

Почвоведение, 2020, № 1, стр. 31-43
Оценка внутрибиогеоценозной изменчивости лесных подстилок и травяно-кустарничковой растительности в еловых насаждениях
О. В. Семенюк a, В. М. Телеснина a, *, Л. Г. Богатырев a, А. И. Бенедиктова a, Я. Д. Кузнецова a
a МГУ им. М.В. Ломоносова
11999 Москва, Ленинские горы, 1, Россия
* E-mail: vtelesnina@mail.ru
Поступила в редакцию 15.03.2019
После доработки 05.06.2019
Принята к публикации 20.06.2019
Аннотация
Выявлены различия в видовом составе живого напочвенного покрова ельника кислично-зеленчукового в зависимости от положения в тессере, под которой понимается совокупность приствольного, подкронового и межкронового пространства, различающихся локальными экологическим условиям из-за воздействия ели как эдификатора. От приствольных пространств к межкроновым в составе напочвенного покрова увеличивается разнообразие экологических свит, общее число видов, долевое участие неморальных, светолюбивых и требовательных к общему содержанию элементов питания видов. Мощность и запасы лесных подстилок уменьшаются от приствольных пространств к окнам. Положение в тессере определяет фракционный состав подстилок, а также соотношение величин запасов горизонтов внутри подстилки. Подстилки приствольных участков характеризуются минимальными значениями рН и минимальными запасами зольных элементов в горизонтах L, а также в легкоразлагаемых компонентах, что соотносится с высоким долевым участием ацидофильных видов и видов, мало требовательных к запасам элементов питания. Показатели, характеризующие скорость разложения органического вещества (доля легкоразлагаемых компонентов в горизонте L подстилки, запасы зольных элементов в подстилке и ее легкоразлагаемых фракциях, соотношение мощностей и запасов горизонтов L, F и Н), напрямую связаны с соотношением в напочвенном покрове видов, характеризующихся разным уровнем трофности, установленным по экологическим шкалам. Ель, как эдификатор, создает в разных компонентах тессеры значимые различия в морфологических особенностях и химических свойствах лесных подстилок, что адекватно отражается видовым составом и эколого-ценотической структурой напочвенного покрова.
ВВЕДЕНИЕ
Подстилка является важнейшим компонентом биогеоценоза и интегральным показателем биологического круговорота, отражает основные направления функционирования экосистем, в своем строении и свойствах подчиняется биоклиматической зональности [7, 58]. Лесные подстилки обусловлены существованием древостоя и являются отражением факторов окружающей среды, основной индикаторной системой лесного биогеоценоза, местообитанием многих видов почвенной фауны, важным источником углекислоты и азота [12–14]. Занимая промежуточное положение между поступившим опадом и собственно почвой и являясь источником органических соединений и резервуаром биогенных элементов, подстилка признана неотъемлемым фактором, формирующим лесные почвы, поэтому ее строение, запасы и химический состав чрезвычайно важны для понимания механизмов развития органопрофиля почв [4, 6, 8, 23, 35]. Сочетая в себе высокую информативность по функционированию биогеоценоза и высокую доступность для наблюдения, подстилки являются важнейшим объектом для изучения [2, 9, 22, 37].
В исследовании лесных подстилок за рубежом в последние годы сформировалось несколько направлений. Первое – традиционное изучение влияния на подстилки различных факторов, например, пожаров [71, 72]. Анализируются процессы переноса наземного детрита с последующей их аккумуляцией в смежных экосистемах, в том числе водных [51]. Уделяется внимание гидрологическим характеристикам подстилок, в том числе специфике удержания ими влаги [57]. Установлено, что со временем содержание ферментов в подстилках возрастает [61], а обогащение подстилки микро- и макроэлементами связано с поступлением их из почвы [65]. Обращается внимание на роль калия в процессах разложения, которая ранее игнорировалась [66, 74]. Проводится анализ пространственного распределения подстилок и запасов углерода в рамках решения глобальных проблем [52]. Возрастает интерес к формированию подстилки в условиях городских экосистем. Показано, что на загрязненных тяжелыми металлами участках процессы деструкции могут происходить более интенсивно по сравнению с естественными экосистемами [62]. Второе направление связано с экспериментальными исследованиями. В частности, подчеркивается роль фотодеградации в процессах деструкции подстилок [55], анализируется роль внесения выбросов микроантропод на интенсивность разложения [59]. Экспериментально изучена роль высоты снегового покрова в процессе минерализации наземного детрита с акцентом на особенности весеннего периода [70]. Сравнительный анализ вытяжек из опада и корневой биомассы [73] показал, что последние характеризуются более высокой кислотностью. В экспериментах показана неоднозначная роль хвои [75], с одной стороны, снижающей скорости разложения за счет низкого содержания азота, с другой – улучшающей водный режим подстилки. Кроме того, хвоя влияет на скорость разложения опада [76]. Детальные исследования посвящены установлению взаимосвязи особенностей подстилок, подстилочной фауны и микробного пула. Нередко подчеркивается роль дождевых червей в процессах разложения [69]. При этом для понимания процессов разложения ключевыми признаны два фактора – разнообразие детрита и разнообразие деструкторов [74]. Общепризнана роль зоомикробного пула, обусловленная типологией подстилок. Типы подстилок детерминируют структуру подстилочной фауны [68], которая может на 50% определять интенсивность разложения растительных остатков, особенно на ранних стадиях сукцессий [56]. Показано влияние уровня увлажнения на скорость разложения [77], при котором длительное и стабильное увлажнение стимулировало разложение лабильной части детрита. Обращают на себя внимание результаты крупномасштабных экспериментальных исследований интенсивности минерализации на основе стандартизированных образцов чая – 65% изменчивости в скоростях разложения приурочено к ранним стадиям [53]. Расширяются масштабы изучения интенсивности разложения при различных сценариях изменения климата [63]. Так, увеличение температуры может оказывать влияние на увеличение содержания лигнина и роста эктомикоризных грибов [64]. Наиболее общим положением, объединяющим исследования, является признание необходимости сочетания долгосрочных натурных и экспериментальных исследований.
Таким образом, исследования показывают, что воздействие на почву растительности и других факторов максимально проявляется в поверхностных и близким к ним горизонтах, следовательно, и наибольшее варьирование свойств наблюдается обычно в верхних горизонтах почвы и подстилках [16, 29, 32, 39, 67]. Поскольку подстилка по сравнению с почвой является более лабильным образованием, ее варьирование в пространстве и чувствительность к локальной пространственной изменчивости факторов почвообразования чрезвычайно высока, что создает определенные сложности при изучении [2, 12, 36, 45]. Изменчивость таких характеристик подстилки, как мощность, запасы, фракционный состав и даже некоторые химические свойства, детерминирована в лесных экосистемах чаще всего парцеллярным строением фитоценоза [16, 27], которое создают эдификаторы древесного яруса. Следует отметить, что понятия парцеллы и особенно тессеры применительно к единице структуры фитоценоза до сих пор являются спорными [16, 17, 25]. Парцелла не во всех случаях является элементарной единицей биогеоценоза (например, парцелла древесная и парцелла окна на месте вывала), тогда как компоненты, совокупность которых образует древесную тессеру – гораздо ближе к этому понятию [31]. При этом не вызывает сомнений тот факт, что живой напочвенный покров, подстилки и почвы под кронами деревьев не одинаковы в разных частях кроны и между кронами. Авторы придерживаются мнения, что наиболее точное понятие тессеры подразумевает совокупность таких элементарных почвенно-фитоценотических единиц, как приствольное пространство, подкроновое и межкроновое. Величина запасов подстилки может изменяться в этих компонентах почти в 2 раза [36], а величина мощностей подстилок – более чем в 5 раз [44]. Тем не менее, до сих пор вопрос о закономерностях варьирования свойств подстилок в пространстве лесного фитоценоза недостаточно хорошо исследован. Очевидно, что изучение подстилки с учетом чрезвычайно высокой временной изменчивости и пространственной неоднородности требует принципиально иного подхода, чем изучение почв. Подстилка, как и почва, часто имеет горизонтное строение [4, 5], причем горизонты существенно различаются по составу и свойствам. Однако часто этот факт игнорируется, и подстилка, несмотря на сложное строение, рассматривается как однородное образование. Это служит причиной того, что нередко общие запасы подстилок приводят без указания их типовой принадлежности, что затрудняет или вовсе исключает возможность сравнения с уже имеющимися литературными сведениями. Живой напочвенный покров в лесных фитоценозах влияет на водный режим и свойства почвы [43], в определенной степени определяет величину опада и депонирование углерода [1], то есть участвует в круговороте. Он является весьма чувствительным показателем лесорастительных условий [20, 21, 47], в том числе особенностей подстилок как наиболее лабильных горизонтов почв. Пока еще мало анализируется взаимосвязь видового состава напочвенного покрова и характера лесных подстилок, хотя именно на уровне тессер [25, 31] происходит взаимодействие трех важных составляющих лесного биогеоценоза – напочвенного покрова, опада и формирующихся в данных условиях подстилок.
Цель настоящей работы – изучить изменение строения и свойств лесных подстилок елового биогеоценоза в системе ствол–крона–окно во взаимосвязи с наиболее важными критериями, основанными на использовании принципов, разработанных в рамках классических экологических шкал [34, 48, 54, 60].
ОБЪЕКТЫ И МЕТОДЫ
Исследованная территория расположена в Солнечногорском районе Московской области, на территории учебного опытного полевого экологического центра “Чашниково”. Среднегодовая температура колеблется около 3.5°С, годовые осадки составляют 600–650 мм [33]. Территория относится к подзоне елово-широколиственных лесов. Наряду с сохранившимися елово-широклиственными лесами, тяготеющими к нераспаханным склоновым ландашфтам, широкое распространение получили мелколиственные леса. Положение исследуемого района в пределах Клинско-Дмитровской гряды обусловливает широкое распространение плоско-волнистого рельефа, который вследствие глубокого расчленения долинной и балочной сетью приобретает ярко выраженный эрозионный характер [11]. Относительно моренных отложений в настоящий период подчеркивается самостоятельность и разновозрастность московского и днепровского оледенения [26]. Непосредственно в районе УОПЦ Чашниково преобладают моренные отложения московского оледенения, с классическими включениями от дресвы до камней, перекрытые покровными суглинками различной мощности [19]. В меньшей степени распространены флювиогляциальные отложения, как правило, приуроченные к ложбине стока ледниковых вод. В пределах всей территории учебно-опытного экологического центра выделяется два крупных массива. Первый характерен для наиболее возвышенной части водораздельных пространств между реками Сходня и Клязьма. Вторая территория включает в себя собственно супераквальные пойменные ландшафты и две надпойменные террасы. В почвенном отношении [10, 19] наиболее общей характеристикой является широкое распространение дерново-подзолистых почв, формирующихся на покровных суглинках, подстилаемых мореной. Многолетние исследования почвенного покрова показывают, что уровень залегания морены значительно варьирует от 50–60 см до 1.5 м [19, 20]. В пределах изучаемого типа леса, приуроченного к склону северной экспозиции, уровень залегания морены составляет 60–70 см. Особенностью почв, развивающихся в условиях склоновых ландшафтов, является оглеение, которое может диагностироваться в пределах иллювиальных горизонтов [42].
Исследуемый еловый фитоценоз расположен на склоне северной экспозиции к первой террасе р. Клязьма. Первый ярус древостоя высотой около 30 м представлен елями примерно 100-летнего возраста (10Е). Второй ярус состоит из одиночных деревьев липы сердцелистной (Tilia cordata) и клена платановидного (Acer platanoides). В подлеске – рябина обыкновенная (Sorbus aucuparia), бересклет бородавчатый (Euonymus verrucosus), черемуха обыкновенная (Padus avium) и лещина обыкновенная (Corylus avellana). Общее проективное покрытие напочвенного покрова, представленного травами, составляет в среднем 40%. В нем преобладают зеленчук желтый (Galeobdolon luteum), звездчатка дубравная (Stellaria nemorum), щитовник мужской (Dryopteris filix-mass).
Почвы – дерново-подзолистые, мощные, глубокоосветленные среднесуглинистые на покровных суглинках, подстилаемых мореной, Albic Retisol (Loamic, Ochric) [18].
Геоботаническое описание произведено на площади 10 × 10 м. В фитоценозе выделены тессеры, состоящие из следующих компонентов: приствольное повышение около ели, подкроновое пространство и межкроновое пространство (окно). Укосы надземной фитомассы напочвенного покрова отобрали с площадок 50 × 50 см, а образцы лесных подстилок (по-горизонтно) с площадок 25 × 25 в 17 тессерах. Учитывали принадлежность видов к эколого-ценотическим свитам, под которыми понимают группу видов, обладающих сходными требованиями к экологическим условиям, и отчасти сходными биологическими особенностями [28]. Биомассу напочвенного покрова выражали в г/м2 на абсолютно сухую навеску. Каждому виду присваивали балл по экологическим шкалам Ландольта [60] и Элленберга [54] или интервал баллов по шкалам Раменского [34] и Цыганова [48]. Использовали следующие показатели: отношение растений к кислотности (Ландольт, Элленберг, Цыганов), к обогащенности азотом (Ландольт, Элленберг, Цыганов) и обогащенности элементами питания, то есть трофность (Цыганов, Раменский). Зная долю биомассы вида в напочвенном покрове, для каждого укоса определили средневзвешенные баллы для точечных шкал и средневзвешенные середины интервалов для диапазонных. Изучали мощность, запасы всех горизонтов подстилок, фракционный состав горизонтов L (соотношение шишек, веток, хвои, коры, ветоши, детрита). К ветоши относили опад травянистых растений. Детрит – мелкие растительные остатки, морфологическая принадлежность которых в отличие от других фракций, уже не может быть установлена. Запасы всех компонентов подстилок рассчитывали на сухую массу (высушенную при 105°C) в г/м2. Запас легкоразлагаемых компонентов рассчитывали как сумму фракции листьев и ветоши. Для подстилок определяли рН водной суспензии (соотношение 1 : 25) и содержание зольных элементов методом сухого озоления (450°С) в пяти тессерах. Обработку результатов проводили с помощью программ Excel и Statistica. Классификационную принадлежность подстилок определяли по классификации Богатырева [5].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Напочвенный покров
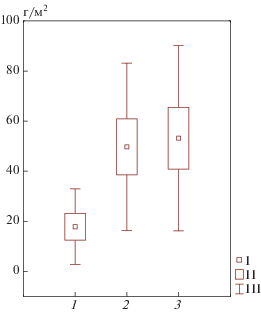
В изученном фитоценозе состав напочвенного покрова определяется положением в системе ствол–крона–окно (табл. 1). Биомасса напочвенного покрова в приствольных пространствах значимо меньше, чем в других компонентах тессеры – в последних она чрезвычайно варьирует в пространстве (рис. 1). В приствольных повышениях выявлено 12 видов, под кронами – 12, а в окнах – 19. Число эколого-ценотических свит – соответственно 6, 6 и 11. Высокое разнообразие видов и свит в окнах связано, по-видимому, с условиями освещения – у стволов и под кронами могут существовать преимущественно теневыносливые. У стволов преобладают виды еловой кисличной свиты (кислица обыкновенная (Oxalis acetosella)), под кронами и в окнах неморальной теневой свиты – зеленчук желтый (Galeobdolon luteum), звездчатка жестколистная (Stellaria holostea). Наличие сорно-рудеральных и нитрофильных видов, мало типичных для хвойного леса, видимо, связано с тем, что выше по склону находится поле, с которого могут поступать удобрения, а также семена подобных растений путем обычного межбиогеоценозного переноса.
Таблица 1.
Особенности напочвенного покрова в разных компонентах тессер
| Параметр | Пространство | ||
|---|---|---|---|
| прист-вольное | подкро-новое | межкро-новое | |
| Число видов | 12 | 12 | 19 |
| Число эколого-ценотических свит | 6 | 6 | 11 |
| Доля видов еловых свит, % | 46.8 | 49.0 | 21.2 |
| Доля неморальных видов, % | 40.0 | 45.9 | 73.8 |
| Доля нитрофилов, % | 8.5 | 5.6 | 4.8 |
| Доля сорно-рудеральных видов, % | 8.8 | 2.7 | 1.4 |
| Доля теневыносливых видов (по Элленбергу), % | 47.0 | 14.7 | 5.8 |
Рис. 1.
Надземная биомасса напочвенного покрова: 1 – приствольные пространства, 2 – подкроновые пространства, 3 – межкроновые пространства. Здесь и далее: I – среднее, II – среднее ± стандартная ошибка, III – среднее ± стандартное отклонение.
Анализ баллов, характеризующих отношение видов к кислотности к обогащенности элементами питания, на основе шкал Ландольта не выявил существенных различий в напочвенном покрове приствольных, подкроновых и межкроновых пространств. Аналогичная оценка напочвенного покрова по шкалам Элленберга, имеющим не четыре, как в шкале Ландольта, а девять градаций, что средневзвешенный балл отношения растений к кислотности почвы значимо увеличивается от приствольного пространства к окнам (рис. 2А). Средневзвешенная середина интервала отношения видов к кислотности по шкале Цыганова также значимо растет от приствольных к межкроновым пространствам, особенно резко изменяясь при переходе от приствольных участков к кронам, что говорит о большей доле видов, предпочитающих кислые почвы, у стволов. Степень варьирования этого показателя в приствольных пространствах значительно меньше, чем в других (рис. 2Б). Средневзвешенный балл по Элленбергу, обозначающий отношение вида к обогащенности субстрата азотом, значимо уменьшается в окнах по сравнению с другими элементами тессеры.
Рис. 2.
Отношение видов напочвенного покрова к кислотности: 1 – приствольный участок, 2 – крона, 3 – окно. А – по шкале Элленберга (средневзвешенный балл), Б – по шкале Цыганова (средневзвешенная середина интервала баллов).

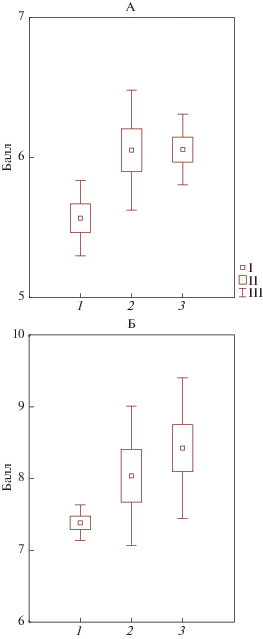
При этом видно, что степень варьирования балла увеличивается от приствольных повышений к другим компонентам тессеры. Аналогичная оценка по шкале Цыганова дала похожий результат (рис. 3) – в приствольных и подкроновых пространствах практически нет различий, тогда как в окнах балл отношения к азоту существенно меньше. Что касается трофности видов, то есть их отношения к общей обогащенности субстрата элементами питания, оцениваемой по шкале Цыганова и Раменского, получена противоположная картина (рис. 4), заключающаяся в том, что средневзвешенная середина диапазона трофности видов напочвенного покрова значимо меньше в приствольном пространстве, чем в других компонентах тессеры. Экологическая оценка напочвенного покрова с помощью комплекса шкал позволила установить закономерности распределения экологических характеристик видов в пределах тессер. По соотношению экологических групп видов диагностируется повышенная кислотность в приствольных пространствах, а также более благоприятные экологические условия для растений в подкроновых и межкроновых пространствах, нежели у стволов.
Рис. 3.
Отношение видов напочвенного покрова к обогащенности субстрата азотом: 1 – приствольный участок, 2 – крона, 3 – окно. А – по шкале Элленберга (средневзвешенный балл), Б – по шкале Цыганова (средневзвешенная середина интервала баллов).
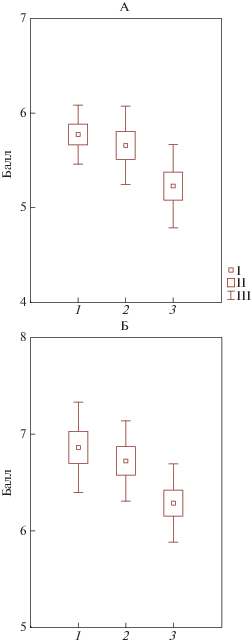
Рис. 4.
Отношение видов напочвенного покрова к обогащенности субстрата элементами питания (трофность): 1 – приствольный участок, 2 – крона, 3 – окно. А – средневзвешенная середина интервала баллов по шкале Цыганова, Б – по шкале Раменского.
Некоторое противоречие выявлено при установлении отношения видов к обогащенности азотом. Доля видов, требовательных к содержанию азота, уменьшается от стволов к межкроновым пространствам. Последний факт никак не объясняется ни составом, ни свойствами подстилок. Вероятно, наличие в подкроновом и приствольном пространстве растений-нитрофилов:- звездчатки средней (Stellaria media), пикульника красивого (Galeopsis speciosa) – имеет другие причины. Почти все нитрофильные виды, ассоциированные с приствольными пространствами, являются эксплерентами, то есть способны эффективно захватывать пространство, где нет или мало других видов. Такими экологическими нишами в данном случае являются приствольные пространства, в которых биомасса и разнообразие видов травяного яруса минимальны.
Морфология лесных подстилок
Анализ полученных данных показал, что подстилки в ельнике представлены деструктивными, ферментативными и гумифицированными типами, доля деструктивного типа подстилок незначительна. Наиболее часто встречаемый тип подстилки – гумифицированный (табл. 2).
Таблица 2.
Встречаемость разных типов подстилок в разных компонентах тессер, %
| Тип подстилки | Пространство | ||
|---|---|---|---|
| приство-льное | подкро-новое | межкро-новое | |
| Деструктивный | 0 | 11 | 0 |
| Ферментативный | 39 | 17 | 61 |
| Гумифицированный | 61 | 72 | 39 |
В большинстве исследуемых подстилок депонировано большое количество органического вещества, что позволяет сделать вывод о низкой скорости разложения. Мощность подстилок уменьшается в ряду ствол–крона–окно (рис. 5), что соответствует литературным данным [3] для еловых фитоценозов. Типичные значения мощностей подстилок варьируют от 5.5 до 12.5 см в приствольном повышении, в подкроновом пространстве – от 2.5 до 5.5 см, в окнах – от 2 до 4.5 см. Установлено, что мощность подстилки у ствола втрое больше, чем в подкроновом и межкроновом пространствах. В двух последних частях тессеры мощность подстилки очень близка. Коэффициент варьирования мощности подстилок во всех точках тессеры составляет около 50%, что соответствует данным других авторов [16, 33]. В пределах тессеры мощность горизонтов L и Н уменьшается от ствола к окну. Мощность горизонта F у ствола вдвое превышает мощность в подкроновом и межкроновом пространствах, различия эти значимы. Подстилки в кроне и окне сходны как по общей мощности, так и по мощностям отдельных горизонтов.
Рис. 5.
Мощность подстилок (А): 1 – приствольный участок, 2 – крона, 3 – окно. Мощности подстилок по горизонтам (Б): 1 – приствольный участок, L; 2 – приствольный участок, F; 3 – приствольный участок, Н; 4 – крона, L; 5 – крона, F; 6 – крона, Н; 7 – окно, L; 8 – окно, F; 9 – окно, Н.
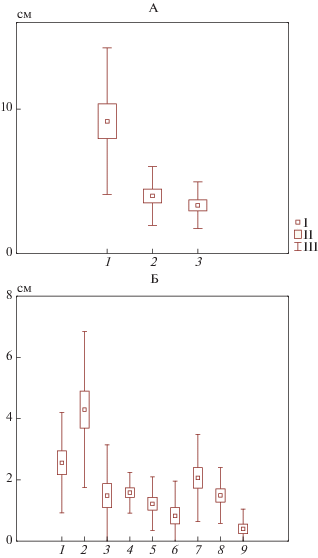
Запасы лесной подстилки, как и мощность, существенно различаются в зависимости от положения в тессере (рис. 6). Максимальные запасы подстилки характерны для приствольных повышений и составляют в среднем около 4500 г/м2. По мере продвижения к кроне и окну общие запасы значимо уменьшаются – 3424 и 2246 г/м2 соответственно. Полученные результаты соответствуют литературным данным по запасам подстилок еловых лесов [16, 36, 44]. Величина запасов в приствольном повышении характеризуется значительным варьированием (V = 70%), в то время как изменчивость величины запасов под кроной и в окне существенно меньше (V = 40%). Полученные данные по варьированию запасов и мощности подстилок показывают, что максимальная степень варьирования характерна для подстилок приствольных участков, представленных в подавляющем большинстве случаев либо мертвопокровными парцеллами [44], либо с двумя–четырьмя видами в напочвенном покрове. Такое варьирование говорит о высоком разнообразии условий формирования подстилок в приствольных пространствах. Анализ распределения запасов подстилок по горизонтам показывает, что во всех участках тессеры запасы органического вещества в горизонте F больше, чем суммарные запасы горизонтов L и Н (рис. 6). Это еще раз подчеркивает некоторую заторможенность круговорота и, в частности, низкую скорость разложения опада. Запасы горизонта L, не имеющие значимого различия в разных участках тессеры, свидетельствуют о близких скоростях разложения опада в поверхностных горизонтах подстилок.
Рис. 6.
Запасы лесных подстилок. А – общие: 1 – приствольный участок, 2 – крона, 3 – окно. Б – по горизонтам: 1 – приствольный участок, L; 2 – приствольный участок, F; 3 – приствольный участок, Н; 4 – крона, L; 5 – крона, F; 6 – крона, Н; 7 – окно, L; 8 – окно, F; 9 – окно, Н.

Различия в условиях формирования подстилок проявляются на более поздних стадиях разложения органического вещества, что отражается в мощности нижележащих горизонтов. Запасы горизонта F значимо уменьшаются от ствола к межкроновому пространству, в то время как запасы горизонта Н лишь в межкроновом пространстве значимо меньше, чем в остальных компонентах тессеры. Изучение фракционного состава горизонтов L в пределах тессеры показало, что у ствола под кроной преобладающими фракциями являются ветки, в межкроновом пространстве – листья (рис. 7). Сокращение поступления веток и хвои в окнах обусловлено влиянием дерева-эдификатора, а увеличение доли листьев – большей вероятностью их накопления в этой точке тессеры. Показано, что содержание фракции детрита в подкроновом и межкроновом пространствах примерно одинаково, в приствольном пространстве – незначительно меньше. Возможно, в сложных подстилках детрит быстро перемещается в нижележащие горизонты. Содержание детрита составляет почти 1/5, что говорит о слабых темпах разложения. С другой стороны, в подстилках ельников содержание детрита может достигать 50% [41].
Рис. 7.
Фракционный состав горизонтов L. А – приствольное повышение, Б – подкроновое пространство, В – межкроновое пространство. 1 – ветки, 2 – листья, 3 – хвоя, 4 – детрит, 5 – шишки, 6 – ветошь.
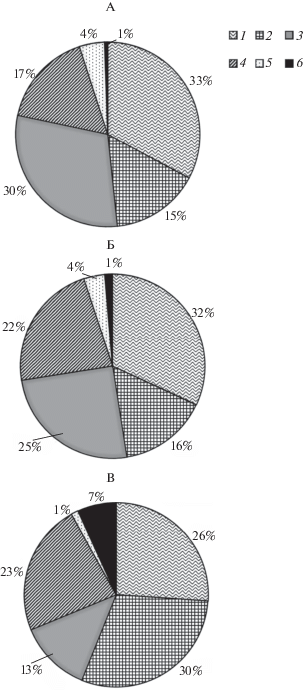
В окнах отмечено повышенное содержание опада трав, что связано с активным ростом травянистых растений из-за оптимальных условий освещенности [45]. В горизонте L доля легкоразлагаемых компонентов (ветошь и листья) увеличивается в 2 раза от ствола к окну. Отношение запасов разных горизонтов подстилок можно использовать для оценки скорости их разложения. Информативность этих показателей определяется тем, что они отражают соотношение поступления опада и его разложения – стадия разложения органического вещества определяет время его пребывания в соответствующем горизонте подстилки [35]. Соотношение запасов горизонтов L и F наиболее высоко в подкроновых пространствах, о минимально – у стволов. То же относится к соотношению L/Н (табл. 3). Низкое значение соответствует повышенному накоплению недоразложенного детрита в составе горизонтов F и Н, в то время как высокое – интенсивному разложению горизонта L без промежуточных стадий в виде других горизонтов. Возможно, стволовые воды, содержащие фенолы и таннины, ингибируют активность микроорганизмов в приствольных пространствах [46], что подтверждено исследованиями [40].
Химические свойства лесных подстилок
Актуальная кислотность – наиболее динамичный показатель, она варьирует в пространстве в зависимости от положения в фитогенном поле эдификатора [24, 30]. Стоит отметить, что кислотность подстилок значительно ниже, чем в еловых лесах, находящихся далеко от очагов антропогенного воздействия – в ельниках Костромской области аналогичного возраста рН составляет 4 и менее [44]. В изучаемом ельнике кислотность подстилок значимо больше в приствольных пространствах, нежели в других частях тессеры, это касается всех горизонтов подстилки (табл. 4).
Таблица 4.
Химические свойства лесных подстилок
| Показатель | Горизонт или его компонент | Ствол | Крона | Окно |
|---|---|---|---|---|
| pH | L | 5.49 ± 0.05* | 5.73 ± 0.08 | 5.72 ± 0.25 |
| F | 5.27 ± 0.10 | 5.57 ± 0.05 | 5.35 ± 0.07 | |
| H | 5.02 ± 0.04 | 5.08 ± 0.05 | 5.17 ± 0.10 | |
| Зольность, % | Листья | 7.2 ± 0.57 | 9.8 ± 0.88 | 8.8 ± 1.15 |
| Ветошь | 6.2 ± 0.85 | 6.8 ± 1.7 | 7.3 ± 2.0 | |
| Хвоя | 8.0 ± 2.61 | 11.3 ± 1.5 | 8.0 ± 0.92 | |
| Детрит | 14.7 ± 4.78 | 21.6 ± 2.7 | 23.2 ± 4.92 | |
| Ветки | 3.8 ± 0.13 | 5.2 ± 1.2 | 4.7 ± 0.71 | |
| Шишки | 3.6 ± 1.1 | 5.1 ± 1.0 | 2.7 ± 0.46 | |
| L (средневзвешенное значение) | 7.26 | 10.8 | 10.7 | |
| F | 18.2 ± 5.0 | 42.6 ± 1.9 | 39.8 ± 0.12 | |
| Н | 39.3 ± 8.3 | 41 ± 1.8 | 40.0 ± 5.9 | |
| Запасы золы, г/м2 | Листья | 4.68 ± 1.6 | 11.0 ± 3.0 | 10.6 ± 2.6 |
| Ветошь | 0.14 ± 0.05 | 0.66 ± 0.1 | 1.97 ± 0.24 | |
| Хвоя | 10.2 ± 2.0 | 20.3 ± 2.3 | 4.1 ± 0.8 | |
| Детрит | 10.3 ± 1.8 | 35.2 ± 1.5 | 21.6 ± 0.9 | |
| Ветки | 5.24 ± 1.5 | 12.1 ± 1.6 | 4.93 ± 1.0 | |
| Шишки | 0.65 ± 0.05 | 1.4 ± 0.1 | 0.14 ± 0.01 | |
| L (сумма) | 31.2 ± 1.2 | 80.6 ± 3.5 | 43.3 ± 2.5 | |
| F | 455 ± 14.5 | 720.4 | 469.6 ± 10.1 | |
| Н | 491 ± 60.0 | 419 ± 11.2 | 164.8 ± 7.2 | |
| Общий запас зольных элементов | 977.2 | 1220 | 677.7 | |
| Запасы зольных элементов (з.э.) в L/запасы з.э. общие | 0.03 | 0.06 | 0.06 | |
| Запасы зольных элементов в легкоразлагаемых компонентах горизонта L, г/м2 | 5.36 | 13.4 | 14.9 | |
Причиной могут быть, в частности, кислотные продукты, поступающие с дождевыми водами и стекающие по стволу и крупным ветвям, а также кислые продукты разложения опада хвои, доля которого максимальна в приствольной части. Различия рН подстилок между кронами и окнами не столь существенны. Во всех компонентах тессеры рН подстилки уменьшается от горизонтов L к горизонтам Н, что связано с процессом разложения опада.
Среди компонентов горизонта L наибольшая зольность выявлена для детрита, несколько меньшая – для хвои ели, листьев и ветоши (6–10%), самая низкая – для ветвей и шишек – 5% и менее, что в целом соответствует данным других авторов [38]. Интересно, что одни и те же компоненты характеризуются несколько разной зольностью в разных компонентах тессеры: максимальная зольность компонентов подстилки отмечена в подкроновых пространствах, в приствольных пространствах значение зольности почти всегда ниже, чем в других. В приствольном пространстве запасы зольных элементов горизонтов L и F отличаются от других участков тессеры в меньшую сторону. Меньшая зольность опада является дополнительным фактором более медленного разложения органического вещества и большей кислотности. Во всех подстилках зольность увеличивается от горизонта L к горизонту Н, что связано с близостью минеральных горизонтов, воздействием мезо- и микрофауны, а также с увеличением степени разложения. Аналогичные результаты получены для ельников другими авторами [41]. Запасы зольных элементов почти во всех компонентах горизонта L характеризуются наименьшим значением в приствольных пространствах, а наибольшим – в подкроновых. Соответственно, запасы зольных элементов в легкоразлагаемых компонентах, вносящих наибольший вклад в биологический круговорот, в приствольных пространствах составляют около 5 г/м2, а в других точках тессеры – более 13–15 г/м2. Зольные элементы и их запас непосредственно определяют скорость разложения опада [49, 50] и, как следствие, характер органопрофиля и лесорастительных условий. В пределах тессеры распределение запасов зольных элементов не соответствует распределению запасов подстилки, видимо, из-за разного соотношения высокозольных и низкозольных фракций. Максимальная величина запаса зольных элементов в подстилке установлена в подкроновых пространствах, где отмечается также максимальная зольность ее компонентов. Запасы зольных элементов в подстилках подкроновых участков в 1.2–1.5 раза больше, чем в других точках тессеры, что, возможно, связано с дополнительным атмосферным поступлением пыли с осадками. По данным Карпачевского [15], в связи с особенностями строения кроны ели, под кроны поступает втрое больше осадков, чем в участки у ствола. Увеличение зольности подстилок за счет перераспределения осадков подтверждается тем, что наибольшие различия в запасах зольных элементов наблюдаются в горизонтах L (до 2 раз) и F (1.5 раза), уменьшаясь в горизонте Н. Значительный запас зольных элементов в подстилках под кронами оказывает влияние на активность микроорганизмов. Дополнительное поступление зольных элементов, а также их значительный запас в виде легкоразлагаемых фракций (табл. 3) стимулирует работу микроорганизмов и активизирует процессы разложения, что приводит к снижению мощности подстилок. Подстилки приствольного и подкронового пространств очень близки по фракционному составу, в том числе по доле хвои, однако их мощность различается более, чем в 2 раза, что сближает последние с подстилками окон.
Эколого-ценотическая структура напочвенного покрова в целом адекватно отражает химические свойства подстилок, а также показатели, связанные со скоростью разложения. Сравнительно повышенные значения актуальной кислотности у стволов хорошо коррелируют с видами, указывающими на эти условия в рамках экологических шкал. Запасы зольных элементов, особенно в легкоразлагаемых фракциях, указывают на существенное отличие приствольных повышений от остальных компонентов тессеры по скорости разложения и, как следствие, интенсивности биологического круговорота и экологическим условиям в целом. Специфика строения и генезиса подстилок для приствольных участков отражается в особенностях напочвенного покрова, характеризующегося преобладанием видов, указывающих в рамках шкал Раменского и Цыганова на значимо пониженные баллы трофности, соответствующие более бедным почвам. Таким образом, показатели лесных подстилок, указывающие на относительную обогащенность элементами питания, такие как зольность, запас зольных элементов в легкоразлагаемых фракциях, погоризонтное соотношение запасов органического вещества и запасов зольных элементов, связаны с характеристиками напочвенного покрова, определенными по экологическим шкалам.
ВЫВОДЫ
1. Подстилки ельника кислично-зеленчукового характеризуются значительной пространственной изменчивостью: в пределах тессеры встречаются деструктивные, ферментативные и гумифицированные подстилки с преобладанием последних. Мощность подстилок варьирует от 3 до 9 см, запасы – от 2000 до 5000 г/м2.
2. Максимальное варьирование свойств отмечается в подстилках приствольных пространств, что свидетельствует о значительном разнообразии условий формирования подстилок в данном участке тессеры. Подстилки приствольных участков характеризуются минимальными значениями рН и минимальными запасами зольных элементов в горизонтах L и легкоразлагаемых компонентах, что соотносится с максимальным долевым участием ацидофильных видов и видов, мало требовательных к элементам питания.
3. Влияние кроны ели через поступление опада проявляется в том, что в горизонтах L подстилок приствольных и подкроновых участков в 2 раза больше долевое участие хвои и в 2 раза меньше доля легкоразлагающихся компонентов подстилки по сравнению с подстилками межкроновых пространств. Мощности подстилок, как и их запасы, уменьшаются в ряду ствол–крона–окно.
4. В результате поступления пылевых частиц из атмосферы и перераспределения атмосферных осадков в пределах кроны, в подкроновых участках отмечены максимальные запасы зольных элементов в подстилках, несмотря на средние общие запасы подстилки. Под кронами также отмечена наибольшая величина надземной биомассы, определяющая повышенное поступление зольных элементов после ее разложения. Совокупность этих факторов создает благоприятные условия для активизации процессов разложения органического вещества и снижения мощности подстилок в данном участке тессеры.
5. Показатели, характеризующие скорость разложения органического вещества (запасы зольных элементов в подстилке и ее легкоразлагаемых фракциях, соотношение мощностей и запасов горизонтов), напрямую связаны с соотношением в напочвенном покрове видов, характеризующихся разным уровнем трофности, установленным по шкалам Л.Г. Раменского и Д.Н. Цыганова.
6. Группировка видов, имеющих различное отношение к кислотности, проведенная на основе использования экологических шкал Элленберга и Цыганова, получила подтверждение в реальных показателях кислотности, характеризующих лесные подстилки.
7. В методическом отношении использование экологических шкал, группирующих виды по уровню отношения к кислотности и общей обогащенности субстрата элементами питания, следует признать дополнительной информационной базой, которую можно использовать при исследовании подстилок.
Список литературы
Аткин А.С., Аткина Л.И. Масса травяно-кустарничкового яруса в лесных фитоценозах // Продуктивность лесных фитоценозов. Красноярск: Изд-во ИЛиД СО АН СССР, 1984. С. 17–27.
Благовещенский Ю.Н., Богатырев Л.Г., Соломатова Е.А., Самсонова В.П. Пространственная изменчивость мощности подстилок в лесах Карелии // Почвоведение. 2006. № 9. С. 1029–1035.
Богатырев Л.Г. Генезис лесных подстилок в различных природных зонах европейской части России [Результаты многолетних исследований в лесах таежной и лесостепной зон] // Лесоведение. 1995. № 4. С. 3–12.
Богатырев Л.Г. Образование подстилок – один из важнейших процессов в лесных экосистемах // Почвоведение. 1996. № 4. С. 501–511.
Богатырев Л.Г. О классификации лесных подстилок // Почвоведение. 1990. № 3. С. 118–127.
Богатырев Л.Г., Демин В.В. Матышак Г.В., Сапожникова В.А. О некоторых теоретических аспектах исследования лесных подстилок // Лесоведение. 2004. № 4. С. 17–29.
Богатырев Л.Г., Сапожникова В.А., Воедило А.А. Трансформация органического вещества в сосновых экосистемах как один из критериев оценки интенсивности круговорота // Вестник Моск. ун-та. Сер. 17, почвоведение. 1999. № 3. С. 13–23.
Богатырев Л.Г., Свентицкий И.А., Шарафутдинов Р.Н., Степанов А.А. Лесные подстилки и диагностика современной направленности гумусообразования в различных географических зонах // Почвоведение. 1998. № 7. С. 783–793.
Богатырев Л.Г., Щенина Т.Г., Комарова М.С. Характеристика лесных подстилок при зарастании вырубок южно-таежной подзоны // Почвоведение. 1989. № 7. С. 106–113.
Васильевкая В.Д., Зборищук Ю.Н., Ульянова Т.Ю. Почвы и почвенный покров УОПЦ Чашниково // Развитие почвенно-экологических исследований. М.: Изд-во Моск. ун-та, 1999. С. 25–30.
Дик Н.Е., Лебедев В.Г., Соловьев А.И., Спиридонов А.И. Рельеф Москвы и Подмосковья. М.: Изд-во географ. лит-ры, 1949. 194 с.
Ефремова Т.Т., Секретенко О.П., Аврова А.Ф., Ефремов С.П. Геостатистический анализ пространственной изменчивости запасов зольных веществ в подстилке болотных березняков Западной Сибири // Почвоведение. 2013. № 1. С. 56–66.
Иванов А.В., Браун М., Замолодчиков Д.Г., Лынов Д.В., Панфилова Е.В. Лесные подстилки как звено цикла углерода хвойно-широколиственных насаждений южного риморья // Почвоведение. 2018. № 10. С. 1226–1233 https://doi.org/10.1134/S0032180X18100052
Ильина Т.М., Сапожников А.П. Лесные подстилки как компонент лесного биогеоценоза // Вестник КрасГАУ, 2007. № 5. С. 45–48.
Карпачевский Л.О. Динамика свойств почвы. М.: ГЕОС, 1997. 169 с.
Карпачевский Л.О. Пестрота почвенного покрова в лесном биогеоценозе. М.: Изд-во Моск. ун-та, 1977. 312 с.
Карпачевский Л.О., Зубкова Т.А., Ташнинова Л.Н., Руденко Р.Н. Почвенный покров и парцеллярная структура лесного биогеоценоза // Лесоведение. 2007. № 6. С. 107–113.
Кириллова Н.П., Силева Т.М., Ульянова Т.Ю., Розов С.Ю., Ильяшенко М.А., Макаров М.И. Цифровая почвенная карта УОПЭЦ Чашниково МГУ им. М.В. Ломоносова // Вестник Моск. ун-та. сер. 17, почвоведение. 2015. № 2. С. 22–29. https://doi.org/10.3103/S0147687415020039
Кириллова Н.П., Силева Т.М., Ульянова Т.Ю., Розов С.Ю., Смирнова И.Е. Цифровая крупномасштабная карта почвообразующих пород и принципы ее составления // Вестник Моск. ун-та. Сер. 17, почвоведение. 2017. № 3. С. 3–9.
Клещева Е.А. Использование экологических шкал для индикации современного состояния лесных сообществ (На примере сосновых лесов) // Экология. 2007. № 2. С. 104–110.
Копцик Г.Н., Багдасарова Т.В., Горленко О.В. Взаимосвязи видового разнообразия растений и свойств почв в экосистемах южной тайги // Бюл. МОИП. отд. биол. 2001. Т. 106. Вып. 2. С. 31–38.
Кузнецов В.А., Рыжова И.М., Стома Г.В. Изменение лесных экосистем мегаполиса под влиянием рекреационного воздействия // Почвоведение. 2019. № 5. С. 633–642. https://doi.org/10.1134/S0032180X1905006X
Лиханова Н.В. Роль растительного опада в формировании лесной подстилки на вырубках ельников средней тайги // Изв. вузов. Лесн. журн. 2014. № 3. С. 52–66.
Лукина Н.В., Горбачева Т.Т., Никонов В.В., Лукина М.А. Пространственная изменчивость кислотности AL-Fe-гумусовых подзолов // Почвоведение. 2002. № 2. С. 163–173.
Лукина Н.В., Орлова М.А., Исаева Л.Г. Плодородие лесных почв как основа взаимосвязи почва-растительность // Лесоведение. 2010. № 5. С. 45–56.
Макарова Н.В., Макаров О.А. стратиграфии четвертичных отложений центра Русской равнины (дискуссионные вопросы) // Вестник Моск. ун-та. Сер. 4, геология. 2004. № 6. С. 19–25.
Надпорожская М.А., Зубкова Е.В., Фролов П.В., Быховец С.С., Чертов О.Г. Соподчиненность почвенных условий и растительных сообществ в сосняках как следствие действия комплекса факторов // Вестник ТвГУ. Сер. Биология и экология. 2018. С. 122–138.
Ниценко А.А. Об изучении экологической структуры растительного покрова // Бот. журн. 1969. Т. 54. № 7. С. 1002–1014.
Орешкина Н.С. Статистические оценки пространственной изменчивости почв. М.: Изд-во Моск. ун-та, 1988. С. 14–18.
Орлова М.А., Лукина Н.В., Смирнов В.Э., Краснов Д.А., Камаев И.О. Плодородие почв еловых лесов Хибинских гор // Почвоведение. 2012. № 6. С. 682–694.
Орлова М.А., Лукина М.В., Смирнов В.Э. Методические подходы к отбору образцов лесной подстилки с учетом мозаичности лесных биогеоценозов // Лесоведение. 2015. № 3. С. 214–221.
Подвезенная М.А., Рыжова И.М. Изменчивость содержания и запасов углерода в почвах лесных биогеоценозов южной тайги // Лесоведение. 2011. № 1. С. 52–60.
Почвенно-агрономическая характеристика Чашниково. Ч. 1. M: Изд-во Моск. ун-та, 1986. С. 93.
Раменский Л.Г., Цаценкин И.А., Чижиков О.Н., Антипов Н.А. Экологическая оценка кормовых угодий по растительному покрову. М.: Сельхозгиз., 1956. 472 с.
Решетникова Т.В. Лесные подстилки как депо биогенных элементов // Вестник КрасГАУ. 2011. № 12. С. 74–82.
Рыжова И.М., Подвезенная М.А. Пространственная вариабельность запасов органического углерода в почвах лесных и степных биогеоценозов // Почвоведение. 2008. № 12. С. 1429–1437.
Сапожников А.П. Лесная подстилка – номенклатура, классификация, индексация // Почвоведение. № 5. 1984. С. 54–63.
Семенюк О.В., Богатырев Л.Г., Ваганова М.А. Характеристика подстилок парковых насаждений исторических ландшафтов на примере музея-усадьбы “Архангельское” // Бюл. Московского общества испытателей природы. Отдел биологический. 2017. Т. 122. № 5. С. 37–49.
Семенюк О.В., Ильяшенко М.А. Пространственная изменчивость почвенных свойств разновозрастных сосняков пейзажной части паркового комплекса “Архангельское” // Вестник Моск. ун-та. Сер. 17, почвоведение. 2013. № 1. С. 23–29.
Сизоненко Т.А., Загирова С.В., Хабибуллина Ф.М. Микробные сообщества в подстилке ельника черничного средней тайги // Почвоведение. 2010. № 10. С. 1221–1228.
Соломатова Е.А. Фракционный и компонентный состав лесных подстилок ельников-черничников восточной Фенноскандии // Лесоведение. 2013. № 6. С. 37–46.
Сорокина Н.П. Региональная модель почвенно-ландшафтных связей (на примере Клинско-дмитровской гряды // Почвоведение. 1998. № 4. С. 389–398.
Сунгурова Н.Р. Изменение видового состава живого напочвенного покрова в культурах сосны и ели // Экологические проблемы севера. 2005. Вып. 8. С. 103–106.
Телеснина В.М., Ваганов И.Е., Карлсен А.А., Иванова А.Е., Жуков М.А., Лебедев С.М. Особенности морфологии и химических свойств пост-агрогенных почв южной тайги на легких отложениях (Костромская область) // Почвоведение. 2016. № 1. С. 115–129.
Телеснина В.М., Семенюк О.В., Богатырев Л.Г. Свойства лесных подстилок во взаимосвязи с напочвенным покровом в лесных экосистемах Подмосковья (на примере УОПЭЦ “Чашниково”) // Вестник Моск. ун-та. Сер. 17, почвоведение. 2017. № 4. С. 11–20.
Фомичева О.А., Полянская Л.М., Никонов В.В., Лукина Н.В., Орлова М.А., Исаева Л.Г., Звягинцев Д.Г. Численность и биомасса почвенных микроорганизмов в коренных старовозрастных северо-таежных еловых лесах // Почвоведение. 2006. № 12. С. 1469–1478.
Ханина Л.Г., Грозовская И.С., Смирнов В.Э. Анализ базы данных по биомассе лесного напочвенного покрова для моделирования его динамики в круговоротных моделях лесных экосистем // Хвойные леса бореальной зоны. 2013. Т. XXXI. № 1–2. С. 22–29.
Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М.: Наука, 1983. 196 с.
Berg B. Litter decomposition and organic matter turnover in northern forest soils // Forest Ecology end Management. 2000. V. 133. № 1–2. P. 13–23.
Berg B., McClaugherty C. Plant litter. Decomposition, humus formation, carbon sequestration. Berlin: Springer-Verlag, 2008. 340 p.
Bilby R.E., Heffner J.T. Factors influencing litter delivery to streams // Forest Ecology and Management. 2016. V. 369. P. 29–37.
Cao B., Domke G.M., Russell M.B., Walters B.F. Spatial modeling of litter and soil carbon stocks on forest land in the conterminous United States // Sci. Total Environ. 2019. V. 654. P. 94–106.
Djukic I., Kepfer-Rojas S., Schmidt I.K., Larsen K.S. Early stage litter decomposition across biomes // Sci. Total Environ. 2018. V. 628–629. P. 1369–1394.
Ellenberg H. Zeigerwerte der Gefasspflanzen Mitteleuropas. Gottingen: Goltze. 1974. 97 p.
Enkhmaa E., Xuehua Y., Congwen W., Zhenying H., Johannes H.C. Cornelissen. Positive and negative effects of UV irradiance explain interaction of litter position and UV exposure on litter decomposition and nutrient dynamics in a semi-arid dune ecosystem // Soil Biol. Biochem. 2018. V. 124. P. 245–254.
Frouz J. Effects of soil macro- and mesofauna on litter decomposition and soil organic matter stabilization // Geoderma. 2018. V. 332. P. 161–172.
Jie D., Jianzhi N., Zhaoliang G., Xiongwen C., Zhijun Z. Effects of rainfall intensity and slope on interception and precipitation partitioning by forest litter layer // Catena. 2019. V. 172. P. 711–718.
Johansson M.B., Berg B., Meentemeyer V. Litter massloss rates in late stages of de-composition in a climatic transect of pine forests. Long-term decomposition in a Scots pine forest. IX // Can. J. Bot. 1995. V. 73. P. 1509–1521.
Joly F.X., Coulis M., Gérard A., Fromin N., Hättenschwiler S. Litter-type specific microbial responses to the transformation of leaf litter into millipede feces // Soil Biol. Biochem. 2015. V. 86. P. 17–23.
Landolt E. Okologische Zeigerwerts zur Sweizer Flora // Veroff. Geobot. Inst. ETH. Zurich. 1977. H. 64. P. 1–208.
Lucas-Borja M.E., J. Hedo J.S., Yang Y., Shen Y., Candel-Pérez D. Nutrient, metal contents and microbiological properties of litter and soil along a tree age gradient in Mediterranean forest ecosystems // Sci. Total Environ. 2019. V. 650. P. 749–758.
Lucisine P., Lecerf A., Danger M., Felten V., Maunoury-Danger F. Litter chemistry prevails over litter consumers in mediating effects of past steel industry activities on leaf litter decomposition // Sci. Total Environ. 2015. V. 537. P. 213–224.
Maes S.L., Blondeel H., Perring M.P., Depauw L., Verheyen K. Litter quality, land-use history, and nitrogen deposition effects on topsoil conditions across European temperate deciduous forests // Forest Ecology and Management. 2019. V. 433. P. 405–418.
Morrison E.W., Pringle A., Linda T.A. van Diepen, Grandy A.S., Frey S.D. Warming alters fungal communities and litter chemistry with implications for soil carbon stocks // Soil Biol. Biochem. 2019. V. 132. P. 120–130.
Mukesh K.G., Kwang-Sik L., Björn B., Byeong-Yeol S., Jeh-Yeong Y. Trends of major, minor and rare earth elements in decomposing litter in a cool temperate ecosystem, South Korea // Chemosphere. 2019. V. 222. P. 214–226.
Ochoa-Hueso R., Delgado-Baquerizo M., Tuan An King P., Benham M., Power S.A. Ecosystem type and resource quality are more important than global change drivers in regulating early stages of litter decomposition // Soil Biol. Biochem. 2019. V. 129. P. 144–152.
Orlova M., Lukina M., Tutubalina O., Smirnov V., Isaeva L., Hofgaard A. Soil nutrients spatial variability in forest-tundra ecotones on the Kola peninsula, Russia // Biogeochemistry. 2013. V. 113. P. 283–305. https://doi.org/10.1007/s10533-012-9756-6
Otsing E., Barantal S., Anslan S., Koricheva J. Litter species richness and composition effects on fungal richness and community structure in decomposing foliar and root litter // Soil Biol. Biochem. 2018. V. 125. P. 328–339.https://doi.org/10.1016/j.soilbio.2018.08.006
Patoine G., Thakur M.P., Friese J., Nock C., Eisenhauer N. Plant litter functional diversity effects on litter mass loss depend on the macro-detritivore community // Pedobiologia. 2017. V. 65. P. 29–42.
Qiqian W. Effects of snow depth manipulation on the releases of carbon, nitrogen and phosphorus from the foliar litter of two temperate tree species // Sci. Total Environ. 2018. V. 643. P. 1357–1365.
Stirling E., Macdonald L.M., Smernik R.J., Cavagnaro T.R. Post fire litters are richer in water soluble carbon and lead to increased microbial activity // Appl. Soil Ecology. 2019. V. 136. P. 101–105.
Stirling E., Smernik R.J., Macdonald L.M., Cavagnaro T.R. The effect of fire affected Pinus radiata litter and char addition on soil nitrogen cycling // Sci. Total Environ. 2019. V. 664. P. 276–282.
Toko T., Saori F., Lijuan S., Yasuhiro H., Nagamitsu M. Leachate from fine root litter is more acidic than leaf litter leachate: A 2.5-year laboratory incubation // Sci. Total Environ. 2018. V. 645. P. 179–191.
Tresch S., Frey D., Renée-Claire Le Bayon, Zanetta A., Moretti M. Litter decomposition driven by soil fauna, plant diversity and soil management in urban gardens // Sci. Total Environ. 2019. V. 658. P. 1614–1629.
Wenya X., Han Y.H. Chen, Praveen K., Chen C., Qingwei G. Multiple interactions between tree composition and diversity and microbial diversity underly litter decomposition // Geoderma. 2019. V. 341. P. 161–171. https://doi.org/10.1111/mec.12481
Xiaoxi Z., Boya W., Zengwen L. Coniferous litter extracts inhibit the litter decomposition of Catalpa fargesii Bur and Eucommia ulmoides Oliver // Acta Oecologica. 2018. V. 93. P. 7–13.
Yajun X., Yonghong X., Huayun X., Xinsheng C., Feng L. The effects of simulated inundation duration and frequency on litter decomposition: A one-year experiment // Limnologica. 2019. V. 74. P. 8–13.
Дополнительные материалы отсутствуют.