

Почвоведение, 2021, № 3, стр. 287-303
Динамика пулов углерода и биологической активности агродерново-подзолов южной тайги в ходе постагрогенной эволюции
И. Н. Курганова a, *, В. М. Телеснина b, В. О. Лопес де Гереню a, В. И. Личко a, Е. И. Караванова b
a Институт физико-химических и биологических проблем почвоведения РАН
142290 Пущино, Московская обл., ул. Институтская, 2, Россия
b МГУ им. М.В. Ломоносова
119991 Москва, Ленинские горы, 1, Россия
* E-mail: ikurg@mail.ru
Поступила в редакцию 17.05.2020
После доработки 19.05.2020
Принята к публикации 26.06.2020
Аннотация
Возобновление естественной растительности на бывших пахотных почвах запускает процессы постагрогенного восстановления экологических функций, присущих почвам той или иной биоклиматической зоны. На примере хроноряда агродерново-подзолов (южная тайга, Костромская область), выведенных из сельскохозяйственного использования 15, 20 и 45 лет назад, проанализирована постагрогенная динамика комплекса почвенных характеристик для верхней части старопахотного горизонта (слои 0–5 и 5–10 см): значений рНKCl, содержания органического углерода (SOC), общего азота (TN), водорастворимых форм углерода и азота (DOC и DON), скорости базального дыхания (BR), углерода микробной биомассы (Сmic) и относительных показателей состояния микробного сообщества. Для почв хроноряда были также оценены абсолютный (HIX1) и относительный (HIX2) индексы гумификации водорастворимого органического вещества (ВОВ) в слое 0–5 см и стратификационное отношение SR(0–5 : 5–10) для некоторых почвенных характеристик. Показано, что за 45 лет постагрогенной сукцессии в верхней части старопахотного горизонта наблюдается: 1 – достоверное уменьшение величины рНKCl и индекса гумификации ВОВ; 2 – рост содержания SOC, TN, Cmic, величины ВR; 3 – заметная стратификация по содержанию SOC, TN и микробным свойствам. Самых высоких значений величина SR (0–5 : 5–10) достигала в почве старовозрастного леса для SOC и TN, а в почве 45-летней залежи значения SR (0–5 : 5–10) были максимальными для микробных свойств. Таким образом, в ходе вторичной залежной сукцессии на агродерново-подзолах южно-таежной зоны происходит постепенное восстановление их основных характеристик и экологических функций.
ВВЕДЕНИЕ
В настоящее время значительные площади пахотных земель на территории России выведены из сельскохозяйственного использования [9]. По данным официальной статистики, посевные площади на территории Российской Федерации в 2017 г. занимали 80 млн га, что составляет 68% от площади посевов в 1990 г. [15]. После прекращения сельскохозяйственного использования, бывшие пахотные земли развиваются в направлении формирования естественных типов экосистем, присущих той или иной природно-климатической зоне [3, 9, 53, 65]. Ведущую роль в постагрогенной трансформации почв играет характер смены растительности [83], который особенно рельефно проявляется в таежной зоне, когда в ходе постагрогенной сукцессии происходит замена травяной растительности на лесную [45, 46, 69, 70, 78]. Общая площадь земель, выведенных из сельскохозяйственного использования и подвергающихся естественному лесовосстановлению, только в европейской части России составляет около 36.3 млн га, из которых более 2/3 расположено в таежной зоне [9]. Так, например, в отдельных регионах южной тайги (Владимирская, Тверская, Ивановская, Костромская, Псковская, Новгородская и др. области) посевная площадь в 2017 г. по сравнению с 1990 г. сократилась в 2.1–3.6 раза [15]. В целом постагрогенные экосистемы в зоне южной тайги занимают более 20% площади [61], и сегодня на бывших пахотных угодьях образовались древостои различного возраста и состава [4, 22]. Вновь сформированные еловые, сосновые или смешанные леса характеризуются значительными запасами фитомассы и образованием мощной подстилки, что значительно трансформирует углеродный баланс территории [18, 21, 61].
Скорость восстановления лесных экосистем на месте сельскохозяйственных угодий определяется как особенностями их использования в сельскохозяйственном производстве, то есть начальным состоянием залежи [56, 70], так и их зональной приуроченностью [9, 66]. При зарастании заброшенных сельскохозяйственных угодий лесной растительностью в значительной степени изменяются основные физико-химические и химические свойства почв [41–43, 45, 46, 84], содержание и состав органического вещества (ОВ) [32, 51, 58, 59], а также их биологическая активность [27, 50, 56, 58, 75]. Наиболее заметные изменения почвенных свойств охватывают главным образом старопахотную толщу почвы [20, 33, 45, 47, 78]. Вместе с тем некоторые признаки освоения почв сохраняются под лесом продолжительное время [7, 9].
Поскольку много факторов одновременно влияет на постагрогенную динамику почвенных свойств в ходе естественного лесовосстановления, то данные, имеющиеся в литературе, часто противоречивы [23, 27, 31, 74]. Постагрогенная динамика запасов углерода зависит, в частности, и от свойств нативных (ненарушенных) почв, которые определяются биоклиматическими, литологическими и геоморфологическими условиями [48, 68]. Так, после вывода из сельскохозяйственного оборота малогумусных пахотных почв запасы органического углерода (soil organic carbon, SOС) в них увеличиваются [54, 57], в то время как в длительно удобряемых сильноокультуренных пахотных почвах запасы SOC могут убывать [9, 16, 66], а в почвах с запасами, близкими к ненарушенным почвам – оставаться на прежнем уровне [41, 69]. Мета-анализ данных по изменению запасов органического углерода (SOC), в бывших пахотных почвах умеренной зоны (273 парных сравнения), показал, что средняя скорость секвестрации углерода после прекращения их сельскохозяйственного использования составляет 0.72 т C на га в год для слоя 0–30 см, и увеличение запасов SOС не лимитировано низкой продуктивностью растительности в залежных экосистемах [48]. Было также показано, что накопление С в бывших пахотных почвах в ходе постагрогенной сукцессии происходит во всех денсиметрических фракциях. Так, при зарастании агродерново-подзолистых почв южной тайги [17, 32] установлено, что со временем увеличивается содержание не только легкой фракции углерода, но и органо-минеральной, при этом доля илистой фракции углерода от SOC все же уменьшается. При постагрогенном восстановлении серых лесных почв увеличение содержания органического углерода в почве выражено главным образом за счет свободной и окклюдированной фракций [40]. В результате изучения постагрогенной динамики органического вещества темно-серых лесных почв и черноземов [66] выявлено постагрогенное накопление органического углерода во всех фракциях на глубине 0–5 см, причем доля свободного органического вещества увеличивалась, тогда как окклюдированная фракция практически не изменялась. По-видимому, тренд динамики соотношения разных денсиметрических фракций углерода в общем пуле SOC по мере восстановления растительности определяется в том числе и биоклиматической (зональной) принадлежностью экосистемы.
При естественном лесовосстановлении по мере роста древостоя идет формирование подстилки, которая представляет существенный пул слаборазложившегося ОВ в почвах. По причине постепенного увеличения запасов поступающего древесного опада [21], а также уменьшения скорости его разложения (при восстановлении хвойных лесов), запасы подстилки в течение сукцессии увеличиваются до 8–10 т/га [17, 69], возрастают сложность их строения и степень пространственного варьирования всех морфологических и химических показателей [21, 80].
Вторым по важности аспектом, помимо получения или дальнейшего уточнения общих оценок потенциального накопления углерода при выведении почв из сельскохозяйственного использования, представляется оценка стабильности вновь образуемого органического вещества [58]. По-видимому, оно в первую очередь будет вовлекаться в процессы минерализации, о чем свидетельствует его более высокая оборачиваемость [30]. Одним из подходов, позволяющих косвенно оценить стабильность растворенных органических веществ по их флуоресценции является индекс гумификации (HIX), предложенный в 1999 г. Zsolnay с соавт. [91]. Величина HIX рассчитывается как отношение интегральной интенсивности флуоресценции в диапазоне 435–480 нм (область флуоресценции более высокомолекулярных гумусированных веществ) к аналогичной величине эмиссии в диапазоне 300–345 нм (низкомолекулярные неспецифические вещества фенольного и белкового типа, включающие ароматические структуры) при возбуждении спектра светом с длиной волны 254 нм. Позднее был предложен еще один способ выражения HIX, отражающий относительную долю флуоресценции, обусловленной гумусовыми веществами, от общей флуоресценции образца [64]. По мнению авторов, он точнее отражает степень гумификации органических веществ при анализе концентрированных растворов. Предложенные индексы обычно определяют для растворимых форм ОВ природных вод, почв или других органических субстратов [49, 63, 89 ]. Его использование для оценки степени гумифицированности (стабильности) SOC в постагрогенных почвах ранее не проводилось и в данной работе выполнено впервые.
При достаточно глубокой проработанности проблемы постагрогенных изменений запасов и фракционного состава SOС в почвах залежей, динамика микробиологических свойств почв в ходе постагрогенной эволюции в подзоне южной тайги к настоящему времени наименее изучена, несмотря на то, что микробные сообщества почв весьма чутко реагируют на любые изменения природной среды [24, 25, 28]. Так, на примере постагрогенных почв в различных природно-климатических зонах было показано, что прекращение сельскохозяйственного использования ведет не только к увеличению их микробиологической активности [38, 39, 58, 60, 65, 86], но и к изменению структуры микробного сообщества [77, 82, 87]. Именно микроорганизмы, утилизируя более или менее устойчивые формы ОВ, определяют краткосрочную динамику лабильного (активного) пула SOC, состоящего из химически и физически незащищенных соединений высокого энергетического и питательного статуса [19, 85]. Вместе с тем в литературе имеется довольно скудная информация о взаимосвязях микробной активности почв с различными пулами SOC в ходе постагрогенной сукцессии.
Задачи настоящего исследования включали: 1 – анализ постагрогенной динамики пулов углерода и азота, индекса гумификации и показателей биологической активности агродерново-подзолов на примере хроноряда, представляющего собой зарастающую пашню в подзоне южной тайги; 2 – оценку взаимосвязей между всеми изученными показателями в ходе пост-агрогенной сукцессии.
ОБЪЕКТЫ И МЕТОДЫ
Регион исследования, растительность и почвы. Исследование проводили в 2019 г. в Мантуровском районе Костромской области (табл. 1) в хроноряду залежных почв различного возраста. Территория относится к северо-восточной подобласти атлантико-континентальной лесной области [2]. Среднегодовая температура воздуха за последние 20 лет составляет 3.6°С, среднегодовое количество осадков – 644 мм [1]. Почвообразующие породы представляют собой разнообразные ледниковые и водно-ледниковые отложения. Чаще всего встречаются двучленные породы, или еще более литологически сложные и неоднородные отложения, хотя верхние 40–45 см, включая старопахотную толщу, довольно однородны и имеют супесчаный гранулометрический состав [79]. Условия почвообразования в районе исследований характерны для южной тайги, где естественные растительные сообщества представлены еловыми травяно-кустарничковыми лесами [13], а преобладающим типом почв являются дерново-подзолистые (Albic Retisol) и подзолы (Retic Albic Podzol).
Таблица 1.
Почвы, растительный покров, поступление опада и запасы лесной подстилки на разных стадиях постагрогенной эволюции агродерново-подзолов
| Стадия зарастания | Координаты | Растительность | Поступление опада, т/(га год) | Запасы лесной подстилки, т/га | Почва [5] |
|---|---|---|---|---|---|
| Пашня | 58°10′55.0″ N 44°28′27.8″ E |
Посевы овса | 1.3 ± 0.1 | – | Агродерново-подзол литобарьерный среднепахотный супесчаный на ледниковых отложениях/Retic Albic Podzol (Anoarenic, Endoloamic, Raptic, Ochric) (P (0–30)–Ehi (30-46)–Ef (46–66)–BF (66–72)–BC (72–100)) |
| Залежь 15 лет | 58°10′52.7″ N 44°28′23.1″ E |
Луг с преобладанием овсяницы красной, ежи сборной и ястребинки зонтичной | 5.0 ± 0.9 | – | Агродерново-подзол реградированный лито-барьерный среднепахотный супесчаный на ледниковых отложениях/Retic Albic Podzol (Anoarenic, Endoloamic, Raptic, Ochric) (AYpa (0–30)–Ehi (30–43)–Ef (43–67)–BF (67–105)) |
| Залежь 20 лет | 58°10′52.8″ N 44°28′21.0″ E |
Практически сформирован древостой из ивы козьей, в травостое овсяница красная, ситник нитевидный, грушанка круглолистная | 3.0 ± 0.1 | 0.85 ± 0.20 | Агродерново-подзол реградированный лито-барьерный глееватый среднепахотный супесчаный на ледниковых отложениях/Retic Stagnic Albic Podzol (Anoarenic, Endoloamic, Raptic, Ochric) (AYpa (0–30)–Ehi (30–42)–Еfg (42–55)–BFg (55–92)) |
| Лес вторичный 45 лет | 58°10′54.0″ N 44°28´21.6″ E | Осиново-березовый черничный лес | 6.0 ± 0.3 | 1.81 ± 0.78 | Дерново-подзол постагрогенный литобарьерный супесчаный на ледниковых отложениях/Retic Albic Podzol (Anoarenic, Endoloamic, Raptic, Ochric) (O (0–2)–Aypa (2–25)–Ehi (25–43)–Ef (43–64)–BF (64–110)) |
| Лес 120 лет | 58°10′56.1″ N 44°28′29.0″ E |
Лес елово-березовый черничный с куртинами мха | 4.7 ± 0.2 | 5.50 ± 0.70
(в окнах)
40.2 ± 3.0 (у стволов) |
Подзол грубогумусированный литобарьерный супесчаный на ледниковых отложениях/Retic Albic Podzol (Anoarenic, Endoloamic, Raptic) (O (0–4)–AH (4–6)–Ehi (6–22)–E (22–48)–Ef (48–60)–BF (60–97)) |
Хроноряд зарастающих пахотных почв представляет площадки многолетнего мониторинга и состоит из следующих стадий: 1) пашня, засеянная овсом или многолетними травами (0-стадия); почва слабо окультурена, в последние годы (как минимум 25–30 лет) на ней проводилась распашка, удобрения не вносились около 15–20 лет; 2) 15-летняя залежь, на которой почва не возделывается с 2005 г.; 3) 20-летняя залежь, где почва не возделывается с 2000 г.; 4) мелколиственный лес 45-летнего возраста; 5) старовозрастный березово-еловый лес, имеющий возраст около 120 лет и выступающий в качестве естественной экосистемы (контроль).
Возраст древостоя определяли методом кернения (бур Пресслера). Данные об истории и продолжительности освоения земель получены от сотрудников экологической станции ИПЭЭ РАН им. А.Н. Северцова. Начиная с 2008 г., на площадках организованы регулярные наблюдения за изменением состава растительности и свойств почвы. Ботаническое описание растительности и отбор биомассы травяного (травяно-кустарничкового) яруса проводили практически ежегодно. Опад древесного яруса собирали непрерывно в течение 2012–2015 гг. с помощью шести опадоуловителей размером 25 × 25 см на каждой площадке. Легкоразлагаемый опад травяно-кустарничкового яруса учитывали как сумму надземной биомассы и 1/3 его корневой биомассы [11]. Образцы лесных подстилок отбирали послойно в 9 повторностях на площади 50 × 50 см. Запасы опада и подстилок рассчитывали на сухую массу [21].
Отбор почвенных проб и определение химических свойств почв. На каждом объекте хроноряда на площади размером 10 × 10 м2 летом 2019 г. из верхней части старопахотного горизонта (слои 0–5 и 5–10 см) методом конверта со стороной ~2 м были отобраны по 3 смешанные почвенные пробы весом 0.2 кг. Образцы почв были высушены до воздушно сухого состояния и просеяны через сито с диаметром ячеек 2 мм. Корни и грубодисперсный органический материал удаляли вручную. Таким образом, для анализов было подготовлено 30 индивидуальных почвенных образцов.
Содержание органического углерода (soil organic carbon, SOC) и общего азота (total nitrogen, TN) определяли на автоматическом CHNS анализаторе (Leco, США) в трехкратной повторности. Определение величины рН проводили в 1 М растворе КСl (соотношение почвы и раствора 1 : : 2.5) на рН-метре Metler-Toledo (Швейцария). Содержание водорастворимых форм углерода (dissolve organic carbon, DOC) и азота (dissolve organic nitrogen, DON) измеряли в водных вытяжках (соотношение почва : вода = 1 : 5) на элементном анализаторе TOC-VCPN (Shimadzu Corporation, Япония).
Определение индексов гумификации водорастворимого ОВ. Для характеристики водорастворимого органического вещества почв были сняты спектры флуоресценции на люминесцентном спектрометре LS-50 (Perkin Elmer, США). Спектры снимали при длине волны возбуждения (λex) 254 нм, изученная область длин волн эмиссии (λem) от 200 до 627.5 нм. Скорость развертки спектра 500 нм/мин, ширина щели монохроматоров для возбуждающего света 10 нм, для излучаемого – 10 нм. При обработке спектров из полученных значений вычитали эмиссию фонового растворителя (дистиллированная вода). За величину наименьшего надежно диагностируемого аналитического сигнала, обусловленного присутствием исследуемых компонентов, принималось значение 6λσ0, где λσ0 – стандартное отклонение величины флуоресценции фонового растворителя. Максимальная величина холостого сигнала для исследуемого диапазона длин волн составляла 0.6 условных единиц интенсивности флуоресценции. Для анализируемых проб значения флуоресценции были больше, как минимум на порядок. При необходимости, во избежание эффекта “внутреннего фильтра”, растворы разбавляли до оптической плотности D ≤ 0.05 [14]. По спектрам флуоресценции рассчитывали абсолютный (HIX1) и относительный (HIX2) индексы гумификации водорастворимого ОВ [64, 91]:
(1)
${\text{HIX1}} = \Sigma {\text{I}}{{\left( {435{\kern 1pt} --{\kern 1pt} 480} \right)} \mathord{\left/ {\vphantom {{\left( {435{\kern 1pt} --{\kern 1pt} 480} \right)} {\Sigma {\text{I}}\left( {300{\kern 1pt} --{\kern 1pt} 345} \right)}}} \right. \kern-0em} {\Sigma {\text{I}}\left( {300{\kern 1pt} --{\kern 1pt} 345} \right)}},$(2)
$\begin{gathered} {\text{HIX2}} = \\ = \,\,\Sigma {\text{I}}{{\left( {435{\kern 1pt} --{\kern 1pt} 480} \right)} \mathord{\left/ {\vphantom {{\left( {435{\kern 1pt} --{\kern 1pt} 480} \right)} {\Sigma {\text{I}}}}} \right. \kern-0em} {\Sigma {\text{I}}}}((300{\kern 1pt} - {\kern 1pt} 345) + \Sigma {\text{I}}(435{\kern 1pt} --{\kern 1pt} 480)), \\ \end{gathered} $Микробиологический анализ почв. Базальное дыхание почв (basal respiration, BR) определяли в трехкратной повторности по интенсивности выделения СО2 из почвы после семи дней предварительной инкубации при увлажнении, соответствующем 70–75% их полной влагоемкости, и температуре 22–24°С по формуле [55]:
(3)
$\begin{gathered} {\text{BR}}\left( {{{{\text{мг С}}} \mathord{\left/ {\vphantom {{{\text{мг С}}} {{\text{кг почвы сут}}}}} \right. \kern-0em} {{\text{кг почвы сут}}}}} \right) = \\ = \,\,\left( {{{С}_{1}}--{{С}_{0}}} \right) \times 12V{\text{флак}}{{ \times 1000} \mathord{\left/ {\vphantom {{ \times 1000} m}} \right. \kern-0em} m} \times 22.4t \times 100, \\ \end{gathered} $Содержание углерода микробной биомассы (Сmic) определяли методом субстрат-индуцированного дыхания [27] в почвенных образцах после измерения BR и внесения 1 мл раствора глюкозы из расчета 10 мг глюкозы на 1 г почвы. Скорость субстрат-индуцированного дыхания (VSIR), отражающую отклик микробного сообщества почв на внесение глюкозы, рассчитывали по формуле (3), выражали в мкл CО2/(г почвы/ч) и затем оценивали содержание Сmic по формуле [27]:
На основе полученных микробных характеристик (BR и Сmic) и содержания SOC для почв хроноряда были оценены относительные индексы функционирования микробного сообщества почвы и доступности ОВ разложению [38, 52, 56, 75, 76]:
1. Микробный метаболический коэффициент (qCO2) или удельное дыхание микробной биомассы, представляющий собой отношение BR/Cmic (мг СО2-С/(г Сmic сут)) и являющийся показателем экофизиологического состояния микробного сообщества почвы [29];
2. Отношение Сmic/SOC (%) как показатель “качества” ОВ почвы [28];
3. Отношение BR/SOC как индикатор доступности (стабильности) органического углерода почвы [55, 58].
Статистическая обработка данных. В таблицах представлены средние значения и стандартная ошибка (SE). Однофакторный дисперсионный анализ применяли для оценки влияния возраста залежи на почвенные свойства; значимые различия показаны разными буквами. Для определения связи между почвенными характеристиками использовали коэффициенты корреляции Спирмена. Визуализация данных выполнялась в программе MS Excel 2013. Статистический анализ данных проводили с использованием STATISTICA 6 при уровне значимости α = 0.05.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Изменение величины рН. По мере восстановления растительности потенциальная кислотность почвы увеличивалась, причем значимые различия между всеми стадиями сукцессии наблюдались как на глубине 0–5 см, так и 5–10 см (рис. 1, А). Наиболее резкая смена значений рН имела место при окончательном переходе лугового фитоценоза 20-летнего возраста в лесной (с 4.3 до 3.8), что в целом совпадает с данными, полученными ранее [78, 79], а также с данными других авторов [8]. Причина этого, помимо прекращения агротехнических мероприятий, заключается в изменении состава опада, поступающего на поверхность почвы, в котором при доминировании древесной растительности увеличивается доля трудноразлагаемых компонентов при одновременном уменьшении зольности опада.
Рис. 1.
Изменение значений рНKСl (А), содержания органического углерода (Б) и общего азота (В) и отношения С/N (Г) в процессе постагрогенного развития агродерново-подзолов: 1 – слой 0–5 см, 2 – слой 5–10 см.
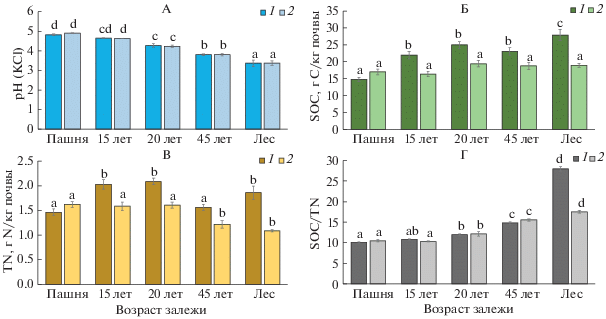
Так, доля травянистых растений в составе опада существенно уменьшается уже на стадии зарослей ивы (залежь 20 лет), а в 120-летнем лесу лиственный опад в значительной мере замещается хвойным [3]. Почва 120-летнего березово-елового леса характеризуется наибольшей степенью пространственного варьирования кислотности (от 3.1 до 3.6), обусловленной усложнением парцеллярной структуры фитоценоза, а именно, появлением выраженных компонентов тессер – приствольных повышений, подкроновых и межкроновых пространств. В ходе постагрогенного развития величина рН демонстрировала значимые отрицательные связи с содержанием SOC, DOC и величинами SOC/TN, DOC/DON (r = –0.79…–0.97, P ≤ 0.05; табл. 2).
Таблица 2.
Коэффициенты корреляции Спирмена между всеми изучаемыми почвенными характеристиками в слое 0–5 см
| Показатель | SOC | TN | SOC/TN | DOC | DON | DOC/DON | DOC/SOC | DON/TN | pH | BR | Сmic | BR/SOC | qCO2 | Cmic/SOC | HIX1 | HIX2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SOC | 1.00 | |||||||||||||||
| TN | 0.60 | 1.00 | ||||||||||||||
| SOC/TN | 0.84 | 0.09 | 1.00 | |||||||||||||
| DOC | 0.89 | 0.48 | 0.82 | 1.00 | ||||||||||||
| DON | 0.33 | 0.54 | 0.07 | 0.37 | 1.00 | |||||||||||
| DOC/DON | 0.73 | 0.15 | 0.83 | 0.79 | –0.26 | 1.00 | ||||||||||
| DOC/SOC | –0.01 | –0.01 | 0.06 | 0.43 | 0.22 | 0.21 | 1.00 | |||||||||
| DON/TN | 0.19 | 0.08 | 0.22 | 0.38 | 0.50 | –0.04 | 0.61 | 1.00 | ||||||||
| pH | –0.80 | –0.07 | –0.97 | –0.79 | –0.08 | –0.79 | –0.09 | –0.27 | 1.00 | |||||||
| BR | 0.64 | 0.39 | 0.55 | 0.51 | 0.05 | 0.47 | –0.04 | 0.23 | –0.62 | 1.00 | ||||||
| Сmic | 0.15 | –0.02 | 0.22 | –0.08 | –0.22 | 0.07 | –0.38 | 0.10 | –0.33 | 0.75 | 1.00 | |||||
| BR/SOC | 0.15 | 0.10 | 0.17 | 0.12 | –0.10 | 0.15 | 0.08 | 0.26 | –0.30 | 0.84 | 0.82 | 1.00 | ||||
| qCO2 | 0.71 | 0.55 | 0.53 | 0.87 | 0.36 | 0.64 | 0.54 | 0.26 | –0.47 | 0.40 | –0.28 | 0.11 | 1.00 | |||
| Cmic/SOC | –0.29 | –0.35 | –0.12 | –0.45 | –0.33 | –0.26 | –0.33 | 0.08 | –0.02 | 0.44 | 0.89 | 0.73 | –0.58 | 1.00 | ||
| HIX1 | –0.66 | –0.72 | –0.42 | –0.63 | –0.51 | –0.35 | –0.14 | –0.30 | 0.51 | –0.58 | –0.31 | –0.39 | –0.45 | 0.02 | 1.00 | |
| HIX2 | –0.53 | –0.70 | –0.24 | –0.44 | –0.56 | –0.10 | 0.02 | –0.30 | 0.33 | –0.28 | –0.14 | –0.07 | –0.19 | 0.10 | 0.86 | 1 |
Изменение содержания SOC, TN и отношения SOC/TN. Отчетливая динамика увеличения содержания SOC в ходе постагрогенной сукцессии выявлена только для слоя 0–5 см – от 14.7 ± 0.6 до 33.9 ± 1.6 г С/кг почвы (рис. 1, Б). Это хорошо согласуется с данными по динамике содержания органического углерода для постагрогенных почв южной тайги, полученных другими исследователями [17, 26, 44, 45]. Увеличение содержания SOC в верхних слоях старопахотной толщи на ранних этапах постагрогенного развития (травяные экосистемы), видимо, связано с постепенным развитием и преобразованием корневой биомассы [74]. На более поздних этапах рост SOС объясняется поступлением листового опада древесного яруса и образованием лесного органопрофиля, для которого типично накопление грубого гумуса в самых верхних горизонтах почвенного профиля. Несколько выпадает из общего тренда стадия мелколиственного леса 45 лет, что, возможно, связано с некоторым временным разрывом между окончанием разложения дернины, образованной в предшествующем луговом фитоценозе, и формированием достаточного запаса лесного детрита. На глубине 5–10 см отчетливого тренда увеличения содержания SOC не наблюдается. Можно видеть лишь некоторую тенденцию возрастания значений этого показателя при переходе к лесным стадиям сукцессии (17–23 до 20–28 г С/кг почвы), а также увеличение пространственного варьирования по мере лесовосстановления. Различия по этому показателю между разными объектами хроноряда не достоверны.
В пахотных почвах единственным источником органического вещества являются части растений, остающиеся после уборки урожая, а также органические удобрения, если они вносятся. В нашем случае степень окультуренности пахотной почвы крайне низка, органические удобрения не вносили по меньшей мере 15 лет. После прекращения распашки и появления травяной экосистемы происходит резкое увеличение поступления растительных остатков на поверхность почвы и корневого опада в верхнюю часть старопахотного горизонта. В результате этих процессов содержание и запасы органического углерода в почвах после прекращения их использования, как правило, увеличиваются [36]. Дальнейшая динамика гумусного состояния зависит от ряда факторов, таких как появление терминальной растительности, к которой будет стремиться в своем развитии постагрогенная экосистема, а также особенности самой почвы [9, 69].
Содержание TN в поверхностном (0–5 см) слое минимально в почве пашни (1.5 ± 0.6 г/кг), затем оно достоверно увеличивается в почвах залежей 15 и 20 лет (рис. 1, В). На стадии 45-летнего леса содержание TN достоверно не отличается от такового на пашне, а в почве старовозрастного леса оно снова увеличивается, в целом повторяя постагрогенную динамику SOC. В слое 5–10 см содержание азота в почвах пашни и залежей 15- и 20-летнего возраста достоверно не отличается и превышает таковое в почвах лесных фитоценозов (рис. 1, Б). Возможная причина этого – более резкое убывание содержания органического вещества по профилю лесных почв, обусловленное процессами оподзоливания. В ходе постагрогенного развития мы ожидаемо наблюдали значимую положительную корреляцию (r = 0.60, P ≤ 0.05) между содержанием SOС и концентрацией TN в слое 0–5 см (табл. 2).
Соотношение C/N в самой верхней части старопахотного горизонта закономерно возрастает на всем протяжении залежной сукцессии (рис. 1, Г): от 10.1 до 18.5, что говорит об уменьшении скорости разложения органического вещества. В слое 0–10 см увеличение отношения С/N при лесовосстановлении достоверно, но не столь выражено как в слое 0–5 см. Обнаружена тесная отрицательная корреляция значений рН и величины SOС/TN в слое 0–5 см (r = –0.97, P < 0.001), что может свидетельствовать о некотором замедлении процессов микробного разложения органических материалов в кислой среде.
Изменение содержания DOC, DON и отношения DOC/DON. Водорастворимый углерод является промежуточным продуктом разложения органического вещества и служит затравочным материалом для процессов минерализации и гумификации [6]. Эта фракцией почвенного углерода является наиболее чувствительной к изменению условий среды и может выступать косвенным показателем потенциальной биологической активности почв [62].
В целом динамика содержания DOC отражает динамику содержания SOC (рис. 2, А): в слое 0–5 см оно значимо возрастает от 160 ± 20 мг С/кг почвы на пашне до 264 ± 27 мг С/кг почвы на 15-летней залежи. В ходе дальнейшей сукцессии до стадии формирования мелколиственно-хвойного леса содержание DOC находится на стабильном уровне. Существенное увеличение запасов лесной подстилки в почве старовозрастного леса (табл. 1) отражается на достоверном увеличении содержания DOC, которое достигает здесь 411 ± 15 мг С/кг почвы. В почвах хроноряда выявлена значимая положительная корреляция (r = 0.82–0.89, P ≤ 0.05) между содержанием DOC, SOC и отношением SOC/TN (табл. 2). Содержание DON в слое 0–5 см сильно варьирует, поэтому его изменение в ходе постагрогенного восстановления почв не являются достоверными (рис. 2, А). Заметное увеличение содержание DON наблюдается на стадии перехода пашни в луговое сообщество (c 15 ± 3 до 18 ± 1.5 мг N/кг почвы). В почве 45-летнего леса снова происходит уменьшение содержания DON до 15 мг N/кг почвы, что, возможно, связано с возрастанием скорости разложения ОВ почвы и подтверждается увеличением именно на этой стадии интенсивности базального дыхания (рис. 3, А). Так, рост биологической активности может сопровождаться повышенным потреблением доступного азота. Отношение DOC/DON в ходе сукцессии растет, хотя достоверные различия по этому показателю между разными стадиями сукцессии также не выявлены (рис. 2, А). Как и в случае с величиной SOС/ТN, в слое 0–5 см обнаружена тесная отрицательная корреляция значений рН и значений DOC/DON (r = –0.79, P < 0.05).
Рис. 2.
Изменение содержания водорастворимого углерода (DOC) и азота (DON), отношения DOС/DON (A) и индексов гумификации HIX1 и HIX2 (Б) в слое 0–5 см в процессе постагрогенного развития агродерново-подзолов.
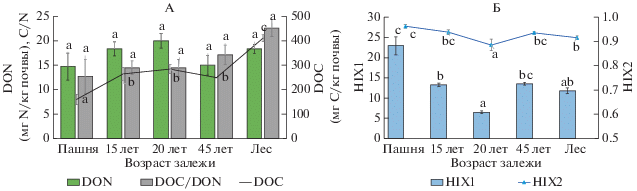
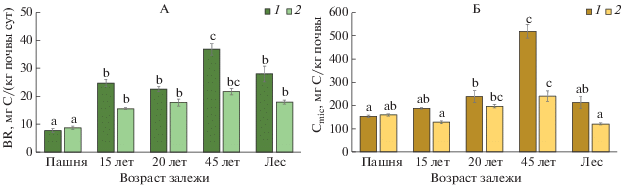
Рис. 3.
Изменение скорости базального дыхания (А) и содержания микробного углерода (Б) в процессе постагрогенного развития агродерново-подзолов: 1 – слой 0–5 см, 2 – слой 5–10 см.
Доля DOC в составе SOC в слое 0–5 см составляет не более 1.22% в течение всей сукцессии, и какой-либо значимой динамики этого показателя не выявлено. В то же время доля DON в составе TN в слое 0–5 см увеличивается от 0.69% в почве пашни до ~1% в почвах 45-летнего и старовозрастного леса.
Изменение индекса гумификации. Чем больше доля в составе водорастворимого ОВ (ВОВ) высокомолекулярных веществ, тем больше величина HIX. Для ВОВ почв, как смеси, включающей разные по структуре соединения, величины HIX можно трактовать как относительную долю в составе общего пула гумусовых веществ, которые отличаются высокими молекулярными массами, конденсированным строением и содержат ароматические циклы, обеспечивающие флуоресценцию. Кроме того, HIX используют как показатель, который коррелирует с соотношением С/Н в структурах органических веществ: чем он больше, тем больше их конденсированность и содержание углерода [90]. Поскольку гумификация сопровождается повышением атомного отношения С/H в составе молекул органических соединений, индекс HIX позволяет оценивать интенсивность этого процесса в сукцессионном ряду почв. Вместе с тем при использовании индекса HIX надо иметь в виду, что есть факторы, способные дополнительно влиять на интенсивность флуоресценции гумусовых веществ в диапазоне 435–480 нм и, соответственно, на величину HIX. Это – конформационные и конфигурационные особенности молекул органического вещества, расположение и тип заместителей в ароматических кольцах, а также возможная вариабельность границ диапазонов длин волн, в которых флуоресценция выражена [72, 73].
В исследованном ряду почв средние величины HIX1 и HIX2 тесно коррелируют между собой (r = = 0.86, P = 0.0001, табл. 2), поэтому основные тренды их изменения в ходе постагрогенной сукцессии рассмотрим на примере величины HIX1 (рис. 2, Б). Значения этого индекса варьируют довольно широко – от 6.5 в 20-летней залежи до 25.2 – на пашне. Считается, что ВОВ поверхностных вод является сильно гумифицированным при значениях HIX1 > 16, в то время как при HIX1 в диапазоне от 6 до 10 в их составе будут преобладать гумусовые соединения и некоторая доля свежеобразованного автохтонного вещества (например, прижизненных экскретов и продуктов разложения водорослей) [37]. Для почвенного ВОВ аналогами этих веществ могут быть неспецифические соединения фенольного или белкового типа, увеличение их доли в составе ВОВ будет сопровождаться уменьшением величины HIX1.
В рассматриваемом сукцессионном ряду наиболее низкие значения HIX1 = 6.5 ± 0.4 характерны для ВОВ почв 20-летней залежи (рис. 2, Б). На поверхности этих почв уже сформирован древостой, представленный ивой, и впервые фиксируется в значительном количестве древесный опад (262 г/м2) [80]. Биохимический состав ивы отличается высоким содержанием фенольных соединений (фенолгликозидов, флавоноидов, дубильных веществ) [10], некоторое количество которых, как и продукты их частичного разложения, переходят из опада в состав образующегося ВОВ. Живые древесные корни также являются типичным источником фенольных соединений, доля которых может составлять до 78% от общего содержания водорастворимого углерода [81].
Наиболее высокое среднее значение HIX1 = = 22.9 ± 2.3 отмечено для почв пашни (рис. 2, Б). Почвы пашни обеднены лабильными формами углерода [58] и свободной фракцией ОВ [32, 65, 66]. Источником углерода в них является фактически только гумус почвы. Поэтому относительная доля гумусовых веществ в составе ВОВ пахотного участка наибольшая среди почв хроноряда, что и подтверждает величина HIX1. Можно предположить, что в структуре ВОВ пахотной почвы будет и более высокое отношение С/Н. Зарастание пашни травянистой растительностью сопровождается заметным уменьшением HIX1 на 15-летней залежи (почти на 10 единиц), что связано, с увеличением корневой биомассы и продукции опада, являющихся источниками неспецифических и слабогумифицированных веществ. В отличие от почв пашни, здесь, согласно нашим более ранним исследованиям, образуется как наземный опад в количестве 338 г/м2, так и заметно (в 3 раза) увеличивается подземный [21, 80]. В почве этого участка часть ВОВ, флуоресцирующих в коротковолновой области спектра, может иметь белковую (аминокислотную) природу, что подтверждает значимая корреляция между величиной HIX2 и количества DON (r = –0.56, P = 0.05; табл. 3). Для ВОВ почв 45-летней залежи характерны чуть более высокие уровни значений HIX1, по сравнению с 15-летней (рис. 2, Б). Это незначительное увеличение величины HIX1 на фоне роста древесного опада можно объяснить увеличением микробиологической активности, приводящей к быстрой биодеструкции наиболее лабильных веществ, в том числе флуоресцирующих низкомолекулярных N-содержащих компонентов, чему соответствует достоверное уменьшение DON (рис. 2, А). На этом фоне в составе ВОВ возрастает доля более стабильных гумусовых веществ, и соответственно, увеличивается величина HIX1.
Таблица 3.
Относительные индексы функционирования микробного сообщества почвы (среднее ± SE) в слоях 0–5 и 5–10 см почв хроноряда на агродерново-подзолах. Буквенные обозначения соответствуют значимости различий между объектами хроноряда BR/SOC (мг С/г Сорг/сут); qCO2 (мг СО2-С/г Сmic/сут); Cmic/SOC (%)
| Объект | 0–5 см | 5–10 см | ||||
|---|---|---|---|---|---|---|
| BR/SOC | qCO2 | Cmic/SOC | BR/SOC | qCO2 | Cmic/SOC | |
| Пашня | 0.55 ± 0.07a | 0.51 ± 0.04a | 1.08 ± 0.12a | 0.51 ± 0.07a | 0.19 ± 0.02a | 0.95 ± 0.04b |
| 15 лет | 1.18 ± 0.08b | 0.95 ± 0.05a | 0.90 ± 0.03a | 0.95 ± 0.05b | 0.45 ± 0.06b | 0.80 ± 0.12b |
| 20 лет | 0.90 ± 0.09ab | 0.89 ± 0.03a | 1.20 ± 0.02a | 0.89 ± 0.09b | 0.37 ± 0.04ab | 0.98 ± 0.15b |
| 45 лет | 1.56 ± 0.05c | 1.15 ± 0.01a | 2.19 ± 0.05b | 1.15 ± 0.08b | 0.34 ± 0.04ab | 1.28 ± 0.15bc |
| Лес | 0.81 ± 0.11ab | 0.94 ± 0.03ab | 0.58 ± 0.06a | 0.94 ± 0.0ab | 0.52 ± 0.02b | 0.60 ± 0.09ab |
Что касается старовозрастного леса, то в этих почвах, судя по индексу HIX1, степень гумификации ВОВ еще немного снижается (до 11.8 ± 0.7), что говорит, скорее всего, об увеличении как абсолютной продукции растворимых фенольных соединений, так и их доли в составе ВОВ. В этот период в древостое уже представлена хвойная растительность, присутствует значительное количество слаборазлагаемого опада, и на поверхности почвы сформирована стратифицированная подстилка (табл. 1). Величина рН и скорость базального дыхания в почве под лесом заметно снижаются (рис. 1, А и 3, А). Эти условия благоприятны для образования и сохранения в составе ВОВ веществ фенольного типа. Растворимые фенольные соединения (РФС), помимо прочего, выступают как регуляторы количества азота в экосистеме, препятствуя его быстрой утилизации. Добавление к почве РФС, экстрагированных из подстилки, достоверно ингибирует минерализацию органического вещества и расход азота [35], что подтверждается наблюдаемым в нашем случае увеличением отношения C/N, в составе ОВ и ВОВ (рис. 1, Г, 2, А). Индекс HIX1 демонстрировал значимую отрицательную корреляцию с содержанием SOC, ТN и DOC и BR, а HIX2 – с содержанием TN и DON (табл. 2). Обратная корреляция с BR отражает увеличение на более поздних стадиях сукцессии количества растворимых лабильных неспецифических веществ, являющихся субстратом питания для микроорганизмов.
Изменение микробных характеристик. Скорость BR и величина Cmic являются наиболее чувствительными параметрами, характеризующими постагрогенную динамику почвенной микробиоты [26, 38, 58]. Наиболее низкие значения интенсивности BR в слое 0–5 см соответствуют почве пашни (7.7 ± 0.7 мг С/(кг сут)), поскольку именно эта экосистема характеризуется дефицитом легко минерализуемых компонентов растительного опада. На двух следующих стадиях сукцессии (залежи 15- и 20-летнего возраста) интенсивность базального дыхания значимо возрастает до 22–25 мг С/(кг сут) (рис. 3, А), достоверно не различаясь между собой. В почве молодого 45-летнего леса скорость BR снова значимо увеличивается, особенно в поверхностном слое старопахотного горизонта до 36 ± 2 мг С/(кг сут), но затем снова уменьшается в почве 120-летнего леса, где старопахотный горизонт практически не дифференцируется, а непосредственно за лесной подстилкой следует горизонт оподзоливания.
Постагрогенная динамика величины BR в слое 5–10 см повторяет таковую для слоя 0–5 см, но при более низких абсолютных значениях параметра (рис. 3, А). Скорость BR в слое 0–5 см демонстрировала значимую положительную связь с содержанием SOC, Cmic и отношением SOC/TN и отрицательную – с величиной рН (табл. 2). Также выявлена положительная корреляция (r = 0.87) интенсивности величины BR в слое 0–5 см с запасами ежегодно поступающего легкоразлагаемого опада [21], который служит главным субстратом для обеспечения жизнедеятельности почвенной микробиоты и поступление которого существенно повышает микробиологическую активность [18].
В ходе сукцессии был выявлен отчетливый тренд увеличения Cmic в самом верхнем (0–5 см) слое почвы (рис. 3, Б): от 153 ± 6 мг С/кг почвы на пашне до 521 ± 30 мг С/кг почвы – в почве 45-летнего леса, что естественно при восстановлении растительности, поскольку распашка почвы почти всегда приводит к уменьшению микробной биомассы [67]. В почве 120-летнего леса произошло значительное (более, чем в 2 раза) уменьшение величины Сmic, что может быть связано с существенным понижением значений рН на этом участке (до 3.4 ± 0.2 ед.), ингибирующими жизнедеятельность микробного сообщества даже при достаточном пуле доступного субстрата [26]. Постагрогенная динамика величины Сmic в слое 5–10 см повторяет таковую для слоя 0–5 см, но при более низких абсолютных значениях параметра (рис. 3, Б). Для слоя 0–5 см обнаружена тесная положительная корреляция между величиной пула Cmic, скоростью BR (r = 0.75, P < 0.01) и отношением BR/SOC (r = 0.82, P < 0.001), отражающим доступность ОВ почвы (табл. 2).
Итак, наиболее значимое изменение показателей микробной активности происходит после прекращения распашки и последующего бурного роста травянистой растительности (стадия 2), а также при окончательном формировании лесного фитоценоза на месте лугового (стадия 4), то есть при существенных моментах изменения характера биологического круговорота, его интенсивности и емкости, а также при изменении качественного состава органического вещества, поступающего в почву.
Динамика относительных показателей функционирования микробного сообщества. Метаболический коэффициент qCO2 служит косвенным индикатором не только активности микробных сообществ, но и обогащенности среды субстратом, необходимым для жизнедеятельности микробиоты, то есть свежего растительного вещества [12, 26, 38]. Самые низкие значения qCO2 (0.51 ± 0.04 мг СО2-С/(г Сmic сут) выявлены для почвы пашни как в слое 0–5 см, так и в слое 5–10 см (табл. 3).
Именно в почве пашни микробные сообщества испытывают недостаток питательных веществ, которые в неудобряемые почвы поступают только с пожнивными остатками. В почве молодой залежи в слое 0–5 см происходит довольно резкий рост qCO2 до 0.95 ± 0.05 мг СО2-С/г Сmic/сут, что связано с увеличением поступления растительного опада, причем биохимически гораздо более разнообразного, нежели в экосистеме пашни. Вместе с тем увеличивается варьирование метаболического коэффициента, что, по-видимому, говорит об увеличении разнообразия экологических ниш при постепенном восстановлении растительности и переходе луговой экосистемы в древесную. Следующий пик qCO2 наблюдается в почве 45-летнего леса (1.15 ± 0.01 мг СО2-С/(г Сmic сут), что совпадает с возрастанием поступления мелколиственного опада и формированием сплошного горизонта лесной подстилки. Постагрогенная динамика величины qCO2 в слое 5–10 см более сглажена, а сами значения метаболического коэффициента существенно ниже, чем в слое 0–5 см.
В отношении коэффициента BR/SOC, являющегося характеристикой доступности ОВ разложению [55], наблюдается похожий тренд, что говорит об уменьшении стабильности органического вещества при прекращении распашки и начале восстановления растительности, а также при формировании оранопрофиля лесного типа, о чем свидетельствуют пики BR/SOC на второй и четвертой стадиях, составляющие 1.18 ± 0.08 и 1.56 ± 0.05 мг С/(г Сорг сут) соответственно. Похожие в целом данные были получены ранее для хроноряда постагрогенных серых лесных почв [56, 58].
Доля Cmic в составе SOC изменяется от 0.6 до 2.2% в слое 0–5 см и 0.6 до 1.3% – в слое 5–10 см, принимая минимальные значения в почве старовозрастного леса, а максимальные – в почве 45-летней залежи, покрытой лесной растительностью. Постагрогенная динамика этого показателя в целом повторяет динамику двух других отношений, рассмотренных выше.
Итак, если смотреть на общие тренды изменения экофизиологических параметров микробного сообщества, то микробиологическая активность в ходе естественного восстановления растительности увеличивается, что в целом совпадает с увеличением содержания и запасов органического вещества в почве [17, 26, 65]. Так, выявлено, что метаболический коэффициент qCO2 в слое 0–5 см положительно коррелировал с SOC, TN, DOC и отношением DOC/DON (r = 0.55–0.87, P ≤ 0.05). Для того же почвенного слоя обнаружена тесная положительная взаимосвязь между величиной BR/SOC и содержанием Cmic (r = 0.82, P < 0.001; табл. 2).
Стратификация почвенных свойств в слое 0–10 см. Наиболее заметные изменения большинства почвенных свойств в различных типах почв в ходе постагрогенной сукцессии имеют место в самой верхней части старопахотного горизонта [46, 65, 66]. Кроме того, после снятия сельскохозяйственной нагрузки и зарастании бывших пахотных угодий естественной растительностью, происходит стратификация старопахотного слоя, то есть существенно меняется распределение почвенных свойств с глубиной [70]. В настоящем исследовании, используя стратификационное отношение (stratification ratio, SR), мы оценили особенности перераспределения некоторых почвенных свойств (SOC, TN, C/N, BR, Cmic, pH) в слое 0–10 см. Величина SR(0–5 : 5–10), рассчитанная как отношение между значением той или иной почвенной характеристики в слоях 0–5 и 5–10 см, показывает, как быстро идет дифференциация почвенных свойств в верхней части старопахотного горизонта при зарастании пашни лесом (рис. 4).
Рис. 4.
Изменение стратификационных отношений (SR(0–5 : 5–10)) для различных почвенных свойств в ходе постагрогенного развития агродерново-подзолов.
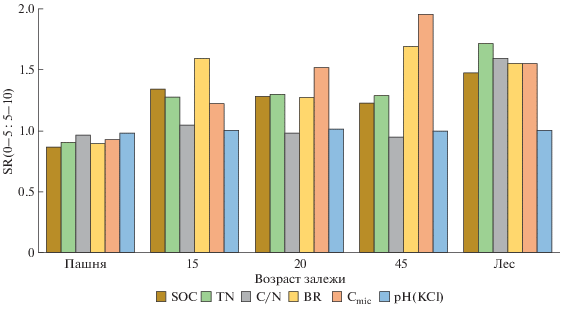
На пашне величина SR для разных свойств варьирует от 0.87 до 0.98, то есть она близка к единице, что закономерно. В ходе постагрогенной сукцессии величина SR(0–5 : 5–10) для рН и отношения С/N остается практически неизменной, и только в почве старовозрастного леса величина SR(0–5 : 5–10) для отношения С/N составляет 1.60. В почвах всех залежей степень дифференциации слоя 0–10 см по содержанию SOC и TN практически одинакова и заметно больше, чем на пашне (SR(0–5 : 5–10) = 1.23–1.30), в то время как в почве старовозрастного леса величина SR(0–5 : : 5–10) для SOC и TN достигает 1.48 и 1.72 соответственно. Наиболее отчетливо стратификация верхней части старопахотного слоя проявляется по интенсивности BR и содержанию Cmic. Величина SR(0–5 : 5–10) c возрастом залежи постепенно растет, достигая максимума в почве 45-летней залежи под молодым лиственным лесом, где SR (0–5 : 5–10) = 1.69 (BR) и SR(0–5 : 5–10) = = 1.96 (Cmic). Интересно отметить, что в почве старовозрастного леса степень дифференциации слоя 0–10 см по всем рассматриваемым свойствам, кроме рН, практически одинакова и величина SR(0–5 : 5–10) варьирует от 1.48 до 1.72. Таким образом, стратификация верхней части старопахотного горизонта по содержанию SOC и TN наиболее выражена в почве старовозрастного леса, а по микробным свойствам – в почве 45-леней залежи.
В почвах под естественной растительностью, как правило, органическое вещество более стратифицировано по глубине по сравнению с пахотными почвами, поэтому при естественном лесовосстановлении величина SR для всех почвенных характеристик, положительно связанных с содержанием SOC, увеличивается, что свидетельствует об улучшении экологического качества почв [52, 70]. Кроме того, cтратификационное отношение часто используют для оценки секвестрации углерода [71, 88]. Величина SR зависит от мощности выбранных почвенных слоев, и наиболее часто используют отношение между слоями 0–10 и 10–20 см [52]. Однако этот выбор часто бывает самым разным и зависит от целей исследования [34]. Заметный рост значений SR(0–5 : 5–10) по содержанию SOC в ходе постагрогенной сукцессии говорит о высоком потенциале залежных почв южно-таежной зоны к секвестрированию углерода. Ранее, на примере двух залежных хронорядов Костромской области с использованием SR(0–10 : 20–30) была продемонстрирована постепенная дифференциация старопахотной толщи по содержанию и запасам органического углерода, которая была максимально выражена в почвах субклимаксных смешанных лесов 90–100 лет [70].
ВЫВОДЫ
1. Восстановление естественной растительности на бывших пахотных почвах в подзоне южной тайги ведет к увеличению поступления свежего органического материала в форме надземного и подземного опада и определяет существенный рост содержания органического углерода и общего азота в верхней части старопахотного слоя.
2. Индекс гумификации водорастворимого почвенного вещества в ходе естественного лесовосстановления уменьшается, принимая самые низкие значения в 20-летней залежи, на которой сформирован древостой из ивы козьей, формирующей опад, богатый фенольными соединениями, способными ингибировать процессы активного преобразования органических субстратов.
3. Вторичная залежная сукцессия индуцирует заметный рост интенсивности BR и увеличение содержания Cmic, наиболее выраженное в слое 0–5 см. Максимальная биологическая активность почв наблюдается в почве 45-летней залежи, на которой сформирован лиственный лес, подстилка которого является источником лабильных форм углерода, представляющих легкодоступный субстрат для микробного сообщества.
4. Наиболее значимое изменение относительных показателей микробной активности происходит после прекращения распашки и последующего бурного роста травянистой растительности (луговая стадия), а также при окончательном формировании лесного фитоценоза на месте лугового, когда происходят существенные изменения качественного состава органического вещества, поступающего в почву.
5. В ходе постагрогенной сукцессии наблюдается заметная стратификация верхней части старопахотного горизонта по содержанию SOC, TN, Сmic и интенсивности BR. Самые высокие значения величина SR(0–5 : 5–10) достигала в почве старовозрастного леса для SOC и TN, а в почве 45-летней залежи значения SR(0–5 : 5–10) были максимальными для микробных свойств.
6. Корреляционный анализ показал наличие тесных связей между различными пулами углерода, азота и отношением C/N в слое 0–5 см с индексами гумификации водорастворимого ОВ, микробной активностью почв и метаболическим коэффициентом qCO2.
Список литературы
Архив климатических данных. http://climatbase.ru/ дата обращения 04.06.2020.
Болысов С.И., Фузеина Ю.Н. Физико-географические условия Костромского Заволжья. Геолого-геоморфологическое устройство // Костромское Заволжье: природа и человек. М., 2001. С. 36–60.
Владыченский А.С., Телеснина В.М., Чалая Т.А. Влияние поступления растительного опада на биологическую активность почв южной тайги // Вестник Моск. ун-та. Сер. 17. Почвоведение. 2012. № 1. С. 3–10.
Ермолова Л.С., Гульбе Я.И., Гульбе Т.А., Гульбе А.Я. Морфологические особенности и сезонное развитие ольхи серой в молодняках на залежах Ярославской области // Лесоведение. 2016. № 4. С. 279–293.
Классификация и диагностика почв России. Смоленск: Ойкумена, 2004.
Колесник А.А. Пространственное распределение общего и водорастворимого гумуса в агрочерноземах красноярской лесостепи при минимизации основной обработки. Инновационные тенденции развития Российской науки. Красноярск, 2017. С. 38–40.
Литвинович А.В., Павлова О.Ю., Чернов Д.В. Изменение гумусового состояния дерново-подзолистой почвы при прекращении антропогенного воздействия // Доклады РАСХН. 2002. № 6. С. 26–28.
Литвинович А.В., Павлова И.А. Изменение кислотно-основных свойств дерново-подзолистой суглинистой почвы в процессе постагрогенной эволюции // Научное обеспечение развития АПК в условиях реформирования. СПб., 2009. С. 160–164.
Люри Д.И., Горячкин С.В., Караваева Н.А., Денисенко Е.А., Нефедова Т.Г. Динамика сельскохозяйственных земель России в ХХ в. и постагрогенное восстановление растительности и почв. М.: ГЕОС, 2010.
Масленникова К.А., Конюхова О.М., Канарский А.В. Фенолгликозиды растений семейства Salicaceae // Вестник КТУ. 2014. Т. 17. № 14. С. 383–387.
Методы изучения биологического круговорота в различных природных зонах. М.: Мысль, 1978. 181 с.
Мостовая А.С., Курганова И.Н., Лопес де Гереню В.О., Хохлова О.С., Русаков А.В., Шаповалов А.С. Изменение микробиологической активности серых лесных почв в процессе естественного лесовосстановления // Вестник ВГУ. Сер. химия, биология, фармация. 2015. № 2. С. 64–72.
Огуреева Г.Н. Ботанико-географическое районирование СССР. М.: Изд-во Моск. ун-та, 1991. 76 с.
Паркер С. Фотолюминесценция растворов. М.: Мир, 1972. 510 с.
Регионы России. Социально-экономические показатели. М.: Росстат., 2018. 1162 с.
Романовская А.А., Коротков В.Н., Карабанов Р.Т., Смирнов Н.С. Динамика элементов баланса углерода на неиспользуемых пахотных угодьях Валдайской возвышенности // Экология. 2012. № 5. С. 347–352. https://doi.org/10.1134/S106741361204011X
Рыжова И.М., Ерохова А.А., Подвезенная М.А. Изменение запасов углерода в постагрогенных экосистемах в результате естественного восстановления лесов в Костромской области // Лесоведение. 2015. № 4. С. 307–317.
Семенов В.М., Иванникова Л.А., Кузнецова Т.В., Семенова Н.А. Роль растительной биомассы в формировании активного пула органического вещества почвы // Почвоведение. 2004. № 11. С. 1350–1359.
Семенов В.М., Когут Б.М. Почвенное органическое вещество. М.: ГЕОС, 2015. 233 с.
Скворцова Е.Б., Баранова О.Ю., Нумеров Г.Б. Изменение микростроения почв при зарастании пашни лесом // Почвоведение. 1987. № 9. С. 101–109.
Телеснина В.М., Богатырев Л.Г., Бенедиктова А.И., Земсков Ф.И., Маслов М.Н. Динамика поступления растительного опада и некоторых свойств лесных подстилок при постагрогенном лесовосстановлении в условиях южной тайги // Вестник. Моск. ун-та. Сер. 17. Почвоведение. 2019. № 4. С. 3–10.
Уткин А.И., Гульбе Я.И., Гульбе Т.А., Гульбе А.Я., Ермолова Л.С. Березняки и сероольшаники центра Русской равнины – экотон между экосистемами хвойных пород и сельскохозяйственными угодьями [О научно-организационной деятельности Сукачева В.Н. на Княжедворском стационаре (Псковская обл.) в начале 20 в.] // Лесоведение. 2005. № 4. С. 64–65.
Фомина А.С. Интенсивность протекания элементарных почвообразовательных процессов в дерново-подзолистых песчаных почвах и пути дальнейшего использования залежных земель // Известия СПбГАУ. 2009. № 13. С. 11–15. https://doi.org/10.1134/S1064229314090117
Anan’eva N.D., Blagodatskaya E.V., Demkina T.S. Estimating the resistance of soil microbial complexes to natural and anthropogenic impacts // Eurasian Soil Science. 2002. V. 35. № 5. P. 514–521.
Anan’eva N.D., Blagodatskaya E.V., Demkina T.S. The effect of drying-moistering and freezing-thawing on soil microbial communities resilience // Eurasian Soil Science. 1997. V. 30. № 9. P. 1010–1014.
Anan’eva N.D., Susyan E.A., Ryzhova I.M., Bocharnikova E.O., Stolnikova E.V. Microbial Biomass Carbon and the Microbial Carbon Dioxide Production by Soddy-Podzolic Soils in Postagrogenic Biogeocenoses and in native Spruce Forests of the Southern Taiga (Kostroma Oblast) // Eurasian Soil Science. 2009. V. 42. № 9. P. 1029–1037. https://doi.org/10.1134/S1064229309090105
Anderson J.P.E., Domsch K.H. A phisiological method for the quantitative measurement of microbial biomass in soils // Soil Biol. Biochem. 1978. V. 10. № 3. P. 215–221.
Anderson T.-H. Physiological analysis of microbial communities in soil: Applications and limitations // Beyond the Biomass. London: Wiley-Sayce Publication, 1994. P. 67–76.
Anderson T.-H., Domsch K.H. The metabolic quotient for CO2 (qCO2) as a specific activity parameter to assess the effects of environmental conditions, such as pH, on the microbial biomass of forest soils // Soils Biol. Biochem. 1993. V. 25. P. 393–395. https://doi.org/10.1016/0038-0717(93)90140-7
Blagodatskaya E., Yuyukina T., Blagodatsky S., Kuzyakov Y. Turnover of soil organic matter and of microbial biomass under C3-C4 vegetation change: consideration of 13C fractionation and preferential substrate utilization // Soil Biology and Biochemistry. 2011. V. 43. № 1. P. 159–166. https://doi.org/10.1016/j.soilbio.2010.09.028
Ermolaev A.M., Shirshova L.T. Influence of weather conditions and management of a sown meadow on the herbage productivity and properties of gray forest soils // Eurasian Soil Science. 2000. V. 33. № 12. P. 1321–1328.
Erokhova A.A., Makarov M.I., Morgun E.G., Ryzhova I.M. Effect of the Natural Reforestation of an Arable Land on the Organic Matter Composition in Soddy-Podzolic Soils // Eurasian Soil Science. 2014. V. 47. № 11. P. 1100–1106. https://doi.org/10.1134/S1064229314110040
Falkengren-Grerup U., ten Brink D.-J., Brunet J. Land use effects on soil N, P, C and pH persist over 40–80 years of forest growth on agricultural soils // Forest Ecol. Manag. 2006. V. 225. № 1–3. P. 74–81. https://doi.org/10.1016/j.foreco.2005.12.027
Ferreira A.O., Carneiro Amado T.J. Stratification Ratio as Soil Carbon Sequestration Indicator in Oxisol and Alfisol Under No-Tillage // CO2 Sequestration and Valorization / Eds.: Claudia do Rosario, Vaz Morgado and Victor Paulo Pecanha Esteves. IntechOpen, 2014. Chapter 6. P. 157–169. https://doi.org/10.5772/57063
Ganjegunte G.K., Condron L.M., Clinton P.W., Davis M.R., Mahieu N. Effects of the addition of forest floor extracts on soil carbon dioxide efflux // Biol Fertil Soils. 2006. V. 43. № 2. P. 199–207. https://doi.org/10.1007/S00374-006-0093-6
Guo L.B., Gifford R.M. Soil carbon stocks and land use change: a metaanalysis // Global Change Biology. 2000. V. 8. P. 345–360. https://doi.org/10.1046/j.1354-1013.2002.00486x
Huguet A., Vacher L. Relexans S., Saubusse S., Froidefond J.M., Parlanti E. Properties of fluorescent dissolved organic matter in the Gironde Estuary // Organic Geochemistry. 2009. V. 40. P. 706–719. https://doi.org/10.1016/j.orggeochem.2009.03.002
Ivashchenko K.V., Ananyeva N.D., Vasenev V.I., Kudeyarov V.N., Valentini R. Biomass and respiration activity of soil microorganisms in anthropogenically transformed ecosystems (Moscow region) // Eurasian Soil Science. 2014. V. 47. № 9. P. 892–903. https://doi.org/10.1134/S1064229314090051
Jia G., Cao J., Wang C., Wang G. Microbial biomass and nutrients in soil at the different stages of secondary forest succession in Ziwulin, northwest China. Forest Ecology and Management. 2005. V. 217(1). P. 117–125. https://doi.org/10.1016/j.foreco.2005.05.055
Kalinina O., Cherkinsky A., Chertov O., Goryachkin S., Kurganova I., Lopes de Gerenyu V., Lyuri D., Kusyakov Ya., Giani L. Post-agricultural restoration: Implication for dynamics of soil organic matterpools // Catena. 2019. V. 181. P. 1–8. https://doi.org/10.1016/j.catena.2019.104096
Kalinina O., Chertov O., Dolgikh A.V., Goryachkin S.V., Lyuri D.I., Vormstein S., Giani L. Self-restoration of post-agrogenic Albeluvisols: Soil development, carbon stocks and dynamics of carbon pools // Geoderma. 2013. V. 207–208. P. 221–233. https://doi.org/10.1016/j.geoderma.2013.05.019
Kalinina O.Yu., Chertov O., Frolov P., Goryachkin S., Kuner P., Küper J., Kurganova I., Lopes de Gerenyu V., Lyuri D., Rusakov A., Kuzyakov Y., Giani L. Alteration process during the post-agricultural restoration of Luvisols of the temperate broad-leaved forest in Russia // Catena. 2018. V. 171. P. 602–612. https://doi.org/10.1016/j.catena.2018.08.004
Kalinina O., Barmin A.N., Chertov O., Dolgikh A.V., Goryachkin S.V., Lyuri D.I., Giani L. Self-restoration of post-agrogenic soils of Calcisol–Solonetz complex: Soil development, carbon stock dynamics of carbon pools // Geoderma. 2014. V. 237–238. P. 117–128. https://doi.org/10.1016/j.geoderma.2014.08.013
Kalinina O., Goryachkin S.V., Karavaeva N.A., Lyuri D.I., Giani L. Dynamics of carbon pools in post-agrogenic sandy soils of southern taiga of Russia. Carbon Balance and Management. 2010. V. 5. № 1. P. 1–9. https://doi.org/10.1186/1750-0680-5-1
Kalinina O., Goryachkin S.V., Karavaeva N.A., Lyuri D.I., Najdenko L., Giani L. Self-restoration of post-agrogenic sandy soils in the southern Taiga of Russia: Soil development, nutrient status, and carbon dynamics // Geoderma. 2009. V. 152. P. 35–42. https://doi.org/10.1016/j.geoderma.2009.05.014
Kalinina O., Goryachkin S.V., Lyuri D.I., Giani L. Post-agrogenic development of vegetation, soils, and carbon stocks under self-restoration in different climatic zones of European Russia // Catena. 2015. V. 129. P. 18–29. https://doi.org/10.1016/j.catena.2015.02.016
Kalinina O., Krause S.E., Goryachkin S.V., Karavaeva N.A., Lyuri D.I., Giani L. Self-restoration of post-agrogenic Chernozems of Russia: Soil development, carbon stocks, and dynamics of carbon pools // Geoderma. 2011. V. 162. P. 196–206. https://doi.org/10.1016/j.geoderma.2011.02.005
Kämpf I., Hölzel N., Störrle M., Broll G., Kiehl K. Potential of temperate agricultural soils for carbon sequestration: A meta-analysis of land-use effects // Science of the Total Environment. 2016. V. 566–567. P. 428–435. https://doi.org/10.1016/j.scitotenv.2016.05.067
Karavanova E.I., Zolovkina D.F., Stepanov A.A. Biochemical Stability of Water-Soluble Organic Matter in Litter of Iron-Illuvial Podzol before and after Interaction with Mineral Horizons // Moscow Univ. Soil Sci. Bull. 2019. V. 74. P. 154–159. https://doi.org/10.3103/S0147687419040057
Karelin D.V., Goryachkin S.V., Kudikov A.V., Lopes de Gerenu V.O., Lunin V.N., Dolgikh A.V., Lyuri D.I. Changes in carbon pool and CO2 emission in the course of postagrogenic succession on gray soils (Luvic Phaeozems) in Europian Russia // Eurasian Soil Science. 2017. V. 50. № 5. P. 559–572. https://doi.org/10.1134/S1064229317050076
Kechaikina I.O., Ryumin A.G., Chukov S.N. Postagrogenic transformation of organic matter in soddy-podzolic soils // Eurasian Soil Science. 2011. V. 44. № 10. P. 1077–1089. https://doi.org/10.1134/S1064229311100061
Kooch Y., Ehsanib S., Akbariniac M. Stratification of soil organic matter and biota dynamics in natural andanthropogenic ecosystems // Soil and Tillage Research. 2020. V. 200. P. 104621. https://doi.org/10.1016/j.still.2020.104621
Kurganova I.N., Lopes de Gerenyu V.O. Assessment and prediction of changes in the reserves of organic carbon in abandoned soils of European Russia in 1990–2020 // Eurasian Soil Science. 2008. V. 41. № 13. P. 1371–1377. https://doi.org/10.1134/S1064229308130048
Kurganova I.N., Lopes de Gerenyu V.O. The stock of organic carbon in soils of the Russian Federation: Updated estimation in connection with land use changes // Doklady Biological Sciences. 2009. V. 426. № 8. P. 219–221. https://doi.org/10.1134/S0012496609030089
Kurganova I.N., Lopes de Gerenyu V.O., Gallardo Lancho J.F., Oehm C.T. Evaluation of the rates of soil organic matter mineralization in forest ecosystems of temperate continental, mediterranean, and tropical monsoon climates // Eurasian Soil Science. 2012. V. 45. № 1. P. 68–79. https://doi.org/10.1134/S1064229312010085
Kurganova I.N., Lopes de Gerenyu V.O., Mostovaya A.S., Ovsepyan L.A., Telesnina V.M., Lichko V.I., Baeva Y.I. The Effect of Reforestation on Microbial Activity in Post-Agrogenic Soils in European Part of Russia // Contemporary Problem of Ecology. 2018. № 7. P. 7–17. https://doi.org/10.1134/S1995425518070089
Kurganova I.N., Lopes de Gerenyu V. O., Shvidenko A.Z., Sapozhnikov P.M. Changes in the organic carbon pool of abandoned soils in Russia (1990–2004) // Eurasian Soil Science. 2010. V. 43. № 3. P. 333–340. https://doi.org/10.1134/S1064229310030129
Kurganova I., Merino A., Lopes de Gerenyu V., Barros N., Kalinina O., Giani L., Kuzyakov Y. Climate dependent mechanisms of carbon sequestration and stabilization by restoration of arable soils after abandonment // Geoderma. 2019. V. 354. P. 113882. https://doi.org/10.1016/j.geoderma.2019.113882
Kurganova I., Yermolaev A., Lopes de Gerenyu V., Larionova A., Kuzyakov Y., Keller T., Lange S. Carbon balance in soils of abandoned lands in Moscow region // Eurasian Soil Science. 2007. V. 40. № 1. P. 50–58. https://doi.org/10.1134/S1064229307010085
Lopes de Gerenyu V., Kurganova I., Kuzyakov Ya. Carbon pools and sequestration in former arable Chernozems depending on restoration period // Ekologjia. 2008. V. 54. № 4. P. 38–44. https://doi.org/10.2478/v10055-008-0034-9
Lyuri D.I., Karelin D.V., Kudikov A.V., Goryachkin S.V. Changes in Soil Respiration in the Course of the Postagrogenic Succession on Sandy Soils in the Southern Taiga Zone // Eurasian Soil Science. 2013. V. 46. № 9. P. 935–947. https://doi.org/10.1134/S1064229313070041
McDowell W.H. Dissolved organic matter in soils–future directions and unanswered questions // Geoderma. 2003. V. 113(3-4). P. 179–186. https://doi.org/10.1016/S0016-7061(02)00360-9
Odintsov P.E., Karavanova E.I., Stepanov A.A. Transformation of Water-Soluble Organic Substances in Litters of Podzols in the Background and Technogenic Areas of the Kola Peninsula // Eurasian Soil Science. 2018. V. 51. P. 955–964. https://doi.org/10.1134/S1064229318080094
Ohno T. Fluorescence inner-filtering correction for determining the humification index of dissolved organic matter // Environmental science and technology. 2002. V. 36. № 4. P. 742–746. https://doi.org/10.1021/es0200692
Ovsepyan L., Kurganova I., Lopes de Gerenyu V., Kuzyakov Ya. Recovery of organic matter and microbial biomass after abandonment of degraded agricultural soils: the influence of climate // Land Degr. Dev. 2019. P. 1861–1874. https://doi.org/10.1002/ldr.3387
Ovsepyan L., Kurganova I., Lopes de Gerenyu V., Rusakov A.V., Kuzyakov Ya. Changes in the Fractional Composition of Organic Matter in the Soils of the Forest–Steppe Zone during Their Postagrogenic Evolution // Eurasian Soil Science. 2020. V. 53. № 1. P. 50–61. https://doi.org/10.31857/SS2180X20010128
Polyanskaya L.M., Zvyagintsev D.G., Lukin S.M. The change in composition of microbial biomass in cultivated soils // Eurasian Soil Science. 1997. V. 30. № 2. P. 172–177.
Post W.M., Kwon K.C. Soil carbon sequestration and land-use change: processes and potential // Global change biology. 2000. V. 6. P. 317–328. https://doi.org/10.1046/j.1365-2486.2000.00308.x
Ryzhova I.M., Erokhova A.A., Podvezennaya M.A. Dynamics and Structure of Carbon Storage in the Postagrogenic Ecosystems of the Southern Taiga // Eurasian Soil Science. 2014. V. 47. № 12. P. 1207–1215. https://doi.org/10.1134/S1064229314090117
Ryzhova I.M., Telesnina V.M., Sitnikova A.A. Dynamic of soil properties and carbon stock structure in postagrogenic ecosystems of Southern taiga during natural reforestation // Eurasian Soil Science. 2020. V. 53. № 2. P. 230–243. https://doi.org/10.1134/S0032180X20020100
Sá J.C.M., Lal R. Stratification ratio of soil organic matter pools as an indicator of carbon sequestration in a tillage chronosequence on a Brazilian Oxisol // Soil and Tillage Research. 2009. V. 103 № 1. P. 46–56. https://doi.org/10.1016/j.still.2008.09.003
Senesi N. Molecular and quantitative aspects of the chemistry of fulvic acid and its interactions with metal ions and organic chemicals. Part II. The fluorescence spectroscopy approach // Analytica Chimica Acta. 1990. V. 232. P. 77–106. https://doi.org/10.1016/S0003-2670(00)81226-X
Senesi N., Miano T., Provenzano M.R., Brunetti G. Characterization, differentiation, and classification of humic substances by fluorescence spectroscopy // Soil Science. 1991. V. 152. № 4. P. 259–271. https://doi.org/10.1097/00010694-199110000-00004
Sorokina O.A. Diagnostic parameters of soil formation in gray forest soils of abandoned fields overgrowing with pine forests in the middle reaches of the Angara river // Eurasian Soil Science. 2010. V. 43. № 8. P. 867–875. https://doi.org/10.1134/S1064229310080041
Spohn M., Novák T.J., Incze J., Giani L. Dynamics of soil carbon, nitrogen, and phosphorus in calcareous soils after land-use abandonment – A chronosequence study // Plant and soil. 2016. V. 401. № 1–2. P. 185–196. https://doi.org/10.1007/s11104-015-2513-6
Stolnikova E.V., Ananyeva N.D., Chernova O.V. The Microbial Biomass and Its Activity and Structure in the Soils of Old Forests In the European Russia // Eurasian Soil Science. 2011. V. 44. № 4. P. 437–452. https://doi.org/10.1134/S1064229311040107
Susyan E.A., Wirth S., Ananyeva N.D., Stolnikova E.V. Forest succession on abandoned arable soils in European Russia – Impacts on microbial biomass, fungal-bacterial ratio, and basal CO2 respiration activity // Eur. J. Soil Biol. 2011. V. 47. P. 169–174. https://doi.org/10.1016/j.ejsobi.2011.04.002
Telesnina V.M., Kurganova I.N., Lopes de Gerenyu V.O., Ovsepyan L.A., Lichko V.I., Ermolaev A.M., Mirin D.M. Dynamics of soil properties and plant composition during postagrogenic evolution in different bioclimatic zones // Eurasian Soil Science. 2017. V. 50. № 12. P. 1515–1534. https://doi.org/10.1134/S1064229317120109
Telesnina V.M., Vaganov I.E., Karlsen A.A., Ivanova A.E., Zhukov M.A., Lebedev S.M. Specific features of the morphology and chemical properties of coarse-textured postagrogenic soils of the southern taiga, Kostroma oblast // Eurasian Soil Science. 2016. V. 49. № 1. P. 102–115. https://doi.org/10.1134/S1064229316010117
Telesnina V.M., Zhukov M.A. The Influence of Agricultural Land Use on the Dynamics of Biological Cycling and Soil Properties in the Course of Postagrogenic Succession (Kostroma Oblast) // Eurasian Soil Science. 2019. V. 52. № 9. P. 1114–1129. https://doi.org/10.1134/S1064229319070135
Uselman S.M., Qualls R.G., Lilienfein J. Quality of soluble organic C, N and P produced by different types and species of litter: Root litter versus leaf litter // Soil Biology & Biochemistry. 2012. V. 54. P. 57–67. https://doi.org/10.1016/j.soilbio.2012.03.021
van der Wal A., van Veen J.A., Smant W., Boschker T.S., Bloem J., Kardol P., van der Putten W.H., de Boer W. Fungal biomass development in a chronosequence of land abandonment // Soil Boil. Biochem. 2006. V. 38. № 1. P. 51–60. https://doi.org/10.1016/j.soilbio.2005.04.017
Vesterdal L., Ritter E., Gundersen P. Change in soil organic carbon following afforestation of former arable land // Forest Ecol. Manag. 2002. V. 169. № 1–2. P. 137–147. https://doi.org/10.1016/S0378-1127(02)00304-3
Vladychenskii A.S., Telesnina V.M., Rumyantseva K.A., Chalaya T.A. Organic Matter and Biological Activity of Postagrogenic Soils in the Southern Taiga Using the Example of Kostroma Oblast // Eurasian Soil Science. 2013. V. 46. № 5. P. 518–529. https://doi.org/10.1134/S1064229313050141
von Lützow M., Kögel-Knabner I., Ludwig B., Matzner E., Flessa H., Ekschmitt K., Guggenberger G., Marschner B., Kalbitz K., Stabilization mechanisms of organic matter in four temperate soils: development and application of a conceptual model // J. Plant Nutr. Soil Sci. 2008. V. 171. P. 111–124. https://doi.org/10.1002/jpln.200700047|
Wang B., Liu G.B., Xue S., Zhu B. Changes in soil physicochemical and microbiological properties during natural succession on abandoned farmland in the Loess Plateau // Environmental Earth Sciences. 2011. V. 62(5). P. 915–925. https://doi.org/10.1007/s12665-010-0577-4
Wang G., Jin J., Chen X., Liu J., Liu X., Herbert S.J. Biomass and catabolic diversity of microbial communities with long-term restoration, bare fallow and cropping history in Chinese Mollisols // Plant, Soil and Environment. 2008. V. 53(4). P. 177–185. https://doi.org/10.17221/2313-PSE
Zhao F., Yang G., Han X., Feng Y., Ren G. Stratification of Carbon Fractions and Carbon Management Index in Deep Soil Affected by the Grain-to-Green Program in China // PLoS ONE. 2014. V. 9. № 6. https://doi.org/10.1371/journal.pone.0099657
Zolovkina D.F., Karavanova E.I., Stepanov A.A. Sorption of Water-Soluble Organic Substances by Mineral Horizons of Podzol // Eurasian Soil Science. 2018. V. 51. P. 1154–1163. https://doi.org/10.1134/S1064229318100162
Zsolnay A. Dissolved organic matter: Artefacts, definitions, and functions // Geoderma. 2003. V. 113. № 3–4. P. 187–209. https://doi.org/10.1016/S0016-7061(02)00361-0
Zsolnay A., Baigar E., Jimenez M., Steinweg B., Saccomandi F. Differentiating with fluorescence spectroscopy the sources of dissolved organic matter in soils subjected to drying // Chemosphere. 1999. V. 38. № 1. P. 45–50. https://doi.org/10.1016/s0045-6535(98)00166-0
Дополнительные материалы отсутствуют.