

Почвоведение, 2019, № 9, стр. 1092-1103
Скорость разложения растений-торфообразователей в олиготрофных болотах южно-таежной подзоны Западной Сибири: оценка влияния уровня болотных вод и температуры торфяной залежи
Л. Г. Никонова 1, Е. А. Головацкая 1, *, И. В. Курьина 1, И. Н. Курганова 2
1 Институт мониторинга климатических и экологических систем СО РАН
634050 Томск, Академический пр-т, 10/3, Россия
2 Институт физико-химических и биологических проблем почвоведения РАН
142290 Московская область, Пущино, ул. Институтская, 2, Россия
* E-mail: golovatskayaea@gmail.com
Поступила в редакцию 06.08.2018
После доработки 13.11.2018
Принята к публикации 30.01.2019
Аннотация
В полевом эксперименте изучали скорость разложения четырех видов растений-торфообразователей, наиболее типичных для болотных фитоценозов южно-таежной подзоны Западной Сибири: Sphagnum fuscum, Chamaedaphne calyculata, Eriophorum vaginatum и Sphagnum angustifolium. Исследования проводили методом частично изолированных проб на двух болотах: Тимирязевском и Бакчарском, различающихся по условиям влаго- и теплообеспеченности. Независимо от химического состава растений-торфообразователей, процесс их разложения характеризовался самой значительной потерей массы в течение первого года исследований. К концу третьего года эксперимента происходило замедление процессов деструкции растительных остатков, и величина констант разложения всех исследуемых видов растений уменьшалась в 1.2–2.8 раза. Наиболее медленно процесс деструкции растительных остатков протекал в более влажных условиях Бакчарского болота, где потери массы для всех исследуемых видов растений за исключением Eriophorum vaginatum были в 1.2–1.6 раз меньше, чем на Тимирязевском болоте. По величине суммарных потерь массы и константам разложения достоверно за 3 года эксперимента отличались Sphagnum fuscum и Chamaedaphne calyculata, а также два вида сфагнума: Sphagnum fuscum и Sphagnum angustifolium. Выявлены значимые положительные корреляционные связи между потерями массы растений-торфообразователей (кроме Sphagnum angustifolium) и суммами температур торфяной залежи, превышающими 0, 5 и 10°С. Понижение уровня болотных вод приводило к достоверному росту интенсивности процессов разложения всех исследуемых видов растений за исключением Eriophorum vaginatum.
ВВЕДЕНИЕ
Занимая около 5% от поверхности суши, болотные экосистемы играют значительную роль в глобальном круговороте углерода (С) [4, 46, 55]. В болотах в виде торфа сосредоточены значительные запасы органического С 120–455 Пг C (1 Пг = = 1015 г), накопленные здесь в течение нескольких тысячелетий [4, 34, 38, 51]. Торфообразование происходит в болотных экосистемах за счет активных процессов консервации растительных остатков в условиях постоянного переувлажнения [14, 15, 30]. В настоящее время исследование процессов разложения органического вещества (ОВ) в болотных экосистемах приобретает особую актуальность, поскольку наблюдаемые и прогнозируемые изменения климата в значительной степени определяют гидротермический режим болот и через него влияют на процессы минерализации как свежих растительных остатков, так и торфяной залежи в целом [44]. Например, понижение уровня болотных вод (УБВ) может привести к увеличению зоны аэрации и дополнительному высвобождению питательных веществ за счет усиления процессов минерализации, что в свою очередь может вызвать увеличение чистой первичной продукции болотных экосистем, обусловленное более активным ростом подземных органов растений [2, 4, 7, 12]. Таким образом, в условиях изменяющегося климата соотношение потоков углерода в болотных экосистемах может меняться, влияя тем самым на глобальный баланс С, ввиду огромных площадей, занятых болотами [53, 54].
На основе анализа многолетних рядов метеорологических данных было показано, что текущие изменения климата на территории Западной Сибири выражаются в общем повышении среднегодовой температуры воздуха за счет более заметного потепления зимнего периода и в увеличении осадков весеннего сезона [22]. К середине текущего столетия в Сибирском регионе прогнозируется дальнейшее увеличение среднегодовой температуры за счет сокращения числа морозных дней, накопления массы снега зимой и его интенсивного таяния весной [5, 22]. Однако прогнозируемое потепление и изменение увлажненности могут разнонаправленно влиять на процессы разложения ОВ торфа и, следовательно, на интенсивность выделения СО2 в атмосферу.
Интенсивность процессов разложения растительных остатков в болотных экосистемах в значительной степени определяется гидротермическим режимом торфяной залежи [6, 39, 42, 48]. Увеличение температуры воздуха и почвы усиливает микробиологическую активность, и соответственно, скорость процесса разложения торфа увеличивается [35, 52]. Вместе с тем рост температуры в сочетании со снижением УБВ способствует ускорению процесса трансформации растительных остатков, в то время как при высоком УБВ роль температуры становится существенно ниже [42]. Наряду с влиянием абиотических факторов, таких как температура и УБВ, который определяет влажность верхней толщи торфяной залежи, скорость деструкции растительных остатков в болотных экосистемах в значительной степени зависит от химического состава самих растений-торфообразователей [6, 16, 29, 31, 32, 48].
Несмотря на несомненную важность влияния температуры и УБВ на скорость разложения торфа, количественные оценки этого влияния, полученные в полевых условиях, весьма малочисленны.
Цель работы заключалась в оценке влияния температуры и УБВ на процессы разложения основных растений-торфообразователей в типичных олиготрофных торфяных почвах (Fibric Histosols) олиготрофных болот южно-таежной подзоны Западной Сибири. Полученные данные представляются важными в свете понимания механизмов устойчивости и продуктивности болотных экосистем и прогноза эволюции болот при изменении климатической и экологической обстановки в регионе.
ОБЪЕКТЫ И МЕТОДЫ
Общая характеристика болотных массивов
Исследования проводили на двух олиготрофных болотах, расположенных в южно-таежной подзоне Западной Сибири: Бакчарском (Бакчарский район Томской области, 56°57′ N, 82°30′ E) и Тимирязевском (Томский район Томской области, 56°26′ N, 84°50′ E). На каждом из болот выбирали по два однотипных фитоценоза: сосново-кустарничково-сфагновый, расположенный на рямовых участках, и осоково-сфагновый, приуроченный к топям (табл. 1). Выбранные для исследования болота являются типичными для территории южно-таежной подзоны Западной Сибири [13, 14, 20]. Верхний слой торфяной залежи низкого ряма Бакчарского болота (1 м) сложен из двух типов сфагновых мхов: магелланикум со средней степенью разложения и фускум с низкой степенью разложения, далее расположен мощный слой переходного торфа двух видов – древесно-осокового и древесно-сфагнового. В основании залежи лежит слой хорошо разложившегося древесно-хвощового и древесного (березового) торфа. Верхний слой торфяной залежи осоково-сфагновой топи (0–100 см) сложен верховыми (ангустифолиум, магелланикум) и сфагново-мочажинным торфами слабой степени разложения. На глубине 100–200 см чередуются слои древесно-осокового, осокового и древесного переходных торфов. В основании залежи 210–290 см лежит слой хорошо разложившегося низинного хвощового торфа, с прослойкой дрепаноклядус-торфа (220–240 см).
Таблица 1.
Характеристика растительного покрова исследуемых болотных экосистем
| Деревья | Кустарнички | Травы | Мхи |
|---|---|---|---|
| Бакчарское болото | |||
| Сосново-кустарничково-сфагновый фитоценоз (низкий рям) – БНР Мощность торфяной залежи 200 см, УБВ (см) средний: –19, макс: –5, мин: –28 |
|||
| P. silvestris f. Litwinowii |
L. palustre L. C. calyculata L. Andromeda polifolia L. Vaccinium uliginosum L. |
E. vaginatum L. R. chamaemorus L. Drosera rotundifolia L. |
Sphagnum fuscum (Schmp.) Klinggr. Sph. angustifolium (Russ.) C. Jens. Sph. magellanicum Brid. |
| (30%) | (65%) | (5%) | (95%) |
| Осоково-сфагновая топь – БТ Мощность торфяной залежи 300 см, УБВ (см) средний: –11, макс: 0, мин: –16 |
|||
| V. oxycoccus L. C. calyculata A. polifolia |
E. vaginatum Carex rostrata Stokes. Carex limosa L. Scheuchzeria palustris L. |
Sph. angustifolium Sph. magellanicum |
|
| (10%) | (50%) | (100%) | |
| Тимирязевское болото | |||
| Сосново-кустарничково-сфагновый фитоценоз (рям) – ТР Мощность торфяной залежи 500 см, УБВ (см) средний: –28, макс: –8, мин: –40 |
|||
| P. silvestris |
L. palustre C. calyculata V. vitis–idea L. V. oxycoccus |
E. vaginatum R. chamaemorus |
Sph. fuscum Sph. angustifolium Sph. magellanicum |
| (90%) | (90%) | (15%) | (96%) |
| Осоково-сфагновая топь – ТТ Мощность торфяной залежи 600 см, УБВ (см) средний: –14, макс: 0 мин: –20 |
|||
| – | – | E. vaginatum C. rostrata C. limosa |
S. angustifolium Sphagnum fallax Klinggr. |
| (50%) | (100%) | ||
Торфяная залежь ряма Тимирязевского болота до глубины 400 см сложена фускум-торфом, далее следуют переходные виды торфа (слой 400–500 см) которые подстилаются низинным торфом. На топи Тимирязевского болота верхний слой торфяной залежи (0–100 см) сложен верховыми (ангустифолиум-торф) и сфагново-мочажинным торфами слабой степени разложения (фаллакс-торф). В настоящее время Тимирязевское болото находится в зоне влияния Томского водозабора, что проявляется в снижении УБВ, обсыхании краевых зон, изменении величины приростов сосны, появлении большого количества зеленых мхов и лишайников в растительном покрове [10, 11]. Бакчарское болото не подвержено заметному антропогенному влиянию, и его можно считать естественно развивающимся. Кроме того, Томский и Бакчарский районы характеризуются разными погодными условиями. Томский район находится южнее, климат здесь более теплый с большим количеством осадков по сравнению с Бакчарским районом (табл. 2). Погодные условия в районе исследований оценивали по данным метеостанций Бакчар и Томск, расположенных в 30 и 10 км от пунктов наблюдения на Бакчарском и Тимирязевском болотах соответственно.
Таблица 2.
Характеристика погодных условий Бакчарского и Томского районов Томской области по данным метеостанций Бакчар и Томск за 2013–2016 гг., среднее значение ± среднее квадратическое отклонение
| Параметр | Метеостанция | |
|---|---|---|
| Бакчар | Томск | |
| Среднегодовая температура воздуха, °С | 0.93 ± 0.93 | 1.80 ± 0.82 |
| Средняя температура воздуха за период вегетации, °С | 13.4 ± 1.04 | 14.2 ± 1.20 |
| Годовая сумма осадков, мм | 544 ± 123 | 639 ± 77 |
| Продолжительность бесснежного периода, дни | 181 ± 14 | 196 ± 5 |
| Сумма активных температур (t > 10°С), °С | 1854 ± 258 | 1953 ± 298 |
| Гидротермический коэффициент Селянинова | 1.02 ± 0.19 | 1.07 ± 0.16 |
| Высота снежного покрова, см | 82 ± 20 | 89 ± 16 |
Определение скорости разложения растений-торфообразователей
Для исследования процессов разложения растений-торфообразователей были выбраны четыре вида-доминанта современного растительного покрова олиготрофных болот. В сосново-кустарничково-сфагновых фитоценозах – это сфагнум бурый (Sphagnum fuscum) и кассандра обыкновенная (Chamaedaphne calyculata), а в осоково-сфагновых – пушица влагалищная (Eriophorum vaginatum) и сфагнум узколистный (Sphagnum angustifolium). Кроме того, выбранные виды сфагновых мхов являются основными растениями-торфообразователями в настоящее время, о чем свидетельствует строение верхних горизонтов торфяных залежей. Пушицевые и кустарничковые виды торфа образуют слои в строении торфяных залежей, образовавшиеся в более ранние периоды развития болотных экосистем [7]. Скорость разложения растений определяли методом частично изолированных проб, который широко используется при изучении процессов трансформации растительных материалов и торфа [6, 9]. С этой целью в сентябре 2013 г. на каждом исследуемом болоте был собран моховой очес сфагновых мхов Sph. fuscum и Sph. angustifolium (10 см от их головок), листья прошлых лет Chamaedaphne calyculata и ветошь Eriophorum vaginatum. Образцы растительных остатков высушивали в лабораторных условиях до воздушно-сухого состояния и помещали в нейлоновые капсулы размером 15 × 15 см. Масса навески растений составляла около 5–6 г. Подготовленные капсулы с растениями закладывали в торфяную залежь на глубину 10 см от поверхности мха (сфагновый очес) в сентябре 2013 г., то есть помещали их в те же условия, в которых произрастали исследуемые растения. Всего было подготовлено 160 капсул (по 40 капсул для каждого вида растительных остатков). Для закладки образцов выбирали участки, максимально приближенные к средней по высоте поверхности болота, для того, чтобы все образцы находились в равных условиях относительно УБВ. То есть на каждом объекте УБВ для всех образцов одинаков. В мае и сентябре 2014 г., а также в сентябре 2015 и 2016 гг. (соответственно через 8, 12, 24, 36 мес. после начала эксперимента) из торфяной залежи извлекали по 10 капсул с каждым видом растительного материала, и затем в лабораторных условиях определяли абсолютно-сухую массу растительного образца. Скорость разложения растительных остатков оценивали, определяя в динамике уменьшение массы растительных образцов.
Потерю массы (ПМ, % от исходной массы растительного образца) рассчитывали в по формуле:
(1)
${\text{П М }}(\% ) = \frac{{{{M}_{{\text{0}}}} - {{M}_{t}}}}{{{{M}_{{\text{0}}}}}} \times 100\% ,$Предполагалось, что процесс разложения растительных остатков происходит по экспоненциальному закону [47]:
где k – константа разложения, год–1; t – время разложения, лет.Исходя из уравнения (2), константу разложения можно рассчитать по формуле:
Для оценки изменения содержания общего С и общего N в исходных образцах исследуемых растений использовали метод одновременного определения общего содержания С и N в торфяных почвах методом Анстета в модификации Пономаревой и Николаевой [28]. Анализы проводили в трехкратной повторности.
Контроль гидротермических параметров во время эксперимента
Температуру торфяной залежи измеряли с помощью автономных измерителей профиля температуры (АИПТ, ИМКЭС СО РАН [17]). Датчики температуры располагали на глубине 15 см от поверхности, временной шаг измерений составлял 15 мин. Данные затем осредняли до среднесуточных значений и на их основании рассчитывали сумму температур выше 0, 5, 10°С на глубине 15 см для каждого вегетационного периода (2013–2016 гг.). На Бакчарском болоте УБВ определяли с помощью дифференциального измерителя уровня болотных вод HOBO Water Level Logger U20-001-01 (США). На Тимирязевском болоте контроль за УБВ вели с помощью датчиков уровня болотных вод, которыми оснащены автономные измерители профиля температуры АИПТ, ИМКЭС СО РАН [17]. Измерения УБВ проводили в автоматическом режиме с периодичностью 1 ч круглогодично. Эти данные затем осредняли до среднесуточных значений, на основании которых рассчитывали средние и минимальные УБВ для каждого вегетационного периода. Чтобы привязать данные по УБВ, полученные с помощью автоматических датчиков, во время экспедиционных работ проводили измерения УБВ по общепринятой методике с использованием водомерных скважин, оборудованных на измерительных пунктах [21].
Статистическую обработку данных (расчет средних значений, стандартных ошибок, коэффициентов корреляции Спирмена) осуществляли с помощью программы Statistica6. Достоверность различий для скорости разложения между разными видами растений оценивали по критерию Стьюдента, при уровне значимости Р = 0.05. Влияние гидротермических условий на скорость разложения растительных остатков оценивали с помощью однофакторного дисперсионного анализа.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Характеристика гидротермических условий за время проведения эксперимента
Гидрологический режим исследуемых болот существенно различался (рис. 1, табл. 3). В результате влияния Томского водозабора на Тимирязевском болоте произошло снижение УБВ [6]. Так, в сосново-кустарничково-сфагновом фитоценозе Тимирязевского болота УБВ в течение вегетационного периода (за исключением периода снеготаяния) в среднем был на 15 см ниже по сравнению с Бакчарским болотом, а на осоково-сфагновой топи эта разница составляла лишь 5 см.
Рис. 1.
Средние значения УБВ в течение вегетационного периода на Бакчарском и Тимирязевском болотах в 2013–2016 гг.: БНР – Бакчарское болото, низкий рям; БТ – Бакчарское болото, осоково-сфагновая топь; ТР – Тимирязевское болото, рям; ТТ – Тимирязевское болото, осоково-сфагновая топь.
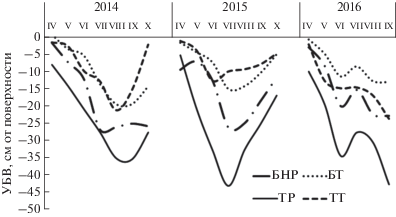
Таблица 3.
Показатели гидротермического режима торфяных залежей в слое 0–20 см в различных болотных экосистемах на Бакчарском и Тимирязевском болотах за 2013–2016 гг., среднее значение ± среднее квадратическое отклонение
| Показатель | Бакчарское болото | Тимирязевское болото | ||
|---|---|---|---|---|
| БНР* | БТ | ТР | ТТ | |
| Среднегодовая температура, °С | 5.07 ± 0.62 | 6.65 ± 0.98 | 5.42 ± 1.31 | 7.00 ± 1.44 |
| Средняя температура за период вегетации, °С | 11.61 ± 0.86 | 14.21 ± 1.31 | 12.56 ± 1.01 | 13.90 ± 0.82 |
| Средний за вегетационный период УБВ, см | –19 ± 4 | –11 ± 3 | –28 ± 7 | –14 ± 3 |
| Средний минимальный УБВ, см | –28 ± 5 | –16 ± 3 | –40 ± 4 | –20 ± 4 |
Средняя влажность торфа, в который были заложены образцы (слой 5–10 см), в сосново-кустарничково-сфагновых рямах на Тимирязевском болоте составляла 85% от полной полевой влагоемкости, а на Бакчарском – 95%. Влажность торфа осоково-сфагновых топей на обоих болотах равнялась 95%. Изменения влажности торфа в слое 5–10 см в течение вегетационного периода были незначительны из-за большой водоудерживающей способности сфагнового торфа. Максимальные значения влажности наблюдались в весенний период, после снеготаяния, а также после выпадения осадков. Поэтому в работе основной акцент сделан на оценке влияния УБВ.
Температурный режим торфяных залежей значимо не различался. В среднем за исследуемый период (сентябрь 2013–сентябрь 2016) самым холодным был верхний (0–20 см) слой торфяной залежи сосново-кустарничково-сфагнового фитоценоза Бакчарского болота (табл. 3). Среднегодовая температура на глубине 15 см за весь период исследования была здесь на 0.35°С ниже, чем на участке с аналогичной растительностью Тимирязевского болота. Среднегодовая температура участков на осоково-сфагновой топи была на 1.5°С выше, чем на рямах, при этом топь Бакчарского болота также была в среднем на 0.35°С холоднее по сравнению с топью Тимирязевского болота. В течение вегетационного периода (апрель–сентябрь) разница температур между Бакчарским и Тимирязевским болотами в среднем составляла 0.95°С – на рямах и 0.31°С – на топях. Топи прогревались сильнее чем рямы, и разница средних температур в период вегетации между ними достигала 2.60 и 1.34°С – на Бакчарском и Тимирязевском болотах соответственно.
Разложение растительных остатков. Растительный материал, который подвергался разложению, сильно различался по содержанию С, N и величине С/N (табл. 4). Самое высокое содержание биогенных элементов и самое узкое отношение С/N было характерно для Chamaedaphne calyculata. Сфагновые мхи (Sph. fuscum и Sph. angustifolium) отличались самым низким содержанием С и N в своем составе при весьма существенной разнице в величине отношения С/N.
Таблица 4.
Содержание углерода, азота и величина C/N в растениях-торфообразователях (n = 3), среднее значение ± среднее квадратическое отклонение
| Растение-торфообразователь | Содержание элементов, % | C/N | |
|---|---|---|---|
| С | N | ||
| Chamaedaphne calyculata | 48.8 ± 0.13 | 1.62 ± 0.02 | 30 |
| Sph. fuscum | 40.7 ± 0.11 | 0.58 ± 0.01 | 70 |
| Eriophorum vaginatum | 43.9 ± 0.26 | 0.81 ± 0.01 | 54 |
| Sph. angustifolium | 35.9 ± 0.14 | 0.68 ± 0.04 | 53 |
Независимо от химического состава растений-торфообразователей, в процессе их разложения самая значительная потеря массы наблюдалась в течение первого года исследований (рис. 2). На Тимирязевском болоте на топяных участках потери массы за первый год исследований составляли в зависимости от вида растений 17–29% от первоначальной массы, а на Бакчарском – от 8.5 до 26%. Самая низкая потеря массы была характерна для Sph. fuscum, а самая высокая – для Chamaedaphne calyculata на Бакчарском болоте и Sph. angustifolium – на Тимирязевском. Высокие скорости разложения на начальном этапе эксперимента объясняются, по-видимому, тем, что в свежем растительном опаде содержится много водорастворимых и легкогидролизуемых веществ, которые потребляются микроорганизмами-деструкторами в первую очередь. В ходе разложения содержание легкодоступных компонентов углеводного и полипептидного комплексов убывает, и соответственно уменьшаются потери органического вещества [3, 31, 36, 40].
Рис. 2.
Потери массы при разложении растительных остатков на Бакчарском и Тимирязевском болотах за 36 мес. эксперимента.

В течение второго года эксперимента для всех исследуемых видов растений было характерно снижение темпов потери массы, за исключением образцов Chamaedaphne calyculata на Тимирязевском болоте, для которой скорость убыли массы сохранилась на том же уровне (~29–30%). За три года эксперимента минимальные потери массы среди исследуемых растений были получены для Sph. fuscum – 20 и 25% от исходного веса для Бакчарского и Тимирязевского болот соответственно. Chamaedaphne calyculata характеризовалась максимальными потерями массы на каждом из болот, составляющими 46–62% от исходного значения. Результаты исследований показали также, что более медленно процесс деструкции растительных остатков протекал во влажных условиях Бакчарского болота, где потери массы для всех исследуемых видов за исключением опада Eriophorum vaginatum были в 1.2–1.6 раз ниже, чем на Тимирязевском болоте. В соответствии с результатами однофакторного дисперсионного анализа, потери массы растений, в зависимости от видовой принадлежности, за 3 года исследований достоверно отличались для образцов Sph. fuscum и Chamaedaphne calyculata (F = 10.6, Р = 0.005), а также для двух видов сфагнума – бурого и узколистного (F = 10.73, P = 0.005).
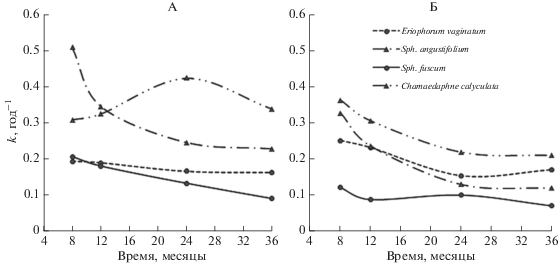
Значения константы разложения одних и тех же видов растений и их временная динамика в разных болотах существенно отличались (рис. 3). На Тимирязевском болоте самые низкие константы разложения и постепенное снижение от 0.20 до 0.17–0.10 год–1 за все время эксперимента показывали Sph. fuscum и Eriophorum vaginatum (рис. 3, А). Chamaedaphne calyculata (Тимирязевское болото) характеризовалась максимальной скоростью разложения (0.42 год–1) в течение второго года эксперимента, которая в конце третьего года разложения уменьшилась до значений, характерных для начальной стадии эксперимента (рис. 3, А). На Бакчарском болоте константы разложения для всех видов изученных растений были ниже, чем на Тимирязевском и не превышали 0.40 год–1 (рис. 3, Б). В течение трех лет эксперимента значения k уменьшались неравномерно. Так, константа разложения пушицы на Бакчарском болоте резко (почти в 2 раза) уменьшалась в течение первых двух лет разложения и практически не менялась в течение третьего года эксперимента (рис. 3, Б). Наиболее значительным снижение скорости разложения было в образцах Eriophorum vaginatum (от 0.33 до 0.11 год–1), а наименее заметным – в образцах Sph. fuscum (от 0.11 до 0.08 год–1).
Рис. 3.
Изменение константы разложения растительных остатков в течение 36 мес. эксперимента: А – Тимирязевское болото, Б – Бакчарское болото.
Результаты проведенного эксперимента в целом соответствуют данным, полученным ранее для территории Западной Сибири [23, 24]. В зависимости от вида сфагнового мха потери массы при их разложении в течение двух лет составляли от 10 до 30%, а наибольшими потерями массы характеризовался Sph. angustifolium. Кроме того, в этих работах [23, 24] показано влияние УБВ на процесс разложения: образцы, расположенные в торфяной залежи на грядах грядово-мочажинных комплексов (в более сухих условиях), разлагались быстрее, чем в более влажных условиях мочажин, что также соответствует полученным нами данным. Для всех видов мхов константы разложения растительных остатков, как правило, были максимальны на начальных этапах эксперимента. Однако при общей тенденции снижения их величин с увеличением времени экспозиции, в ходе экспериментов наблюдались колебания, обусловленные условиями, в которых протекало разложение [6, 24, 45].
Известно, что скорость разложения растительных остатков в значительной степени зависит от таких, свойств как концентрация азота, соотношение C/N, содержание лигнина и целлюлозы [19, 27, 33, 49, 56]. Невысокая скорость разложения сфагнума бурого (Sph. fuscum) была, по-видимому, обусловлена особенностями его биохимического состава. Так, в сфагновых мхах присутствуют специфические фенольные соединения – сфагнолы, которые препятствуют бактериальному разложению органического вещества [25, 31, 37]. Значение имеет состав полисахаридов клеточных стенок сфагновых мхов, который также ингибирует процесс разложения [40, 50]. Суммарное количество аминокислот и азотсодержащих соединений в составе растений, о содержании которых можно косвенно судить по концентрации общего азота и отношению C/N, также оказывает существенное влияние на процессы разложения растительных остатков [25, 31]. Однако до сих пор не существует однозначного мнения по поводу влияния соединений азота на процессы минерализации углерода растительных органических материалов [19]. Низкие концентрации азота в растениях, как правило, приводят к снижению скорости трансформации растительных остатков. Наименьшим отношением C/N и самым высоким содержанием азота отличается Chamaedaphne calyculata и соответственно, максимальные потери массы в нашем эксперименте были получены для растительных остатков Chamaedaphne calyculata. В то же время потери массы Sph. fuscum были минимальными при минимальном содержании азота и максимальной величине C/N.
Вместе с тем сведения, имеющиеся в литературе часто противоречивы. Сообщается о том, что увеличение концентрации азота в растительном опаде может как ускорять, так и замедлять процессы разложения [19, 41, 43]. В некоторых исследованиях влияние азота на скорость разложения растительных остатков либо не выявлялось, либо показывало очень слабую положительную зависимость [43, 56]. В нашем эксперименте наибольшие константы разложения были характерны для Chamaedaphne calyculata, имеющей самое узкое отношение С/N = 30, а наименьшая скорость разложения наблюдалась у Sph. fuscum, имеющего самое высокое отношение С/N = 70. Таким образом, для изученных растений-торфообразователей наблюдалась четкая отрицательная корреляция между средними значениями константы разложения и отношением С/N (R2 = 0.64, Р = 0.02).
Влияние гидротермических условий на скорость разложения растительных остатков
Наряду с особенностями состава растительных остатков, не менее важными факторами, влияющими на скорость их разложения, являлись температура и влажность торфяной залежи, которая в существенной степени определялась УБВ. В сосново-кустарничково-сфагновых фитоценозах исследуемых болот, где разложению подвергались Sph. fuscum и Chamaedaphne calyculata, УБВ существенно различался. В осоково-сфагновых топях, где закладывали на разложение Eriophorum vaginatum и Sph. angustifolium, разница УБВ была не столь существенна (рис. 1). Проведенные исследования показали, что образцы растений, помещенные в торфяную залежь Тимирязевского болота с более глубоким УБВ, разлагаются быстрее по сравнению с образцами, заложенными в более влажный торф Бакчарского болота. Так, на Тимирязевском болоте в сосново-кустарничково-сфагновом фитоценозе, капсулы с растениями в течение всего вегетационного периода находились выше УБВ, тогда как на Бакчарском болоте капсулы с растительным материалом практически все время располагались либо в водоносном слое (ниже УБВ), либо – на границе УБВ, периодически попадая то в аэробную, то в анаэробного зону.
Выявлено, что влажность субстрата, в котором протекают процессы разложения растительных остатков, оказывает заметное влияние на их скорость, регулируя доступность питательных элементов для микроорганизмов-деструкторов [1, 8, 18, 26]. Результаты корреляционного анализа показали, что и средний, и минимальный УБВ (за период вегетации) оказывали достоверное влияние на потери массы растительных остатков всех исследуемых видов растений за исключением Sph. angustifolium (табл. 5). Наиболее тесная отрицательная связь между скоростью разложения и УБВ наблюдалась для образцов Chamaedaphne calyculata (рис. 4), в то время как более интенсивное разложение растительных остатков Eriophorum vaginatum наблюдалось при высоких значениях УБВ. В целом, в болотных экосистемах повышенная влажность, как правило, замедляет деструкцию растительных остатков, что ведет к их консервации и росту торфяной залежи [14, 15]. В более ранних исследованиях [6, 24, 48] также отмечалось, что, как правило, при снижении УБВ наблюдается увеличение интенсивности процессов разложения органического вещества, за счет увеличения зоны аэрации и активизации аэробных микроорганизмов.
Таблица 5.
Коэффициенты корреляции* между потерями массы растительных остатков и гидротермическими условиями (n = 8)
| Растение-торфообразователь | УБВ за вегетацию | Ʃt | |||
|---|---|---|---|---|---|
| средний | минимальный | >0 °C | >5°C | >10°C | |
| Eriophorum vaginatum | 0.58 | 0.87 | 0.81 | 0.79 | 0.73 |
| Sph. angustifolium | –0.18 | 0.23 | 0.39 | 0.37 | 0.32 |
| Сhamaedaphne calyculata | –0.46 | –0.40 | 0.79 | 0.76 | 0.77 |
| Sph. fuscum | –0.40 | –0.28 | 0.77 | 0.74 | 0.75 |
Рис. 4.
Зависимость потери массы растительных остатков от суммы температур >0°C на глубине 10 см (А) и среднего значения УБВ за вегетационный период (Б). Все регрессионные зависимости значимы при P < 0.05.
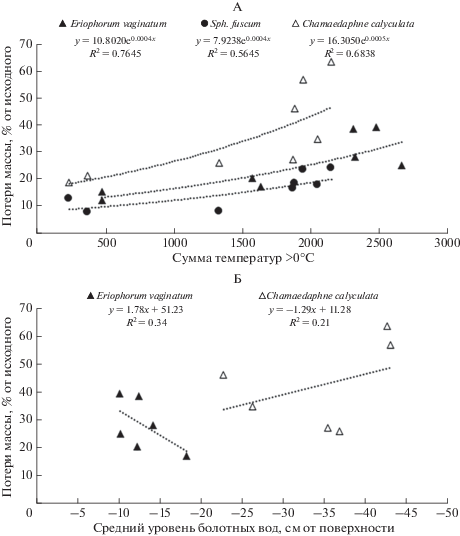
Для всего массива данных были выявлены положительные корреляционные связи между величиной потери массы растительных остатков и суммами температур торфяной залежи, превышающими 0, 5 и 10°С. Показано, что сумма температур в течение вегетационного периода оказывала достоверное влияние на скорость разложения всех изученных видов растительных остатков за исключением Sphagnum angustifolium, для которого коэффициенты корреляции были не значимы при P < 0.05.
Таким образом, результаты длительного полевого эксперимента показали, что температура торфяной залежи и УБВ, наряду с особенностями биохимического состава растений оказывали значимое влияние на скорость разложения растительных остатков. Однако сочетание этих факторов может неоднозначное влиять на деструкционные процессы ОВ. В исследованиях Домиш с соавт. [35] показано, что при повышении температуры усиливалась активность микроорганизмов-деструкторов и активность процессов разложения увеличивалась. Кроме того, с ростом температуры, в результате потепления климата, чувствительность микроорганизмов к температурному фактору была больше в северных регионах, и интенсивность процесса разложения увеличивалась здесь еще сильнее. Наиболее явно влияние температуры и влажности проявлялось на начальных стадиях разложения, когда происходила деструкция лабильных компонентов органического вещества [18]. Выявленные в настоящем исследовании закономерности влияния гидротермического режима на скорость разложения растительных остатков могут быть дополнены модельными экспериментами, проводимыми в контролируемых лабораторных условиях. Это позволит получить более детальную оценку влияния абиотических факторов (температуры, влажности) на процессы деструкции растений торфообразователей.
ВЫВОДЫ
1. Получены количественные оценки скорости разложения четырех видов современных растений-торфообразователей, типичных для болотных фитоценозов южно-таежной подзоны Западной Сибири: сосново-кустарничково-сфагновых (Sphagnum fuscum, Chamaedaphne calyculata) и осоково-сфагновых (Eriophorum vaginatum и Sphagnum angustifolium), в течение 36 мес. полевого эксперимента. Наибольшей устойчивостью к разложению характеризовались образцы Sph. fuscum, потери массы которого за три года эксперимента составили 18–24%. Наименьшую устойчивость к разложению проявляли растительные образцы Chamaedaphne calyculata с максимальными потерями массы – 46–62%. Константы скорости разложения достоверно различались у Sph. fuscum и Chamaedaphne calyculata и Sph. fuscum и Sph. angustifolium.
2. Наиболее интенсивная деструкция растительных остатков наблюдалась в первые 8–12 мес. эксперимента. Затем скорость разложения органического вещества оставалась либо на том же уровне, либо снижалась (в 1.2–2.8 раза). Скорость разложения растительных остатков во многом определялась содержанием в них общего азота и отношением C/N. Максимальная скорость разложения Chamaedaphne calyculata среди исследуемых видов, вероятно, была связана с самым узким отношением C/N и самым высоким содержанием N в составе этого растения. Скорость разложения Sph. fuscum, характеризующегося минимальным содержанием азота и максимальной величиной C/N, была самой низкой в ряду изученных видов растений.
3. Интенсивность процессов разложения растительных остатков в значительной степени определялась гидрологическими условиями вегетационного периода. В более влажных условиях Бакчарского болота интенсивность разложения растительных остатков была в 1.3–1.9 раз ниже, чем в более сухом Тимирязевском болотном массиве. Наиболее чувствительны к изменению уровня болотных вод были Chamaedaphne calyculata и Eriophorum vaginatum.
4. Температурный режим торфяной залежи также оказывал существенное влияние на скорость процессов разложения растительных остатков, причем значимая корреляционная связь для большинства видов изученных растений была выявлена между потерями массы и суммой температур выше 0°C, что косвенно свидетельствует о жизнедеятельности микроорганизмов-деструкторов при низких температурах. Наиболее тесная корреляция величины потери массы с суммами температур в верхнем слое торфяной залежи наблюдалась для образцов Eriophorum vaginatum и Сhamaedaphne calyculata.
БЛАГОДАРНОСТЬ
Авторы выражают благодарность Е.А. Дюкареву и С.В. Смирнову за помощь в организации и проведении экспедиционных исследований.
ФИНАНСИРОВАНИЕ РАБОТЫ
Работа выполнялась при частичной финансовой поддержке РФФИ (проект № 17-34-50041), Программы Президиума РАН № 51 “Изменение климата: причины, риски, последствия, проблемы адаптации и регулирования” и рамках госбюджетных заданий № АААА-А17-117013050031-8 и АААА-А18-118013190177-9.
Список литературы
Авров О.Е. Влияние температуры и влажности на разложение соломы // Использование соломы как органического удобрения. М.: Наука, 1980. С. 103–113.
Бамбалов Н.Н. Баланс органического вещества торфяных почв и методы его изучения. Минск: Наука, 1984. 175 с.
Бамбалов Н.Н., Хоружик А.В., Лукошко Е.С., Стригуцкий В.П. Превращение отмерших растений в болотных биогеоценозах // Эксперимент и математическое моделирование в изучении биогеоценозов лесов и болот. М.: Наука, 1990. С. 53–63.
Вомперский С.Э. Роль болот в круговороте углерода // Биогеоценотические особенности болот и их рациональное использование. М.: Наука, 1994. С. 5–37.
Второй оценочный доклад Росгидромета об изменениях климата и их последствиях на территории Российской Федерации. М.: Росгидромет, 2014. 1004 с.
Головацкая Е.А., Никонова Л.Г. Влияние уровня болотных вод на процессы трансформации сфагновых мхов в торфяной почве олиготрофных болот // Почвоведение. 2017. № 5. С. 606–613. https://doi.org/10.7868/S0032180X17030030
Головацкая Е.А. Потоки углерода в болотных экосистемах Западной Сибири: дис. … д-ра биол. наук. Томск, 2013. 326 с.
Гришкан И.Б. Разложение растительного опада в основных сообществах бассейна верховий Колымы // Экология. 1995. № 4. С. 9–12.
Дементьева Т.В., Богданова О.Ю., Шинкеева Н.А. Физикохимия и биология торфа // Руководство по методам изучения трансформации органического вещества торфов: методическое пособие. Томск, 2011. 68 с.
Дюкарев А.Г., Пологова Н.Н., Базанов В.В., Льготин В.А., Свечников Д.А., Чернова Н.А., Горбов Е.В. Состояние природной среды в зоне действия Томского водозабора // Тр. междунар. конф. E-NVIROMIS-2002. Томск, 2002. Т. 2. С. 244–251.
Дюкарев А.Г., Пологова Н.Н. Водный режим почв в зоне влияния Томского водозабора // Вестник ТГУ. 2009. № 324. С. 363–371.
Ефимов В.Н. Торфяные почвы и их плодородие. Ленинград: Агропромиздат, 1986. 264 с.
Инишева Л.И. Болота Западной Сибири. Новосибирск: Принтинг, 2007. 60 с.
Инишева Л.И., Кобак К.И., Инишев Н.Г. Заболачивание на Васюганском Болоте // Сибирский экологический журнал. 2017. № 2. С. 119–126.
Инишева Л.И., Шайдак Л., Сергеева М.А. Динамика биохимических процессов и окислительно-восстановительных условий в геохимически сопряженных ландшафтов олиготрофных болот // Почвоведение. 2016. № 4. С. 505–513. https://doi.org/10.7868/S0032180X16040055
Козловская Л.С., Медведева В.М., Пьявченко Н.И. Динамика органического вещества в процессе торфообразования. Ленинград: Наука, 1978. 172 с.
Кураков С.А., Крутиков В.А., Ушаков В.Г. Автономный измеритель профиля температуры АИПТ // Приборы и техника эксперимента. 2008. № 5. С. 166–167.
Ларионова А.А., Мальцева А.Н., Лопес де Гереню В.О., Квиткина А.К., Быховец С.С., Золотарева Б.Н., Кудеяров В.Н. Влияние температуры и влажности на минерализацию и гумификацию лиственного опада в модельном инкубационном эксперименте // Почвоведение. 2017. № 4. С. 438–448. https://doi.org/10.7868/S0032180X17020083
Ларионова А.А., Квиткина А.К., Быховец С.С., Лопес-де-Гереню В.О., Колягин Ю.Г., Каганов В.В. Влияние азота на минерализацию и гумификацию лесных опадов в модельном эксперименте // Лесоведение. 2017. № 2. С. 128–139.
Лисс О.Л., Абрамова Л.И., Аветов Н.А., Березина Н.А., Инишева Л.И., Курнишкова Т.В., Слука З.А., Толпышева Т.Ю., Шведчикова Н.К. Болотные системы Западной Сибири и их природоохранное значение. Тула: Гриф и К°, 2001. 584 с.
Наставления гидрометеорологическим станциям и постам. Ленинград: Гидрометеоиздат, 1989. 201 с.
Оценочный доклад об изменениях климата и их последствиях на территории Российской Федерации // Изменения климата. М.: Росгидромет, 2008. Т. 1. 227 с.
Паршина Е.К. Разложение растительного вещества в лесотундре // Сиб. экол. журн. 2007. № 5. С. 781–787.
Паршина Е.К. Деструкция растительного вещества в болотных экосистемах таежной и лесотундровой зон Западной Сибири: Автореф. дис. … канд. биол. наук. Томск, 2009. 23 с.
Раковский В.Е., Пигулевская Л.В. Химия и генезис торфа. М.: Недра, 1987. 231 с.
Рахно П.Х. Микробиология. М.: Наука, 1960. Т. 2. № 2. С. 229–233.
Семенов В.М., Иванникова Л.А., Кузнецова Т.А., Семенова Н.А. Роль растительной биомассы в формировании активного пула органического вещества почвы // Почвоведение. 2007. № 11. С. 1350–1359.
Теория и практика химического анализа почв / Под ред. Воробьевой Л.А. М.: ГЕОС, 2006 г. 400 с.
Теппер Е.З., Комаревцева Л.Т., Симакова Н.С. Процессы минерализации растительных остатков, характерных для подзоны Южной тайги // Известия ТСХА. 1975. В. 1. С. 121–126.
Тюремнов С.Н. Торфяные месторождения. М.: Недра, 1976. 487 с.
Функционирование микробных комплексов в верховых торфяниках – анализ причин медленной деструкции торфа. М.: Товарищество научных изданий КМК, 2013. 128 с.
Aerts R. Climate, Leaf Litter Chemistry and Leaf Litter Decomposition in Terrestrial Ecosystems: A Triangular Relationship // Oikos. 1997. V. 79. P. 439–449. https://doi.org/10.2307/3546886
Berg B. Decomposition patterns for foliar litter: A theory for influencing factors // Soil Biol.Biochemi. 2014. V. 78. P. 222–232. https://doi.org/10.1016/j.soilbio.2014.08.005
Bolin B. How Much CO2 will remain in the Atmosphere? // The greenhouse effect, climatic change, and ecosystems. 1986. P. 93–155.
Domisch T., Finér L., Laine J., Laiho R. Decomposition and nitrogen dynamics of litter in peat soils from two climatic regions under different temperature regimes // Europ. J. Soil Biol. 2006. V. 42. P. 74–81. https://doi.org/10.1016/j.ejsobi.2005.09.017
Fioretto A., Di Nardo C., Papa S., Fuggi A. Lignin and cellulose degradation and nitrogen dynamics during decomposition of three leaf litter species in a Mediterranean ecosystem // Soil Biol. Biochem. 2005. V. 37. P. 1083–1091. https://doi.org/10.1016/j.soilbio.2004.11.007
Freeman C., Ostle N.J., Fenner N., Kang H. A regulatory role for phenol oxidase during decomposition in peatlands // Soil Biol. Biochem. 2004. V. 36. P. 1663–1667. https://doi.org/10.1016/j.soilbio.2004.07.012
Gorham E. Northern Peatlands: role in carbon cycle and probable responses to climatic warming // Ecological Appl. 1991. V. 1. P. 182–195. https://doi.org/10.2307/1941811
Guo X., Lu X., Tong S., Dai G. Influence of environment and substrate quality on the decompostion of wetland plant root in the Sanjiang Plain, Northeast China // J. Environ. Sci. 2008. V. 20. P. 1445–1452. https://doi.org/10.1016/S1001-0742(08)62547-4
Hajek T., Ballance S., Limpens J., Zijlstra M., Verhoeven J.T.A. Cell-wall polysaccharides play an important role in decay resistance of Sphagnum and actively depressed decomposition in vitro // Biogeochemistry. 2011. V. 103. P. 45–57. https://doi.org/10.1007/s10533-010-9444-3
Hobbie S., Eddy W., Buyarski C., Adair C., Ogdahl M., Weisenhorn P. Response of decomposing litter and its microbial community to multiple forms of nitrogen enrichment // Ecological Monographs. 2012. V. 82. P. 389–405. https://doi.org/10.1890/11-1600.1
Hogg E.H., Lieffers V.J., Wein R.W. Potential carbon losses from peat Profiles: Effects of Temperature, Drought Cycles, and Fire // Ecological Appl. 1992. V. 2. № 3. P. 298–306.
Knorr M., Frey S.D., Curtis P.S. Nitrogen additions and litter decomposition: a meta-analysis // Ecology. 2005. V. 86. P. 3252–3257. https://doi.org/10.1890/05-0150
Limpens J., Berendse F., Blodau C., Canadell J.G., Freeman C., Holden J., Roulet N., Rydin H., Schaepman-Strub G. Peatlands and the carbon cycle: from local processes to global implications – a synthesis // Biogeosciences. 2008. V. 5. P. 1475–1491. https://doi.org/10.5194/bg-5-1475-2008
Moor T.R., Trofimow J.U.A., Prescott C.E., Titus B.D. Can short-term litter-bag measurements predict long-term decomposition in northern forests? // Palnt and soil. 2017. V. 416. P. 419–426. https://doi.org/10.1007/s11104-017-3228-7
Nilsson M., Sagerfors J., Buffam I., Laudon H., Eriksson T., Grelle A., Klemedtsson L., Weslien P., Lindroth A. Contemporary carbon accumulation in a boreal oligotrophic minerogenic mire – A significant sink after accounting for all C-fluxes // Global Change Biol. 2008. V. 14. P. 2317–2332. https://doi.org/10.1111/j.1365-2486.2008.01654.x
Olson J.S. Energy storage and the balance of producers and decomposers in ecological systems // Ecology. 1963. V. 44. P. 322–331. https://doi.org/10.2307/1932179
Peltoniemi K., Strakova P., Fritze H., Iraizoz P.A., Pennanen T., Laiho R. How water-level drawdown modified litter-decomposing fungal and actinobacterial communities in boreal peatlands // Soil Biol. Biochem. 2012. V. 51. P. 20–34. https://doi.org/10.1016/j.soilbio.2012.04.013
Prescott C. Litter decomposition: what controls it and how can we alter it to sequester more carbon in forest soils // Biogeochemistry. 2010. V. 101. P. 133–149. https://doi.org/10.1007/s10533-010-9439-0
Turetsky M.R., Crow S.E., Evans R.J., Vitt D.H., Wieder R.K. Trade-offs in resource allocation among moss species control decomposition in boreal peatlands // J. Ecology. 2008. V. 96. P. 1297–1305. https://doi.org/10.1111/j.1365-2745.2008.01438.x
Turunen J., Tomppo E., Tolonen K., Einikainen A. Estimating carbon accumulation rates of undrained mires in Finland – application to boreal and subarctic regions // Holocene. 2002. V. 12. P. 69–80.
Vanhala P., Karhu K., Tuomi M. Björklöf K., Fritze H., Liski J. Temperature sensitivity of soil organic matter decomposition in southern and northern areas of the boreal forest zone // Soil Biol. Biochem. 2008. V. 40. P. s1758–1764. https://doi.org/10.1016/j.soilbio.2008.02.021
Vygodskaya N.N., Groisman P.Ya., Tchebakova N.M., Kurbatova J.A., Panfyorov O., Parfenova E.I., Sogachev A.F. Ecosystems and climate interactions in the boreal zone of northern Eurasia // Environ. Res. Lett. 2007. V. 2. P. 45033.
Wieder R.K., Canadell J., Limpens J., Moore T., Roulet N., Schaepman-Strub G.: Peatlands and the carbon cycle: From local processes to global implications // EOS. 2007. V. 88. P. 295. https://doi.org/10.1029/2007EO290005
Yu Z., Beilman D.W., Frolking S., MacDonald G.M., Roulet N.T., Camill P., Charman D.J. Peatlands and Their Role in the Global Carbon Cycle // EOS. 2011. V. 92. P. 97–98. https://doi.org/10.1029/2011EO120001
Zhang D., Hui D., Luo Y., Zhou G. Rates of litter decomposition in terrestrial ecosystems: global patterns and controlling factors // J. Plant Ecology. 2008. V. 1. P. 85–93. https://doi.org/10.1093/jpe/rtn002
Дополнительные материалы отсутствуют.