

Почвоведение, 2019, № 7, стр. 827-842
Влияние растительности на характеристики лесных почв Республики Карелия
Н. В. Лукина 1, *, М. А. Орлова 1, О. Н. Бахмет 2, Е. В. Тихонова 1, Д. Н. Тебенькова 1, А. И. Казакова 1, А. М. Крышень 2, А. В. Горнов 1, В. Э. Смирнов 1, М. П. Шашков 3, В. В. Ершов 4, С. В. Князева 1
1 Центр по проблемам экологии и продуктивности лесов РАН
117997 Москва, ул. Профсоюзная, 84/32, стр. 14, Россия
2 Институт леса Карельского научного центра РАН
185910 Республика Карелия, Петрозаводск, ул. Пушкинская, 11, Россия
3 Институт физико-химических и биологических проблем почвоведения, Пущинский НЦ РАН
142290 Московская область, Пущино, ул. Институтская, 2, Россия
4 Институт проблем промышленной экологии Севера КНЦ РАН
184200 Апатиты, Мурманской обл, ул. Ферсмана, 7а, Россия
* E-mail: lukina@cepl.rssi.ru
Поступила в редакцию 05.07.2017
После доработки 05.10.2018
Принята к публикации 28.11.2018
Аннотация
Дана оценка влияния растительности на характеристики почв (кислотность, отношение C/N, содержание обменных форм элементов питания) северо- и средне-таежных лесов Республики Карелия, формирующихся в автономных позициях ландшафтов на почвообразующих породах варьирующего состава. Объектами исследований послужили распространенные в лесных биогеоценозах Карелии альфегумусовые почвы, приуроченные к автоморфным условиям на рыхлых четвертичных отложениях. Преобладают подзолы – Albic Podzols. Различия в характеристиках органогенных горизонтов почв, обнаруживаемые на уровне типов и формаций лесов в пределах и между подзонами, объясняются как варьированием состава почвообразующих пород и количеством осадков, так и влиянием растительности. К механизмам влияния растительности на характеристики почв, наряду с воздействием химического состава активной фракции опада растений доминирующих функциональных групп, можно отнести регулирование кронами деревьев разных видов и разного строения объема осадков, проникающих под полог, и соответственно интенсивности выноса соединений элементов из почв.
ВВЕДЕНИЕ
Растительность относится к биологическим факторам почвообразования. Влиянию растений на свойства лесных почв посвящен ряд работ прошлого и текущего столетия. Из механизмов влияния древесных и других лесных растений на почвы к наиболее известным, получившим экспериментальное подтверждение, относятся такие, как выветривание горных пород, формирование растениями доминирующих функциональных групп опада разного качества, определяющего состав органогенных горизонтов и скорость разложения [31, 32, 35], формирование ветровально-почвенных комплексов [7], регулирование количества и состава осадков, прошедших сквозь лесной полог [19]. Рассматривалось комбинированное влияние различных механизмов [10, 14, 25, 30].
Особенности почвообразования в Республике Карелия на породах различного генезиса исследованы довольно подробно [2–5, 12, 27]. Однако работы, посвященные оценке влияния растительности на наиболее динамично меняющиеся свойства лесных почв республики, необходимые для понимания взаимосвязей лесная растительность – почва, немногочисленны [15, 21–23].
Цель работы – оценить влияние растительности на характеристики почв северо- и средне-таежных лесов Республики Карелия, формирующихся в автономных позициях ландшафтов.
ОБЪЕКТЫ И МЕТОДЫ
Объекты исследований. Данные получены авторами в ходе работ в рамках международной программы ICP Forests в 2009–2010 гг. [46 ] . В Республике Карелия пункты постоянного наблюдения (ППН) заложены Российским центром защиты леса в узлах регулярной сети с шагом 32 км [6, 34]. ППН – кластеры с четырьмя круговыми пробными площадками, на которых произрастает не менее шести деревьев преобладающей породы. Круговые площадки расположены на расстоянии 25 м и ориентированы по сторонам света от центра ППН (центральное дерево). Проведен анализ данных по 69 ППН Республики Карелия, характеризующих растительность и почвы в автоморфных позициях ландшафта, среди которых 36 ППН расположены в северной тайге и 33 ППН – в средней тайге (рис. 1).

Средняя годовая температура в северной тайге Республика Карелия – около 0°C, а в средней тайге может достигать 4°C. Общее количество осадков в северной тайге составляет 450–500 мм, а в средней тайге на 150–200 мм больше. Республика Карелия отличается высокой неоднородностью почвообразующих пород [24]. В целом преобладают почвообразующие породы легкого гранулометрического состава с низкой влагоемкостью, высокой водопроницаемостью, бедные элементами питания, с разнообразными первичными минералами [18].
Объектами исследований послужили распространенные в лесных биогеоценозах Карелии альфегумусовые почвы, приуроченные к автоморфным условиям на рыхлых четвертичных отложениях. Преобладают подзолы: Albic Podzols (47 ППН). Подбуры (Entic Podzols) идентифицированы в северо-таежных ельниках черничных (2 ППН), березняке черничном (1 ППН), а также в средне-таежных сосняках брусничных (3 ППН), ельниках мелкотравных (4 ППН), ельниках черничных (4 ППН) и березняке черничном (5 ППН). Также в березовых лесах на двух ППН идентифицированы подзолистые почвы, на одном ППН – дерново-подзолистые почвы (Albic Retisols). Для объяснения изменений характеристик почв привлекались данные по объемам снеговых, дождевых кроновых и стволовых, а также почвенных вод, полученные в северо-таежных лесах Мурманской обл. на границе с Республикой Карелия с 2003 по 2012 гг. на пробных площадках, заложенных авторами статьи в соответствии с методами международной программы ICP Forests [17].
Методы исследований. Сравнительная характеристика растительности проводилась на экосистемном (типы леса) и видовом уровнях на ППН [17]. Номенклатура сосудистых растений дана по [29], листостебельных мхов – по [37], печеночных мхов – по [39], лишайников – по [26]. Растительность описывали на четырех площадках каждая площадью 100 м2, заложенных случайным образом в пределах каждого ППН. Для каждого из ярусов растительности проведена визуальная оценка общего проективного покрытия (в процентах). В работе принята классификация функциональных групп растений, основанная на делении видов растений по их таксономической принадлежности и жизненным формам [44]: 1 – злаки и осоки; 2 – травы – все травянистые растения, за исключением п. 1; 3 – кустарнички – низкорослые, не имеющие главного ствола многолетники с одревесневшими побегами; 4 – зеленые мхи; 5 – лишайники. Для каждой площадки составлены полные списки видов, затем списки четырех учетных площадок объединены в общий список для ППН с нахождением средних значений обилия видов.
Сравнение свойств почв проводили с учетом типов и формаций леса в пределах подзон северной и средней тайги. На четырех площадках на каждом ППН по горизонтам А0, E, B и BC отобраны по четыре образца почв, которые смешивались в одну пробу в соответствии с горизонтами на каждой площадке, то есть получали по одной смешанной пробе для каждого горизонта в пределах ППН. Почвообразующую породу характеризовали с использованием показателей кислотности, гранулометрического и валового состава гор. ВС. Объем дождевых кроновых и стволовых вод оценивали с применением осадкоприемников специальной конструкции [17], объем почвенных вод – с использованием гравитационных лизиметров, заложенных под различными горизонтами почв [17]. Ниже обсуждаются данные по объему вод, прошедших по почвенному профилю через органогенные, элювиальные и иллювиальные горизонты. Объем снеговых вод оценивали на основе данных снегосъемки в период максимального снегонакопления.
В лабораторных условиях почвенные образцы высушивали, просеивали через сито 2 мм. Аналитической обработке подвергалась фракция <2 мм, рН водной и солевой (0.01 M CaCl2) вытяжек определяли потенциометрически, обменную кислотность и обменный водород – титрованием до pH 7.8 [17]. Содержание органического углерода и общего азота определялось на CHNS-анализаторе PE-2040 фирмы Perkin Elmer (США).
Для оценки содержания обменных форм Ca, Fe, K, Mg, Mn, Na, Al в органогенных и верхних минеральных горизонтах почв в вытяжке 0.1 М BaCl2, рН 7.8 [17] использовали метод атомно-абсорбционной спектрометрии (ААС) на приборе AAnalist 800. Гранулометрический состав почв определяли по методу ISO/DIS 11277, согласно которому выделяют следующие гранулометрические фракции: <0.002 мм – ил; 0.002–0.063 мм – пыль; 0.063–2 мм – песок.
В образцах, отобранных из гор. ВС, методом ААС определяли валовое содержание металлов (Ca, Mg, K, Na, Al, Fe). Для разложения образцов использовали смесь фтористоводородной (плавиковой) и серной кислот.
Для оценки влияния преобладающей древесной породы на почвенные характеристики (на примере гор. A0) и функциональные группы растений (зеленые мхи, лишайники, кустарнички, травы, злаки) применяли v-критерий [36]. Для каждой категории q переменной номинального типа (в нашем случае это преобладающая порода, категории – сосна, ель, береза) и для каждой количественной переменной X, чьи значения поделены на категории, рассчитывается статистика критерия:
Для исследования связи между составом почвообразующих пород, органогенных и минеральных горизонтов проведен корреляционный анализ Спирмена. Достоверность различий по характеристикам почв оценивали с использованием U-критерия Манна–Уитни.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Характеристика растительности. Состав растительности северо- и средне-таежных лесов имеет как общие черты, так и существенные различия. В северной тайге Республики Карелия преобладают сосновые леса (72%), доля еловых и березовых лесов значительно меньше – 21 и 7.5% пробных площадей соответственно. Эти соотношения сопоставимы с полученными сведениями по данным лесной таксации [40]. В средней тайге Карелии вклад еловых и сосновых лесов в структуру растительности сопоставим (39 и 37% соответственно), возрастает доля березовых лесов (24%). Средний возраст древостоев сосны в северной и средней тайге различаются на один класс возраста (80 и 100 лет соответственно). Еловые древостои в северной тайге, напротив, старше, чем в средней (137 и 100 лет в среднем). Березовые древостои значительно моложе, чем сосновые и еловые, в среднем их возраст составлял 27 лет в северной тайге, 42 г. – в средней тайге, а предельный возраст не превышал 70 лет (табл. 1). Бонитет древостоев в средней тайге выше, запасы существенно больше, при этом число стволов на 1 га в сосновых и березовых древостоях средней тайги существенно меньше, чем в северной, а в средне-таежных и северо-таежных еловых лесах не различалось.
Таблица 1.
Таксационные характеристики объектов исследования
| Насаждения | Средний возраст, лет | Средняя высота, м | Средний диаметр, см | Бонитет | Число стволов*, шт./га | Запас, м3/га |
|---|---|---|---|---|---|---|
| Северная тайга | ||||||
| Сосняки | $\frac{{80}}{8}$ | $\frac{{13.6}}{{0.8}}$ | $\frac{{16.8}}{{1.2}}$ | $\frac{{3.6}}{{0.2}}$ | $\frac{{1009}}{{124}}$ | $\frac{{113.5}}{{11.8}}$ |
| Ельники | $\frac{{137}}{{14}}$ | $\frac{{14.0}}{{1.7}}$ | $\frac{{19.7}}{{2.2}}$ | $\frac{{4.6}}{{0.2}}$ | $\frac{{582}}{{150}}$ | $\frac{{133.6}}{{36.1}}$ |
| Березняки | $\frac{{27}}{1}$ | $\frac{{8.0}}{{0.8}}$ | $\frac{{8.0}}{{0.0}}$ | $\frac{{4.0}}{{0.0}}$ | $\frac{{2874}}{{1480}}$ | $\frac{{61.7}}{{17.8}}$ |
| Средняя тайга | ||||||
| Сосняки | $\frac{{100}}{{17}}$ | $\frac{{19.2}}{{1.0}}$ | $\frac{{22.6}}{{2.4}}$ | $\frac{{2.8}}{{0.3}}$ | $\frac{{552}}{{109}}$ | $\frac{{165.9}}{{19.9}}$ |
| Ельники | $\frac{{101}}{6}$ | $\frac{{22.4}}{{1.1}}$ | $\frac{{27.1}}{{1.9}}$ | $\frac{{2.8}}{{0.2}}$ | $\frac{{494}}{{76}}$ | $\frac{{233.6}}{{17.7}}$ |
| Березняки | $\frac{{42}}{3}$ | $\frac{{15.8}}{{1.1}}$ | $\frac{{13.6}}{{1.2}}$ | $\frac{{2.5}}{{0.3}}$ | $\frac{{1659}}{{374}}$ | $\frac{{151.8}}{{18.2}}$ |
* Деревья 1–3 классов Крафта. Примечание. Здесь и в табл. 3, 4: над чертой – среднее, под чертой – ошибка среднего.
Анализ типологической структуры лесов северной тайги показал доминирование сосняков кустарничково-зеленомошных с преобладанием бруснично-зеленомошных типов (49%), тогда как доля сосняков зеленомошно-лишайниковых и лишайниковых значительно меньше (11 и 7.5% соответственно), а вклад ельников кустарничково-зеленомошных, среди которых представлены черничные типы, составляет всего 15%.
Типологическая структура лесов средней тайги Карелии имеет выраженные различия со структурой лесов в северной тайге. Наиболее представлены ельники кустарничково-зеленомошные (33%), среди которых преобладают ельники черничные. Заметно участие мелкотравных типов. Доля сосняков кустарничково-зеленомошных значительно меньше (13%) и сопоставима с участием березняков – кустарничково-зеленомошных (11%). Сосняки лишайниковые не идентифицированы.
Структура напочвенного покрова в северной и средней тайге также различается. В северной тайге среднее проективное покрытие кустарничков больше, чем в средней, почти вдвое (44%), а проективное покрытие лишайников – более чем в 3 раза (11%). В средней тайге проективное покрытие видов группы трав в 3 раза больше, чем в северной (рис. 2). Как и следовало ожидать, флористическое разнообразие оказалось обильнее в средней тайге. Сходство видового состава сообществ северной и средней тайги, определенное с использованием индекса Съеренсена, составляло 0.68. В северной тайге отмечено 107 видов сосудистых растений, 66 видов мхов и 19 видов лишайников. В средней тайге – 164, 94, и 24 видов соответственно. Особенно значительные различия отмечены в группе трав [44]: в средней тайге обнаружено 100 видов, тогда как в северной – только 57 видов.
Рис. 2.
Проективное покрытие напочвенного покрова в северной (1) и средней (2) тайге: А – сосняк брусничный, Б – сосняк черничный, В – ельник черничный.

Большинство видов-доминантов имеют значительное распространение как в северной, так и в средней тайге – черника обыкновенная (Vaccinium myrtillus), брусника обыкновенная (Vaccinium vitis-idaea), Плевроциум Шребера (Pleurozium schreberi), Гилокомиум блестящий (Hylocomium splendens), Дикранум многоножковый (Dicranum polysetum), Дикранум метловидный (Dicranum scoparium).
В северной тайге отсутствуют (или встречены единично) многие неморальные и бореально-неморальные виды, часть из которых в средней тайге имеет высокую встречаемость: ландыш майский (Convallaria majalis) – 41%, дудник лесной (Angelica sylvestris) – 24%, кочедыжник женский (Athyrium filix-femina) – 20%. Встречаемость ряда видов из групп “злаки” и “травы” в средней тайге существенно большее, чем в северной тайге: вейник тростниковый (Calamagrostis arundinacea), щучка дернистая (Deschampsia cespitosa), малина обыкновенная (Rubus idaeus), майник двулистный (Maianthemum bifolium) (63 и 17%), кислица обыкновенная (Oxalis acetosella) (44 и 6%), грушанка малая (Pyrola minor) (17 и 2%), костяника (Rubus saxatilis) (52 и 13%), щитовник картузианский (Dryopteris carthusiana) (52 и 9%). Среди мхов необходимо отметить виды, имеющие большее распространение в средней тайге: мниумы (Mnium sp.), мхи из родов брахитециум (Brachythecium sp.) и плагиотециум (Plagiothecium sp.).
В то же время в северной тайге большую встречаемость имеют многие виды лишайников – Кладония лесная (Cladonia arbuscula), Кладония оленья (Cladonia rangiferina), Кладония звездчатая (Cladonia stellaris) и др. Среди видов сосудистых растений Водяника черная (Empetrum nigrum s. l.) имеет значительно большее распространение в северной тайге.
Оценка влияния древесных пород на функциональные группы растений с использованием v-критерия показала (табл. 2), что сосна имеет положительную связь с покрытием кустарничков и лишайников и отрицательную – с покрытием трав и злаков. Для ели и березы эти связи имеют противоположный знак; в сосновых лесах покрытия мхов и лишайников значимо больше среднего, в еловых и березовых – значимо меньше. Необходимо иметь в виду, что совместное произрастание древесных видов с определенными видами напочвенного покрова может быть связано со сходными требованиями к условиям местообитаний или конкурентоспособностью. Например, лишайниковые типы леса характерны для самых сухих местообитаний, где сосна с ее глубокой корневой системой является наиболее конкурентоспособной из всех видов деревьев [11].
Таблица 2.
Статистические показатели, связанные с v-критерием, для функциональных групп растений нижних ярусов и характеристик гор. А0 на уровне формаций
| Показатель | v-критерий | Среднее по формации | Стандартное отклонение по формации | р | Общее среднее | Общее стандартное отклонение | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| формация n(сосновая) = 38, n(еловая) = 21, n(березовая) = 10 | ||||||||||||||
| сосновая | еловая | березовая | сосновая | еловая | березовая | сосновая | еловая | березовая | сосновая | еловая | березовая | |||
| Проективное покрытие лишайников, % | 2.9 | –1.9 | –1.6 | 11.5 | 1.7 | 0.1 | 18.2 | 4.5 | 0.1 | 0.004 | 0.054 | 0.115 | 6.9 | 14.6 |
| Проективное покрытие зеленых мхов, % | 1.9 | 0.9 | –3.9 | 52.3 | 51.3 | 19.6 | 23.5 | 19.2 | 14.3 | 0.056 | 0.357 | 0.000 | 47.3 | 24.0 |
| Проективное покрытие кустарничков, % | 4.0 | –2.4 | –2.5 | 46.9 | 28.5 | 22.5 | 18.8 | 18.2 | 13.5 | 0.000 | 0.015 | 0.012 | 37.8 | 20.7 |
| Проективное покрытие злаков, % | –3.9 | 2.0 | 2.8 | 1.2 | 4.6 | 6.5 | 2.3 | 5.5 | 3.1 | 0.000 | 0.043 | 0.005 | 3.0 | 4.2 |
| Проективное покрытие трав, % | –4.1 | 3.2 | 1.7 | 1.2 | 13.1 | 12.0 | 1.8 | 17.0 | 7.9 | 0.000 | 0.001 | 0.098 | 6.4 | 11.5 |
| pH водный | –3.5 | 1.1 | 3.6 | 4.25 | 4.52 | 4.92 | 0.24 | 0.46 | 0.67 | 0.000 | 0.276 | 0.000 | 4.43 | 0.46 |
| Обменный Al, смоль(+)/кг | 2.8 | –1.6 | –1.8 | 1.19 | 0.67 | 0.47 | 1.01 | 0.52 | 0.38 | 0.006 | 0.109 | 0.072 | 0.93 | 0.87 |
| Обменный Ca, смоль(+)/кг | –4.3 | 3.3 | 1.7 | 9.18 | 15.51 | 14.94 | 3.49 | 6.00 | 7.21 | 0.000 | 0.001 | 0.084 | 11.94 | 5.88 |
| Обменный Mg, смоль(+)/кг | –4.7 | 1.9 | 4.1 | 2.00 | 3.33 | 4.64 | 0.82 | 1.46 | 1.61 | 0.000 | 0.054 | 0.000 | 2.79 | 1.52 |
| Обменный K, смоль(+)/кг | –1.8 | 0.2 | 2.3 | 2.05 | 2.19 | 2.54 | 0.50 | 0.55 | 0.65 | 0.071 | 0.832 | 0.023 | 2.16 | 0.56 |
| BS, % | –4.7 | 3.0 | 2.7 | 71 | 84 | 87 | 11 | 9 | 7 | 0.000 | 0.003 | 0.007 | 77 | 12 |
| С общ, % | 2.9 | –1.3 | –2.3 | 41.4 | 36.3 | 32.1 | 6.9 | 9.3 | 12.1 | 0.004 | 0.191 | 0.019 | 38.5 | 9.3 |
| N общ, % | –4.7 | 3.9 | 1.6 | 0.87 | 1.23 | 1.15 | 0.21 | 0.29 | 0.22 | 0.000 | 0.000 | 0.116 | 1.02 | 0.29 |
| C/N | 4.7 | –3.2 | –2.5 | 51 | 30 | 28 | 19 | 7 | 10 | 0.000 | 0.001 | 0.012 | 41 | 19 |
Примечание. Здесь и в табл. 4: BS (base saturation) – насыщенность основаниями, %; n – число ППН.
Характеристика почвообразующих пород и горизонта ВС. Почвообразующими породами на объектах исследований являются четвертичные отложения, покрывающие более 60% территории Карелии. В составе коренных пород принимают участие кварц, плагиоклаз, калиевые полевые шпаты и слюды [8].
О различиях в составе почвообразующих пород между подзонами тайги свидетельствуют данные валового и гранулометрического состава гор. ВС (рис. 3, табл. 3). Валовое содержание кальция в гор. ВС почв средне-таежных лесов всех типов леса существенно (р < 0.01) меньше, чем в северо-таежных, тогда как магния содержится значительно (р < 0.05) больше в гор. ВС почв средне-таежных ельников. Актуальная кислотность гор. ВС почв северо-таежных и средне-таежных сосновых лесов сходных типов (аналогов), так же как и pH солевой вытяжки, не различалась. При этом актуальная кислотность гор. ВС почв средне-таежных ельников черничных достоверно (p < 0.05) больше по сравнению с их северо-таежными аналогами.
Рис. 3.
Гранулометрический состав минеральных горизонтов почв в северной (1) и средней (2) тайге; среднее и ошибки среднего содержания фракций (%) в трех горизонтах (E, B, BC) на уровне формаций: сосна – сосновые леса, ель – еловые леса.

Таблица 3.
Данные о pH и валовом содержании металлов в гор. ВС
| Тип леса (повторность) | pH | Ca | Mg | К | Na | Al | Fe | |
|---|---|---|---|---|---|---|---|---|
| CaCl2 | H2O | г/кг | ||||||
| Северная тайга | ||||||||
| Сосняк зеленомошно-лишайниковый (n = 8) | $\frac{{4.93}}{{0.16}}$ | $\frac{{5.44}}{{0.22}}$ | $\frac{{14.8}}{{1.6}}$ | $\frac{{8.0}}{{2.8}}$ | $\frac{{10.5}}{{0.8}}$ | $\frac{{13.4}}{{1.2}}$ | $\frac{{41.7}}{{2.2}}$ | $\frac{{19.0}}{{4.9}}$ |
| Сосняк брусничный (n = 11) | $\frac{{4.95}}{{0.06}}$ | $\frac{{5.75}}{{0.10}}$ | $\frac{{17.0}}{{1.5}}$ | $\frac{{6.2}}{{0.7}}$ | $\frac{{8.5}}{{0.3}}$ | $\frac{{13.6}}{{0.6}}$ | $\frac{{42.6}}{{1.9}}$ | $\frac{{19.0}}{{1.5}}$ |
| Сосняк черничный (n = 5) | $\frac{{4.98}}{{0.06}}$ | $\frac{{5.30}}{{0.41}}$ | $\frac{{18.0}}{{1.6}}$ | $\frac{{8.5}}{{0.6}}$ | $\frac{{8.7}}{{0.7}}$ | $\frac{{14.7}}{{0.9}}$ | $\frac{{36.1}}{{1.5}}$ | $\frac{{15.3}}{{1.4}}$ |
| Ельник черничный (n = 5) | $\frac{{4.96}}{{0.11}}$ | $\frac{{5.74}}{{0.07}}$ | $\frac{{16.8}}{{1.6}}$ | $\frac{{7.5}}{{1.1}}$ | $\frac{{9.2}}{{0.5}}$ | $\frac{{13.7}}{{1.3}}$ | $\frac{{40.2}}{{5.5}}$ | $\frac{{18.1}}{{3.5}}$ |
| Березняк черничный (n = 2) | $\frac{{4.60}}{{0.20}}$ | $\frac{{5.18}}{{0.30}}$ | $\frac{{18.9}}{{1.7}}$ | $\frac{{9.4}}{{0.8}}$ | $\frac{{7.9}}{{2.7}}$ | $\frac{{14.3}}{{0.1}}$ | $\frac{{32.7}}{{2.0}}$ | $\frac{{15.3}}{{1.1}}$ |
| Средняя тайга | ||||||||
| Сосняк брусничный (n = 7) | $\frac{{4.84}}{{0.14}}$ | $\frac{{5.80}}{{0.09}}$ | $\frac{{12.3}}{{0.8}}$ | $\frac{{6.3}}{{0.7}}$ | $\frac{{10.4}}{{0.6}}$ | $\frac{{13.2}}{{1.0}}$ | $\frac{{35.3}}{{2.2}}$ | $\frac{{14.9}}{{0.9}}$ |
| Сосняк черничный (n = 3) | $\frac{{4.90}}{{0.12}}$ | $\frac{{5.50}}{{0.36}}$ | $\frac{{11.3}}{{1.6}}$ | $\frac{{6.5}}{{2.2}}$ | $\frac{{10.4}}{{1.0}}$ | $\frac{{12.1}}{{1.4}}$ | $\frac{{36.4}}{{0.4}}$ | $\frac{{13.9}}{{1.9}}$ |
| Ельник черничный (n = 8) | $\frac{{4.68}}{{0.08}}$ | $\frac{{5.23}}{{0.22}}$ | $\frac{{11.2}}{{0.3}}$ | $\frac{{9.9}}{{1.9}}$ | $\frac{{11.0}}{{0.4}}$ | $\frac{{12.1}}{{0.9}}$ | $\frac{{36.4}}{{0.8}}$ | $\frac{{17.6}}{{3.0}}$ |
| Ельник мелкотравный (n = 4) | $\frac{{4.23}}{{0.23}}$ | $\frac{{4.58}}{{0.39}}$ | $\frac{{13.8}}{{0.6}}$ | $\frac{{13.9}}{{3.9}}$ | $\frac{{10.3}}{{0.5}}$ | $\frac{{11.5}}{{0.9}}$ | $\frac{{38.3}}{{0.7}}$ | $\frac{{22.8}}{{3.0}}$ |
| Березняк черничный (n = 7) | $\frac{{4.34}}{{0.14}}$ | $\frac{{5.27}}{{0.09}}$ | $\frac{{13.1}}{{1.1}}$ | $\frac{{6.2}}{{1.0}}$ | $\frac{{10.4}}{{0.3}}$ | $\frac{{14.1}}{{0.8}}$ | $\frac{{36.9}}{{2.0}}$ | $\frac{{14.9}}{{1.4}}$ |
Различия в гранулометрическом составе гор. ВС между подзонами обнаружены только в сосновых лесах: содержание фракции песка в этих горизонтах почв в северо-таежных лесах значительно меньше (р < 0.01), а содержание пылеватых и илистых фракций (до 0.063 мм) существенно больше (р < 0.05), чем в средней тайге. В гор. ВС еловых лесов достоверные различия в гранулометрическом составе между подзонами не выявлены, можно лишь отметить сходную с сосновыми лесами тенденцию по фракции пыли.
При сравнении в пределах подзон между типами леса обнаруживается, что в средней тайге гор. ВС почв ельников, развивающихся на подбурах, отличаются высокой кислотностью и содержат существенно больше магния и железа по сравнению с другими типами лесов. Среднее содержание тонких фракций в этих горизонтах еловых и сосновых лесов сопоставимо.
Характеристика органогенных горизонтов почв. Характеристики органогенных гор. А0 почв отличаются высокой вариабельностью в пределах подзон тайги, что связано с принадлежностью лесов к разным формациям и типам. В северной тайге актуальная и обменная кислотность органогенных горизонтов почв формации сосновых лесов достоверно (p < 0.05) больше, а содержание обменного кальция и магния в них меньше, чем в органогенных горизонтах почв еловых лесов (рис. 4, табл. 4). Содержание азота и степень насыщенности основаниями существенно больше, а отношение C/N значительно уже в органогенных горизонтах почв еловых и березовых лесов по сравнению с почвами сосновых лесов.
Рис. 4.
С/N, среднее и ошибки среднего содержания обменных форм элементов питания и отношения C/N в гор. А0 в северной (1) и средней (2) тайге; формации: сосна – сосновые леса, ель – еловые леса, береза – березовые леса.
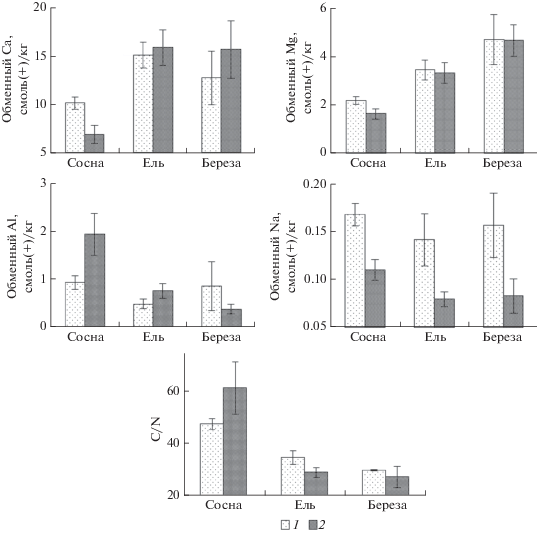
Таблица 4.
Кислотность, содержание общего углерода, азота, обменных форм элементов в гор. А0, Е, В
| Тип леса (повторность) | Гори-зонт | С общ | N общ | C/N | pH | Обменная кислот- ность | Обмен- ный Al | Обмен- ный Н | BS, % | Обмен- ный Ca | Обмен- ный Mg | Обмен- ный К | Обмен- ный Na | Обмен- ное Fe | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % | H2O | CaCl2 | смоль(+)/кг | смоль(+)/кг | |||||||||||
| Северная тайга | |||||||||||||||
| Сосняк зеленомошно-лишайниковый (n = 9) | А0 | $\frac{{40.4}}{{1.5}}$ | $\frac{{0.91}}{{0.07}}$ | $\frac{{47}}{4}$ | $\frac{{4.30}}{{0.06}}$ | $\frac{{3.27}}{{0.08}}$ | $\frac{{5.05}}{{0.53}}$ | $\frac{{0.99}}{{0.22}}$ | $\frac{{4.09}}{{0.39}}$ | $\frac{{72}}{3}$ | $\frac{{8.91}}{{0.76}}$ | $\frac{{1.99}}{{0.20}}$ | $\frac{{1.94}}{{0.17}}$ | $\frac{{0.19}}{{0.03}}$ | $\frac{{0.17}}{{0.05}}$ |
| Е | $\frac{{1.3}}{{0.2}}$ | $\frac{{0.03}}{{0.01}}$ | $\frac{{66}}{{13}}$ | $\frac{{3.74}}{{0.11}}$ | $\frac{{4.50}}{{0.16}}$ | $\frac{{1.13}}{{0.10}}$ | $\frac{{1.13}}{{0.10}}$ | $\frac{{0.25}}{{0.04}}$ | $\frac{{21}}{4}$ | $\frac{{0.19}}{{0.04}}$ | $\frac{{0.10}}{{0.03}}$ | $\frac{{0.06}}{{0.02}}$ | $\frac{{0.03}}{{0.00}}$ | $\frac{{0.07}}{{0.01}}$ | |
| В | $\frac{{1.2}}{{0.2}}$ | $\frac{{0.04}}{{0.01}}$ | $\frac{{39}}{9}$ | $\frac{{4.86}}{{0.11}}$ | $\frac{{5.44}}{{0.07}}$ | $\frac{{0.71}}{{0.44}}$ | $\frac{{0.90}}{{0.46}}$ | $\frac{{0.09}}{{0.02}}$ | $\frac{{49}}{5}$ | $\frac{{0.22}}{{0.08}}$ | $\frac{{0.10}}{{0.05}}$ | $\frac{{0.04}}{{0.02}}$ | $\frac{{0.03}}{{0.01}}$ | $\frac{{0.05}}{{0.01}}$ | |
| Сосняк брусничный (n = 13) | А0 | $\frac{{41.5}}{{1.9}}$ | $\frac{{0.88}}{{0.05}}$ | $\frac{{49}}{3}$ | $\frac{{4.25}}{{0.09}}$ | $\frac{{3.36}}{{0.06}}$ | $\frac{{4.91}}{{0.41}}$ | $\frac{{0.97}}{{0.25}}$ | $\frac{{4.01}}{{0.24}}$ | $\frac{{74}}{3}$ | $\frac{{10.45}}{{1.18}}$ | $\frac{{2.21}}{{0.24}}$ | $\frac{{2.25}}{{0.11}}$ | $\frac{{0.17}}{{0.01}}$ | $\frac{{0.12}}{{0.04}}$ |
| Е | $\frac{{0.8}}{{0.2}}$ | $\frac{{0.01}}{{0.00}}$ | $\frac{{61}}{{25}}$ | $\frac{{3.80}}{{0.09}}$ | $\frac{{4.44}}{{0.18}}$ | $\frac{{1.34}}{{0.13}}$ | $\frac{{1.07}}{{0.11}}$ | $\frac{{0.22}}{{0.03}}$ | $\frac{{23}}{5}$ | $\frac{{0.24}}{{0.06}}$ | $\frac{{0.08}}{{0.01}}$ | $\frac{{0.03}}{{0.00}}$ | $\frac{{0.03}}{{0.00}}$ | $\frac{{0.05}}{{0.00}}$ | |
| В | $\frac{{0.7}}{{0.1}}$ | $\frac{{0.05}}{{0.02}}$ | $\frac{{19}}{4}$ | $\frac{{5.05}}{{0.06}}$ | $\frac{{5.49}}{{0.10}}$ | $\frac{{0.18}}{{0.03}}$ | $\frac{{0.26}}{{0.03}}$ | $\frac{{0.07}}{{0.01}}$ | $\frac{{62}}{4}$ | $\frac{{0.15}}{{0.02}}$ | $\frac{{0.07}}{{0.00}}$ | $\frac{{0.02}}{{0.00}}$ | $\frac{{0.03}}{{0.00}}$ | $\frac{{0.03}}{{0.00}}$ | |
| Сосняк черничный (n = 6) | А0 | $\frac{{43.1}}{{0.9}}$ | $\frac{{1.00}}{{0.09}}$ | $\frac{{46}}{5}$ | $\frac{{4.33}}{{0.05}}$ | $\frac{{3.33}}{{0.07}}$ | $\frac{{4.26}}{{0.24}}$ | $\frac{{0.73}}{{0.16}}$ | $\frac{{3.79}}{{0.29}}$ | $\frac{{78}}{2}$ | $\frac{{10.81}}{{1.10}}$ | $\frac{{2.25}}{{0.49}}$ | $\frac{{2.38}}{{0.10}}$ | $\frac{{0.13}}{{0.01}}$ | $\frac{{0.05}}{{0.01}}$ |
| Е | $\frac{{0.8}}{{0.1}}$ | $\frac{{0.03}}{{0.00}}$ | $\frac{{32}}{5}$ | $\frac{{3.72}}{{0.14}}$ | $\frac{{4.35}}{{0.27}}$ | $\frac{{1.40}}{{0.40}}$ | $\frac{{1.05}}{{0.28}}$ | $\frac{{0.28}}{{0.08}}$ | $\frac{{25}}{6}$ | $\frac{{0.16}}{{0.01}}$ | $\frac{{0.08}}{{0.00}}$ | $\frac{{0.03}}{{0.00}}$ | $\frac{{0.03}}{{0.00}}$ | $\frac{{0.05}}{{0.01}}$ | |
| В | $\frac{{0.9}}{{0.1}}$ | $\frac{{0.03}}{{0.00}}$ | $\frac{{31}}{6}$ | $\frac{{5.02}}{{0.08}}$ | $\frac{{5.35}}{{0.25}}$ | $\frac{{0.20}}{{0.03}}$ | $\frac{{0.27}}{{0.03}}$ | $\frac{{0.07}}{{0.01}}$ | $\frac{{62}}{5}$ | $\frac{{0.23}}{{0.08}}$ | $\frac{{0.10}}{{0.03}}$ | $\frac{{0.03}}{{0.00}}$ | $\frac{{0.03}}{{0.00}}$ | $\frac{{0.03}}{{0.00}}$ | |
| Ельник черничный (n = 6) | А0 | $\frac{{41.1}}{{1.9}}$ | $\frac{{1.23}}{{0.12}}$ | $\frac{{35}}{3}$ | $\frac{{4.52}}{{0.13}}$ | $\frac{{3.57}}{{0.16}}$ | $\frac{{3.84}}{{0.55}}$ | $\frac{{0.48}}{{0.10}}$ | $\frac{{3.40}}{{0.44}}$ | $\frac{{84}}{3}$ | $\frac{{14.95}}{{1.34}}$ | $\frac{{3.42}}{{0.41}}$ | $\frac{{2.37}}{{0.21}}$ | $\frac{{0.14}}{{0.03}}$ | $\frac{{0.05}}{{0.02}}$ |
| Е | $\frac{{10.5}}{{2.8}}$ | $\frac{{0.18}}{{0.04}}$ | $\frac{{36}}{8}$ | $\frac{{3.50}}{{0.11}}$ | $\frac{{4.51}}{{0.13}}$ | $\frac{{2.15}}{{0.52}}$ | $\frac{{1.66}}{{0.44}}$ | $\frac{{0.40}}{{0.10}}$ | $\frac{{14}}{4}$ | $\frac{{0.07}}{{0.00}}$ | $\frac{{0.06}}{{0.01}}$ | $\frac{{0.06}}{{0.02}}$ | $\frac{{0.03}}{{0.01}}$ | $\frac{{0.05}}{{0.01}}$ | |
| В | $\frac{{1.2}}{{0.7}}$ | $\frac{{0.05}}{{0.01}}$ | $\frac{{38}}{8}$ | $\frac{{4.93}}{{0.17}}$ | $\frac{{5.33}}{{0.21}}$ | $\frac{{0.61}}{{0.30}}$ | $\frac{{0.57}}{{0.26}}$ | $\frac{{0.10}}{{0.02}}$ | $\frac{{50}}{8}$ | $\frac{{0.23}}{{0.06}}$ | $\frac{{0.11}}{{0.03}}$ | $\frac{{0.03}}{{0.00}}$ | $\frac{{0.03}}{{0.00}}$ | $\frac{{0.05}}{{0.01}}$ | |
| Березняк черничный (n = 2) | А0 | $\frac{{38.6}}{{6.5}}$ | $\frac{{1.30}}{{0.23}}$ | $\frac{{30}}{0}$ | $\frac{{4.41}}{{0.04}}$ | $\frac{{3.65}}{{0.05}}$ | $\frac{{3.96}}{{0.12}}$ | $\frac{{0.85}}{{0.51}}$ | $\frac{{3.53}}{{0.14}}$ | $\frac{{83}}{4}$ | $\frac{{12.60}}{{2.74}}$ | $\frac{{4.67}}{{1.04}}$ | $\frac{{2.75}}{{0.64}}$ | $\frac{{0.16}}{{0.03}}$ | $\frac{{0.04}}{ - }$ |
| Е | 0.7 | 0.05 | 13 | 3.90 | 4.40 | 0.92 | 0.65 | 0.19 | 34 | 0.20 | 0.18 | 0.06 | 0.03 | 0.05 | |
| В | $\frac{{5.5}}{{4.0}}$ | $\frac{{0.13}}{{0.12}}$ | $\frac{{92}}{{53}}$ | $\frac{{4.42}}{{0.18}}$ | $\frac{{5.44}}{{0.08}}$ | $\frac{{1.00}}{{0.45}}$ | $\frac{{0.84}}{{0.43}}$ | $\frac{{0.16}}{{0.08}}$ | $\frac{{60}}{{19}}$ | $\frac{{2.17}}{{1.93}}$ | $\frac{{0.56}}{{0.48}}$ | $\frac{{0.08}}{{0.05}}$ | $\frac{{0.04}}{{0.01}}$ | $\frac{{0.08}}{{0.02}}$ | |
| Средняя тайга | |||||||||||||||
| Сосняк брусничный (n = 7) | А0 | $\frac{{38.3}}{{3.7}}$ | $\frac{{0.78}}{{0.09}}$ | $\frac{{52}}{6}$ | $\frac{{4.21}}{{0.10}}$ | $\frac{{3.04}}{{0.07}}$ | $\frac{{5.98}}{{0.78}}$ | $\frac{{2.13}}{{0.59}}$ | $\frac{{3.50}}{{0.50}}$ | $\frac{{59}}{4}$ | $\frac{{6.02}}{{1.18}}$ | $\frac{{1.54}}{{0.27}}$ | $\frac{{1.61}}{{0.23}}$ | $\frac{{0.10}}{{0.01}}$ | $\frac{{0.18}}{{0.04}}$ |
| Е | $\frac{{0.6}}{{0.1}}$ | $\frac{{0.03}}{{0.00}}$ | $\frac{{26}}{7}$ | $\frac{{3.70}}{{0.22}}$ | $\frac{{4.67}}{{0.15}}$ | $\frac{{1.98}}{{0.50}}$ | $\frac{{1.41}}{{0.36}}$ | $\frac{{0.37}}{{0.07}}$ | $\frac{{11}}{4}$ | $\frac{{0.09}}{{0.02}}$ | $\frac{{0.04}}{{0.01}}$ | $\frac{{0.03}}{{0.01}}$ | $\frac{{0.03}}{{0.00}}$ | $\frac{{0.09}}{{0.02}}$ | |
| В | $\frac{{1.5}}{{0.4}}$ | $\frac{{0.04}}{{0.01}}$ | $\frac{{39}}{6}$ | $\frac{{4.64}}{{0.11}}$ | $\frac{{4.97}}{{0.16}}$ | $\frac{{0.76}}{{0.18}}$ | $\frac{{0.52}}{{0.12}}$ | $\frac{{0.25}}{{0.07}}$ | $\frac{{20}}{5}$ | $\frac{{0.06}}{{0.01}}$ | $\frac{{0.03}}{{0.00}}$ | $\frac{{0.02}}{{0.00}}$ | $\frac{{0.02}}{{0.00}}$ | $\frac{{0.05}}{{0.01}}$ | |
| Сосняк черничный (n = 3) | А0 | $\frac{{47.8}}{{6.6}}$ | $\frac{{0.66}}{{0.12}}$ | $\frac{{53}}{4}$ | $\frac{{4.07}}{{0.08}}$ | $\frac{{3.03}}{{0.19}}$ | $\frac{{6.03}}{{1.41}}$ | $\frac{{1.47}}{{0.49}}$ | $\frac{{4.29}}{{1.07}}$ | $\frac{{68}}{6}$ | $\frac{{8.60}}{{0.41}}$ | $\frac{{1.74}}{{0.40}}$ | $\frac{{1.93}}{{0.02}}$ | $\frac{{0.14}}{{0.00}}$ | $\frac{{0.17}}{{0.02}}$ |
| Е | $\frac{{2.5}}{{0.7}}$ | $\frac{{0.29}}{{0.19}}$ | $\frac{{21}}{9}$ | $\frac{{3.73}}{{0.22}}$ | $\frac{{4.66}}{{0.20}}$ | $\frac{{2.17}}{{0.47}}$ | $\frac{{1.45}}{{0.18}}$ | $\frac{{0.57}}{{0.29}}$ | $\frac{{13}}{3}$ | $\frac{{0.15}}{{0.04}}$ | $\frac{{0.06}}{{0.01}}$ | $\frac{{0.04}}{{0.01}}$ | $\frac{{0.03}}{{0.01}}$ | $\frac{{0.08}}{{0.02}}$ | |
| В | $\frac{{1.2}}{{0.2}}$ | $\frac{{0.04}}{{0.01}}$ | $\frac{{36}}{{12}}$ | $\frac{{4.67}}{{0.13}}$ | $\frac{{5.01}}{{0.22}}$ | $\frac{{0.79}}{{0.37}}$ | $\frac{{0.52}}{{0.25}}$ | $\frac{{0.19}}{{0.09}}$ | $\frac{{24}}{9}$ | $\frac{{0.08}}{{0.03}}$ | $\frac{{0.05}}{{0.01}}$ | $\frac{{0.03}}{{0.00}}$ | $\frac{{0.02}}{{0.00}}$ | $\frac{{0.06}}{{0.03}}$ | |
| Ельник черничный (n = 11) | А0 | $\frac{{33.9}}{{2.1}}$ | $\frac{{1.19}}{{0.09}}$ | $\frac{{30}}{2}$ | $\frac{{4.48}}{{0.17}}$ | $\frac{{3.51}}{{0.15}}$ | $\frac{{3.82}}{{0.55}}$ | $\frac{{0.77}}{{0.20}}$ | $\frac{{3.05}}{{0.37}}$ | $\frac{{83}}{3}$ | $\frac{{15.30}}{{2.06}}$ | $\frac{{3.37}}{{0.49}}$ | $\frac{{2.25}}{{0.17}}$ | $\frac{{0.08}}{{0.01}}$ | $\frac{{0.09}}{{0.03}}$ |
| Е | $\frac{{1.2}}{{0.3}}$ | $\frac{{0.05}}{{0.02}}$ | $\frac{{35}}{8}$ | $\frac{{3.69}}{{0.10}}$ | $\frac{{4.34}}{{0.13}}$ | $\frac{{2.61}}{{0.30}}$ | $\frac{{2.00}}{{0.26}}$ | $\frac{{0.46}}{{0.03}}$ | $\frac{{12}}{1}$ | $\frac{{0.19}}{{0.04}}$ | $\frac{{0.09}}{{0.03}}$ | $\frac{{0.04}}{{0.01}}$ | $\frac{{0.03}}{{0.01}}$ | $\frac{{0.12}}{{0.02}}$ | |
| В | $\frac{{1.2}}{{0.3}}$ |  $\frac{{0.08}}{{0.02}}$ $\frac{{0.08}}{{0.02}}$ |
$\frac{{24}}{5}$ | $\frac{{4.45}}{{0.10}}$ | $\frac{{5.14}}{{0.14}}$ | $\frac{{1.21}}{{0.26}}$ | $\frac{{0.94}}{{0.21}}$ | $\frac{{0.31}}{{0.04}}$ | $\frac{{38}}{7}$ | $\frac{{0.72}}{{0.40}}$ | $\frac{{0.45}}{{0.31}}$ | $\frac{{0.06}}{{0.02}}$ | $\frac{{0.04}}{{0.01}}$ | $\frac{{0.06}}{{0.01}}$ | |
| Ельник мелкотравный (n = 4) | А0 | $\frac{{35.5}}{{9.1}}$ | $\frac{{1.31}}{{0.18}}$ | $\frac{{27}}{4}$ | $\frac{{4.65}}{{0.24}}$ | $\frac{{3.75}}{{0.19}}$ | $\frac{{2.70}}{{0.78}}$ | $\frac{{0.71}}{{0.25}}$ | $\frac{{1.85}}{{0.53}}$ | $\frac{{86}}{6}$ | $\frac{{16.92}}{{4.34}}$ | $\frac{{3.06}}{{1.04}}$ | $\frac{{1.74}}{{0.24}}$ | $\frac{{0.08}}{{0.01}}$ | $\frac{{0.14}}{{0.06}}$ |
| В | $\frac{{3.9}}{{2.4}}$ | $\frac{{0.41}}{{0.35}}$ | $\frac{{22}}{6}$ | $\frac{{3.98}}{{0.26}}$ | $\frac{{4.59}}{{0.26}}$ | $\frac{{3.76}}{{2.26}}$ | $\frac{{1.09}}{{0.18}}$ | $\frac{{0.72}}{{0.37}}$ | $\frac{{28}}{9}$ | $\frac{{0.32}}{{0.20}}$ | $\frac{{0.19}}{{0.13}}$ | $\frac{{0.03}}{{0.00}}$ | $\frac{{0.05}}{{0.01}}$ | $\frac{{0.06}}{{0.01}}$ | |
| Березняк черничный (n = 8) | А0 | $\frac{{30.5}}{{4.8}}$ | $\frac{{1.12}}{{0.08}}$ | $\frac{{27}}{4}$ | $\frac{{5.05}}{{0.26}}$ | $\frac{{4.09}}{{0.27}}$ | $\frac{{2.92}}{{0.64}}$ | $\frac{{0.37}}{{0.10}}$ | $\frac{{2.25}}{{0.55}}$ | $\frac{{88}}{3}$ | $\frac{{15.52}}{{2.96}}$ | $\frac{{4.63}}{{0.65}}$ | $\frac{{2.49}}{{0.24}}$ | $\frac{{0.08}}{{0.02}}$ | $\frac{{0.05}}{{0.02}}$ |
| Е | $\frac{{0.75}}{{0.09}}$ | $\frac{{0.03}}{{0.02}}$ | $\frac{{64}}{{47}}$ | $\frac{{3.80}}{{0.20}}$ | $\frac{{4.25}}{{0.45}}$ | $\frac{{1.54}}{{0.11}}$ | $\frac{{1.14}}{{0.15}}$ | $\frac{{0.26}}{{0.00}}$ | $\frac{{15}}{3}$ | $\frac{{0.16}}{{0.08}}$ | $\frac{{0.06}}{{0.02}}$ | $\frac{{0.02}}{{0.00}}$ | $\frac{{0.03}}{{0.01}}$ | $\frac{{0.07}}{{0.01}}$ | |
| В | $\frac{{1.1}}{{0.2}}$ | $\frac{{0.15}}{{0.09}}$ | $\frac{{18}}{3}$ | $\frac{{4.26}}{{0.12}}$ | $\frac{{5.08}}{{0.18}}$ | $\frac{{1.03}}{{0.11}}$ | $\frac{{0.74}}{{0.09}}$ | $\frac{{0.25}}{{0.04}}$ | $\frac{{33}}{9}$ | $\frac{{0.24}}{{0.05}}$ | $\frac{{0.06}}{{0.01}}$ | $\frac{{0.05}}{{0.01}}$ | $\frac{{0.02}}{{0.00}}$ | $\frac{{0.05}}{{0.01}}$ | |
Сходные, но еще более ярко выраженные различия в характеристиках органогенных горизонтов почв между формациями лесов выражены в средней тайге.
Результаты оценки влияния разных древесных пород на характеристики почв на объектах исследований с использованием v-критерия демонстрируют, что сосна способствует формированию более кислых органогенных горизонтов, бедных азотом, что объясняет широкое отношение C/N. Содержание обменного кальция и магния в этих горизонтах почв существенно меньше, чем в среднем для всех трех формаций лесов (табл. 2). Береза, как и ель, напротив, обусловливает формирование органогенных горизонтов с относительно высоким содержанием общего азота и узким отношением C/N. При этом содержание обменного алюминия достоверно меньше, чем в среднем для органогенных горизонтов рассматриваемых формаций лесов, а содержание обменного калия и кальция, наоборот, больше, что объясняет относительно высокую степень насыщенности этих горизонтов почв основаниями. Следует заметить, что при разделении данных на северо-таежные и средне-таежные леса выявляются яркие различия между органогенными горизонтами почв сосновых, еловых и березовых лесов (рис. 4).
Различия в кислотности и содержании обменных катионов в органогенных горизонтах почв лесов разных формаций связаны, прежде всего, с химическим составом активной фракции опада доминирующих древесных растений. Как известно, стареющая хвоя ели богата кальцием [14], сосны – натрием [45], а листья березы – основаниями и азотом [9, 16]. Свой вклад вносит и опад растений нижних ярусов, например, кустарнички рода Vaccinium известны как концентраторы кальция [38]. Кроме того, существенное накопление элементов питания в органогенных горизонтах почв еловых лесов по сравнению с сосновыми можно объяснить различиями в строении крон двух доминирующих в бореальных лесах древесных пород – ели и сосны [13–15, 21, 33]. Плотная и протяженная крона ели препятствует формированию значительных объемов стволовых вод и выносу с почвенными водами из верхних почвенных горизонтов соединений элементов, тогда как сосна функционирует как “воронка” для стока вод по стволу (рис. 5). Большой объем стволовых вод может способствовать формированию гораздо более значительных объемов вод из минеральных горизонтов в сосняках по сравнению с ельниками.
Рис. 5.
Отношение объема воды в сосняках кустарничково-лишайниково-зеленомошных и ельниках кустарничково-зеленомошных в северной тайге. Обозначения: 1 – почвенные воды между кронами, 2 – почвенные воды под кронами, 3 – стволовые воды.

Еще одним фактором может быть более активная жизнедеятельность грибов, биомасса которых в органогенных горизонтах почв березовых и еловых лесов значительно больше, чем сосновых [20, 28], что приводит к увеличению содержания доступных для биоты, в том числе обменных, форм соединений элементов.
Таким образом, оценку различий в составе органогенных горизонтов почв между подзонами тайги целесообразно проводить с учетом формаций леса. При сравнении обнаруживается, что органогенные горизонты почв северо-таежных сосновых лесов содержат достоверно больше (p < 0.01–0.05) обменного кальция, натрия и меньше обменного алюминия, чем их аналоги в средней тайге. Для еловых и березовых лесов достоверные различия между северо-таежными и средне-таежными аналогами выявлены для обменного натрия, содержание которого, как и в случае сосновых лесов, оказалось достоверно (p < 0.05) больше в органогенных горизонтах почв северо-таежных лесов (рис. 4). Средние значения отношения C/N в органогенных горизонтах почв еловых и березовых лесов средне-таежных лесов меньше, чем северо-таежных, но различия для березовых лесов статистически не достоверны. В сосновых лесах эти показатели между подзонами не различались.
Различия в содержании обменного кальция в органогенных горизонтах почв сосновых лесов двух подзон можно объяснить составом почвообразующих пород, о чем свидетельствует валовое содержание кальция в гор. ВС. Поскольку сосна отличается глубокой корневой системой, что позволяет ей получать элементы питания из более глубоких минеральных горизонтов почв, состав опада сосны может отражать состав почвообразующих пород. Различия в содержании обменного натрия в органогенных горизонтах не связано с его содержанием в почвообразующих породах и могут объясняться накоплением натрия в опаде доминирующих растений северной тайги, что приводит к увеличению содержания обменного натрия в органогенных горизонтах. Необходимо подчеркнуть, что, как хорошо известно, возраст опадающих частей растений, составляющих фракцию активного опада (хвоя, листья и др.), в северной тайге значительно выше. Поскольку натрий относится к элементам, не способным к ретранслокации внутри растений, уровень его накопления в опадающих органах растений северной тайги существенно выше, чем средней. Следует заметить, что в северной тайге на территории Карелии доминируют сосновые леса, а хвоя сосны накапливает натрий с возрастом в больших количествах [45].
Более выраженные различия в отношении C/N в органогенных горизонтах почв обнаруживаются между формациями лесов в пределах подзон, по сравнению с различиями между подзонами при сравнении аналогичных типов леса. Достоверные различия соотношения C/N и других показателей плодородия почв также выявлены в органогенных горизонтах между типами лесов Каяндера в Финляндии на регулярной сети ICP Forests [44]. Отношение C/N в органогенных горизонтах еловых и березовых лесов существенно уже, чем сосновых, в обеих подзонах (табл. 4). Различия между подзонами по обменному кальцию и магнию в органогенных горизонтах почв ельников не обнаружены, несмотря на то, что его валовое содержание в гор. ВС значительно больше в еловых лесах северной тайги, а магния – в средней. Отсутствие различий между подзонами можно объяснить тем, что в отличие от сосны, у ели поверхностная корневая система, поэтому состав хвои ели – основного компонента активной фракции опада, в меньшей степени, чем хвои сосны, зависит от состава минеральных горизонтов, отражающих особенности почвообразующих пород.
В целом можно заключить, что различия в характеристиках органогенных горизонтов почв, обнаруживаемые при сравнениях на уровне формаций и типов лесов и на уровне подзон, связаны с различиями в составе почвообразующих пород, особенно, в сосновых лесах, так и с влиянием растительности, что особенно ярко выражено в еловых и березовых лесах. При этом наиболее выраженные различия обнаруживаются в пределах подзон между формациями лесов. К механизмам влияния лесной растительности на характеристики органогенных горизонтов почв, наряду с химическим составом опада, можно отнести влияние различий в строении крон деревьев ели и сосны (их плотности и протяженности), регулирующих количество проникающих под полог осадков и интенсивность выноса соединений элементов питания из почв.
ХАРАКТЕРИСТИКА ВЕРХНИХ МИНЕРАЛЬНЫХ ГОРИЗОНТОВ ПОЧВ
Гранулометрический состав. Различия в гранулометрическом составе верхних минеральных горизонтов почв между подзонами тайги часто не соответствуют различиям, выявленным для гор. ВС (рис. 3). В сосновых лесах наблюдается соответствие различий между подзонами в гор. ВС и В только по фракции ила. В отличие от гор. ВС, в элювиальных и, особенно, иллювиальных горизонтах почв средне-таежных ельников содержание ила оказалось больше, чем у их северо-таежных аналогов. Это может быть связано с более интенсивными процессами почвообразования в средне-таежных еловых лесах, приводящими к возрастанию фракции тонких частиц в минеральных горизонтах почв. Следует отметить, что в северной тайге между сосновыми и еловыми лесами не обнаруживается различий в содержании фракции ила ни в гор. ВС, ни в верхних минеральных горизонтах. В средней тайге ситуация другая: содержание фракции ила в гор. В еловых лесов существенно больше, чем в сосновых. По фракции пыли во всех горизонтах почв еловых лесов и в верхних минеральных гор. Е и В почв сосновых лесов различий между подзонами не обнаруживается. Только в гор. ВС почв сосновых лесов содержание пыли достоверно (p < 0.05) больше в северной тайге. При этом в северной тайге содержание пыли в гор. Е почв еловых лесов существенно больше, чем сосновых, тогда как в гор. В и ВС почв северо-таежных лесов и во всех горизонтах почв средне-таежных лесов различия не достоверны.
Таким образом, хотя гор. ВС сосновых северо-таежных лесов содержат больше фракций ила и пыли, чем средне-таежных, сходные различия в верхней части минерального профиля отмечаются только в гор. В и только для фракции ила. При невыраженных различиях по гранулометрическому составу между подзонами в гор. ВС почв еловых лесов, в гор. Е и В проявляются различия по фракции ила, содержание которого больше в средней тайге (рис. 2). В иллювиальных горизонтах почв средне-таежных еловых лесов содержание ила существенно больше, чем сосновых.
Сравнение характеристик элювиальных горизонтов почв. Различия в актуальной кислотности элювиальных горизонтов почв между сосновыми и еловыми лесами в пределах обеих подзон не выявлены, но обнаружены различия по другим показателям (табл. 4). При сопоставимых значениях обменной кислотности, содержание обменного магния оказалось достоверно (p < 0.05) больше в гор. Е средне-таежных ельников черничных по сравнению с сосняками этого же типа, что соответствует различиям в валовом составе гор. ВС.
В северной тайге обменная кислотность, содержание обменного алюминия и водорода в гор. Е почв сосновых лесов оказались меньше, а сумма оснований (за счет обменного кальция и магния) и степень насыщенности их основаниями больше, чем в еловых лесах при сопоставимой актуальной кислотности, то есть обнаруживаются обратные по сравнению с органогенными горизонтами закономерности. При повышенном содержании обменного кальция и магния в органогенных горизонтах почв еловых лесов и сходном валовом содержании этих элементов в гор. ВС почв северо-таежных сосновых и еловых лесов, эти различия можно объяснить менее ярко выраженными процессами выноса оснований из органогенных горизонтов почв еловых лесов по сравнению с сосновыми из-за менее интенсивного потока воды из органогенных горизонтов почв еловых лесов в минеральные (рис. 5).
Содержание азота в гор. Е отличалось существенной вариабельностью внутри типов леса, поэтому достоверные различия между формациями/типами леса внутри подзон не установлены. Содержание углерода в гор. Е средне-таежных ельников черничных оказалось достоверно больше (p < 0.05), чем в других типах леса. Отношение C/N в элювиальных горизонтах почв между типами леса внутри подзон не различалось.
При сравнении между подзонами тайги обнаруживается более высокое содержание обменного магния в элювиальных горизонтах почв средне-таежных еловых лесов по сравнению с их северо-таежными аналогами, что соответствует различиям между подзонами в составе почвообразующих пород. При этом, несмотря на большее валовое содержание кальция в почвообразующих породах ельников северной тайги, содержание его обменных форм значительно больше в элювиальных горизонтах почв средне-таежных ельников, что объясняет и повышенную степень насыщенности их основаниями.
Содержание общего азота в элювиальных горизонтах почв сосновых лесов северной тайги меньше, чем в средней тайге. Отношение C/N в элювиальных горизонтах почв средне-таежных еловых и сосновых лесов сопоставимо при невыраженных различиях в содержании углерода и высокой вариабельности содержания азота.
Содержание обменного алюминия оказалось меньше в элювиальных горизонтах почв северо-таежных сосновых лесов по сравнению с их средне-таежными аналогами, что при сопоставимых валовых содержаниях в почвообразующих породах можно объяснить более активными процессами выветривания, вероятно, обусловленными жизнедеятельностью микроорганизмов в средне-таежных лесах. Это подтверждается и тем, что содержание обменного алюминия и в северной, и средней тайге оказалось меньше в гор. Е почв сосновых лесов по сравнению с этими же горизонтами почв еловых лесов, в которых биомасса грибов существенно больше [20, 28]. Установлено также, что концентрация низкомолекулярных органических кислот в почвенных водах еловых лесов больше, чем сосновых [1, 42], особенно под кронами елей, что может являться результатом экссудации корневых выделений и влиять на концентрацию обменного алюминия в почвах [41].
Таким образом, различия в характеристиках элювиальных горизонтов почв между подзонами могут объясняться как составом почвообразующих пород, так и различиями в интенсивности выноса соединений элементов из почв, регулируемых деревьями разного строения, принадлежащих к разным видам.
Сравнение характеристик иллювиальных горизонтов почв. На основе полученных данных установлены различия между формациями/типами лесов в характеристиках иллювиальных горизонтов почв в пределах подзон (табл. 4). Обменная кислотность и содержание обменного водорода и алюминия достоверно (p < 0.01–0.05) больше в иллювиальных горизонтах почв еловых лесов по сравнению с сосновыми лесами в обеих подзонах, в основном за счет обменного алюминия, так же как в гор. Е почв северной тайги.
По содержанию обменных оснований в иллювиальных горизонтах почв северной тайги различий между еловыми и сосновыми лесами не обнаружено. В средней тайге содержание обменного кальция и магния в иллювиальных горизонтах почв еловых лесах достоверно (p < 0.05) больше по сравнению с сосновыми, что соответствует различиям в органогенных горизонтах почв. По содержанию органического углерода в иллювиальных горизонтах почв различий между сосновыми и еловыми лесами сходного типа ни в северной, ни в средней тайге не обнаружено. По содержанию азота в иллювиальных горизонтах северо-таежные еловые и сосновые леса сходного типа достоверно также не различались, а в средней тайге содержание азота, как и обменных оснований, существенно (p < 0.05) больше в иллювиальных горизонтах почв еловых лесов, что соответствует различиям в органогенных горизонтах почв. Иллювиальные горизонты почв средне-таежных березовых лесов, так же как и еловых, оказались существенно богаче обменным кальцием и общим азотом, чем сосновых, что также соответствует различиям в органогенных горизонтах почв, из которых эти соединения мигрируют.
Между подзонами статистически значимые различия обнаруживаются в актуальной кислотности иллювиальных горизонтов почв сосновых лесов, которая оказалась достоверно (p < 0.05) больше в средней тайге. Достоверные (p < 0.05) различия сходного характера обнаруживаются для pH солевой вытяжки: кислее иллювиальные горизонты почв средне-таежных сосновых лесов по сравнению с их северо-таежными аналогами. Для pH солевой вытяжки в иллювиальных горизонтах почв еловых лесов установлены сходные различия. Обменная кислотность и содержание обменного водорода и алюминия также достоверно (p < 0.01–0.05) больше в иллювиальных горизонтах почв средне-таежных сосновых и еловых лесов по сравнению с их северо-таежными аналогами. Высокую кислотность иллювиальных горизонтов почв средне-таежных сосновых и еловых лесов можно объяснить более интенсивным кислотообразованием, в том числе грибным из-за большей биомассы грибов.
Содержание обменного кальция существенно больше в иллювиальных горизонтах почв северо-таежных сосновых лесов по сравнению с их средне-таежными аналогами, что можно объяснить составом почвообразующих пород. Однако в иллювиальных горизонтах почв северо-таежных еловых лесов содержание обменного кальция, напротив, существенно меньше, несмотря на высокое валовое содержание кальция в гор. ВС. Это может быть связано с большим количеством осадков и более высокой интенсивностью процессов вымывания оснований из почв в средней тайге. Содержание общего углерода существенно больше в иллювиальных горизонтах почв средне-таежных сосновых лесов по сравнению с их северо-таежными аналогами, а по содержанию азота достоверных различий не выявлено. В еловых лесах, напротив, по общему углероду различий не обнаруживается, а содержание общего азота существенно больше в средней тайге. Относительно высокое содержание органического углерода и общего азота в иллювиальных горизонтах почв средне-таежных лесов может быть связано с более интенсивным, чем в северной тайге, выносом их соединений осадками.
Таким образом, различия в характеристиках иллювиальных горизонтов почв, так же как и элювиальных, могут объясняться как составом почвообразующих пород, так и количеством атмосферных выпадений, регулирующих вынос соединений из верхних органогенных горизонтов почв.
Полученные результаты демонстрируют, что для сравнения почв северной и средней тайги в целом необходимо учитывать вклад тех или иных формаций/типов лесов в общую структуру лесного покрова подзоны. При таком сравнении обнаруживается, что органогенные горизонты почв средне-таежных лесов, в составе которых высока доля ельников и березняков, содержат меньше углерода и больше общего азота и отличаются узким соотношением C/N от этих горизонтов почв северо-таежных лесов. Органогенные горизонты почв северо-таежных лесов содержат значительно больше обменного натрия, который концентрируется в хвое сосны, формирующей доминирующие в северной тайге леса. Элювиальные горизонты почв средне-таежных лесов по сравнению с их северо-таежными аналогами отличаются высоким содержанием обменного алюминия и общего азота, узким соотношением C/N,а иллювиальные горизонты почв этих лесов содержат больше углерода, азота, обменного алюминия и водорода. Наблюдаемые различия связаны не только с составом почвообразующих пород, но с интенсивностью выноса соединений элементов из почв, определяемой количеством осадков, проникающих под кроны деревьев разных видов и разного строения, и с активностью почвенной микробиоты, определяющей интенсивность обменных процессов в почвах.
ЗАКЛЮЧЕНИЕ
Характеристики почв северо-таежных и средне-таежных лесов Республики Карелия, формирующихся в автоморфных ландшафтах, варьируют в зависимости от состава растительности и почвообразующих пород. Анализ состава растительности лесов северной и средней тайги выявил как общие черты, так и существенные различия между ними в формационной и типологической структуре, таксационных характеристиках и возрасте древостоев. В северной тайге доминируют сосновые леса с большой долей кустарничков, особенно брусники, в напочвенном покрове, в средней тайге высока доля еловых и березовых лесов с более значительным вкладом трав и злаков. Возраст древостоев ели больше в северной тайге, а сосны и березы – в средней.
Валовой состав гор. ВС свидетельствует о том, что почвообразующие породы северо-таежных еловых, сосновых и березовых лесов отличаются более высоким валовым содержанием кальция по сравнению с подзоной средней тайги, тогда как почвообразующие породы средне-таежных ельников содержат больше валового магния. Данные гранулометрического состава демонстрируют, что гор. ВС северо-таежных сосновых лесов содержит больше тонких фракций, чем средне-таежных. Обнаружено, что различия между подзонами северной и средней тайги Республики Карелия в гранулометрическом составе верхних минеральных горизонтов (элювиальных и иллювиальных) часто не совпадают с различиями в почвообразующих породах, что объясняется разной интенсивностью почвообразования в лесах разного типа средней и северной тайги. В верхних минеральных горизонтах почв средне-таежных еловых лесов, по сравнению с сосновыми лесами, достоверно больше содержание фракции ила.
Различия в характеристиках органогенных горизонтов почв, обнаруживаемые при сравнениях на уровне формаций лесов, объясняются разным составом почвообразующих пород, особенно, в сосновых лесах, и влиянием древесной породы. Различия в характеристиках почв между формациями в пределах двух подзон выражены более ярко, чем между подзонами. В обеих подзонах органогенные горизонты почв еловых и березовых лесов оказались богаче обменным кальцием, магнием, общим азотом и характеризуются низкой кислотностью и узким отношением C/N по сравнению с сосновыми лесами.
К механизмам влияния растительности на характеристики органогенных горизонтов почв, наряду с воздействием химического состава активной фракции опада растений доминирующих функциональных групп, относится влияние строения крон деревьев разных видов, регулирующих объем осадков, проникающих под полог леса, и соответственно, интенсивность выноса соединений элементов из почвенных горизонтов.
Различия в характеристиках элювиальных и иллювиальных горизонтов почв внутри и между подзонами объясняются составом почвообразующих пород, особенностями строения крон доминирующих древесных растений и количеством осадков, регулирующих вынос соединений элементов из верхних органогенных горизонтов почв.
Благодарность. Сбор информации проводился в рамках темы государственного задания ЦЭПЛ РАН “Сохранение и восстановление экологических функций лесных почв” (0110-2014-0004) и программы Президиума РАН “Биоразнообразие природных систем”, проект: “Лесообразующие виды России: пространственное распределение, запасы, вклад в экосистемные функции и услуги” (0110-2015-0005). Химический анализ почв выполнен за счет средств гранта Российского фонда фундаментальных исследований (проект 15-29-02697 офи_м). Обработка результатов исследования выполнена за счет гранта Российского научного фонда (проект № 16-17-10284).
Список литературы
Артемкина Н.А., Горбачева Т.Т., Лукина Н.В. Низкомолекулярные органические кислоты в почвенных водах лесов Кольского полуострова // Лесоведение. 2008. № 6. С. 37–44.
Ахметова Г.В. Содержание микроэлементов в почвообразующих породах и лесных почвах озерно-ледниковых равнин среднетаежной подзоны Карелии // Лесной вестник. 2008. № 2. С. 16–20.
Бахмет О.Н. Особенности органического вещества почв в различных ландшафтах Карелии // Лесоведение. 2012. № 2. С. 19–27.
Бахмет О.Н. Особенности почв скальных ландшафтов карельского побережья белого моря // Уч. зап. Петрозаводского гос. ун-та. Сер. Естественные и технические науки. 2013. № 6(135). С. 55–59.
Бахмет О.Н., Морозова Р.М. Почвенный покров // Разнообразие биоты Карелии: условия формирования, сообщества, виды. Петрозаводск: Карельский научный центр РАН. 2003. С. 34–37.
Бахмет О.Н., Федорец Н.Г., Крышень А.М. Исследования по международной программе ICP-forests в Карелии // Тр. Карельского научного центра РАН. 2011. № 2. С. 133–139.
Васенев И.И., Таргульян В.О. Ветровал и таежное почвообразование (режимы, процессы, морфогенез почвенных сукцессий). М.: Наука, 1995. 247 с.
Геология Карелии / Под ред. В.А. Соколова. Л.: Наука, 1987. 231 с.
Казимиров Н.И., Морозова Р.М. Биологический круговорот веществ в ельниках Карелии. Л.: Наука, 1973. 175 с.
Карпачевский Л.О. Лес и лесные почвы. М.: Лесная промышленность, 1981. 262 с.
Кучеров И.Б., Зверев А.А. Лишайниковые сосняки средней и северной тайги Европейской России // Вестник Томского гос. ун-та. Биология. 2012. № 3(19). С. 46–80.
Лесовая С.Н., Горячкин С.В., Заварзин А.А., Погожее Е.Ю., Полеховский Ю.С. Специфика бореального почвообразования и выветривания на плотных породах (на примере заповедника “Кивач”, Карелия) // Вестник Санкт-Петербургского ун-та. Сер. 3. Биология. 2006. № 1. С. 106–118.
Лукина Н.В., Горбачева Т.Т., Никонов В.В., Лукина М.А. Пространственная изменчивость кислотности Al–Fe-гумусовых подзолов // Почвоведение. 2002. № 2. С. 163–176.
Лукина Н.В., Орлова М.А., Исаева Л.Г. Плодородие лесных почв как основа взаимосвязи почва-растительность // Лесоведение. 2010. № 5. С. 45–56.
Лукина Н.В., Полянская Л.М., Орлова М.А. Питательный режим почв северотаежных лесов. М.: Наука, 2008. 341 с.
Манаков К.Н., Никонов В.В. Биологический круговорот минеральных элементов и почвообразование в ельниках Крайнего Севера. Л.: Наука, 1981. 196 с.
Методика мониторинга лесов по международной программе ICPForests М.: ФГУ “Российский центр защиты леса”, ЦЭПЛ РАН, 2008. 46 с.
Морозова Р.М. Географические закономерности формирования почвенного покрова Карелии // Биогеография Карелии. Тр. КарНЦ РАН. Сер. Биология. 2001. № 2. С. 12–18.
Никонов В.В., Лукина Н.В. Влияние ели и сосны на кислотность и состав атмосферных выпадений в северотаежных лесах индустриально-развитого региона // Экология. 2000. № 2. С. 82–90.
Никонов В.В., Лукина Н.В., Полянская Л.М., Паникова А.Н. Особенности распространения микроорганизмов в Al–Fe-гумусовых подзолах северо-таежных еловых лесов: природные и техногенные аспекты // Микробиология. 2001. Т. 70. № 3. С. 374–383.
Орлова М.А., Лукина Н.В., Смирнов В.Э., Артемкина Н.А. Влияние ели на кислотность и содержание элементов питания в почвах северотаежных ельников кустарничково-зеленомошных // Почвоведение. 2016. № 11. С. 1355–1367.
Орлова М.А., Лукина Н.В., Смирнов В.Э., Исаева Л.Г. Плодородие почв березовых лесов на северном пределе распространения // Почвоведение. 2014. № 3. С. 1–13.
Орлова М.А., Лукина Н.В., Смирнов В.Э., Краснов Д.А., Камаев И.О. Плодородие почв еловых лесов Хибинских гор // Почвоведение. 2012. № 6. С. 682–694.
Разнообразие биоты Карелии: условия формирования, сообщества, виды / Под ред. А.Н. Громцева, С.П. Китаева, В.И. Крутова, О.Л. Кузнецова, Т. Линдхольм, Е.Б. Яковлева. Петрозаводск: Карельский научный центр РАН, 2003. 262 с.
Ремезов Н.П. Еще о роли леса в почвообразовании // Почвоведение. 1956. № 4. С. 70–79.
Урбанавичюс Г.П., Урбанавичене И.Н. Лишайники заповедников России // Современное состояние биологического разнообразия на заповедных территориях России. Вып. 3. Лишайники и мохообразные. М.: Гриф и К, 2004. С. 5–243.
Федорец Н.Г., Морозова Р.М., Солодовников А.Н. Лесные почвы Карелии и оценка их продуктивности // Тр. Карельского научного центра РАН. 2003. № 5. С. 108–120.
Фомичева О.А., Полянская Л.М., Никонов В.В., Лукина Н.В., Орлова М.А., Исаева Л.Г., Звягинцев Д.Г. Численность и биомасса почвенных микроорганизмов в коренных старовозрастных северо-таежных еловых лесах // Почвоведение. 2006. № 12. С. 1469–1478.
Черепанов С.К. Сосудистые растения России и сопредельных государств. СПб.: Мир и семья, 1995. 992 с.
Binkley D., Giardina C. Why do tree species affect soil? The warp and woof of tree-soil interaction // Biogeochemistry. 1998. V. 42. P. 89–106.
Cornelissen J.H.C. et al. Global negative vegetation feedback to climate warming responses of leaf litter decomposition rates in cold biomes // Ecology letters. 2007. V. 10. № 7. P. 619–627.
Cornelissen J.H.C., Thompson K. Functional leaf attributes predict litter decomposition rate in herbaceous plants // The New Phytologist. 1997. V. 135. № 1. P. 109–114.
Ershov V.V., Lukina N.V., Orlova M.A., Zukert N.V. Dynamics of snowmelt water composition in conifer forests exposed to air-born industrial pollution // Russian J. Ecology. 2016. V. 47. № 1. P. 46–52.
Forest condition in Europe: 2011 technical report of ICP Forests and FutMon. Work Report of the Institute for World Forestry // ICP Forests. Hamburg, 2011. 212 p.
Hansson K., Olsson B.A., Olsson M., Johansson U., Kleja D.B. Differences in soil properties in adjacent stands of Scots pine, Norway spruce and silver birch in Sweden // Forest Ecology and Management. 2011. V. 262. № 3. P. 522–530.
Husson F., Le S., Pagиs J. Exploratory multivariate analysis by example using R / 2nd edition. Chapman & Hall/CRC. 2017. 248 p.
Ignatov M.S., Afonina O.M., Ignatova E.A., Abolina A., Akatova T.V., Baisheva E.Z., Bardunov L.V., Baryakina E.A., Belkina O.A., Bezgodov A.G., Boychuk Ya., Cherdantseva V.M.A., Czernyadjeva I.V., Doroshina G.Ya., Dyachenko A.P., Fedosov V.E., Goldberg I.L., Ivanova E.I., Jukoniene I., Kannukene L., Kazanovsky S.G., Kharzinov Z.Kh., Kurbatova L.E., Maksimov A.I., Mamatkulov U.K.A.I., Manakyan V.A., Maslovsky O.M., Napreenko M.G., Otnyukova T.N., Partyka L.Ya, Pisarenko O.Yu, Popova N.N., Rykovsky G.F., Tubanova D.Ya, Zheleznova G.V., Zolotov V.I. Check list of mosses of East Europe and North Asia // Arctoa. 2006. V. 16. P. 1–130.
Ingestad T. Mineral nutrient requirements of Vaccinium vitis idaea and V. myrtillus // Physiologia plantarum. 1973. V. 29. № 2. P. 239–246.
Konstantinova N.A., Bakalin V.A., Andrejeva E.N., Bezgodov A.G., Borovichev E.A., Dulin M.V., Mamontov Yu.S. Checklist of liverworts (Marchantiophyta) of Russia // Arctoa. 2009. V. 18. P. 1–63.
Lukina N.V., Orlova M.A., Gornov A.V., Kuznetsov P.V., Knyazeva S.V., Smirnov V.E., Isaeva L.G. Assessment of sustainable forest management criteria using indicators of the international program mme ICP forests // Contemporary problems of ecology. 2013. V. 6. № 7. P. 734–745.
Murakami T., Utsunomiya S., Yokoyama T., Kasama T. Biotite dissolution processes and mechanisms in the laboratory and in nature: early stage weathering environment and vermi culitization // Am. Mineral. 2003. V. 88. P. 377–386.
Ohta T., Hiura T. Erratum: Root exudation of low-molecular-mass-organic acids by six tree species alters the dynamics of calcium and magnesium in soil // Canadian J. Soil Science. 2016. V. 96. № 4. P. 516–516.
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria, 2017. URL: http://www.R-project.org.
Salemaa M., Derome J., Nцjd P. Response of boreal forest vegetation to the fertility status of the organic layer along a climatic gradient // Boreal Environment Research. 2008. V. 13. P. 48–66.
Steinnes E., Lukina N., Nikonov V., Aamlid D., Royset O. A gradient study of 34 elements in the vicinity of a copper-nickel smelter in the Kola Peninsula // Environmental monitoring and assessment. 2000. V. 60. № 1. P. 71–88.
Дополнительные материалы отсутствуют.