

Почвоведение, 2020, № 10, стр. 1247-1261
Свободное железо и железовосстанавливающие микроорганизмы в почвах и многолетнемерзлых отложениях северо-востока Сибири
Е. М. Ривкина a, *, Д. Г. Федоров-Давыдов a, А. Г. Захарюк b, В. А. Щербакова b, Т. А. Вишнивецкая a, c
a Институт физико-химических и биологических проблем почвоведения РАН
142290 Московская область, Пущино, ул. Институтская, 2, Россия
b Институт биохимии и физиологии микроорганизмов им. Г.К. Скрябина
142290 Московская область, Пущино, Институтская, 5, Россия
c Университет штата Теннесси
TN 37996-1605 Ноксвилл, США
* E-mail: rivkina@issp.psn.ru
Поступила в редакцию 04.03.2020
После доработки 21.04.2020
Принята к публикации 27.04.2020
Аннотация
Соответствие между содержанием аморфного (оксалаторастворимого) железа и морфохроматическими признаками глея говорит о протекании современного глееобразования в тундровых почвах Колымской низменности. Надмерзлотный восстановленный глеевый горизонт, вытаивающий из мерзлоты в наиболее теплые годы, мы рассматриваем как реликт более теплого и влажного этапа почвообразования. На основании комплексных анализов содержания подвижного железа и аннотаций метагеномов показано, что в многолетнемерзлых отложениях, сформировавшихся в гидроморфных условиях, и в современной минеральной почве доминируют микроорганизмы, относящиеся к филуму Proteobacteria, способные осуществлять восстановление железа. В лабораторных экспериментах процесс микробного восстановления железа проходил активнее при 5°С, чем при 20°С, что указывает на большую активностью психрофильной части микробного сообщества. На этом основании можно предположить, что основная часть культивируемых сообществ железовосстанавливающих бактерий адаптирована к низким температурам Арктики. При потеплении климата, увеличении количества атмосферных осадков, повышении температуры мерзлоты, увеличении мощности сезонно-талого слоя и высокого валового содержания железа процессы железоредукции будут играть еще большую роль.
ВВЕДЕНИЕ
Процессы микробиологического восстановления железа так же, как сульфатредукция и денитрификация, играют важную роль в трансформации органического вещества и создании благоприятных окислительно-восстановительных условий для образования метана - одного из важнейших парниковых газов. Наблюдаемое в последние десятилетия изменение климата привело на территории Субарктики к увеличению мощности сезонно-активного слоя и повышению температуры мерзлоты ниже слоя годовых теплооборотов [9, 10, 14], что, очевидно, повлияет на активизацию биологических процессов.
В почвах и донных осадках двухвалентное железо является донором электронов для железоокисляющих микроорганизмов, а Fe(III) может функционировать в качестве конечного акцептора электронов для железоредукторов [13, 25, 27]. В Субарктике соотношение ионов Fe(III) и Fe(II) часто является маркером условий формирования и криоконсервации отложений. Показано, что в многолетнемерзлых отложениях (ММО) и почвах, образование которых проходило преимущественно в гидроморфных условиях, содержание закисного железа заметно больше, чем окисного [6, 7]. В этих публикациях продемонстрирована зависимость между содержанием Fe(II) и метана в озерных, озерно-аллювиальных и морских отложениях голоценового и плейстоценового возраста. Известно, что микробиологические процессы железоредукции, наряду с денитрификацией и сульфатредукцией, создают благоприятные окислительно-восстановительные условия для прохождения терминального анаэробного процесса - биогенного метанообразования.
Ранее нами показано, что в многолетнемерзлых голоценовых (аласных) отложениях возраста от 7 до 10 тыс. лет сохраняют жизнеспособность бактерии, способные осуществлять восстановление трехвалентного железа Fe(III) до Fe(II) при температуре инкубации 15°С [19]. Численность железоредукторов составляла 2 × 103 кл./г при содержании двухвалентного железа 0.008-0.027 г/100 г почвы. В залегающих ниже плиоцен-нижнеплейстоценовых отложениях олерской свиты возраста более 600 тыс. лет, несмотря на значительно большее содержание Fe(II), от 0.156 до 0.240 г/100 г почвы, жизнеспособные железо-восстанавливающие бактерии не обнаружены. Из криопэгов (гиперсоленых прослоев незамерзших грунтов и линз воды внутри многолетнемерзлых отложений) полуостровов Варандей и Ямал были выделены и описаны психроактивные сульфатредукторы Desulfovibrio arcticus [18] и Desulfovibrio gilichinskyi [21], восстанавливающие Fe(III). Культуры психрофильных и психроактивных бактерий, способные использовать только трехвалентное железо в качестве источника энергии для роста, в многолетнемерзлых отложениях, не выделены.
Теоретические и экспериментальные данные [12] указывают на то, что в природе прямое, без предварительного растворения Fe(III)-содержащих соединений, восстановление оксидов железа является широко распространенным процессом. В лабораторных условиях железоредукторы в качестве акцептора электрона чаще используют растворенные соединения трехвалентного железа такие, как цитрат Fe(III) и ЭДТА Fe(III). Показано, что представители рода Shewanella, Geobacter, Aeromonas могут осуществлять реакцию восстановления непосредственно на поверхностях железосодержащих минералов, что связано с расположением фермента железоредуктазы во внешней мембране этих бактерий.
В настоящее время из морских осадков Арктики и Антарктиды выделены единичные представители родов Shewanella, Geobacter, Desulfuromonas и Desulfuromusa [11, 24], способные восстанавливать железо при температуре около 4°С. Все эти бактерии принадлежат к одному семейству Geobacteraceae и восстанавливают Fe(III), окисляя ацетат и другие органические субстраты до диоксида углерода.
Цель исследования - описание процессов, связанных с образованием двухвалентной формы железа в современных тундровых почвах и многолетнемерзлых отложениях. В рамках поставленной цели решали следующие задачи:
- оценка содержания и распределения несиликатного железа, в том числе его подвижной формы, в тундровых почвах и подстилающих ММО;
- представление и обсуждение результатов сравнительного метагеномного анализа современной тундровой почвы и трех образцов многолетнемерзлых отложений позднеплейстоценового возраста, позволяющего выявить гены белков и ферментов, связанных с циклом железа, а также гены бактерий, участвующих в процессе восстановления железа;
- выявление процесса микробного восстановления железа в лабораторных условиях.
ОБЪЕКТЫ ИССЛЕДОВАНИЯ
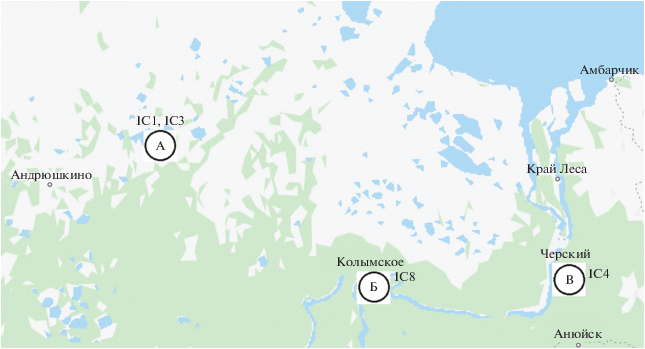
Ключевой участок, где проводили почвенные исследования, расположен в западной части Колымской низменности в непосредственной близости от северной границы тайги в районе Олерского неотектонического поднятия (рис. 1, А). На этом участке, расположенном в среднем течении р. Алазеи, мощность пород едомной свиты не превышает 10-20 м. Ниже в разрезе залегают плиоцен-нижнеплейстоценовые озерно-аллювиальные суглинистые отложения олерской свиты, подстилаемые поздненеогеновыми эпикриогенными песками томус-ярской свиты. Наличие небольших термоэрозионных цирков на крутых склонах едомных останцов говорит об их непрекращающемся разрушении.
Рис. 1.
Районы исследования: А - р. Алазея, Б - устье р. Омолон, В - протока Амболиха (карты Google).
Зональная растительность на водоразделах представлена кустарничково-травянисто-зеленомошными ассоциациями. Бугорковатый микрорельеф и интенсивно протекающее пятнообразование определяют структуру почвенного покрова.
Изучение содержания несиликатного железа проводили в почвах трех катен, включающих крутой (25°-30°) склон северной экспозиции, пологий (7°-15°) склон северной экспозиции и пологий склон южной экспозиции в бассейне р. Алазеи. Помимо этого, исследовали керны из скважин, заложенных в трех географических точках: на ключевом участке в бассейне р. Алазеи (Al3-15 и 2-98), на берегу протоки Амболихи (4-07) и в районе устья р. Омолон (2-07). Скважина 4-07, пробуренная в районе протоки Амболихи, вскрыла многолетнемерзлые отложения старичного озера возраста 34 тыс. лет [20], подстилаемые грубыми пескам бегуновской свиты. Отложения, вскрытые скважиной 2-07 в районе устья р. Омолон, были представлены сильнольдистыми суглинками позднеплейстоценового ледового комплекса.
Для выявления генов микроорганизмов и белков, связанных с процессами восстановления железа, получили метагеномы и провели микробиологические исследования.
МЕТОДЫ ИССЛЕДОВАНИЯ
Колонковое бурение скважин осуществляли установкой УКБ 12/25 без продувки и промывки буровыми растворами с соблюдением требований стерильности, как описано ранее [22].
Отбор почвенных образцов для определения несиликатного железа проводили по основным горизонтам из профилей под задернованными бугорками и из мерзлого дна разреза. В случае разреза 518-07 образцы из неоттаявшей толщи отбирали до глубины 15 см от подошвы сезонноталого слоя, глубже которой залегал чистый лед, по-видимому, полигонально-жильной природы.
Определение потери при прокаливании, рН, гидролитической кислотности, содержания обменных оснований и СО2 карбонатов проводили в химико-аналитическом комплексе ИФХиБПП РАН. Определение валового углерода выполняли в ЦКП “Лаборатория радиоуглеродного датирования и электронной микроскопии” Института географии РАН методом сухого сжигания. Содержание органического углерода рассчитывали по разности между валовым его содержанием и содержанием углерода карбонатов, определенным ацидиметрическим методом.
Определение железа. Фракционный состав несиликатного железа определяли по схеме Зонна [3] в лаборатории отдела географии и эволюции почв Института географии РАН. В полевых условиях определяли содержание подвижных форм железа, извлекаемых 0.1 н. серной кислотой из свежих образцов [1]. Аналогичное определение мобильного железа в образцах многолетнемерзлых отложений осуществляли в лаборатории, куда буровые керны доставляли в мерзлом состоянии. Для определения подвижного железа использовали методику, разработанную для природных вод [5] с раздельным титрованием трех- и двухвалентных форм 0.01 н. трилоном Б. В лабораторных условиях содержание трехвалентного железа определяли путем титрования, а двухвалентного - путем колориметрического определения с 0.5% раствором α-α'-дипиридила по методике Виригиной-Аринушкиной (ГОСТ 27395-87). Для почв крутого северного склона (разрезы 508-07, 509-07 и 510-07), где содержание оксалаторастворимого железа по Тамму и кислоторастворимого железа определяли для одних и тех же профилей, можно точно сказать, какую часть подвижные формы составляют от общего содержания аморфного железа. Для плакоров и пологих склонов, где соответствующие анализы выполняли для почв разных катен, доля подвижного железа может быть оценена только приблизительно.
Молекулярные методы. Для выделения тотальной ДНК из образцов почвы и мерзлой породы использовали набор PowerSoil® (MO BIO Laboratories Inc., USA), очистку и концентрирование полученной ДНК осуществляли с помощью набора Genomic DNA Clean and Concentrator® Kit (Zymo Research Corporation, USA). Секвенирование метагенома выполняли в центре геномных исследований г. Барселоны (Centre for Genomic Regulation, Barcelona, Spain) на платформе Illumina HiSeq 2000™ и в университете Теннесси (США) на платформе Illumina MiSeq. Необработанные данные секвенирования, около 19.8 Гб, что составляет 143.7-150.0 млн последовательностей со средней длиной 150 пар оснований, загружали на сервер MG-RAST [16] для получения геномных аннотаций. Более 95% последовательностей прошли контроль качества.
Микробиологические методы. Для получения накопительных культур железовосстанавливающих бактерий (ЖВБ) использовали модифицированную среду [29], в которую добавляли (мл/л) раствор микроэлементов [23] - 1.0, раствор витаминов - 10.0 [28] и дрожжевой экстракт “Difco”- 0.002 г/л. В качестве источника углерода и донора электронов использовали ацетат натрия (конечная концентрация 20 мМ), в качестве акцептора электронов - цитрат Fe(III) (конечная концентрация 10 мМ). Приготовление минеральной среды и культивирование микроорганизмов проводили в строго анаэробных условиях под N2 (100% в газовой фазе), рН среды 7.0-7.2. Инкубировали в темноте при 5 и 20°С в течение 40 сут. В качестве химического контроля использовали минеральную среду без инокуляции. Восстановление Fe(III) определяли колориметрическим методом по образованию стабильного окрашенного комплекса двухвалентного железа с феррозином [26].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Строение и химические свойства изучаемых почв. Почвы ключевого участка в бассейне р. Алазеи (рис. 1, А) имеют средне- или тяжелосуглинистый состав и характеризуются отсутствием элювиально-иллювиальной дифференциации. Под положительными элементами микрорельефа типично наличие гумусовых затеков в верхней части почвенного профиля и криотурбационных включений торфа - в средней и нижней его частях. Наиболее дренированные позиции на вершинах водоразделов и в верхней части пологих склонов занимают криоземы надмерзлотно-глееватые. В средней части пологого склона северной, в нижней части пологого склона южной экспозиции и на крутом склоне из сменяют глееземы криотурбированные. В верхней части профиля склоновых почв обычно присутствуют грубогумусовые (АО) или перегнойные (Н) горизонты. На вершине водоразделов из-за частых циклов пятнообразования подобные горизонты не всегда успевают сформироваться, чаще органо-аккумулятивная часть представлена оторфованными гумусово-слаборазвитыми (Wт) горизонтами. Верхняя и средняя части профиля в криоземах представлены серией неоглеенных или слабооглеенных горизонтов (Bhi, Вcrm, B@ и др.), нижняя - глееватыми горизонтами (Bg). Наконец, в надмерзлотной части профиля, в интервале глубин 50-70 см присутствует глеевый горизонт (Gт), частично вытаивающий из мерзлоты в наиболее теплые летние сезоны. Ему свойственны сизая или серо-сизая окраска с редкими желтыми или охристыми примазками, высокая влажность, часто тиксотропность, резкий переход от вышележащих глееватых горизонтов, отсутствие структуры, плотное сложение, высокое содержание гумуса, наличие субгоризонтальных торфяных линз и прослоев, часто двух-четырех, залегающих параллельно друг другу. Криоземы и глееземы различаются соотношением неоглееной и оглеенной частей профиля. Если в криоземах неоглеенные горизонты занимают от двух третей до половины деятельного слоя, то в глееземах их мощность уменьшается, и в вариантах у подножья склона составляет всего 3-6 см.
Верхним горизонтам почв водоразделов свойственна сильнокислая реакция среды (табл. 1), глубже она сменяется среднекислой. Величина гидролитической кислотности снижается по профилю от 6-19 до 2-3 × 10-2 моль/кг, а степень насыщения почвенного поглощающего комплекса (ППК) возрастает с 25-46 до 68-80%. Почвы пологих склонов отличаются господством сильнокислой среды по всему профилю, кроме глеевого горизонта, где она среднекислая. Насыщенность ППК основаниями повышается по профилю с 25-30 до 52-73%. В составе обменных оснований здесь, как и на плакоре, преобладает кальций. Почвы крутых склонов характеризуются господством среднекислой среды, которая в нижней частим профиля может меняться на слабокислую, а в глеевом горизонте - на нейтральную. Степень насыщения ППК основаниями невелика лишь в органогенных горизонтах, а в минеральных составляет 61-96%. В составе обменных оснований преобладают кальций и магний.
Таблица 1.
Химическое состояние тундровых почв бассейна р. Алазеи
| Горизонт | Глубина, см | Сорг, % | рН | Гидролитическая кислотность, 10-2 моль/кг | Содержание обменных оснований, 10-2 моль/кг | Степень насыщенности ППК, % | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| H2O | KCl | Ca | Mg | Na | K | |||||
| Разрез 502-07, вершина водораздела | ||||||||||
| Wт | 0-4 | 12.52 | 4.2 | 3.6 | 18.80 | 5.62 | 4.38 | 0.35 | 0.85 | 24.78 |
| Bhi | 4-8 | Не опр. | 4.9 | 3.8 | 6.30 | 4.72 | 4.28 | 0.42 | 0.49 | 46.19 |
| В | 8-17 | 1.47 | 5.8 | 4.7 | 2.70 | 6.96 | 4.90 | 0.36 | 0.26 | 70.83 |
| B@ | 17-36 | 1.23 | 6.0 | 4.9 | 2.00 | 9.66 | 4.94 | 0.30 | 0.30 | 79.81 |
| Bg1 | 36-41 | 1.19 | 5.9 | 4.8 | 2.08 | 6.88 | 4.68 | 0.26 | 0.37 | 75.47 |
| Bg2 | 41-51 | 1.94 | 5.7 | 4.6 | 2.96 | 6.94 | 4.40 | 0.29 | 0.46 | 68.46 |
| Gт | 51-64 | 3.15 | 5.5 | 4.4 | 4.09 | 6.96 | 3.90 | 0.27 | 0.59 | 60.58 |
| Разрез 504-07, середина пологого южного склона | ||||||||||
| AO | 0-4 | 13.59 | 4.3 | 3.5 | 19.81 | 6.26 | 4.12 | 0.63 | 0.75 | 24.90 |
| Вcrm | 4-7 | Не опр. | 4.7 | 3.5 | 6.71 | 4.50 | 2.90 | 0.23 | 0.33 | 38.85 |
| В@ | 7-21 | 1.61 | 5.3 | 4.1 | 3.89 | 5.30 | 4.06 | 0.22 | 0.28 | 57.11 |
| Bg1 | 23-30 | 1.21 | 5.4 | 4.3 | 2.93 | 4.88 | 3.80 | 0.25 | 0.21 | 62.08 |
| Bg2 | 30-46 | 1.76 | Не опр. | Не опр. | 2.76 | 6.56 | 4.24 | 0.28 | 0.29 | 68.43 |
| Gт | 58-64 | 4.60 | 5.5 | 4.6 | 4.47 | 9.02 | 4.14 | 0.27 | 0.44 | 61.97 |
| Разрез 507-07, нижняя часть пологого южного склона | ||||||||||
| АО | 0-6 | 14.36 | 4.2 | 3.3 | 19.96 | 5.48 | 3.92 | 0.54 | 0.77 | 23.16 |
| Вcrm | 6-9 | 1.32 | 5.2 | 4.0 | 3.8 | 4.78 | 3.66 | 0.20 | 0.30 | 55.38 |
| Bg2 | 18-36 | 1.52 | 5.3 | 4.0 | 3.92 | 6.10 | 3.62 | 0.22 | 0.20 | 57.35 |
| Bg3 | 38-50 | 2.26 | 5.3 | 4.2 | 5.06 | 6.18 | 3.86 | 0.32 | 0.25 | 52.44 |
| Gт | 51-59 | 1.41 | 5.8 | 4.9 | 1.90 | 6.32 | 3.06 | 0.20 | 0.36 | 73.40 |
| Разрез 508-07, верхняя часть крутого северного склона | ||||||||||
| H | 0-2 | 8.71 | 5.2 | 4.5 | 17.98 | 7.66 | 7.42 | 0.39 | 1.44 | 34.28 |
| Bcrm | 3-6 | Не опр. | 5.5 | 4.5 | 2.16 | 4.58 | 4.44 | 0.29 | 0.42 | 70.78 |
| B@ | 6-14 | 0.96 | 5.7 | 4.8 | 2.05 | 5.24 | 4.40 | 0.31 | 0.27 | 72.48 |
| Bg1 | 14-35 | 0.79 | 5.9 | 4.8 | 1.74 | 5.56 | 4.40 | 0.23 | 0.23 | 75.74 |
| Bg2 | 22-45 | 1.66 | 6.0 | 5.1 | 1.95 | 6.78 | 3.20 | 0.22 | 0.28 | 73.85 |
| Bg3 | 45-60 | Не опр. | 6.2 | 5.3 | 1.22 | 5.46 | 4.06 | 0.24 | 0.29 | 81.20 |
| Gт | 60-69 | 0.27 | 7.0 | 6.1 | 0.20 | 5.12 | 3.90 | 0.23 | 0.30 | 96.13 |
Распределение форм железа по почвенным профилям. В почвах склонов наблюдается преимущественно аккумулятивное распределение содержания общего несиликатного (дитиониторастворимого) железа и во всех случаях - аккумулятивное распределение его окристаллизованной формы. Такая картина определяется, в первую очередь, биогенным накоплением элемента в верхней части профиля и выносом его из нижней части, в особенности из надмерзлотного горизонта, в результате глеевого процесса. Средневзвешенное содержание дитиониторастворимого железа по профилям почв пологого склона изменяется в пределах 1.90-2.10%.
Содержание аморфного (оксалаторастворимого) железа обычно распределено бимодально с минимумом в средней части профиля. В нижних глееватых и, тем более, в глеевом горизонте доля этой фракции в составе несиликатного железа возрастает. Наиболее резкое расширение отношения концентраций оксалато- и дитиониторастворимого железа ([Feox]/[Fedt], коэффициента Швертмана) в пределах минеральной толщи наблюдается в почвах крутого склона, для которых значения этого показателя сверху вниз по профилю расширяются от 0.37-0.42 до 0.72-0.93. В почвах пологих склонов значения коэффициента Швертмана в верхних и нижних минеральных горизонтах различаются не столь сильно: 0.34-0.37 и 0.49-0.59 соответственно. Лишь у подножья масштаб увеличения этого показателя по профилю (с 0.47 до 0.83) приближается к таковому в случае почв крутого склона. В органо-аккумулятивных горизонтах значения коэффициента Швертмана сравнительно невелики (0.37-0.61), что может быть связано с низкими температурами в верхней части профиля, способствующими дегидратации и кристаллизации аморфных соединений железа. В нижней части пологого и крутого склонов (разрезы 507-07 и 510-07), где создаются оптимальные условия для снегонакопления, отношение концентраций для органо-аккумулятивных горизонтов (0.51-0.61) заметно шире, чем в почвах других позиций (0.37-0.44). Высокое содержание пирофосфаторастворимого железа по Баскомбу (0.47-0.81%) имеет место в органо-аккумулятивных горизонтах, в пределах минеральной части профиля его значения изменяются в пределах 0.07-0.33%.
В водораздельной почве (разрез 502-07) наблюдается небольшая аккумуляция дитиониторастворимого железа в органо-аккумулятивном горизонте, а в пределах минерального профиля - увеличение содержания сверху вниз с резким уменьшением в надмерзлотной зоне. Средневзвешенное содержание его по профилю составляет 1.89%. Распределение оксалаторастворимого железа повторяет таковое для дитиониторастворимого, но в более сглаженном виде. Аккумуляция в горизонте Wт выглядит не столь отчетливо, снижение содержания в надмерзлотном горизонте здесь не так заметно. Величина коэффициента Швертмана меняется по профилю в узких пределах (от 0.43 до 0.53), а в глеевом горизонте резко увеличивается (до 0.86). Пирофосфаторастворимое железо распределено бимодально, его концентрационные максимумы приурочены к горизонтам с высоким содержанием органического вещества (табл. 1, 2): органо-аккумулятивному и надмерзлотному глеевому. Здесь на долю железа в комплексе с гумусовыми веществами приходится примерно половина от общего содержания его аморфных форм. В минеральной части профиля (рис. 2) содержание пирофосфаторастворимого железа имеет тенденцию к увеличению в глубь по профилю.
Таблица 2.
Фракционный состав несиликатного железа в тундровых почвах бассейна р. Алазеи (рассчитано на прокаленную почву), %
| Гори-зонты | Глубина, см | Содержание Fe | Коэффи-циент Швертмана | |||||
|---|---|---|---|---|---|---|---|---|
| дитио-нит- раство-римого | окристал-лизован-ного | аморфного общего (оксалато- раство-римого) | аморфного железоор-ганического (пирофосфато-расраство-римого) | аморфного минераль-ного | подвижного кислото-растворимого | |||
| Разрез 502-07, вершина водораздела | ||||||||
| Wт | 0-4 | 1.33 | 0.65 | 0.68 | 0.34 | 0.34 | Не опр. | 0.51 |
| В | 8-17 | 1.13 | 0.53 | 0.60 | 0.11 | 0.49 | » | 0.53 |
| B@ | 17-36 | 1.56 | 0.88 | 0.68 | 0.11 | 0.57 | » | 0.43 |
| Bg1 | 36-41 | 1.70 | 0.92 | 0.78 | 0.12 | 0.66 | » | 0.46 |
| Bg2 | 41-51 | 1.84 | 0.88 | 0.98 | 0.17 | 0.81 | » | 0.53 |
| Gт | 51-64 | 0.80 | 0.11 | 0.69 | 0.36 | 0.33 | » | 0.86 |
| Разрез 504-07, середина пологого южного склона | ||||||||
| AO | 0-4 | 1.90 | 1.20 | 0.70 | Не опр. | Не опр. | Не опр. | 0.37 |
| В@ | 7-21 | 1.41 | 0.89 | 0.52 | » | » | » | 0.37 |
| Bg1 | 23-30 | 1.29 | 0.83 | 0.46 | » | » | » | 0.36 |
| Bg2 | 30-46 | 1.34 | 0.73 | 0.61 | » | » | » | 0.46 |
| Gт | 58-64 | 1.36 | 0.69 | 0.67 | » | » | » | 0.49 |
| Разрез 505-07, нижняя часть пологого южного склона | ||||||||
| Wт | 0-3 | 1.51 | 0.90 | 0.61 | Не опр. | Не опр. | Не опр. | 0.40 |
| В | 5-15 | 1.59 | 1.05 | 0.54 | » | » | » | 0.34 |
| Bg2 | 15-20 | 1.49 | 0.92 | 0.57 | » | » | » | 0.38 |
| Bg3 | 21-47 | 1.61 | 0.95 | 0.66 | » | » | » | 0.41 |
| Gт | 47-57 | 1.07 | 0.44 | 0.63 | » | » | » | 0.59 |
| Разрез 507-07, нижняя часть пологого южного склона | ||||||||
| АО | 0-6 | 1.74 | 0.86 | 0.88 | 0.56 | 0.32 | Не опр. | 0.51 |
| Вcrm | 6-9 | 1.32 | 0.70 | 0.62 | 0.17 | 0.45 | » | 0.47 |
| Bg2 | 18-36 | 1.47 | 0.75 | 0.72 | 0.18 | 0.54 | » | 0.49 |
| Bg3 | 38-50 | 1.23 | 0.54 | 0.69 | 0.23 | 0.46 | » | 0.56 |
| Gт | 51-59 | 0.76 | 0.13 | 0.63 | 0.22 | 0.41 | » | 0.83 |
| Разрез 508-07, верхняя часть крутого северного склона | ||||||||
| АО | 0-5 | 1.36 | 0.85 | 0.51 | Не опр. | Не опр. | Не опр. | 0.37 |
| G | 2-14 | 1.77 | 0.99 | 0.78 | » | » | 0.335 | 0.44 |
| B@ | 6-14 | 1.10 | 0.73 | 0.37 | » | » | 0.031 | 0.34 |
| Bg1 | 14-35 | 0.95 | 0.57 | 0.38 | » | » | 0.043 | 0.40 |
| Bg2 | 22-47 | 1.14 | 0.63 | 0.51 | » | » | 0.027 | 0.45 |
| Gт | 55-69 | 0.69 | 0.05 | 0.64 | » | » | 0.043 | 0.93 |
| Разрез 509-07, середина крутого северного склона | ||||||||
| АО | 0-5 | 1.54 | 0.96 | 0.58 | Не опр. | Не опр. | НО | 0.37 |
| B@ | 9-14 | 1.28 | 0.75 | 0.53 | » | » | 0.015 | 0.41 |
| Bg1 | 14-35 | 1.12 | 0.70 | 0.42 | » | » | 0.047 | 0.38 |
| Bg2 | 35-57 | 1.17 | 0.70 | 0.47 | » | » | 0.045 | 0.40 |
| Gт | 62-74 | 0.93 | 0.24 | 0.69 | » | » | 0.712 | 0.74 |
| Разрез 510-07, нижняя часть крутого северного склона | ||||||||
| AO | 2-6 | 1.67 | 0.66 | 1.01 | Не опр. | Не опр. | 0.242 | 0.61 |
| B@ | 11-28 | 1.21 | 0.70 | 0.51 | » | » | 0.048 | 0.42 |
| Bg | 32-41 | 1.24 | 0.27 | 0.97 | » | » | 0.228 | 0.79 |
| Gт | 41-53 | 1.07 | 0.29 | 0.78 | » | » | 0.553 | 0.72 |
В пределах склона наблюдается увеличение содержания оксалаторастворимого железа, прежде всего, для глееватых горизонтов, вниз по катене, то есть в направлении нарастания морфохроматических признаков оглеения. В нижней части склона наблюдается большее содержание пирофосфаторастворимого железа по сравнению с водораздельным профилем из-за увеличения доли органических форм в составе аморфного Fe2O3.
В почвах крутого склона средневзвешенное по профилю содержание несиликатного (дитиониторастворимого) железа составляет 1.57, 1.67 и 1.78% для разрезов 508-07, 509-07 и 510-07 соответственно, что в среднем на 15% меньше, чем в почвах водораздела и пологого склона. В верхней неоглееной части профиля уменьшение общего содержания несиликатного железа по сравнению с плакорным вариантом происходит за счет аморфной формы, а для горизонтов, затронутых оглеением, - как аморфной, так и окристаллизованной. На фоне общего обезжелезнения почв крутых склонов также наблюдается тенденция к перераспределению вниз по катене несиликатного железа, в первую очередь оксалаторастворимого. Поэтому почвы верхней и средней частей крутых склонов обезжелезнены в наибольшей степени. В нижней же части склона меняется соотношение между аморфным и окристаллизованным железом, коэффициент Швертмана увеличивается до 0.42-0.79.
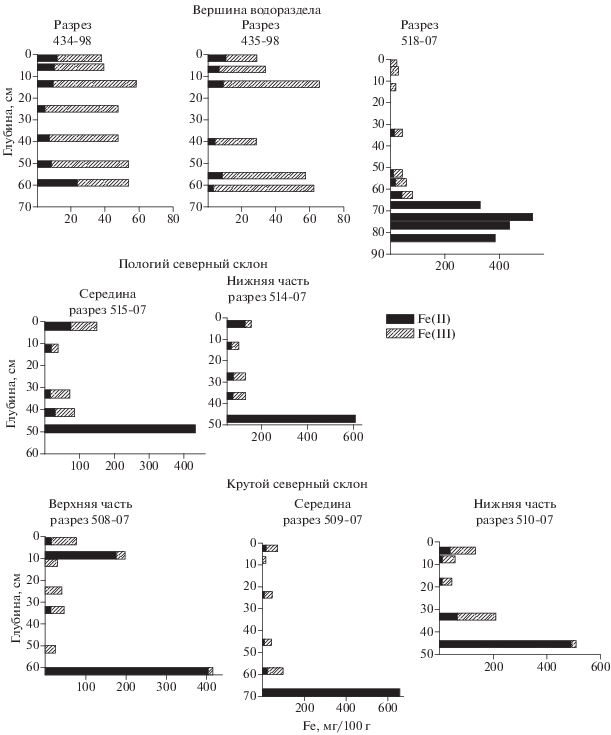
Содержание подвижного железа, экстрагируемого 0.1 н. H2SO4 вытяжкой из свежих образцов, по профилю склоновых почв распределено бимодально (рис. 2). Верхний максимум приурочен к перегнойным или грубогумусовым горизонтам (71-148 мг Fe/100 г), а в профиле 508-07 - к горизонту поверхностного оглеения (197 мг Fe/100 г). Нижний, гораздо более значительный, максимум (417-659 мг Fe/100 г) соответствует надмерзлотным глеевым горизонтам. Помимо этого, экстрагируемость железа существенно меняется при переходе от верхних минеральных Вhi, Bcrm, B@ и др. к глееватым горизонтам Bg и резко изменяется при переходе от них к глеевым горизонтам Gт, где все или почти все экстрагируемое железо представлено двухвалентной формой. Увеличение содержания кислоторастворимого железа вниз по профилю хорошо согласуется с нарастанием проявления морфохроматических признаков подвижности этого элемента. Отношение [Fe(II)]/[Fe(III)] в верхней части профиля может быть несколько расширено, что отчасти определяется двухвалентной формой содержания железа в неразложившихся растительных тканях. Наибольших значений (>1.00) оно достигает в горизонте поверхностного оглеения (разрез 508-07), в грубогумусовом горизонте АО (разрез 514-07) и перегнойном горизонте Н (разрез 515-07). В минеральной толще оно изменяется в широких пределах (0.08-0.90), а в надмерзлотном горизонте Gт - стремится к бесконечности.
Доля кислоторастворимого железа от оксалаторастворимого в пределах минеральных горизонтов несколько увеличивается вниз по профилю и резко увеличивается при переходе к глеевому горизонту. Для органо-аккумулятивных горизонтов она примерно может быть оценена в 17-29%, для горизонта поверхностного оглеения - в 43%, для неоглееных минеральных горизонтов - в 3-10%, для глееватых - в 5-24%, а для глеевых - в 67-100%. В последнем случае все аморфное железо надмерзлотной части некоторых профилей находится в виде подвижных соединений Fe(II). Распределение кислоторастворимого железа по профилю плакорных почв отличается от такового в склоновых вариантах отсутствием верхнего максимума, что связано с особенностями органо-аккумулятивной частью профиля на водоразделах, представленной здесь в большинстве случаев гумусово-слаборазвитыми горизонтами. Возрастание содержания вниз по профилю по сравнению со склоновыми почвами выглядит более сглажено. В неоттаявшей части глеевого горизонта содержание подвижного железа еще выше, чем в оттаявшей. Отношение [Fe(II)]/[Fe(III)] в неоглеенных горизонтах изменяется в пределах 0.10-0.60, а в глееватых - в пределах 0.18-1.13. Доля суммарного кислоторастворимого железа от оксалаторастворимого в пределах профиля может быть оценена в 4-12%, а в глеевом горизонте - 52-83%.
Для почв, как пологих, так и крутых склонов, сверху вниз по мезорельефу имеет место увеличение содержания суммарного кислоторастворимого железа, согласующееся с усилением морфологических признаков глея. Различия по этому показателю между профилями внутри катен определяются как двухвалентной, так и трехвалентной формой элемента, но роль двухвалентной формы существенно выше, что особенно заметно в случае пологого склона. Наиболее существенные различия здесь наблюдаются для надмерзлотных глеевых горизонтов, где все подвижное железо двухвалентно.
Соответствие между увеличением содержания кислоторастворимого железа вниз по профилю почв так же, как и вниз по катенам, и нарастанием морфохроматических признаков подвижности железа, на наш взгляд, говорит в пользу того, что последние отвечают протекающим в современных условиях процессам глееобразования. Несмотря на преобладание окислительных условий на дренированных плакорах, оглеение входит в комплекс почвообразовательных процессов, формирующих профиль.
В нижней части пологих склонов в связи с их большим увлажнением глеевый процесс становится господствующим. Сверху вниз по мезорельефу в почвах происходит увеличение содержания оксалаторастворимого и кислоторастворимого железа, а также доли двухвалентной формы в составе последнего. В этом же направлении уменьшаются значения рН, возрастает потенциальная кислотность, снижается насыщенность ППК основаниями.
Почвы крутых склонов развиваются в условиях непрекращающейся плоскостной эрозии, определяющей их молодость. Им свойственны более холодные тона фоновой окраски и меньшая кислотность, которую в данном случае можно рассматривать в качестве показателя степени педогенной трансформации исходной породы. В нижних горизонтах реакция среды может быть близкой к нейтральной или нейтральной. Их обезжелезнение - снижение содержания дитиониторастворимого железа по сравнению с почвами плакоров и пологих склонов, - по нашим представлениям, связана с вымыванием в условиях свободного дренажа значительной части подвижного железа, в больших количествах содержащегося в едомных и тем более олерских отложениях. В пределах склона, особенно нижней его части также происходит перераспределение оксалато- и кислоторастворимого железа. Эти почвы испытывают постоянный приток влаги из-за вытаивания неглубоко расположенных ледяных жил, поэтому на наиболее крутых склонах создаются условия наибольшего переувлажнения и развиваются наиболее оглеенные разности. Схожий тип катен ранее описан нами на мысу Медвежьем [8].
Наиболее интересным и сложным для интерпретации объектом является глеевый горизонт в надмерзлотной части большинства описанных почв. Помимо морфологического своеобразия этот горизонт отличают обезжелезнение (потеря общего несиликатного и окристаллизованного железа); господство аморфного железа над окристаллизованным; высокое содержание комплексных железоорганических соединении; преобладание, иногда полное, мобильных форм в составе аморфного Fe2O3; почти всегда исключительно двухвалентный состав подвижного железа, указывающий на постоянство восстановительных условий; большое содержание водорастворимого и обменного Fe(II), дающего интенсивную качественную реакцию в природных условиях.
Несоответствие наличия восстановленного глеевого горизонта мезофильной тундровой растительности, особенно на дренированных водоразделах и в верхней части пологих склонов; резкий переход к горизонту Gт от вышележащей глееватой толщи; законсервированность нижней его части в многолетней мерзлоте позволяют предполагать реликтовую природу этого горизонта. Однако повсеместное распространение указывает на то, что возраст его не старше возраста современного мезорельефа. Есть основания считать, что горизонт Gт генетически связан с органогенным надмерзлотно-аккумулятивным горизонтом, описанным в тундровой зонe Колымской низменности [2, 4]. Последний включен в этот глеевый горизонт, и мы склонны рассматривать их как единое целое.
Как показывают наши наблюдения в бассейне р. Алазеи (с 1998 г.) и на территории Колымской низменности вообще (с 1984 г.) глеевый горизонт может годами, возможно десятилетиями, существовать в замерзшем состоянии, за пределами слоя сезонного протаивания и сферы действия современных процессов почвообразования. Нерегулярные и кратковременные вхождения Gт в состав деятельного слоя вызывают некоторую трансформацию верхней его части, выражающуюся в частичном изменении окраски, в появлении желто-охристых примазок, в формировании даже окисно-глеевого горизонта (Gox) над ним (профили 434-98 и 435-98), а также в несколько пониженном содержании кислоторастворимого железа в протаивающей части горизонта Gт по сравнению с непротаивающей. Однако полной окислительной деградации его не происходит. В качестве одной из причин столь высокой окислительно-восстановительной буферности глеевого горизонта, скорее всего, выступают как раз включения больших объемов торфяного материала турбационной природы.
Сопоставление экстрагируемости железа 0.1 н. H2SO4, определенной в холодном 1998 и теплом 2007 гг. на соседних плакорах, показало, что увеличение мощности деятельного слоя и вытаивание глеевого горизонта изменяют конфигурацию профильного распределения подвижного железа, резко увеличивают его запас в почвенном профиле и смещают соотношение двух- и трехвалентной форм (рис. 2).
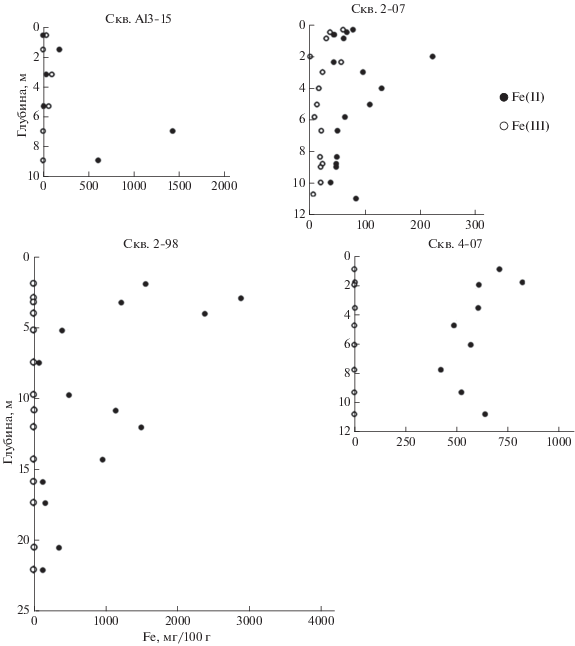
Распределение форм железа в многолетнемерзлых отложениях. На ключевом участке бассейна р. Алазеи содержание подвижного железа в верхней части геологического разреза, представленной ММО едомной свиты (скважина Al 3-15), изменяется в широких пределах. Серые и сизо-серые горизонты, весьма богатые железом (185-1432 мг Fe/100 г), представленном почти исключительно двухвалентной формой, чередуются здесь с неоднородными по окраске горизонтами со сравнительно невысоким его содержанием (39-138 мг Fe/100 г), где преобладает трехвалентная форма - отношение [Fe(II)]/[Fe(III)] составляет 0.08-0.38. В большинстве случаев значения этого показателя больше, чем в минеральных горизонтах современных почв, исключая надмерзлотный глеевый. Подстилающие отложения аллювиальной олерской свиты, вскрытые скважиной 2-98, характеризуются весьма высокой экстрагируемостью железа (до 2366-2865 мг Fe/100 г) и практически полным отсутствием трехвалентной формы в его составе, что указывает на господство восстановительных условий по всей изученной толще.
Определение содержания двух- и трехвалентного железа в мерзлых отложениях скважин 2-07 и 4-07 выявило, что в обеих Fe(II) превалирует над Fe(III), причем в отложениях голоценовой поймы и позднеплейстоценового старичного озера (скважина 4-07) трехвалентное железо полностью отсутствовало, а в породах едомной свиты (ледового комплекса) (скважина 2-07) присутствовало в небольших количествах (рис. 3). В этой же скважине во всех исследованных образцах фиксировался биогенный метан, тогда как в скважине 2-07 его не обнаружено [20]. Преобладание двухвалентной формы подвижного железа над трехвалентной в большинстве изученных образцов ММО указывает на превалирование процесса редукции железа и позволяет предполагать наличие ЖВБ в составе микробных сообществ многолетней мерзлоты и специфических белков, связанных с их жизнедеятельностью.
Анализ метагеномов. Одним из наиболее информативных методов оценки разнообразия природных микробных сообществ являются метагеномные исследования. Этот же подход позволяет оценить функциональные гены белков, ответственных за основные биогеохимические процессы. Анализ аннотированных метагеномов выявил, что в них наиболее широко представлены микроорганизмы 9 бактериальных и одного архейного филума: Proteobacteria, Actinobacteria, Firmicutes, Chloroflexi, Bacteroidetes, Acidobacteria, Cyanobacteria, Verrucomicrobia, Planctomycetes и Euryarchaeota (рис. 4). Количество микроорганизмов, относящихся к другим филумам, было менее 1%. Гены микроорганизмов, относящихся к филуму Proteobacteria, доминируют в образцах старичного озера (IC4) и глееватом горизонте Bg (глубина 50-60 см) водораздельного криозема надмерзлотно-глееватого тяжелосуглинистого (IC1), Actinobacteria более широко представлены в отложениях позднеплейстоценового ледового комплекса из скважин 2-07 и Al3-15 (образцы IC8 и IC3)). Известно, что большинство ЖВБ относится к филуму Proteobacteria [15]. Нами показано, что в образце IC4 присутствует больше микроорганизмов, связанных с метановым циклом, чем в образце из ледового комплекса (IC8) [20]. Анализ четырех метагеномов позволил выявить и сравнить количество последовательностей генов 16S рРНК, принадлежащих трем основным родам железовосстанавливающих бактерий: Shewanella, Geobacter и Desulfuromonas (рис. 5, А). В отложениях старичного озера (IC4) и глееватом горизонте современной тундровой почвы (IC1) эти микроорганизмы были представлены большим количеством генов 16S рРНК, чем в отложениях ледового комплекса (IC8 и IC3).
Рис. 4.
Анализ метагеномов на уровне филума (данные нормализованы). Показаны только филумы, содержание которых превышало 1%.
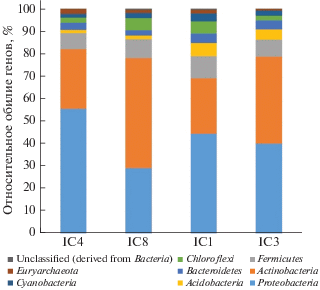
Рис. 5.
Количество 16S рРНК генов железовосстанавливающих микроорганизмов в метагеномах (А); образование двухвалентного железа в накопительных культурах при разных температурах (Б). Образцы, представленные на рис. 5, Б, отобраны из скв. 4-07, протока Амболихи (рис. 1, В, табл. 3): М2 с глубины 1.4 м, М4 с 17 м и М5 с 22 м.
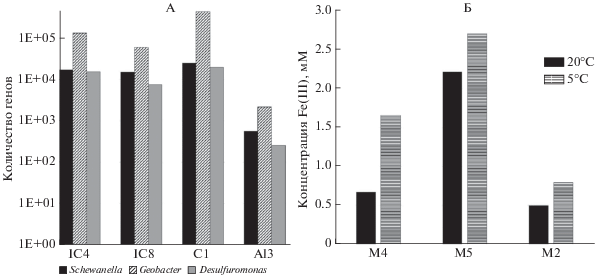
Таблица 3.
Места отбора образцов на микробиологические и метагеномные исследования
| Образец | Район исследования | Место расположения и скважины | Глубина, м | Описание |
|---|---|---|---|---|
| IC1 | А (69.338869° N, 154.996948° E) |
р. Алазея | 0.5-0.6 | Современная почва (криозем надмерзлотно-глееватый тяжелосуглинистый, горизонт Bg) |
| IC3 | р. Алазея, скв. Al3-15 |
3.5 | Оторфованный горизонт в позднеплейстоценовом ледовом комплексе | |
| IC8 | Б (68.693947° N, 158.710142° E) |
р. Омолон, скв. 2-2007 |
16 | Ледовый комплекс, поздний плейстоцен, суглинок |
| IC4 | В (68.711290° N, 161.434493° E) |
Протока
Амболиха, скв. 4-2007 |
1.4 17.5 22.0 |
Старичное озеро, поздний плейстоцен, суглинок |
Примечание. Районы А, Б и В - представлены на рис. 1.
Как известно, железо необходимо для всех живых организмов, включая бактерии. Этот элемент входит в состав белков, осуществляющих важнейшие метаболические функции. В составе исследованных метагеномов выявлены гены, кодирующие белки, участвующие в ассимиляционных и диссимиляционных процессах восстановления и окисления железа (табл. 4). Видно, что в IC1 (глееватый горизонт современной тундровой почвы) по сравнению с образцом IC3 из подстилающих ММО превалируют белки, ответственные за процессы цикла железа, включая транспорт этого элемента в микробную клетку. Сравнивая белки в образцах IC4 и IC8, можно заметить, что набор белков, необходимых для восстановления железа, представлен большим количеством копий в IC4, чем в IC8, тогда как белки ассимиляционной железоредукции представлены большим количеством генов в IC8, чем в IC4. Эти данные являются еще одним подтверждением потенциал задающей роли биогеохимических реакций, осуществляемых микроорганизмами в процессе формирования почв и осадочных отложений.
Таблица 4.
Функциональные белки, связанные с циклом железа (число белок-кодирующих генов в метагеномах современной почвы и образца многолетнемерзлой породы)
| Номен-клатура | Функциональные белки | Р. Алазея | Р. Амболиха и Омолон | ||
| IC1 | IC3 (Al3-15) | IC4 (4-07) | IC8 (2-07) | ||
| Fe-содержащие белки | |||||
| EC 1.2.7.1 | Pyruvate: ferredoxin oxidoreductase | 27 291 | 155 | 9588 | 1858 |
| EC 1.2.7.8 | Indolepyruvate oxidoreductase | 15 875 | 159 | 18 473 | 2051 |
| EC 1.2.7.4* | Carbon monoxide dehydrogenase | 195 098 | 1479 | 51 839 | 51 725 |
| EC 1.17.1.9* | Formate dehydrogenase-O, iron-sulfur | 2066 | 11 | 798 | 278 |
| EC 1.18.1.3 | Naphthalene 1,2-dioxygenase system ferredoxin-NAD(+) reductase | 17 | 1 | 16 | 68 |
| ЕС | 4Fe-4S ferredoxin, iron-sulfur binding | 12 623 | 7 | 10 511 | 7212 |
| EC 1.7.7.1 | Ferredoxin—nitrite reductase | 2874 | 22 | 584 | 108 |
| EC 1.18.1.2 | Ferredoxin—NADP(+) reductase | 3118 | 1 | 1531 | 310 |
| EC 1.8.7.1 | Ferredoxin—sulfite reductase (assimilatory) | 6842 | 35 | 919 | 858 |
| Белки ассимиляционной железоредукции | |||||
| EC 1.5.1.30 | Flavin reductase | 588 | 21 | 958 | 420 |
| EC 7.1.1.8* | Ubiquinol-cytochrome c reductase | 26 319 | 858 | 20 376 | 23 046 |
| ЕС | Ferredoxin, 2Fe-2S | 26 660 | 142 | 2667 | 4219 |
| Белки диссимиляционной железоредукции | |||||
| Periplasmic decaheme cytochrome c, MtrA,D | 1567 | 5 | 75 | 40 | |
| EC 1.18.6.1 | Nitrogenase (molybdenum-iron) | 2810 | 19 | 1380 | 22 |
| EC 1.18.6.1 | Nitrogenase (vanadium-iron) | 4 | 9 | 7 | 0 |
| Белки транспорта Fe(III) и Fe(II) | |||||
| EC 4.99.1.1 | Ferrochelatase, protoheme ferro-lyase | 32 978 | 690 | 15 682 | 13 913 |
| Ferrous iron transport protein B | 56 840 | 380 | 14 643 | 2605 | |
| Белки, регулирующие транспорт Fe(III) и Fe(II) | |||||
| Ferric uptake regulation protein FUR | 19 736 | 120 | 2526 | 975 | |
Микробиологические исследования. Из образцов скважины 4-07, отобранных в районе протоки Амболихи, были получены накопительные культуры, в которых при разных температурах культивирования зафиксирован процесс восстановления трехвалентного железа. В накопительных культурах, инкубируемых при температуре 20°С (рис. 5, Б), количество восстановленного железа варьировало в пределах 0.5-2.21 мМ. Максимальное количество ионов Fe(II) (2.21 мМ) отмечено в накопительной культуре М5, полученной из образца ММО Колымской низменности, отобранного с глубины 22.2-22.3 м, а минимальное в культуре М2 с глубины 1.4 м, где содержание новообразованного двухвалентного железа не превышало 0.5 мМ. В накопительной культуре М4, полученной из образца той же скважины, но с глубины 17.5-17.6 м, концентрация ионов Fe(II) составила 0.67 мМ.
В накопительных культурах, инкубируемых в течение 40 сут при 5°С, концентрация ионов Fe(II) изменялась от 0.79 до 2.70 мМ. Микробные сообщества трех накопительных культур в процессе роста восстанавливали цитрат Fe(III) с образованием наибольшего количества Fe(II). Максимальная концентрация Fe(II) (2.70 мМ) обнаружена в накопительной культуре М5, полученной из образца ММО Колымской низменности. В накопительной культуре М4, полученной из образца арктических отложений, взятых из той же скважины с глубины 17.5-17.6 м, количество восстановленного железа составило 1.65 мМ.
В результате экспериментов показано, что сформировавшиеся микробные сообщества ЖВБ, полученные из разных природных образцов и культивируемые при 5°С, восстанавливали трехвалентное железо более активно в отличие от сообществ, полученных из этих же образцов, но выращенных при 20°С. Зарубежными коллегами в 2017 г. было обнаружено [17], что скорость восстановления железа накопительными культурами, полученными из подледниковых отложений, отобранных в различных географически отдаленных друг от друга ледниках, была выше при 4°С по сравнению с 15°С.
ЗАКЛЮЧЕНИЕ
Получены данные, указывающие на возможность протекания глеевого процесса в водораздельных и склоновых почвах, формирующихся в условиях континентального климата тундровой зоны Колымской низменности. В почвах нижней части пологих склонов и крутых склонов этот процесс является преобладающим.
Периодически вытаивающий из мерзлоты в наиболее теплые годы глеевый горизонт (Gт) характеризуется преобладанием двухвалентной формы подвижного железа в составе несиликатного, что способствует обезжелезнению этого горизонта. По всей видимости, он представляет собой реликт более теплого и влажного этапа почвообразования. Несмотря на некоторые изменения в верхней части глеевого горизонта, полной окислительной деградации при его протаивании не происходит, в том числе из-за буферной роли содержащихся в Gт торфяных прослоев и линз.
На основании анализа аннотаций метагеномов выявлены гены микроорганизмов, ответственных за железоредукцию. Показано, что в отложениях, сформировавшихся в более гидроморфных условиях и глееватом горизонте современной водораздельной почвы, доминирует филум Proteobacteria, к которым относятся железовосстанавливающие микроорганизмы. Три выбранных нами рода: Shewanella, относящийся к классу Gammaproteobacteria, Geobacter и Desulfuromonas (класс Deltaproteobacteria) - были широко представлены в этих образцах.
Выявлен процесс микробного восстановления железа в образцах (IC4) старичного озера, причем большая активность наблюдалась при температуре инкубирования 5°С, чем при 20°С. Можно предположить, что основная часть культивируемых сообществ ЖВБ адаптирована к низким температурам многолетнемерзлых отложений Арктики. Такие психрофильные сообщества играют важную роль в цикле железа и деструкции органического вещества в холодных экосистемах.
Показано, что железоредукция и осуществляющие ее микроорганизмы являются важным фактором формирования окислительно-восстановительных условий в позднем плейстоцене и современных почвах. Можно предположить, что при потеплении климата, увеличении количества атмосферных осадков, повышении температуры мерзлоты и мощности сезонноталого (активного) слоя все биогеохимические процессы, включая железоредукцию будут играть еще большую роль.
Список литературы
Аринушкина Е.В. Руководство по химическому анализу почв. М.: Изд-во Моск. ун-та, 1970. 488 с.
Губин С.В., Лупачев А.В. Надмерзлотные горизонты аккумуляции грубого органического вещества в криоземах тундр Северной Якутии // Почвообразование. 2018. № 7. С. 815-825.
Зонн С.В. Железо в почвах (генетические и географические аспекты). М.: Наука, 1982. 209 с.
Лупачев А.В., Губин С.В. Органогенные надмерзлотно-аккумулятивные горизонты криоземов тундр Севера Якутии // Почвоведение. 2012. № 1. С. 57-68.
Муликовская Е.П., Резников А.А., Соколов И.Ю. Методы анализа природных вод. М.: Недра, 1970. 488 с.
Ривкина Е.М., Краев Г.Н., Кривушин К.В., Лауринавичюс К.С., Федоров-Давыдов Д.Г., Холодов А.Л., Щербакова В.А. Гиличинский Д.А. Метан в вечномерзлых отложениях северо-восточного сектора Арктики // Криосфера Земли. 2006. Т. 10. № 3. С. 23-41.
Федоров-Давыдов Д.Г., Губин С.В., Макеев О.В. Содержание подвижного железа и возможность оглеения в почвах Колымской низменности // Почвоведение. 2004. № 2. С. 159-170.
Федоров-Давыдов Д.Г., Мергелов Н.С., Морозов М.М. Почвенный покров приморских едом Колымской низменности // Мат-лы Междунар. конф. “Криогенные ресурсы полярных регионов” (г. Салехард, июнь 2007 г.). Т. 2. С. 113-116.
Федоров-Давыдов Д.Г., Давыдов С.П., Давыдова А.И., Остроумов В.Е., Холодов А.Л., Сороковиков В.А., Шмелев Д.Г. Температурный режим почв северной Якутии // Криосфера Земли. 2018. Т. 22. № 4. С. 15-24.
Abramov A., Davydov S., Ivashchenko A., Karelin D., Kholodov A., Kraev G., Lupachev A. et al. Two decades of active layer thickness monitoring in northeastern Asia // Polar Geography. 2019. P. 1-17. https://doi.org/10.1080/1088937X.2019.1648581
Bowman J.P., McCammon,S.A., Nichols D.S., Skerratt J.H., Rea S.M., Nichols P.D., McMeekin T.A. Shewanella gelidimarina sp. nov., Shewanella frigidimarina sp. nov., novel Antarctic species with the ability to produce eicosapentaenoic acid (20: 5ω3) and grow anaerobically by dissimilatory Fe (III) reduction // Int. J. System. Evolut. Microbiol. 2005. V. 47. V. 4. P. 1040-1047.
Canfield D., Kristensen E., Thamdrup B. Aquatic Geomicrobiology1st Edition. 2005. V. 48. 656 p. ISBN: 9780080575407
Cowart R.E. Reduction of iron by extracellular iron reductases: implications for microbial iron acquisition // Archives Biochem. Biophys. 2002. V. 2. P. 273-281.
Fyodorov-Davydov D.G., Kholodov A.L., Ostroumov V.E., Kraev G.N., Sorokovikov V.A., Davydov S.P., Merekalova A.A. Seasonal thaw of soils in the North Yakutian ecosystems // Proceedings of the Ninth International conference on permafrost (June 29-July 3, 2008, University of Alaska, Fairbanks). Fairbanks, 2008. V. 1. P. 481-486.
Hedrich S., Schlömann M., Johnson D.B. The iron-oxidizing proteobacteria // Microbiology. 2011. V. 157(6). P. 1551-1564.
Meyer F., Paarmann D., D’Souza M., Olson R., Glass E.M., Kubal M., Paczian T., Rodriguez A., Stevens R., Wilke A., Wilkening J. The metagenomics RAST server-a public resource for the automatic phylogenetic and functional analysis of metagenomes // BMC Bioinformatics. V. 9. P. 386. https://bmcbioinformatics.biomedcentral.com/ articles/10.1186/1471-2105-9-386
Nixon S.L., Telling J.P., Wadham J.L., Cockel C.S. Viable cold-tolerant iron-reducing microorganisms in geographically diverse subglacial environments // Biogeosciences. 2017. V. 14(6). P. 1445-1455. https://doi.org/10.5194/bg-14-1445-2017
Pecheritsyn S.A., Rivkina E.M., Akimov V.N., Shcherbakova V.A. Desulfovibrio arcticus sp. nov., a psychrotolerant sulphate-reducing bacterium from a cryopeg // Int. J. System. Evolu. Microbiol. 2012. V. 62(1). P. 33-37.
Rivkina E., Gilichinsky D., Wagener S., Tiedje J., McGrath J. Biogeochemical activity of anaerobic microorganisms from buried permafrost sediments // Geomicrobiology J. 1998. V. 15(3). P. 187-193. https://doi.org/10.1080/01490459809378075
Rivkina E., Petrovskaya L., Vishnivetskaya T., Krivushin K., Shmakova L., Tutukina M., Meyers A., Kondrashov F. Metagenomic analyses of the late Pleistocene permafrost-additional tools for reconstruction of environmental conditions // Biogeosciences. 2016. V. 13(7). P. 2207-2219.
Ryzhmanova Y., Abashina T., Petrova D., Shcherbakova V. Desulfovibrio gilichinskyi sp. nov., a cold-adapted sulfate-reducing bacterium from a Yamal Peninsula cryopeg // Int. J. Syst. Evol. Microbiol. 2019. V. 69(4). P. 1081-1086. https://doi.org/10.1099/ijsem.0.003272
Shi T., Reeves R.H., Gilichinsky D.A., Friedmann E.I. Characterization of viable bacteria from Siberian permafrost by 16S rDNA sequencing // Microbial Ecology. 1997. V. 33(3). P. 169-179.
Slobodkin A.I., Wiegel J. Fe(III) as an electron acceptor for H2 oxidation in thermophilic anaerobic enrichment cultures from geothermal areas. Extremophiles. 1997. V. 1(2). P. 106-109.
Vandieken V., Mussmann M., Niemann H., Jørgensen B.B. Desulfuromonas svalbardensis sp. nov., Desulfuromusa ferrireducens sp. nov., psychrophilic, Fe(III)-reducing bacteria isolated from Arctic sediments, Svalbard // Int. J. System. Evolut. Microbiol. 2006. V. 56(5). P. 1133-1139. https://doi.org/10.1099/ijs.0.63639-0
Ventura B.A., González F., Ballester A., Blázquez M.L., Muñoz J.A. Bioreduction of iron compounds by Aeromonas hydrophila // Int. Biodeter. Biodegrad. 2015. V. 103. P. 69-76. https://doi.org/10.1016/j.ibiod.2015.03.034
Viollier E., Inglett P.W., Hunter K., Roychoudhury A.N., Van Cappellen P. The ferrozine method revisited: Fe(II)/Fe(III) determination in natural waters // Appl. Geochem. 2000 V. 15(6). P. 785-790.
Weber K.A., Achenbach L.A., Coates J.D. Microorganisms pumping iron: anaerobic microbial iron oxidation and reduction // Nature Rev. Microbiol. 2006. V. 4(10). P. 752-764. https://doi.org/10.1038/nrmicro1490
Wolin E.A., Wolin M., Wolfe R.S. Formation of methane by bacterial extracts // J. Biological Chemistry. 1963. V. 238(8). P. 2882-2886.
Zhang C., Stapleton R.D., Zhou J., Palumbo A.V., Phelps T.J., Iron reduction by psychrotrophic enrichment cultures // FEMS Microbiol. Ecol. 1999. V. 30(4). P. 367-371. https://doi.org/10.1111/j.1574-6941.1999.tb00664.x
Дополнительные материалы отсутствуют.