

Почвоведение, 2021, № 10, стр. 1262-1273
Эмиссия СО2 из почвы при введении краткосрочной сидерации в паровое поле в условиях лесостепной зоны Прибайкалья
Л. Г. Соколова a, С. Ю. Зорина a, *, Е. Н. Белоусова a, А. В. Поморцев a, Н. В. Дорофеев a
a Сибирский институт физиологии и биохимии растений СО РАН
664033 Иркутск, ул. Лермонтова, 132, Россия
* E-mail: zorina@sifibr.irk.ru
Поступила в редакцию 30.12.2020
После доработки 05.03.2021
Принята к публикации 27.03.2021
Аннотация
Изучены особенности эмиссии СО2 из почвы при краткосрочной летней сидерации в паровом поле (35–40 дней от посева до запашки редьки масличной Raphanus sativus var. oleifera Metzg), используемой в качестве нестандартного приема земледелия в лесостепной зоне Прибайкалья. В трехлетних (2015–2017 гг.) полевых опытах на серой лесной среднесуглинистой почве выявлены изменения в сезонной динамике скорости эмиссии СО2 из почвы при использовании данного приема по сравнению с традиционным черным паром. В двух независимых севооборотах показано увеличение интенсивности выделения СО2 с поверхности почвы после запашки зеленой массы. Суммарный (май–сентябрь) поток СО2 из почвы в варианте с сидерацией во все годы оказался в 1.3–1.6 раза больше, чем при традиционной обработке. Эффект усиления процессов минерализации за счет вновь поступившего органического вещества составил 38–131% по отношению к черному пару. Потерям подвергалось 19–79% углерода, аккумулированного в чистой первичной продукции редьки масличной. Размах колебаний суммарного потока СО2 из почвы определялся комплексом факторов: гидротермическими условиями сезона; содержанием Сорг и Nобщ в почве; отношением С : N, содержанием N и долей воды в тканях сидерата, а также запасами углерода в корневых и пожнивных остатках предшествующей культуры. Плотность связи между величиной потока СО2 и исследуемыми факторами уменьшалась в ряду: качественные характеристики зеленой массы редьки масличной > исходное плодородие почвы > пул углерода растительных остатков предшественника. Полученные данные показали, что введение краткосрочной сидерации в паровое поле увеличивает эмиссию СО2 из серой лесной почвы за теплый сезон.
ВВЕДЕНИЕ
Интерес к приемам земледелия как инструменту, позволяющему регулировать поток СО2 из почвы, обусловлен вниманием, которое уделяется сокращению выбросов парниковых газов в сельскохозяйственном производстве [37, 49, 53]. Возможности снижения газообразных потерь почвенного углерода и его секвестрации за счет ресурсосберегающих агротехнологий широко обсуждаются [28, 33, 36, 40]. Введение сидеральных культур в севообороты рассматривается в качестве фактора, способного оказать влияние на запасы почвенного органического вещества (ПОВ) и величину газообразных потерь углерода из почв [30]. Чаще указывают на усиление эмиссии СО2 при поступлении сидеральной массы [29, 48]. Выявлено, что сроки, способ и глубина заделки биомассы сидеральной культуры обусловливают величину потока СО2 [3, 34, 42]. Показана тесная прямая связь между количеством поступивших растительных остатков и эмиссией углекислого газа из почв [27]. При этом считается, что минерализация/стабилизация органического вещества в почвах детерминируются сочетанием характера землепользования и климата [11]. На примере анализа больших массивов данных установлено, что вновь внедряемые методы ведения сельского хозяйства могут вызывать совершенно противоположный отклик в процессах трансформации углерода, включая его потери, в зависимости от условий среды, поэтому требуется оценка их эффективности на экосистемном уровне [46, 54].
На территории лесостепной зоны Прибайкалья все чаще происходит замена полей, обрабатываемых по типу черного пара, на сидеральные пары. В качестве нестандартного приема земледелия предложено использование быстрорастущих капустных культур непосредственно в паровом поле. Это позволяет сочетать положительные для регионального земледелия свойства чистых (черных) и сидеральных паров. Первые дают возможность эффективно бороться с многолетними сорняками и накапливать влагу, а также благодаря минерализации органического вещества улучшают режим питания для последующих культур севооборотов. Вторые оптимизируют физические свойства почвы, осуществляют почвопокровную функцию, обеспечивают поступление свежего органического вещества и значительного пула питательных элементов [24]. Кроме того, за счет значительной скорости роста капустные культуры успешно конкурируют с сорняками за влагу и элементы питания, что усиливает фитосанитарную роль данного поля в севообороте.
Вопрос о влиянии краткосрочной сидерации парового поля капустными культурами на эмиссию СО2 из почвы остается открытым. Надо отметить, что этот агроприем отличается от других видов сидерации во многих аспектах, которые могут иметь значение для динамики потерь углерода из почвы. Вне периода возделывания капустной культуры почва остается незанятой, выступая нетто-источником СО2. Прямые оценки потерь углерода, в том числе за счет поступления свежей фитомассы, при введении данного приема земледелия отсутствуют. Не изучено влияние капустных культур на процессы трансформации углерода, хотя зависимость минерализации углерода, в том числе почвенного органического вещества, от видовых особенностей сидератов известна [51].
Цель работы – изучение особенностей эмиссии СО2 из серой лесной почвы при краткосрочной сидерации парового поля в условиях лесостепи Прибайкалья и выявление факторов, влияющих на величину газообразных потерь углерода из почвы при использовании редьки масличной как сидерата в паровом поле.
ОБЪЕКТЫ И МЕТОДЫ
Характеристика района исследований. Агроэкологический стационар СИФИБР СО РАН расположен в северо-западной части лесостепной зоны Прибайкалья (53°33′58.75″ N и 102°35′23.90″ E). Климат региона – резко континентальный, умеренно холодный [2]. Среднемноголетнее годовое количество осадков за 1981–2010 гг., которые, по современным представлениям, следует использовать для оценки климатической нормы [35], составляет 388 мм, из которых 323 мм выпадает в период с апреля по октябрь. Несмотря на то, что распределение осадков в регионе отличается значительной меж- и внутригодовой вариабельностью, начало теплого периода засушливое [5]. Среднегодовая температура воздуха низкая (‒1.1°С за 1981–2010 гг.). Весенний переход среднесуточной температуры воздуха через +10°С происходит к концу второй декады мая, а осенний – в середине сентября. Динамика среднесуточных температур теплого периода характеризуется выраженным пиком в его середине. Самые высокие среднесуточные температуры наблюдаются в июле–августе. Максимальные значения показателя в это период достигают 21.3°С, а минимальные – 11.1°С, что попадает в пределы величин, благоприятных для минерализации органического вещества [4].
Почва экспериментального участка – серая лесная среднесуглинистая (Luvic Retic Greyzemic Phaeozem (Loamic, Aric) [38]) относится к наиболее широко представленному в пахотном фонде региона (≈70%) типу почв. Почвы опытных полей характеризуются следующими свойствами: плотность сложения – 1.0–1.2 г/см3, содержание Сорг – 1.9–2.5%, Nобщ – 0.13–0.26%, величина рНвод – 6.9–7.1, сумма обменных оснований – 25.9–26.4 смоль(экв)/кг. Основные запасы почвенного углерода сосредоточены в пахотном слое (0–20 см), что обусловлено небольшой мощностью гумусового горизонта (25–30 см) [8].
Организация экспериментов. Полевые опыты проводили в течение 2015–2017 гг. Схема экспериментов включала варианты: 1 – черный пар (контроль), 2 – пар с краткосрочным (35–40 сут) посевом редьки масличной. Оба варианта ежегодно размещались в первом поле двух независимых севооборотов: четырехпольного (черный/сидеральный пар–соя–яровая пшеница–яровой ячмень; вторая ротация) и пятипольного (черный/сидеральный пар–картофель–кукуруза–пшеница–ячмень; первая ротация). Соответственно опыты I и II. Учетная площадь в опыте I составляла 1000 м2, в опыте II – 80 м2. В качестве предшествующей культуры в опытах выступал яровой ячмень. Только в 2015 г. опыт II был заложен по черному пару, в связи с началом освоения опытного поля. Содержание Сорг в опыте I было больше, чем в опыте II. Соответственно 2.5–2.8 и 1.5–2.1%, что, видимо, объясняется природной почвенной неоднородностью и отличиями по ротациям.
В качестве сидеральной культуры возделывали редьку масличную (Raphanus sativus var. oleifera Metzg или Raphanus sativus var. oleiformis Pers.). Посев проводили в третьей декаде июня, когда температура, освещенность и влагообеспеченность оптимальны для интенсивного роста растений. Накопленную за 35–40 сут от посева сидеральной культуры биомассу заделывали в почву с помощью отвальной вспашки (глубина 22 см). Паровой участок экспериментальных полей обрабатывали по типу черного пара с осенней зяблевой вспашкой, промежуточными культивациями и второй глубокой обработкой в начале августа, что совпадало с моментом запашки сидеральной культуры. До ее посева почва варианта с сидерацией обрабатывали так же, как вариант черного пара.
Определение интенсивности выделения СО2 из почвы. Скорость эмиссии СО2 из почвы исследуемых вариантов измеряли ежегодно с конца апреля по сентябрь включительно (шаг 15 сут) адсорбционным методом [26]. На поверхность почвы устанавливали емкости диаметром 5 см, содержащие по 10 мл 1 M раствора NaOH. В качестве изолятора использовали непрозрачные белые сосуды диаметром 10 см и высотой 15 см, которые врезали в почву на глубину 2–3 см. Время экспозиции составляло 24 ч, затем остаток щелочи титровали 0.2 н. раствором H2SO4. Повторность определений для каждого варианта пятикратная. Одновременно проводили холостой опыт, в котором емкость со щелочью помещали в герметичный сосуд аналогичного объема. Скорость эмиссии СО2 из почвы определяли в точках, равномерно размещенных по площади опыта. Во время вегетации сидеральной культуры камеры с поглотителем располагали между растениями. Скорость эмиссии СО2 рассчитывали по формуле:
(1)
${\text{С}}{{{\text{О}}}_{2}}\left( {{{\text{г}} \mathord{\left/ {\vphantom {{\text{г}} {{{{\text{м}}}^{{\text{2}}}}}}} \right. \kern-0em} {{{{\text{м}}}^{{\text{2}}}}}}\,\,{\text{сут}}} \right) = \left( {А{\text{ }}--{\text{ }}В} \right) \times 44N \times {{10} \mathord{\left/ {\vphantom {{10} S}} \right. \kern-0em} S},$Суммарную эмиссию СО2 для всего сезона наблюдений и отдельных ее периодов (до посева сидеральной культуры, ее вегетации и после заделки зеленой массы в почву) определяли на основе проведенных измерений в пределах выделенных интервалов методом линейной интерполяции. Дыхание корней вычисляли по разности между эмиссией СО2 из почвы с растениями и из парующегося участка за период вегетации сидерата (разностный метод).
На всех экспериментальных площадках одновременно с определением скорости эмиссии СО2 из почвы анализировали влажность пахотного горизонта (0–20 см) и среднесуточную температуру воздуха в прикорневой зоне. Влажность почвы измеряли весовым методом, а температуру воздуха в прикорневой зоне – с использованием датчика THC-4 1.2.1 (Xuzhou Youbo Electrics Co., Ltd.).
Эффект усиления процессов минерализации за счет вновь поступившего органического вещества рассчитывали по приросту суммарной эмиссии СО2 из почвы варианта с сидерацией (СО2сид, г С/м2) относительно варианта черного пара (СО2пар, г С/м2) за период после заделки зеленой массы в почву по формуле:
(2)
$\begin{gathered} {\text{Прирост С}}{\kern 1pt} --{\kern 1pt} {\text{С}}{{{\text{О}}}_{2}}\left( \% \right) = \\ = \,\,{{\left( {{\text{С}}{{{\text{О}}}_{{2{\text{сид}}}}}--{\text{С}}{{{\text{О}}}_{{2{\text{пар}}}}}} \right)} \mathord{\left/ {\vphantom {{\left( {{\text{С}}{{{\text{О}}}_{{2{\text{сид}}}}}--{\text{С}}{{{\text{О}}}_{{2{\text{пар}}}}}} \right)} {{\text{С}}{{{\text{О}}}_{{2{\text{пар}}}}}}}} \right. \kern-0em} {{\text{С}}{{{\text{О}}}_{{2{\text{пар}}}}}}} \times 100. \\ \end{gathered} $Потери свежего органического вещества растительного происхождения (РОВ) за счет дыхания почвенной микрофлоры оценивали как долю суммарной эмиссии СО2 в варианте с сидерацией в период после заделки зеленой массы в почву, выраженную в г С/м2, от величины чистой первичной продукции (ЧПП) редьки масличной.
ЧПП сидеральной культуры и возврат углерода с остатками предшественника. Величину ЧПП посева редьки масличной (г С/м2) рассчитывали на основе данных сухого веса надземной биомассы и корней, а также содержания в них углерода. Учет надземной массы редьки масличной для каждого из опытов проводили методом укосов (0.25 м2; n = 5) непосредственно перед ее заделкой в почву. Биомассу корней определяли одновременно с помощью ручного отбора из почвенного монолита (10 × 10 × 30 см) [15]. Пробы зеленой массы и корней высушивали при температуре 60°С, взвешивали и готовили к анализу на содержание углерода. Средние фактические показатели содержания Сорг в тканях редьки масличной (52.2% для надземной части и 48.7% для корней) использовали для расчетов.
Поступление углерода в почву с растительными остатками предшествующей культуры (ячменя) оценивали на основе универсального пересчетного коэффициента (0.45) и биомассы пожнивных остатков и корней. Биомассу рассчитывали по известным эмпирическим зависимостям [14], исходя из данных фактического урожая зерна.
Свойства почвы и растений. Для анализа свойств почвы использовали общепринятые методы [1]. Плотность сложения определяли буровым методом; актуальную кислотность (pHвод) – потенциометрически при соотношении почва : раствор (1 : : 2.5); сумму обменных оснований – по методу Каппена–Гильковица. Содержание Сорг в почвенных и растительных образцах устанавливали методом Тюрина, а Nобщ – по методу Кьельдаля.
Статистическая обработка результатов. Полученные данные обрабатывали с использованием программы Sigma Plot14.0. Для оценки нормальности распределения результатов наблюдений использовали критерий Шапиро–Уилка. Поскольку распределение значений скорости эмиссии СО2 и ее суммарных показателей в части выборок отличалось от нормального, для статистической обработки данных применяли непараметрические методы анализа: тест Манна–Уитни и ранговый дисперсионный анализ. Корреляционный анализ данных выполняли с использованием критерия Спирмена. В таблицах и на рисунке приведены значения медианы, а также 25 и 75% процентилей. Статистически значимыми принимали различия при уровне значимости (p) меньше 0.05.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Гидротермические условия в годы исследования. Сумма активных температур (Такт > 10°С) в годы исследования оказалась несколько выше среднемноголетних значений (1904–1992°C, против 1732°C за 1981–2010 гг.). Количество осадков за сезон наблюдений при этом колебалось в пределах среднемноголетних величин (179–263 и 235 мм соответственно). Основные отличия исследуемых лет определялись сочетанием условий тепла и влаги в отдельные периоды наблюдений (табл. 1). Период до посева сидеральной культуры (май–вторая декада июня) более благоприятным оказался в 2017 г. за счет большей суммы активных температур при незначительном дефиците влаги. Этот период в 2015 г. отличался резкой нехваткой осадков, особенно в мае, а в 2016 г. – недобором тепла. Посев сидеральной культуры (третья декада июня) только в 2015 г. был в условиях хорошей влагообеспеченности. Сумма осадков за декаду составляла 25 мм, а в 2016 и 2017 гг. – 0 и 3 мм. В период активной вегетации редьки масличной, приходящийся на июль, теплообеспеченность и количество осадков во все годы исследований были близки. Гидротермические условия августа и сентября (период после заделки зеленой массы в почву) более тепло- и влагообеспеченными оказались в 2015 и 2016 гг., по сравнению с 2017 г., когда осадков и особенно тепла было существенно меньше. В целом годы исследования позволили продемонстрировать широкий спектр характерных для региона гидротермических условий, которые могут оказать влияние на формирование потоков углерода в агроценозах, особенно в сочетании с агротехническими мероприятиями.
Таблица 1.
Метеорологическая характеристика сезона наблюдений в годы исследований
| Месяц | 2015 г. | 2016 г. | 2017 г. | Среднее за 1981–2010 гг.* |
||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | |
| Май | 178 | 10 | 85 | 28 | 271 | 18 | 173 | 34 |
| Июнь | 505 | 53 | 516 | 36 | 540 | 30 | 441 | 54 |
| Июль | 616 | 84 | 616 | 96 | 596 | 79 | 562 | 80 |
| Август | 558 | 65 | 503 | 90 | 467 | 41 | 462 | 72 |
| Сентябрь | 136 | 43 | 185 | 22 | 65 | 12 | 111 | 17 |
| Весь сезон | 1992 | 253 | 1904 | 263 | 1939 | 179 | 1732 | 235 |
Примечание. Данные по метеостанции Залари [www.pogodaklimat.ru; индекс станции 30606]; 1 – сумма активных температур (Такт > 10°С), 2 – количество осадков, мм. * Значения характеризуют климатическую норму для района исследований [35].
Сезонная динамика интенсивности выделения СО2 из почвы при краткосрочной сидерации. Краткосрочная сидерация парового поля вносила существенные изменения в формирование сезонной динамики скорости эмиссии СО2 с поверхности почвы по сравнению с черным паром во все годы исследования (рис. 1). В черном пару отмечался подъем интенсивности выделения СО2 из почв к середине сезона и затем постепенный ее спад, что характерно для паровых участков на серых лесных почвах региона [18, 19]. В варианте с сидерацией фиксировалось повышение эмиссии СО2 в период вегетации растений в 1.1–2.5 раза относительно черного пара. Отмеченное усиление эмиссии СО2 в период вегетации сидеральной культуры происходило, по-видимому, за счет повышения вклада корневого дыхания и дыхательной активности ризосферной микрофлоры по мере активного прироста биомассы и корневой экскреции [6, 25, 45].
Рис. 1.
Динамика скорости эмиссии СО2 с поверхности почвы черного пара (1) и при краткосрочной сидерации в паровом поле (2) в опытах на серой лесной почве лесостепи Прибайкалья. Представлены значения медианы, барами показаны 25 и 75% процентили. Прямоугольным контуром выделен период вегетации сидеральной культуры.
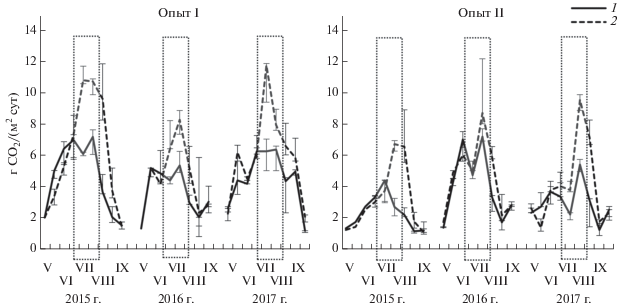
После запашки сидеральной культуры интенсивность выделения СО2 из почвы превышала эмиссионную активность черного пара в 1.1–3.0 раза. Независимо от года, эмиссия СО2 из почв постепенно уменьшалась, и к концу периода наблюдений различия между вариантами нивелировались. Надо отметить, что интенсивность выделения СО2 с поверхности почвы в варианте с сидерацией в опыте I оказались больше, чем в опыте II, где исходный уровень плодородия почвы был меньше.
Активизация процессов выделения СО2 из почвы после запашки сидеральной массы обусловлена, по-видимому, включением свежего РОВ в процессы минерализации. Известно, что растительные остатки являются более предпочтительным для почвенной микрофлоры субстратом [39]. В момент перепашки и сразу после нее не исключался вклад почвенной составляющей в поток СО2 за счет эффекта перемешивания. Но поскольку оба варианта опыта (черный и сидеральный пары) обрабатывались одновременно, полученные межвариантные различия показателей эмиссии СО2, вероятно, следует относить за счет поступившей биомассы. Нивелирование различий между вариантами уже через 3–4 недели после запашки, причем независимо от года опыта, могли быть обусловлены двумя причинами: исчерпанием легкодоступного для почвенных микроорганизмов субстрата или уменьшением их метаболической активности в связи с окончанием теплого периода.
Зависимость скорости разложения органического вещества от температуры известна. При этом характер связи чаще бывает линейным или экспоненциальным [10, 12]. В вариантах черного пара линейная связь скорости эмиссии СО2 со среднесуточной температурой воздуха оказалась прямой и сильной (r = 0.6–0.7; р < 0.05), а с влажностью почвы – обратной и слабой (r = –0.15–0.29; р < 0.05). На наличие аналогичных связей между скоростью эмиссии СО2 и гидротермическими факторами в агроэкосистемах лесостепной зоны Прибайкалья неоднократно указывалось [17–20].
Краткосрочная сидерация парового поля оказывала существенное влияние на направленность и тесноту связи между изучаемыми показателями. Так, в течение вегетации сидеральной культуры связь между скоростью эмиссии СО2 и влажностью почвы не только усиливалась, но и меняла направленность (r = 0.60 и 0.58, против –0.17 и ‒0.29 в черном пару на опытах I и II; p < 0.05). По-видимому, по сравнению с черным паром, выступающим в роли влагонакопителя [3, 24], в корнеобитаемом слое редьки масличной содержание влаги изменялось более значительно, из-за высокой транспирационной активности данной культуры [16]. В период после запашки биомассы сидерата в почву отмечалось усиление связи между скоростью эмиссии СО2 и температурой (r = 0.52 и 0.85, против r = 0.18 и 0.48 в чистом пару на опытах I и II соответственно), что подтверждает известную зависимость скорости разложения растительного опада от температуры, особенно на начальном этапе [13].
Данных об особенностях разложения и скоростях минерализации растительных остатков капустных культур, к сожалению, найти не удалось. При этом известно, что видовые особенности растений оказывают значительное влияние на минерализуемость их тканей [23]. Чаще сопоставляют скорости минерализации остатков бобовых и небобовых культур и/или побегов и корней [41, 44]. Физиологическая зрелость остатков также влияет на скорость их минерализации. По данным Семенова с соавт. [22], в начальный период трансформации зеленая масса злаковых культур (кукуруза, овес) по сравнению с их соломой отличалась преобладанием быстроразлагаемых соединений. Соответственно, минерализационные потери углерода зеленой биомассы были больше. По-видимому, максимальная скорость выделения СО2 из почвы в наших экспериментах непосредственно после заделки сидерата была обусловлена тем, что редька масличная поступала в почву преимущественно в виде зеленой массы [47]. Интенсивную начальную минерализацию РОВ связывают с быстрым оборотом наиболее легко разлагаемых компонентов растительного сырья, таких как свободные аминокислоты, аминосахара, углеводы и некоторые другие вещества клетки. Содержание их в молодых активно вегетирующих растениях обычно больше. Вступление в реакцию медленно разлагаемых составляющих (клеточных стенок и структурных компонентов) способствует замедлению скорости оборота биомассы [52]. Подобная динамика, связанная с замедлением интенсивности выделения СО2 при отдалении от момента запашки растительной массы, наблюдалась и в наших экспериментах. Полученные данные подтверждают предположение об исчерпании легкодоступного субстрата для почвенных микроорганизмов. Возможно, имело место взаимодействие между параметрами минерализуемости растительных остатков и гидротермическими условиями, но для их выявления требуются дополнительные исследования.
Суммарный поток СО2 из почвы при краткосрочной сидерации парового поля. Суммарный за период наблюдений (май–сентябрь) поток углекислоты из почвы в варианте с краткосрочной сидерацией во все годы оказался значимо (p < 0.001) больше, чем черном пару (рис. 2). В опыте I пределы его колебаний за годы эксперимента составляли соответственно 266–301 и 166–210 г С/м2. В опыте II эта закономерность сохранялась, но при меньшей величине суммарного потока СО2 (143–214, против 91–173 г С/м2). Существенных отличий по годам по этому показателю внутри варианта для каждого из опытов не найдено (p > 0.05). Исключение составляла суммарная эмиссия СО2 для опыта II в 2015 г. В обоих вариантах она оказалась наименьшей (р < 0.01). Причиной могли быть разные предшественники, поскольку в связи с началом освоения участка в 2015 г. опыт был заложен по черному пару, а в последующие годы – по яровому ячменю.
Рис. 2.
Суммарная эмиссия СО2 с поверхности серой лесной почвы в вариантах черного пара (1) и при краткосрочной сидерации в паровом поле (2). Средние значения представлены в виде медианы, контуром показан межквартильный размах между 25 и 75% процентилями, барами обозначены 10 и 90% процентили. Разными буквами отмечены выборки данных, имеющие статистически значимые различия при p ≤ 0.05, одинаковыми – в которых различия не обнаружены. Строчными буквами показаны различия внутри варианта по годам, прописными – сравнение вариантов в рамках одного года.
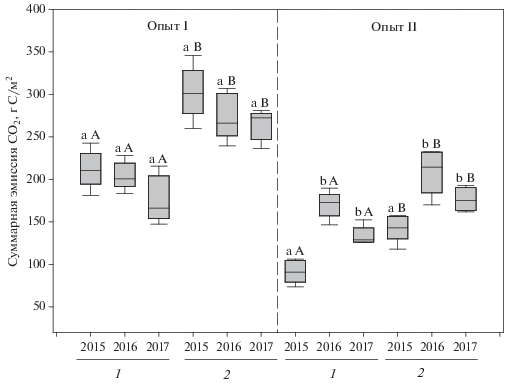
Увеличение суммарных потоков СО2 с поверхности почвы происходило за счет периода вегетации сидеральной культуры и периода после запашки зеленой массы в почву. До посева редьки масличной показатели суммарной эмиссии СО2 для исследуемых вариантов в каждом из опытов различалась незначительно (p > 0.05), что подтверждает относительную выровненность опытных участков и корректность выбранных контролей.
Суммарный поток СO2 за период вегетации редьки масличной по отношению к контрольным вариантам возрастал во все годы (108–130, против 70–81 г С/м2 в опыте I и 59–85, против 42–77 г С/м2 в опыте II). Вклад дыхания корней и ризосферной микрофлоры в его формирование, оцененный на основе различий между вариантами с растениями и без них (разностный метод), колебался в пределах 40–63% от суммарной за вегетацию эмиссии СО2. Данные сопоставимы с существующими оценками по дыханию корней (до 50% от общего потока) [9]. Необходимо отметить, что применение разностного метода давало близкие оценки только при равномерном посеве. В условиях изреженного посева (опыт II в 2016 г.) относительный прирост эмиссии СО2 в варианте с сидерацией достигал всего 10% по отношению к черному пару.
После запашки сидерата в почву суммарный поток СО2 с ее поверхности повышался в 1.4–2.3 раза по сравнению с контролем, причем различия во все годы оказались статистически значимыми (p < 0.05). Эффект усиления минерализационных потерь углерода при поступлении свежего органического вещества существенно варьировал в зависимости от опыта и года – 38–131% относительно черного пара (табл. 2). Представление о том, что добавление легкодоступного субстрата в почву способствует переключению микробного метаболизма на его использование [50], дало основание отнести эффект усиления газообразных потерь углерода после запашки редьки масличной за счет ее минерализации. В пользу этого предположения свидетельствуют исследования, согласно которым вклад почвенного органического вещества в поток СО2 с поверхности почвы при высоком содержании в ней легкодоступного субстрата является незначительным [31, 39]. На основе данных допущений провели оценку доли свежего РОВ, которое подвергалось процессам минерализации в течение текущего сезона. Согласно расчетам, минимальными (19 и 30% от ЧПП редьки масличной, соответственно в опытах I и II) относительные показатели потерь РОВ оказались в условиях недостатка тепла (2017 г.), что корреспондировало с низким эффектом усиления минерализационных потерь углерода из почвы. В более благоприятных условиях в процессы минерализации вовлекалось 42–59% от ЧПП редьки масличной. При запашке изреженного посева сидеральной культуры (опыт II в 2016 г.) эффект усиления эмиссии СО2 оказался низким, что, вероятно, связано с меньшим поступлением органического вещества в почву. При этом потери РОВ достигали 79% от ЧПП.
Таблица 2.
Эффект усиления процессов минерализации после поступления зеленой массы редьки масличной в почву и ее потери в полевых опытах
| Год | Суммарная эмиссия СО2 за период после запашки, г C/м2 | ΔСО2 (СИД – ЧП), г C/м2 | Прирост C–CO2, % | ЧПП редьки масличной, г/м2 | Потери РОВ, % C–CO2 от ЧПП | |
|---|---|---|---|---|---|---|
| сидерация (СИД) | черный пар (ЧП) | |||||
| Опыт I | ||||||
| 2015 | 102 (87; 112)* | 49 (38; 61) | 53 | 108 | 172 (144; 194) | 59 |
| 2016 | 95 (80; 104) | 46 (35; 57) | 49 | 107 | 206 (109; 272) | 46 |
| 2017 | 72 (70; 75) | 52 (50; 52) | 20 | 38 | 372 (317; 404) | 19 |
| р | 0.061 | 0.210 | 0.03 | |||
| Опыт II | ||||||
| 2015 | 67 (56; 79) | 29 (25; 31)** | 38 | 131 | 158 (124; 183) | 42 |
| 2016 | 72 (66; 91) | 52 (43; 54) | 20 | 38 | 91 (55; 95)** | 79 |
| 2017 | 63 (60; 64) | 42 (38; 45) | 21 | 50 | 206 (184; 246) | 31 |
| р | 0.160 | 0.005 | 0.01 | |||
В среднем за 3 года потери углерода свежего РОВ в течение текущего сезона составили 41 и 50% от ЧПП редьки масличной в опытах I и II соответственно. Следовательно, половина запасов углерода РОВ, аккумулированного в ходе краткосрочной сидерации, сохранялась в качестве резерва для процессов минерализации–иммобилизации, обеспечивающих эффективное плодородие почвы в последующие годы. В соответствии с данными, полученными в опытах с меченной 14С биомассой редьки масличной, ~40% углерода ее растительных остатков сохранялось через 18 мес. после внесения в песчаную почву в более теплых и влажных условиях Дании [43]. Долгосрочный прогноз, выполненный этими авторами на основе модели CN-SIM carbon, предсказал сохранение 8–10% углерода из остатков РМ в песчаной почве и через 30 лет. Для более суровых погодных условий лесостепной зоны резкоконтинентального климата секвестрация углерода при краткосрочной сидерации редькой масличной, как и время его закрепления в суглинистых почвах должны быть значительно больше.
Взаимосвязь между показателями суммарной эмиссии СО2 и свойствами почвы и растительных остатков. Корреляционный анализ, проведенный для всего ряда полученных в обоих опытах данных, позволил выявить наиболее значимые факторы, участвующие в формировании суммарного потока СО2 при краткосрочной сидерации парового поля (табл. 3). Установлены статистически значимые связи умеренной плотности между суммарным за сезон потоком СО2 и содержанием Сорг и Nобщ в почве (r = 0.55 и 0.61; р < 0.005). При этом уровень связи в варианте с сидерацией был слабее, чем в черном пару (r = 0.70 и 0.78 для показателей Сорг и Nобщ в почве соответственно), по-видимому, за счет расширения спектра воздействующих факторов. Данные подтверждают важную роль элементов плодородия почвы в формировании эмиссии СО2 с ее поверхности [11]. В период после запашки зеленой массы в варианте с сидерацией коэффициент корреляции между суммарной за данный отрезок времени эмиссией СО2 и содержанием Nобщ в почве сохранялся (r = = 0.59; р = 0.008), тогда как с содержанием Сорг снизился до 0.07. Это можно рассматривать как косвенное подтверждение снижения вклада ПОВ в формирование потока СО2 из почвы в период высокой доступности легкоминерализуемого субстрата. Сохранение тесной связи величины потока СО2 из почвы с содержанием азота доказывает роль последнего в процессах минерализации. Известно, что при разложении почвенного органического вещества микроорганизмы преимущественно иммобилизуют азот, а почвенный углерод подвергается потерям. То же, вероятно, наблюдается и на начальных этапах разложения свежего РОВ, вплоть до достижения динамического равновесия между наличием доступного углерода и потребностью микробного комплекса в азоте [21].
Таблица 3.
Коэффициенты корреляции Спирмена между величиной суммарной эмиссии СО2 из почв и свойствами почвы и растительных остатков при краткосрочной сидерации парового поля
| Параметр | Сорг в почве | Nорг в почве | С : N в почве | Возврат С с ос-татками ячменя | ЧПП РМ | С : N в тка-нях РМ | N в тканях РМ | Вода в тканях РМ, % | С– СО2 за весь сезон | СО2 за период вегетации РМ | СО2 за период после запашки |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Сорг в почве | 1 | ||||||||||
| Nорг в почве | 0.06 | 1 | |||||||||
| С : N в почве | 0.68** | –0.61** | 1 | ||||||||
| Возврат С с остатками ячменя | 0.74** | 0.29 | 0.47 | 1 | |||||||
| ЧПП РМ | 0.38 | 0.21 | 0.15 | 0.13 | 1 | ||||||
| С : N в тканях РМ | 0.38 | 0.75** | –0.17 | 0.42 | 0.39 | 1 | |||||
| N в тканях РМ | –0.39 | –0.75** | 0.19 | –0.43 | –0.32 | –0.97** | 1 | ||||
| Вода в тканях РМ, % | –0.56** | –0.46* | –0.27 | –0.72** | –0.61* | –0.65** | 0.62** | 1 | |||
| СО2 за весь сезон | 0.55** | 0.61** | 0.06 | 0.46 | 0.30 | 0.92** | –0.91** | –0.63** | 1 | ||
| СО2 за период вегетации РМ | 0.59** | 0.49 | 0.20 | 0.50* | 0.50* | Не опр. | Не опр. | Не опр. | 0.83** | 1 | |
| СО2 за период после запашки | 0.07 | 0.52** | 0.27 | 0.55* | 0.11 | 0.68** | –0.64** | –0.42* | 0.56** | 0.34 | 1 |
Анализ связи суммарной эмиссии СО2 и величиной ЧПП редьки масличной выявил статистически значимую корреляцию только в период вегетации редьки масличной (r = 0.50; р < 0.05), что подтверждает роль корневого и ризомикробного дыхания растений в формировании суммарного потока СО2 за этот период. Для периода после запашки зеленой массы коэффициент корреляции между показателями ЧПП и суммарной эмиссии СО2 был значительно меньше (r = 0.11; р > 0.05). При этом связь суммарного потока СО2 из почв с величиной С : N в тканях растений оказалась прямой и сильной (r = 0.68; р < 0.01). Близкий уровень плотности связи, но противоположный по направленности обнаружен между потоком СО2 и содержанием азота в тканях растений (r = = –0.63; р < 0.01). Найдены статистически значимые коэффициенты корреляции между эмиссией СО2 и содержанием воды в тканях зеленой массы (r = –0.42; р < 0.05). Данные дают основание полагать, что формирование потока СО2 при краткосрочной сидерации больше зависело не от количества, а от качества зеленой массы редьки масличной.
Как известно, параметры качества растительных остатков, в частности отношение С : N и содержание N в тканях, существенно влияют на активность их разложения в почве [22, 23, 29, 41]. Чаще указывают на отрицательную корреляцию между величиной С : N и количеством углерода, который теряется в результате минерализации. Однако существуют данные и о повышении минерализационных потерь С при внесении в почву РОВ с более широким отношением C : N [32]. Вероятно, неоднозначность обусловлена значительным усложнением углеродно-азотных взаимодействий в почве при внесении растительных остатков [21]. Выявленная в наших экспериментах отрицательная связь между величиной эмиссии СО2 и содержанием азота в тканях редьки масличной могла быть следствием оптимизации процессов иммобилизации. Известно, что эндогенный азот растительных остатков иммобилизуется наиболее активно [7], а редька масличная является азотонакопителем. На менее плодородных участках данная культура способна мобилизовать азот из нижележащих горизонтов почвы [16], что, по-видимому, наблюдалось в наших экспериментах. Связь между содержанием азота в тканях редьки масличной и пахотном слое почвы оказалась плотной, но отрицательной (r = –0.75; р < 0.001). Соответственно в менее обогащенную азотом почву поступала растительная масса с более узким отношением С : N. По-видимому, это сглаживало дисбаланс между поступившим углеродом и потребностью микробного комплекса в азоте, и как следствие, показатели суммарной эмиссии СО2 оставались низкими.
Статистически значимая связь средней плотности (r = 0.46–0.55; р < 0.01) между суммарной эмиссией СО2 и количеством углерода в пожнивных и корневых остатков ячменя в варианте с сидерацией позволила подтвердить, что пул углерода растительных остатков предшествующей культуры влияет на величину потерь углерода из почв с внесением свежего органического вещества. В качестве довода можно привести более слабый уровень связи между исследуемыми параметрами в варианте черного пара (r = 0.33; р < 0.05). Также в пользу этого предположения выступают выявленные различия величины эффекта усиления минерализационных потерь С при наличии предшествующей культуры и без нее. Влияние могло быть обусловлено тем, что углерод растительных остатков ячменя являлся самостоятельным дополнительным ресурсом для почвенной микрофлоры. При одновременном наличии растительных остатков ячменя и свежей биомассы редьки масличной процессы минерализации–иммобилизации углерода могли существенно усложняться за счет стимулирования или подавления различных групп почвенного микробиома, участвующего в их трансформации. К сожалению, понимание происходящих изменений невозможно без применения меченых субстратов. Решение вопроса осложняется тем, что растительные остатки ячменя и редьки масличной находились на разной стадии разложения.
Данные корреляционного анализа свидетельствуют, что величина газообразных потерь углерода при краткосрочной сидерации парового поля посевом редьки масличной в условиях лесостепной зоны Прибайкалья обусловлена комплексом факторов, складывающихся в конкретной системе почва–растения. Наиболее значимыми из них можно считать качество растительных остатков сидерата, а также исходный уровень плодородия почвы, особенно обеспеченность ее азотом, и количество углерода остатков предшествующей культуры.
ЗАКЛЮЧЕНИЕ
Введение в паровое поле краткосрочной сидерации в летний период существенно изменяет динамику эмиссии СО2 с поверхности серой лесной почвы в условиях лесостепной зоны Прибайкалья. Усиление эмиссии СО2 может наблюдаться на протяжении вегетации сидеральной культуры и в течение 3–4 недель после заделки зеленой массы в почву. На интенсивность выделения углекислого газа существенное влияние оказывают гидротермические условия конкретного сезона. Скорость эмиссии углерода из почвы в течение вегетации редьки масличной больше зависит от условий увлажнения, а в период после запашки зеленой массы в почву – от температуры. Для уточнения специфики разложения свежей биомассы капустных культур в почве, в том числе при изменении температуры, требуются дополнительные исследования в условиях лабораторного эксперимента.
Суммарный поток СО2 с поверхности почвы при введении в севооборот сидеральной капустной культуры будет увеличиваться. Эффект усиления процессов минерализации за счет поступления свежего органического вещества может варьировать в широких пределах (38–131%). По усредненным оценкам примерно половина углерода, аккумулированного в ЧПП редьки масличной, может теряться в текущем сезоне. Величина газообразных потерь углерода определяется комплексом факторов, складывающихся в конкретной системе почва–растения. Наряду с гидротермическими факторами, важно влияние качества зеленой массы, исходного плодородия почвы, количества растительных остатков предшествующей паровому полю культуры.
В целом замена чистых паров на краткосрочные сидеральные в условиях лесостепной зоны Прибайкалья может иметь как положительное, так отрицательное влияние на формирование потоков углерода в агроэкосистемах региона. Отрицательное влияние связано с усилением эмиссии СО2 из почвы, интенсивность которой будет зависеть от комплекса почвенно-климатических условий в конкретной агроэкосистеме. Положительным можно считать почвопокровную и фотосинтетическую роль сидеральной культуры. Исходя из того, что потерям подвергается лишь часть углерода, аккумулированного в фитомассе редьки масличной, судьба его остаточного количества представляет отдельный научный интерес.
Список литературы
Агрохимические методы исследования почв. М.: Наука,1975.
Атлас. Иркутская область. Экологические условия развития. М.–Иркутск: Изд-во ИГ СО РАН, Минтранс РФ, Федеральное агентство геодезии и картографии, 2004. 90 с.
Берзин А.М., Шпедт А.А. Использование зеленых удобрений в Красноярском крае // Агрохимия. 2001. № 5. С. 27–32.
Благодатская Е.В., Семенов М.В., Якушев А.В. Активность и биомасса почвенных микроорганизмов в изменяющихся условиях окружающей среды. М.: ООО Товарищество научных изданий КМК, 2016. 243 с.
Густокашина Н.Н. Многолетние изменения основных элементов климата на территории Предбайкалья. Иркутск: Изд-во Ин-та географии СО РАН, 2003. 107 с.
Евдокимов И.В. Динамика ризосферного эффекта в почве // Почвоведение. 2013. № 6. С. 715–724.
Квиткина А.К., Ларионова А.А., Быховец С.С. Влияние экзогенного и эндогенного азота на скорость минерализации растительных остатков кукурузы // Агрохимия. 2014. № 9. С. 48–57.
Колесниченко В.Т. Озимая пшеница Заларинка в Иркутской области (результаты полевых опытов). М.: Промэкобезопасность, 2003. 306 с.
Кузяков Я.В., Ларионова А.А. Вклад ризомикробного и корневого дыхания в эмиссию СО2 из почвы (обзор) // Почвоведение. 2006. № 7. С. 842–854.
Курганова И.Н., Лопес де Гереню В.О., Галлардо Ланчо Х.Ф., Ем К.Т. Оценка скорости минерализации органического вещества почв в лесных экосистемах внутриконтинентального умеренного, среднеземноморского и тропического муссонного климата // Почвоведение. 2012. № 1. С. 82–94.
Курганова И.Н., Семенов В.М., Кудеяров В.Н. Климат и землепользование как ключевые факторы стабильности органического вещества в почвах // Доклады АН. 2019. № 489(6). С. 646–650.
Ларионова А.А., Квиткина А.К., Евдокимов И.В., Быховец С.С., Стулин А.Ф. Влияние температуры на интенсивность разложения лабильного и устойчивого органического вещества агрочернозема // Почвоведение. 2013. № 7. С. 803–812. https://doi.org/10.7868/S0032180X13070046
Ларионова А.А., Мальцева А.Н., Лопес де Гереню В.О., Квиткина А.К., Быховец С.С., Золотарева Б.Н., Кудеяров В.Н. Влияние температуры и влажности на минерализацию и гумификацию лиственного опада в инкубационном эксперименте // Почвоведение. 2017. № 4. С. 438–448.
Методические рекомендации по оценке качества и классификации земель по их пригодности для использования в сельском хозяйстве. М.: Госземкадастрсъемка – ВИСХАГИ, 2003. 170 с.
Методы изучения корневых систем растений в поле и лаборатории. М.: Изд-во Моск. гос. ун-та леса, 2004. 40 С.
Пешкова А.А., Дорофеев Н.В. Биологические особенности и технологии возделывания редьки масличной. Иркутск, 2008. 146 с.
Помазкина Л.В. Мониторинг эмиссии СО2 и содержания микробной биомассы в агроэкосистемах на серой лесной почве Предбайкалья в условиях загрязнения фторидами // Почвоведение. 2015. № 8. С. 1003–1016.
Помазкина Л.В., Котова Л.Г., Лубнина Е.В. Биогеохимический мониторинг и оценка режимов функционирования агроэкосистем на техногенно загрязняемых почвах. Новосибирск: Наука, 1999. 209 с.
Помазкина Л.В., Котова Л.Г. Лубнина Е.В., Зорина С.Ю., Лаврентьева А.С. Устойчивость агроэкосистем к загрязнению фторидами. Иркутск: Изд-во ИГ СО РАН, 2004. 225 с.
Помазкина Л.В., Семенова Ю.В. Воздействие климатических изменений и загрязнения тяжелыми металлами разных типов почв на трансформацию соединений углерода в агроэкосистемах лесостепи Прибайкалья // Почвоведение. 2018. № 5. С. 617–629.
Семенов В.М. Функции углерода в минерализационно-иммобилизационном обороте азота в почвах // Агрохимия. 2020. № 6. С. 78–96. https://doi.org/10.31857/S0002188120060101
Семенов В.М., Иванникова Л.А., Кузнецова Т.В., Семенова Н.А. Роль растительной биомассы в формировании активного пула органического вещества почвы // Почвоведение. 2004. № 11. С. 1350–1359.
Семенов В.М., Паутова Н.Б., Лебедева Т.Н., Хромычкина Д.П., Семенова Н.А., Лопес де Гереню В.О. Разложение растительных остатков и формирование активного органического вещества в почве инкубационных экспериментов // Почвоведение. 2019. № 10. С. 1172–1184.
Солодун В.И., Цвынтарная Л.А. Влияние чистых и сидеральных паров на засоренность зерновых культур // Вестник ИрГСХА. 2016. Вып. 72. С. 22–27.
Титлянова A.A., Булавко Г.И., Кудряшова С.Я., Наумов A.B., Смирнов В.В., Танасиенко A.A. Запасы и потери органического углерода в почвах Сибири // Почвоведение. 1998. № 1. С. 51–59.
Шарков И.Н. Метод оценки потребности в органических удобрениях для создания бездефицитного баланса углерода в почве пара // Агрохимия. 1986. № 2. С. 109–118.
Шарков И.Н., Шепелев А.Г., Мишина П.В. Продуцирование СО2 пашней на черноземе выщелоченном в условиях лесостепи Западной Сибири // Агрохимия. 2013. № 5. С. 51–57.
Abdalla M., Hastings A., Helmy M., Prescher A., Osborne B., Lanigan G., Forristal D. et al. Assessing the combined use of reduced tillage and cover crops for mitigating greenhouse gas emissions from arable ecosystem // Geoderma. 2014. V. 223. P. 9–20. https://doi.org/10.1016/j.geoderma.2014.01.030
Almagro M., Martínez-Mena M. Litter decomposition rates of green manure as affected by soil erosion, transport and deposition processes, and the implications for the soil carbon balance of a rainfed olive grove under a dry Mediterranean climate // Agricult., Ecosyst. Environ. 2014. V. 196. P. 167–177. https://doi.org/10.1016/j.agee.2014.06.027
Bhattacharya S.S., Kim K.-H., Das S., Uchimiya M., Jeon B.H., Kwon E., Szulejko J.E. A review on the role of organic inputs in maintaining the soil carbon pool of the terrestrial ecosystem // J. Environ. Managem. 2016. V. 167. P. 214–227. https://doi.org/10.1016/j.jenvman.2015.09.042
Blagodatskaya E.V., Blagodatsky S.A., Anderson T.H., Kuzyakov Y. Priming effects in Chernozem induced by glucose and N in relation to microbial growth strategies // Appl. Soil Ecol. 2007. V. 37(1–2). P. 95–105. https://doi.org/10.1016/j.apsoil.2007.05.002
Dannehl T., Leithold G., Brock C. The effect of C:N ratios on the fate of carbon from straw and green manure in soil // Eur. J. Soil Sci. 2017. V. 68. P. 988–998. https://doi.org/10.1111/ejss.12497
Dhaliwal S.S., Naresh R.K., Gupta R.K., Panwar A.S., Mahajan N.C., Singh R., Mandal A. Effect of tillage and straw return on carbon footprints, soil organic carbon fractions and soil microbial community in different textured soils under rice-wheat rotation: a review // Rev. Environ. Sci. Bio-Technol. 2020. V. 19(1). P. 103–115. https://doi.org/10.1007/s11157-019-09520-1
Gómez J.A., Guzmán M.G., Giráldez J.V., Fereres E. The influence of cover crops and tillage on water and sediment yield, and on nutrient, and organic matter losses in an olive orchard on a sandy loam soil // Soil Till. Res. 2009. V. 106. P. 137–144. https://doi.org/10.1016/j.still.2009.04.008
Commission for Climatology (CCI) – Sixteenth session: Abridged final report with resolutions and recommendations (3–8 July 2014; Heidelberg, Germany). WMO, 2014 (WMO-№ 1137). 79 p.
He L., Zhang A., Wang X., Li J., Hussain Q. Effects of different tillage practices on the carbon footprint of wheat and maize production in the Loess Plateau of China // J. Cleaner Production. 2019. V. 234. P. 297–305. https://doi.org/10.1016/j.jclepro.2019.06.161
IPCC, 2014: Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. IPCC, Geneva, Switzerland, 151 p.
IUSS Working Group WRB: World Reference Base for Soil Resources 2014, Update 2015. World Soil Resources Reports 106. Rome: FAO, 2015.
Kuzyakov Y. Sources of CO2 efflux from soil and review of partitioning methods // Soil Biol. Biochem. 2006. V. 38. P. 425–48. https://doi.org/10.1016/j.soilbio.2005.08.020
Langeroodi A.R.S., Osipitan O.A., Radicetti E. Benefits of sustainable management practices on mitigating greenhouse gas emissions in soybean crop (Glycine max) // Sci. Total Environ. 2019. V. 660. P. 1593–1601. https://doi.org/10.1016/j.scitotenv.2019.01.074
Li F., Sorensen P., Li X., Olesen J. Carbon and nitrogen mineralization differ between incorporated shoots and roots of legume versus non-legume based cover crops // Plant and Soil. 2020. V. 446. P. 243–257. https://doi.org/10.1007/s11104-019-04358-6
Mitchell E., Scheer C., Rowlings D., Conant R., Cotru-fo M.F., van Delden L., Grace P. The influence of above-ground residue input and incorporation on GHG fluxes and stable SOM formation in a sandy soil // Soil Biol. Biochem. 2016. V. 101. P. 104–113. https://doi.org/10.1016/j.soilbio.2016.07.008
Mutegi J.K., Petersen B.M., Munkholm L.J. Carbon turnover and sequestration potential of fodder radish cover crop // Soil Use Managem. 2013. V. 29. P. 191–198. https://doi.org/10.1111/sum.12038
Mwafulirwa L., Baggs E.M., Morley N., Paterson E. Ryegrass root and shoot residues differentially affect short-term priming of soil organic matter and net soil C-balance // Eur. J. Soil Biol. 2019. V. 93. P. 103096. https://doi.org/10.1016/j.ejsobi.2019.103096
Sey B.K., Manceur A.M., Whalen J.K., Gregorich E.G., Rochette Ph. Root-derived respiration and nitrous oxide production as affected by crop phenology and nitrogen fertilization // Plant and Soil. 2010. V. 326. P. 369–379. https://doi.org/10.1007/s11104-009-0018-x
Singh. H., Northup B.K., Baath G.S., Gowda P.P., Kakani V.G. Greenhouse gas mitigation strategies for agronomic and grazing lands of the US Southern Great Plains (October, https://doi.org/10.1007/S11027-019-09894-1, 2019) // Mitigation and Adaptation Strategies for Global Change. 2020. V. 25. P. 855–855. https://doi.org/10.1007/s11027-019-09902-4
Sokolova L.G., Zorina S.Y., Pomortsev A.V., Dorofeev N.V. Carbon balance in case of short-term green manure of a fallow in the conditions of the Baikal forest-steppe // IOP Conference Series: Earth and Environmental Science: paper of International Conference on Agribusiness, Environmental Engineering and Biotechnologies – AGRITECH-2019 (Krasnoyarsk, June 20-22, 2019). 2019. V. 315. Art. 052050. https://doi.org/10.1088/1755-1315/315/5/052050
Sosa-Rodrigues A.B., Garcia-Vivas Y.S. Emission of greenhouse gases in the soil under the green manure effect // Agronomia Mesoamericana. 2019. V. 30. P. 767–782. https://doi.org/10.15517/am.v30i3.36103
Soussana J.F., Lutfalla S., Ehrhardt F., Rosenstock T., Lamanna Ch., Havlik P., Richards M. et al. Matching policy and science: Rationale for the '4 per 1000-soils for food security and climate' initiative // Soil Till. Res. 2019. V. 188. P. 3–15. https://doi.org/10.1016/j.still.2017.12.002
Sparling G.P., Fermor T.R., Wood D.A. Measurement of the microbial biomass in composted wheat straw, and the possible contribution of the biomass to the nutrition of Agaricus bisporus // Soil Biol. Biochem. 1982. V. 14. P. 609–611.
Wanic M., Zuk-Golaszewska K., Orzech K. Catch crops and the soil environmental – a review of the literature // J. Elementology. 2018. V. 24. P. 31–45. https://doi.org/10.5601/jelem.2018.23.3.1638
Watkins N., Barraclough D. Gross rates of N mineralization associated with the decomposition of plant residues // Soil Biol. Biochem. 1996. V. 28. P. 169–175.
Wollenberg E., Richards M., Smith P., Havlik P., Obersteiner M., Tubiello F., Herold M. et al. Reducing emissions from agriculture to meet the 2 degrees C target // Global Change Biol. 2016. V. 22. P. 3859–3864. https://doi.org/10.1111/gcb.13340
Zhao X., Liu B.-Y., Liu S.-L., Qi J.-Y., Wang X., Pu Ch., Li S.-S. et al. Sustaining crop production in China’s cropland by crop residue retention: A meta-analysis // Land Degrad. Developm. 2020. V. 31. P. 694–709. https://doi.org/10.1002/ldr.3492
Дополнительные материалы отсутствуют.