенный интерес представляет спектр жизненных форм коллембол. Их соотношение оказалось довольно схожим на участках 2 и 4, располагающихся в пойме, где преобладали гемиэдафические виды, что соответствует данным литературы [20]. Похожая картина, но с доминированием поверхностно обитающих видов ногохвосток отмечена на участках 1 и 3 на надпойменной террасе. Доля почвенных видов во всех исследованных нами биотопах низка, особенно в постагрогенных экосистемах, где она не превышала 22%. Из литературы [29, 45] известно, что разработка пашни (участки 1 и 2) вызывает уплотнение почвы, что приводит к сокращению жизненного пространства для эуэдафических видов коллембол.
В целом, для микроартропод агроценозов характерно обеднение видового состава, набора жизненных форм и общей численности по сравнению с естественными экосистемами [3]. Однако эта тенденция по-разному проявляется в разных систематических группах мелких почвенных беспозвоночных, а также в зависимости от конкретных почвенно-климатических условий [25, 34, 54]. Наблюдаемая средняя численность микроартропод на участке 1 была сопоставима с приводимой другими авторами средней плотностью для почв под многолетними травами и естественными луговыми биоценозами, располагающихся на водоразделе. А на участке 2 – с таковой в пахотных почвах и пойменных экосистем, а также близка к показателям более южных широт [1, 2, 5, 8, 15, 26, 51].
Исследования не выявили связи между почвенными параметрами и численностью различных групп микроартропод, но обнаружили их разделение в зависимости от типа исследованных участков (тундры или постагрогенные экосистемы). Нами показано, что на численность панцирных клещей и коллембол оказывал влияние тип биотопа, что подтверждается и данными литературы [47]. Связь между плотностью мезостигматических клещей и растительностью выражена слабо, что, по-видимому, обусловлено их биологией. Предполагается, что данные хищники являются оппортунистами со слабыми предпочтениями к типу местообитания, и их изменения тесно связаны с изменениями в сообществе панцирных клещей и коллембол, которые являются их потенциальной добычей. В то же время орибатиды и ногохвостки имеют сильные пищевые предпочтения к определенным видам растений и микроорганизмов, в особенности к грибам [44], что, возможно, и определяет их связь с типом растительности [47]. С биомассой грибов может быть связано разнообразие сообществ коллембол [55], которое также обнаружило зависимость от типа исследованных участков. Учитывая, что признаки освоения в почве сохраняются продолжительное время, в частности повышенное содержание питательных элементов и гумуса [21], то на видовой состав ногохвосток может оказывать влияние внесение органических и минеральных удобрений [5]. Кроме того, установлено влияние типа биотопа и на ряд почвенных параметров. Почти нейтральная реакция почвы, нехарактерная для целинных тундровых почв, является результатом многолетнего внесения известковых материалов в почву пахотного угодья, и даже спустя 15–20 лет после снятия агрорежима в постагрогенных экосистемах она сохранилась. Известно, что сообщества коллембол очень чувствительны к изменению рН почвы [49, 56]. Например, виды W. anophthalma, M. pygmaea, предпочитающие почвы с рН менее 4 [38], в постагрогенных экосистемах отсутствовали или являлись редкими. Напротив, обилие P. notabilis возрастало на участках 1 и 2 до 20% по сравнению с тундрами, где его доля в сообществе ногохвосток не превышала 4%. Высокие значения отношения C : N в почвах тундр свидетельствует о низкой обогащенности почвенного органического вещества азотом и слабой степени его разложения [13]. Однако на участке 1 отмечена высокая аккумуляция общего и аммонийного азота, а также фосфора [7] что, по-видимому, является следствием внесения неустановленных доз удобрений в начале 90-х годов. Высокие концентрации P приводят к менее разнообразному составу растительного покрова [41], что подтверждается данными литературы [12], а большие значения N не способствуют инициации процесса восстановления исходной экосистемы [41]. В настоящее время данный участок находится на этапе адаптации (устойчивое состояние фитоценоза в новых условиях) [12]. Восстановление сообществ микроартропод может длиться десятилетия или столетия, в зависимости от возможности рассеивания вида и близости возможных источников иммиграции более адаптированных видов, а также отсутствия новых изменений в землепользовании за это время [49].
ЗАКЛЮЧЕНИЕ
Сельскохозяйственное освоение тундры привело к смене типа растительности, свойств почвы, состава микробоценоза и, соответственно, сообществ микроартропод. Результаты исследований показали, что в постагрогенных почвах, сформированных на месте возделывания многолетних и однолетних культур в тундровой зоне европейского северо-востока, доминирующей группой среди микроартропод являлись коллемболы. Сообщества панцирных клещей, напротив, находились в угнетенном состоянии, хотя в тундровых почвах они преобладали. Плотность населения гамазовых клещей в рассмотренных участках не претерпевала существенных изменений. Сравнение видового состава коллембол исследованных почв свидетельствует о существенном отличии в структуре комплексов доминирующих видов (индекс Жаккара равен 33%), однако резко специализированной фауны на участках 1 и 2 не выявлено. В дерново-луговой и аллювиальной гумусовой постагрогенной почвах зарегистрировано 33 вида, разнообразие которых в основном складывалось из элементов фауны тундр. Исследования эдафических факторов не выявили связи с численностью различных групп микроартропод, но обнаружили их разделение в зависимости от типа исследованных почв (тундровые или постагрогенные).
Необходим дальнейший мониторинг изменений, так как таких прецедентов, как почти полувековое существование рукотворных луговых сообществ за Полярным кругом, насколько нам известно, не существует.
БЛАГОДАРНОСТЬ
Авторы признательны сотрудникам экоаналитической лаборатории за проведенные химические анализы и анонимному рецензенту за ценные замечания по рукописи статьи.
ФИНАНСИРОВАНИЕ РАБОТЫ
Работа выполнена в рамках госбюджетной темы НИР отдела экологии животных “Распространение, систематика и пространственная организация фауны и населения животных таежных и тундровых ландшафтов и экосистем европейского северо-востока России”, номер государственной регистрации АААА-А17-117112850235-2.
Список литературы
Алейникова М.М., Мартынова Е.Ф. Ландшафтно-экологический обзор фауны почвенных ногохвосток (Collembola) Среднего Поволжья // Pedobiologia. 1966. Bd. 6. № 1. P. 35–64.
Андриевский В.С., Якутин М.В., Шепелев А.И. Трансформация зоомикробиологического комплекса в процессе естественной эволюции пойменных почв в таежной зоне Западной Сибири // Евразиатский энтомол. журн. 2013. № 12(5). С. 425–430.
Гиляров М.С. Зоологический метод диагностики почв. М.: Наука, 1965. 275 с.
Гиляров М.С. Биогеоценология и агроценология. Пущино, 1980. 16 с.
Капин Г.Ю. Распределение коллембол в неудобренной и удобренной пахотной дерново-подзолистой почве // Фауна и экология ногохвосток (Collembola). М.: Наука, 1984. С. 179–186.
Ковалева В.А., Денева С.В., Панюков А.Н. Микробиологические показатели почвы постагрогенного биоценоза в тундровой зоне // Вестник Института биологии Коми НЦ УрО РАН. 2016. № 4(198). С. 2–9.
Ковалева В.А., Хабибуллина Ф.М., Арчегова И.Б., Панюков А.Н. Характеристика биоты постагрогенной экосистемы в тундровой зоне // Известия Коми научного центра УрО РАН. 2014. № 3(19). С. 70–74.
Колесников В.Б. Орибатиды как биоиндикаторы при мониторинге состояния малоиспользуемых сельскохозяйственных земель Воронежской области // Известия Самарского научного центра РАН. 2010. Т. 12. № 1(4). С. 1133–1138.
Криволуцкий Д.А., Казадаев А.А., Пономаренко А.В. Влияние хозяйственной деятельности человека на комплексы панцирных клещей // Вестник зоологии. 1977. № 6. С. 7–12.
Леонов В.Д. Панцирные клещи (Acari, Oribatida) тундровых почв Кольского п-ва. Автореф. дис. … канд. биол. наук. М., 2016. 26 с.
Миронов С.Ю. Население ногохвосток (Collembola) пахотных черноземов Курской области // Зоол. журн. 2006. Т. 85. № 11. С. 1321–1326.
Новаковский А.Б., Панюков А.Н. Анализ сукцессионной динамики сеянного луга при помощи системы жизненных стратегий Раменского-Грайма // Экология. 2018. № 2. С. 110–118.
Орлов Д.С. Гумусовые кислоты почв и общая теория гумификации. М.: Изд-во Моск. ун-та, 1990. 325 с.
Панюков А.Н. Однолетние агрофитоценозы в тундровой зоне – создание и трансформация // Аграрная наука Евро-Северо-Востока. 2009. № 4(15). С. 39–42.
Петрова-Никитина А.Д., Бутовский Р.О., Минор М.А. Влияние автодороги на комплекс микроартропод агроценоза. Сообщение 1. Структура комплекса // Агрохимия. 1994. № 10. С. 116–125.
Посттехногенные территории Севера. СПб.: Наука, 2002. 160 с.
Рыжова И.М., Ерохова А.А., Подвезенная М.А. Изменение запасов углерода в постагрогенных экосистемах в результате естественного восстановления лесов в Костромской области // Лесоведение. 2015. № 4. С. 307–317.
Стебаева С.К. Жизненные формы ногохвосток (Collembola) // Зоол. журн. 1970. Т. 49. Вып. 10. С. 1437–1455.
Стриганова Б.Р. Влияние эдафического фактора на формирование животного населения почв агроценозов // Зоол. журн. 2003. Т. 82. № 2. С. 178–187.
Таскаева А.А., Лаптева Е.М., Дегтева С.В. Изменение комплекса ногохвосток аллювиальных почв в процессе сукцессии растительности в пойменных ландшафтах таежной зоны // Вестник Поморского ун-та. Сер. Естественные и точные науки. 2005. № 2(8). С. 36–47.
Телеснина В.М., Ваганов И.Е., Климович Е.Ю., Чалая Т.А. Некоторые особенности биологического круговорота в постагрогенных экосистемах южной тайги и их влияние на химические свойства и биологическую активность почв // Вестник Моск. ун-та. Сер. 17, почвоведение. 2013. № 2. С. 43–51.
Титова В.И., Митянин И.О., Ветчинников А.А., Вершинина И.В. Оценка возможности использования сеяных трав для консервации деградированных пахотных земель и рекультивации техногенно нарушенных почв // Агрохимический вестник. 2011. № 2. С. 24–26.
Хантимер И.С. Залужение – основа обеспечения кормами молочного животноводства в тундре // Сообщества Крайнего Севера и человек. 1985. М.: Наука, С. 115–133.
Хантимер И.С. Сельскохозяйственное освоение тундры. Л.: Наука, 1974. 227 с.
Чернова Н.М. Экологические сукцессии при разложении растительных остатков. М.: Наука, 1977. 200 с.
Чернова Н.М. Комплексы микроартропод пахотных почв Подмосковья // Почвенные беспозвоночные Московской области. М.: Наука, 1982. С. 107–118.
Штирц А.Д., Задорожная Г.А., Кунах О.Н., Жуков А.В. Пространственная организация сообщества панцирных клещей (Acari: Oribatida) в почве сельскохозяйственного поля в условиях степной зоны Украины // Изв. Харьк. Энтомол. о-ва. 2013. Т. 21. № 1. С. 49–60.
Экологические основы управления продуктивностью агрофитоценозов восточноевропейской тундры. 1991. Л.: Наука, 152 с.
Bardgett R.D., Cook R. Functional aspects of soil animal diversity in agricultural grasslands // Appl. Soil Ecol. 1998. V. 10. P. 263–276.
Behan-Pelletier V. Oribatid mite biodiversity in agroecosystems: role for bioindication // Agric. Ecosyst. Environ. 1999. V. 74. P. 411–423.
Brussaard L., de Ruiter P.C., Brown G.G. Soil biodiversity for agricultural sustainability // Agriculture, ecosystems and environment. 2007. V. 121. P. 233–244. https://doi.org/10.1016/j.agee.2006.12.013
Cluzeau D.,Guernion M., Chaussod R., Martin-Laurent F., Villenave C., Cortet J., Ruiz-Camacho N., Pernin C., Mateille T., Philippot L., Bellido A., Rougé L., Arrouays D., Bispo A., Pérès G. Integration of biodiversity in soil quality monitoring: Baselines for microbial and soil fauna parameters for different land-use types // Europ. J. Soil Biol. 2012. V. 49. P. 63–72. https://doi.org/10.1016/ j.ejsobi.2011.11.003
Coulson S.J., Fjellberg A., Melekhina E.N., Taskaeva A.A., Lebedeva N.V., Belkina O.A., Seniczak S., Seniczak A., Gwiazdowicz D.J. Microarthropod communities of industrially disturbed or imported soils in the High Arctic; the abandoned coal mining town of Pyramiden, Svalbard // Biol. Conservat. 2015. V. 24. P. 1671–1690. https://doi.org/10.1007/s10531-015-0885-9
D’Hose T., Molendijk L., Van Vooren L., van den Berg W., Hoek H., Runia W., van Evert F., ten Berge H., Spiegel H., Sandèn T., Grignani C., Ruysschaert G. Responses of soil biota to non-inversion tillage and organic amendments: An analysis on European multiyear field experiments // Pedobiologia – J. Soil Ecol. 2018. V. 66. P. 18–28. https://doi.org/10.1016/j.pedobi.2017.12.003
Engelmann H.-D. Zur dominanz klassifizierung von Boden arthropoden // Pedobiologia. 1978. Bd. 18. S. 378–380.
Fjellberg A. The Collembola of Fennoscandia and Denmark // Fauna entomologica Scandinavica. 1998. V. 35. 184 p.
Fjellberg A. The Collembola of Fennoscandia and Denmark. Part II: Entomobryomorpha and Symphypleona // Fauna Entomologica Scandinavica. 2007. V. 42. Leiden: Brill. 264 p.
Hagvar S., Amrahamsen G. Colonization by Enchytraeidae, Collembola and Acari in sterile soil samples wih adjusted pH levels // Oikos. 1980. V. 34. P. 245–258.
Hendrickx F., Maelfait J.-P., van Wingerden W., Schweiger O., Speelmans M., Aviron S. et al. How landscape structure, land-use intensity and habitat diversity affect components of total arthropod diversity in agricultural landscapes // J. Appl. Ecol. 2007. V. 44. № 2. P. 340–351. https://doi.org/10.1111/j.1365-2664.2006.01270.x
Holland J.M. The environmental consequences of adopting conservation tillage in Europe: reviewing the evidence // Agric. Ecosyst. Environ. 2004. V. 103. P. 1–25. doi .https://doi.org/10.1016/j.agee.2003.12.018
Holtkamp R., Kardol P., van der Wal A., Dekker S.C., van der Putten W.H., de Ruiter P.C. Soil food web structure during ecosystem development after land abandonment // Appl. Soil Ecol. 2008. V. 39. P. 23–34. doi https://doi.org/10.1016/j.apsoil.2007.11.002
Hooker T.D., Compton J.E. Forest ecosystem carbon and nitrogen accumulation during the first century after agricultural abandonment // Ecol. Appl. 2003. V. 13. № 2. P. 99–313. https://doi.org/10.1890/1051-0761(2003)013[0299:FECANA]2.0.CO;2
Kovac L. Effects of soil type on collembolan communities in agroecosystems // Acta Zool. Fenn. 1994. № 195. P. 89–93.
Kurakov A.V., Davydova M.A., Byzov B.A. Microarthropods as regulators of the communities of microscopic fungi and biological activity in the litter of a mixed forest // Eurasian Soil Sci. 2006. V. 39. № 8. P. 838–847.
Larsen T., Schjønning P., Axelsen J. The impact of soil compaction on euedaphic Collembola // Appl. Soil Ecol. 2004. V. 26. P. 273–281. https://doi.org/10.1016/j.apsoil.2003.12.006
Melekhina E.N., Markarova M.Y., Shchemelinina T.N., Anchugova E.M., Kanev V.A. Secondary successions of biota in oil-polluted peat soil upon different biological remediation methods // Eurasian Soil Sci. 2015. V. 48. № 6. P. 643–653. https://doi.org/10.1134/S1064229315060071
Nielsen U.N., Osler G.H.R., Campbell C.D., Burs-lem D.F.R.P., van der Wal R. The influence of vegetation type, soil properties and precipitation on the composition of soil mite and microbial communities at the landscape scale // J. Biogeogr. 2010. V. 37. P. 1317–1328. https://doi.org/10.1111/j.1365-2699.2010.02281.x
Olejniczak I., Boniecki P., Kaliszewicz A., Panteleeva N. The response of tundra springtails (Collembola, Hexapoda) to human activity on the Murman coast of the Kola Penisula, Russia // Polar Sci. 2018. V. 15. P. 99–103. https://doi.org/10.1016/j.polar.2017.12.005
Ponge J.F., Gillet S., Dubs F., Fedoroff E., Haese L., Sousa J.P., Lavelle P. Collembolan communities as bioindicators of land use intensification // Soil Biol. Biochem. 2003. V. 35. P. 813–826. https://doi.org/10.1016/S0038-0717(03)00108-1
Potapov M. Synopses on Palaearctic Collembola. V. 3. Isotomidae. Görlitz, 2001. 601 p.
Taskaeva A.A. Springtail (Collembola) assemblages in floodlands of the taiga zone of the Republic of Komi // Entomological Rev. 2009. V. 89. № 8. P. 965–974.
Thies C., Tscharntke T. Landscape structure and biological control in Agroecosystems // Science. 1999. V. 285. P. 893–895. https://doi.org/10.1126/science.285.5429.893
Tscharntke T., Klein A.M., Kruess A., Steffan-Dewenter I., Thies C. Landscape perspectives on agricultural intensification and biodiversity – ecosystem service management // Ecology Lett. 2005. V. 8. P. 857–874. https://doi.org/10.1111/j.1461-0248.2005.00782.x
van Capelle C., Schrader S., Brunotte J. Tillage-induced changes in the functional diversity of soil biota – A review with a focus on German data // Europ. J. Soil Biol. 2012. V. 50. P. 165–181. https://doi.org/10.1016/j.ejsobi.2012.02.005
Vandegehuchte M.L., Raschein U., Schütz1 M., Gwiazdowicz D.J., Risch1 A.C. Indirect short- and long-term effects of aboveground invertebrate and vertebrate herbivores on soil microarthropod communities // Plos one. 2015. P. 1–22. https://doi.org/10.1371/journal.pone.0118679
van Straalen N.M. Evaluation of bioindicator systems derived from soil arthropod communities // Applied soil ecology. 1998. V. 9. P. 429–437.
Vesterdal L., Ritter E., Gundersen P. Change in soil organic carbon following afforestation of former arable land // Forest Ecology and Management. 2002. V. 169. P. 137–147.
Дополнительные материалы отсутствуют.