

Почвоведение, 2021, № 4, стр. 436-450
Динамика биологической активности и водорастворимого органического вещества в почвах горной тундры Хибин на склонах разной экспозиции
М. Н. Маслов a, *, О. А. Токарева a, Е. И. Караванова a, О. А. Маслова a, Е. И. Копеина b
a МГУ им. М.В. Ломоносова
119991 Москва, Ленинские горы, 1, Россия
b Полярно-альпийский сад-институт им. Н.А. Аврорина Кольского научного центра РАН
184209 Апатиты, мкр. Академгородок, 18, Россия
* E-mail: maslov.m.n@yandex.ru
Поступила в редакцию 29.06.2020
После доработки 17.08.2020
Принята к публикации 27.10.2020
Аннотация
Оценена динамика содержания и свойств водорастворимого органического вещества и биологической активности почв кустарничкового и травяного биогеоценозов горной тундры на склонах разной экспозиции. Выявлено варьирование большинства изученных показателей в течение вегетационного периода, что подтверждает важность динамических наблюдений при оценке роли почв в меняющихся условиях окружающей среды. Установлено, что почвы на юго-западном склоне характеризуются более высокой среднесуточной температурой и меньшей влажностью на протяжении всего вегетационного периода. Несмотря на это, экспозиция склона не оказывает влияния на содержание водорастворимого органического вещества и потенциальную дыхательную активность почвенных микроорганизмов. В то же время установлено, что в более теплых и сухих условиях органическое вещество почв горной тундры менее устойчиво к микробиологической трансформации. Естественный градиент температуры и влажности почвы существенным образом не сказывается на эффективности использования источников углерода и общем функционировании микробных сообществ. На основании полученных данных спрогнозирована адаптация почвенного микробного сообщества при плавном повышении температуры в Субарктике без резкого увеличения скорости минерализации почвенного органического вещества микроорганизмами.
ВВЕДЕНИЕ
В почвах арктических и субарктических экосистем аккумулировано около 14% мирового почвенного углерода [16, 32, 44], что оценивается в 190 Гт [48]. Столь значительные запасы, а также высокая чувствительность северных экосистем к воздействию внешних факторов, определяют интерес к изучению механизмов регулирования биогеохимического цикла углерода. Минерализация органического вещества сдерживается такими факторами, как кислая реакция почвенной среды, переувлажнение, низкая обеспеченность доступными формами азота и высокое содержание трудноразлагаемых субстратов [20, 24, 37, 38]. Однако главным фактором, затрудняющим разложение органического вещества в тундровых почвах, является низкая температура окружающей среды [52]. В то же время рост температуры воздуха в высоких широтах в 2 раза превышает среднемировые значения [15, 19] и в этих условиях температурные ограничения на процессы разложения могут быть смягчены, что подвергает часть почвенного органического вещества риску активной минерализации [31, 45].
Сочетание разнообразных орографических и микроклиматических условий в горной тундре создает предпосылки для формирования на сравнительно небольших площадях экосистем, различающихся по составу растений [13] и микроорганизмов, а также особенностям биологического круговорота и обеспеченности азотом [3]. Очевидно, что реакция контрастных по условиям функционирования экосистем на изменение факторов окружающей среды может различаться как по силе проявления, так и по направлению. В связи с этим прогнозирование степени, в которой потепление климата изменит минерализационную активность микроорганизмов, зависит от механистического понимания того, как потепление может взаимодействовать с другими факторами окружающей среды, воздействуя на весь комплекс почвенных условий [30, 39]. Для изучения будущих изменений в горно-тундровых экосистемах может быть применен метод экологических градиентов, в частности изучение экосистем и почв на склонах разной экспозиции, что позволяет установить изменение лабильных свойств почвы в условиях слабого естественного градиента климатических условий. Использование такого подхода для оценки реакции экосистем на изменение температуры имеет преимущество перед манипуляционными экспериментами, поскольку позволяет оценить происходящие в экосистеме процессы в более длительных временны́х рамках [46, 54]. Получение такого рода данных является неотъемлемой частью понимания того, как тундровые экосистемы будут функционировать по мере постепенного изменения климата.
Цель работы – исследование динамики содержания и свойств водорастворимого органического вещества и микробиологической активности в двух контрастных типах почв горной тундры в зависимости от экспозиции склона.
ОБЪЕКТЫ И МЕТОДЫ
Исследование проводили в Хибинах (Мурманская область). Образцы почвы отбирали в течение 2019 г. в 3 этапа, соответствующих началу (июнь), середине (конец июля–начало августа) и концу (сентябрь) вегетационного периода. Образцы почвы отбирали на северо-восточном и юго-западном склонах г. Вудъяврчорр на высоте 600–650 м над ур. м. Исследовали почвы двух горно-тундровых экосистем – кустарничковой тундры и злакового луга.
Кустарничковая тундра занимает пологие склоны и функционирует в условиях умеренного снегонакопления в зимний период. Растительный покров представлен листопадным кустарником Betula nana, а также вечнозелеными (Empetrum hermaphroditum, Vaccinium vitis-idaea, Harrimanella hypnoides, Linnaea borealis, Loiseleuria procumbens, Phyllodoce caerulea) и листопадными (Vaccinium myrtillus, Vaccinium uliginosum) кустарничками. Значительно реже здесь встречаются злаки (Agrostis canina, Anthoxanthum alpinum, Deschampsia flexuosa, Festuca ovina, Nardus stricta), осоки (Carex bigelowii) и разнотравье (Solidago lapponica). Моховый покров представлен Polytrichum juniperinum и Pleurozium sp. Среди лишайников доминируют Cetraria islandica и Cladonia arbuscula.
Злаковый луг занимает выположенные подножия склонов в условиях значительного зимнего снегонакопления. Доминирующими видами являются злаки (Avenella flexuosa, Anthoxanthum alpinum, Festuca ovina, Nardus stricta) и осоки (Carex bigelowii). В этом сообществе встречаются кустарнички Empetrum hermaphroditum и Vaccinium myrtillus. Среди разнотравья преобладают Solidago lapponica, Taraxacum sp. и др. Наиболее распространенным видом мхов является Racomitrium macrocarpon. Лишайники представлены Cetraria islandica, Cetrariella delisei, Stereocaulon alpinum.
Под кустарничковой тундрой формируется сухоторфяно-подбур иллювиально-гумусовый с профилем TJ–BH–C [4] или Entic Folic Podzols [29]. Под злаковым лугом формируется литозем перегнойно-темногумусовый потечно-гумусовый с профилем O–AH–Chi–С [4] или Haplic Leptosols [29]. Обе почвы сформированы на элювии нефелиновых сиенитов.
Температуру почвы на глубине 10 см в каждой из исследованных экосистем фиксировали каждые 4 ч с июня по сентябрь с помощью логгеров Thermochron ibutton™ (Dallas Semiconductor Corporation, TX, USA) с точностью 0.5°C. Влажность почвенных горизонтов определяли весовым методом при отборе проб почвы.
В воздушно-сухих образцах почвы исследовали общее содержание углерода (Cобщ) и азота (Nобщ) на элементном анализаторе Elementar Vario III (Германия). Значение рН определяли для водной и солевой (1 М KCl) почвенных суспензий потенциометрическим методом с помощью рН-метра Hanna HI 83141 со стеклянным электродом HI 1230B с точностью 0.01 ед. рН.
Свежие образцы почвы доставляли в лабораторию и анализировали не позднее, чем через 2 сут после отбора согласно рекомендациям [7]. В свежих образцах определяли содержание углерода и азота водорастворимого органического вещества (ВОВ). Экстракцию проводили бидистиллированной водой при температуре 20оС при соотношении почва : раствор 1 : 10. Полученные растворы фильтровали через мембранный фильтр с размером пор 0.45 мкм (ацетат целлюлозы; OE 67, Schleicherand Schuell). Определение концентрации углерода (СВОВ) и азота ВОВ проводили на автоматическом анализаторе liqui TOC Elementar (Германия). В растворах определяли абсорбцию в ультрафиолетовом спектре при длине волны 254 нм (A254) на спектрофотометре Specord 50 (Analitic Jena, Германия). Для определения степени гидрофобности и ароматичности ВОВ рассчитывали показатель удельной ультрафиолетовой абсорбции SUVA254 = (А254/СВОВ) × 100, л/(мг м) [28, 51]. Спектры флуоресценции измеряли на люминесцентном спектрометре LS-45 (Perkin Elmer, США) при длине волны возбуждения 310 нм, по ним рассчитывали индекс свежести ВОВ (β/α), равный отношению интенсивности флуоресценции при 380 нм (β) к максимуму в диапазоне 420–435 нм (α) [53]. В природных поверхностных водах, для которых первоначально был предложен данный индекс, флуоресценция β-типа обусловлена наличием автохтонного органического вещества и продуктов начальных стадий их разложения, флуоресценция α-типа – гумусовыми веществами [27, 40]. Таким образом, индекс свежести отражает долю в составе растворенных органических веществ новообразованных негумифицированных компонентов. Для ВОВ почв величина β/α характеризует долю неспецифических лабильных веществ, потенциально подверженных дальнейшей микробной трансформации.
Содержание углерода (Смикр) и азота (Nмикр) микробной биомассы определяли методом фумигации–экстракции [17, 50] как разницу между концентрацией экстрагируемого углерода и азота в фумигированных и нефумигированных навесках исследуемого образца. Экстракцию углерода и азота проводили 0.05 М раствором K2SO4. Концентрацию C и N определяли на автоматическом анализаторе liqui TOC Elementar (Германия). В вытяжках из нефумигированных образцов исследовали содержание минерального азота. Аммонийный азот определяли салицилат-нитропрусидным методом, нитратный – после восстановления нитратов до нитритов на кадмиевой колонке с получением окрашенного азосоединения при реакции с сульфаниламином и N-(1-нафтил)-этилендиамин-дигидрохлоридом. Колориметрирование окрашенных растворов проводили на спектрофотометре Genesys TM 10 UV (США).
Базальное дыхание (БД) почвы определяли после инкубации образцов почвы при полевой влажности при температуре +20°С в течение 24 ч. Скорость субстрат-индуцированного дыхания (СИД) исследовали при аналогичных температурных условиях через 3 ч после внесения в почву 0.2 мл 10%-ного раствора глюкозы. Концентрацию выделившегося С–CO2 определяли на газовом хроматографе с детектором по теплопроводности (колонка Haye Sep N с внутренним диаметром 2 мм и длиной 2000 мм, адсорбент – 80/100 Porapak Q, температура колонки 60°C, скорость потока газа-носителя (гелий) 20 мл/мин; объем вводимой пробы газа – 1 мл). Показатели БД и СИД использовали для характеристики экофизиологического состояния микробных комплексов почв [1], для чего определяли величину метаболического коэффициента qCO2 = БД/Смикр и коэффициента микробного дыхания QR = БД/СИД [2, 14].
Все эксперименты проводили в 4-кратной повторности для каждого срока отбора образцов (12-кратная повторность для каждой почвы). В таблицах и на графиках данные приведены как средние величины ± ошибка среднего, расчет выполнен на вес сухой почвы (105°С, 12 ч). Для статистической обработки результатов использовали программы Microsoft Excel и Statistica 10.0. Все измеренные показатели были оценены на нормальность распределения (тест Колмогорова–Смирнова) и гомогенность и при необходимости логарифмированы (в таблицах и на графиках данные приведены без преобразования). Для оценки достоверности различий температуры почвы в разных экосистемах использовали t-критерий для независимых переменных. Результаты проанализированы с использованием факторного дисперсионного анализа (ANOVA) с выявлением эффектов экспозиции склона, времени отбора почвенного образца, типа экосистемы и почвенного горизонта, а также их взаимодействия. Влияние факторов признавалось статистически значимым при p < 0.05. Для анализа части данных использовали метод регрессионного анализа с последовательным включением факторов.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Температура и влажность почвы. Средняя температура почвы кустарничковой тундры за вегетационный период составляет 6.9 ± 1.2°С на северо-восточном и 9.7 ± 1.9°С на юго-западном склонах. Для почвы злакового луга этот показатель больше и составляет 8.0 ± 2.0 и 10.1 ± 1.8°С соответственно (рис. 1). Максимальная температура почвы на глубине 10 см зафиксирована в июле и августе. В начале августа наблюдалось снижение температуры почвы, связанное с вторжением холодных арктических воздушных масс, что в целом характерно для климата региона. Сухоторфяно-подбур под кустарничковой тундрой характеризуется наибольшими перепадами температуры в течение вегетационного сезона по сравнению с литоземом под злаковым лугом. Для почв одноименных экосистем на разных склонах установлены статистически достоверные различия в среднесуточной температуре (p < 0.001). Также значимые различия установлены между температурой сухоторфяно-подбура и литозема на северо-восточном склоне (p < 0.001), в то время как для почв на юго-западном склоне различия не зафиксированы (p = 0.199).
Рис. 1.
Динамика среднесуточной температуры и влажности почвы кустарничковой тундры (А) и злакового луга (Б): 1 – температура почвы на северо-восточном склоне; 2 – температура почвы на юго-западном склоне; 3 – влажность горизонта TJ сухоторфяно-подбура на северо-восточном склоне; 4 – влажность горизонта TJ сухоторфяно-подбура на юго-западном склоне; 5 – влажность горизонта ВН сухоторфяно-подбура на северо-восточном склоне; 6 – влажность горизонта ВН сухоторфяно-подбура на юго-западном склоне; 7 – влажность горизонта О литозема на северо-восточном склоне; 8 – влажность горизонта О литозема на юго-западном склоне; 9 – влажность горизонта AH литозема на северо-восточном склоне; 10 – влажность горизонта АН литозема на юго-западном склоне; 11 – влажность горизонта Chi литозема на северо-восточном склоне; 12 – влажность горизонта Chi литозема на юго-западном склоне.
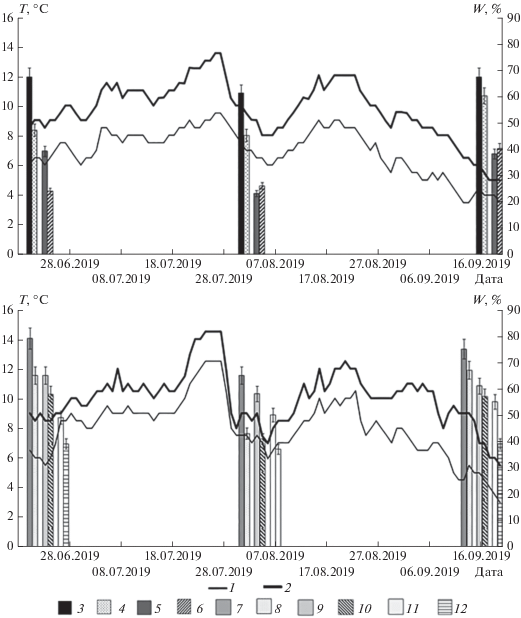
Влажность поверхностных горизонтов в почвах составляет 60–70% на северо-восточном склоне и 40–60% на юго-западном. С глубиной влажность снижается до 20–40% в минеральных горизонтах. Наименьшая влажность почвы зафиксирована в середине вегетационного сезона. Дисперсионный анализ (табл. 1) показал влияние факторов экспозиции склона, сезона вегетационного периода, типа экосистемы и горизонта на влажность почвы.
Таблица 1.
Влияние факторов (P-value) на общие свойства и влажность почв горной тундры (ANOVA)
| Фактор | Собщ, % | Nобщ, % | Cобщ : Nобщ | ${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$ | pHKCl | W, % |
|---|---|---|---|---|---|---|
| Время отбора (1) | – | – | – | <0.001 | 0.019 | <0.001 |
| Экспозиция (2) | <0.001 | <0.001 | <0.001 | 0.575 | <0.001 | <0.001 |
| Экосистема (3) | <0.001 | 0.265 | <0.001 | 0.006 | <0.001 | <0.001 |
| Горизонт (4) | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| 1 × 2 | – | – | – | 0.040 | 0.129 | 0.001 |
| 1 × 3 | – | – | – | 0.655 | 0.980 | 0.161 |
| 2 × 3 | 0.028 | 0.769 | 0.419 | <0.001 | <0.001 | 0.006 |
| 1 × 4 | – | – | – | 0.022 | 0.335 | 0.324 |
| 2 × 4 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.011 |
| 3 × 4 | <0.001 | 0.496 | <0.001 | <0.001 | <0.001 | <0.001 |
| 1 × 2 × 3 | – | – | – | 0.426 | 0.032 | 0.075 |
| 1 × 2 × 4 | – | – | – | 0.158 | 0.072 | 0.427 |
| 1 × 3 × 4 | – | – | – | 0.505 | 0.240 | 0.428 |
| 2 × 3 × 4 | <0.001 | 0.007 | 0.081 | 0.063 | 0.001 | <0.001 |
| 1 × 2 × 3 × 4 | – | – | – | 0.312 | 0.883 | 0.254 |
Физико-химические свойства почв. Горно-тундровые почвы на обоих склонах характеризуются кислой реакцией среды (табл. 2). При этом литозем под злаковым лугом имеет большие значения рН, чем сухоторфяно-подбур под кустарничковой тундрой, что может быть связано с различием в качестве поступающего растительного опада. Ранее опубликованы схожие данные по кислотности почв горной тундры Фенноскандии [10] и Хибин [11]. Экспозиция склона не влияет на величину рН водной суспензии почвы, в то время как для потенциальной кислотности не установлено значимого влияния периода вегетационного сезона.
Таблица 2.
Физико-химические свойства почв
| Экосистема | Горизонт | Собщ, % | Nобщ, % | C : N | ${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$ | pH KCl |
|---|---|---|---|---|---|---|
| Северо-восточный склон | ||||||
| КТ | TJ | 32.2 ± 2.2 | 1.23 ± 0.05 | 26.3 ± 2.0 | 4.8 ± 0.3 | 3.8 ± 0.1 |
| BH | 5.3 ± 1.2 | 0.33 ± 0.08 | 19.2 ± 1.2 | 5.5 ± 0.2 | 4.8 ± 0.1 | |
| ЗЛ | O | 41.7 ± 2.9 | 1.61 ± 0.20 | 31.4 ± 2.5 | 5.3 ± 0.1 | 4.4 ± 0.1 |
| AH | 14.6 ± 2.6 | 1.20 ± 0.18 | 15.9 ± 0.7 | 5.3 ± 0.1 | 4.7 ± 0.2 | |
| Chi | 6.9 ± 0.7 | 0.45 ± 0.07 | 19.0 ± 0.8 | 5.6 ± 0.1 | 5.1 ± 0.1 | |
| Юго-западный склон | ||||||
| КТ | TJ | 28.9 ± 1.4 | 1.47 ± 0.21 | 22.9 ± 1.5 | 4.9 ± 0.1 | 4.0 ± 0.1 |
| BH | 6.6 ± 0.9 | 0.50 ± 0.12 | 19.6 ± 0.5 | 5.7 ± 0.2 | 4.7 ± 0.1 | |
| ЗЛ | O | 35.2 ± 3.5 | 1.94 ± 0.20 | 20.9 ± 1.2 | 5.3 ± 0.1 | 4.3 ± 0.2 |
| AH | 17.3 ± 1.1 | 1.62 ± 0.11 | 13.1 ± 0.8 | 5.0 ± 0.1 | 4.1 ± 0.1 | |
| Chi | 6.4 ± 0.5 | 0.41 ± 0.09 | 17.7 ± 1.0 | 5.3 ± 0.1 | 4.5 ± 0.1 | |
Для тундровых почв характерно высокое общее содержание углерода и азота. Валовое содержание С в почве составляет около 30–40% в поверхностных органогенных горизонтах и снижается до 6–8% в минеральных горизонтах (табл. 2). Высокое содержание углерода в минеральных горизонтах, с одной стороны, объясняется миграцией органических веществ из вышележащих горизонтов в условиях промывного водного режима, а, с другой стороны, связано с присутствием большого количества подземной фитомассы (в первую очередь, тонких функциональных корней) во всей глубине почвенного профиля. Экспозиция склона оказывает влияние на общее содержание углерода в почвах (p < 0.001): на северо-восточном склоне его содержание выше, чем на юго-западном, что может быть связано с более интенсивной трансформацией растительных остатков в наиболее теплых и сухих условиях юго-западного склона.
Содержание общего азота в поверхностных горизонтах почв составляет 1.2–1.9% и с глубиной сокращается до 0.3–0.5%. Схожие данные по содержанию азота отмечены в ранее опубликованных работах в почвах горной [3, 9] и зональной [12] тундры. Экспозиция склона влияет (p < 0.001) на общее содержание азота в почвах. Однако в противоположность Собщ содержание Nобщ больше в почвах юго-западного склона. Соотношение C : N в органическом веществе почв варьирует в широких пределах от 13 до 31 в зависимости от склона и горизонта. Низкое обогащение азотом свидетельствует о слабой степени гумифицированности почвенного органического вещества. При этом органическое вещество почв на юго-западном склоне в большей степени обогащено азотом, чем в аналогичных почвах на северо-восточном склоне.
Почвы горной тундры бедны минеральным азотом (рис. 2). Аналогичные результаты были опубликованы ранее для схожих почв горной тундры северной Швеции [10]. Преобладающей формой минерального N является аммонийный азот, что в целом характерно для тундровых почв [6, 12, 23, 43]. Экспозиция склона, сезон вегетационного периода, а также тип экосистемы и почвенного горизонта оказывают значимое влияние (p < 0.001) на содержание как аммонийного, так и нитратного азота в почвах (табл. 3). Почвы на юго-западном склоне отличаются наибольшим содержанием минерального азота, что объясняется более оптимальным температурно-влажностным режимом. Роль температуры, как лимитирующего фактора процессов минерализации соединений азота в тундровых почвах, показана ранее [10, 22, 42]. Максимальное содержание минерального азота в почвах наблюдается в начале вегетационного сезона, что может быть связано с влиянием весенних циклов промораживания–оттаивания [5, 23, 25, 33]. В середине вегетации содержание минерального азота в почвах минимальное, что связано с его потреблением растениями и микроорганизмами.
Рис. 2.
Сезонная динамика содержания аммонийного и нитратного азота: 1 – начало вегетации (июнь); 2 – середина вегетации (июль–август); 3 – окончание вегетации (сентябрь); 4 – содержание аммонийного азота в почвах на северо-восточном склоне; 5 – содержание аммонийного азота в почвах на юго-западном склоне; 6 – содержание нитратного азота в почвах на северо-восточном склоне; 7 – содержание нитратного азота в почвах на юго-западном склоне.
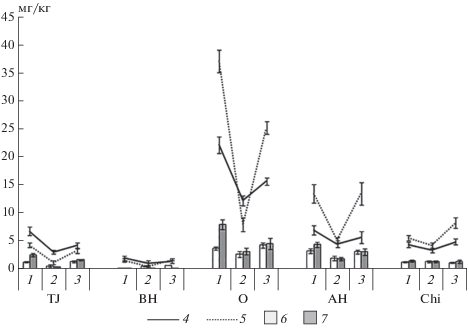
Таблица 3.
Влияние факторов (P-value) на содержание и свойства водорастворимых компонентов почв горной тундры (ANOVA)
| Фактор | СВОВ, мг/кг | СВОВ : NВОВ | N-${\text{NH}}_{4}^{ + }$, мг/кг | N-${\text{NO}}_{3}^{ - }$ мг/кг | SUVA280, л/(мг С м) | β/α |
|---|---|---|---|---|---|---|
| Время отбора (1) | 0.004 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Экспозиция (2) | 0.189 | 0.004 | <0.001 | 0.004 | <0.001 | <0.001 |
| Экосистема (3) | 0.005 | 0.014 | <0.001 | <0.001 | <0.001 | <0.001 |
| Горизонт (4) | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| 1 × 2 | 0.664 | 0.004 | <0.001 | <0.001 | 0.025 | <0.001 |
| 1 × 3 | 0.526 | 0.002 | <0.001 | 0.061 | 0.123 | 0.124 |
| 2 × 3 | 0.088 | <0.001 | <0.001 | 0.001 | 0.742 | <0.001 |
| 1 × 4 | <0.001 | <0.001 | <0.001 | <0.001 | 0.101 | <0.001 |
| 2 × 4 | 0.021 | 0.001 | <0.001 | <0.001 | 0.639 | <0.001 |
| 3 × 4 | <0.001 | 0.023 | <0.001 | 0.625 | 0.003 | <0.001 |
| 1 × 2 × 3 | 0.039 | <0.001 | <0.001 | <0.001 | 0.686 | <0.001 |
| 1 × 2 × 4 | 0.129 | <0.001 | <0.001 | <0.001 | 0.465 | <0.001 |
| 1 × 3 × 4 | 0.876 | <0.001 | <0.001 | <0.001 | 0.501 | <0.001 |
| 2 × 3 × 4 | 0.976 | 0.003 | <0.001 | <0.001 | 0.327 | 0.482 |
| 1 × 2 × 3 × 4 | 0.054 | 0.002 | <0.001 | <0.001 | 0.110 | 0.443 |
Содержание и свойства ВОВ. Содержание углерода ВОВ в горизонте TJ сухоторфяно-подбура варьирует в пределах 220–470 мг/кг и снижается в минеральном горизонте до 45–150 мг/кг (рис. 3). Для литозема характерно большее содержание ВОВ: в подстилке от 500 до 800 мг/кг, в нижележащих горизонтах – от 100 до 250 мг/кг. Содержание водорастворимого углерода в почвах наиболее тесно коррелирует с содержанием общего углерода в почве (r = 0.78, p < 0.05), при этом доля углерода водорастворимой фракции в общем пуле относительно постоянна и не превышает 0.10–0.25%. По данным множественного пошагового регрессионного анализа, вне зависимости от экспозиции склона и типа экосистемы, содержание ВОВ в почвах определяется двумя факторами: температурой почвы и общим содержанием углерода.
Рис. 3.
Сезонная динамика содержания (А) и спектральные характеристики свойств (Б) водорастворимого органического вещества почв: 1 – начало вегетации (июнь); 2 – середина вегетации (июль–август); 3 – окончание вегетации (сентябрь); 4 – содержание СВОВ в почвах на северо-восточном склоне; 5 – содержание СВОВ в почвах на юго-западном склоне; 6 – соотношение С : NВОВ в почвах на северо-восточном склоне; 7 – соотношение С : NВОВ в почвах на юго-западном склоне; 8 – индекс SUVA254 водорастворимого органического вещества почв северо-восточного склона; 9 – индекс SUVA254 ВОВ почв юго-западного склона; 10 – индекс свежести β/α ВОВ почв северо-восточного склона; 11 – индекс свежести β/α водорастворимого органического вещества почв юго-западного склона.
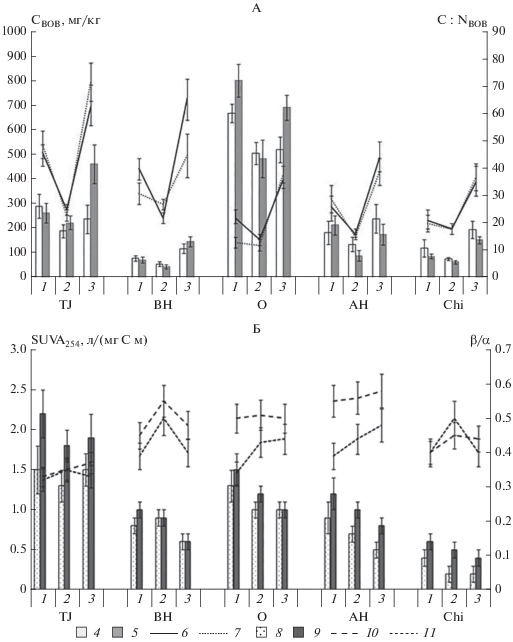
Экспозиция склона не оказывает влияния на содержание CВОВ в почвах (табл. 3). Сезон вегетационного периода оказывает влияние на содержание ВОВ: во всех почвах и горизонтах минимальное содержание CВОВ зафиксировано в середине вегетации, что связано с активизацией микроорганизмов, потребляющих ВОВ. Наибольшее содержание CВОВ в почвах приурочено к окончанию периода вегетации и связано с началом разложения свежего растительного опада. Осенний всплеск содержания ВОВ в поверхностных горизонтах почв более выражен на юго-западном склоне, что связано с более высокой температурой почвы, способствующей лучшему разложению поступающих растительных остатков.
Процессы поступления свежего растительного опада и его микробиологической трансформации определяют соотношение C : N в ВОВ горно-тундровых почв. Бóльшие значения соотношения C : N ВОВ, наблюдаемые в течение вегетационного периода во всех почвенных горизонтах, кроме подстилки злакового луга, по сравнению со средними значениями этого показателя для валового органического вещества, свидетельствуют о том, что большая часть азота почвы сохраняется в составе слаботрансформированных растительных остатков или малорастворимых органических соединений. Напротив, в подстилке злакового луга соотношение C : N водорастворимой фракции сравнимо или ниже соотношения в общем органическом веществе, что свидетельствует о слабой защищенности органического вещества от микробиологической трансформации.
Показатель SUVA254, характеризующий долю различных, преимущественно гидрофобных, веществ ароматического строения в составе ВОВ [41], в поверхностных горизонтах почв составляет 1–3 л/(мг С м), а в минеральных от 0.3 до 1 л/(мг С м). Подобное профильное распределение отражает формирование компонентов ВОВ из разлагающегося растительного опада и связано с разной степенью микробиологической трансформации ВОВ. Следует отметить, что полученные для поверхностных горизонтов почв значения ниже, чем ранее опубликованные данные для торфяных почв сфагнового болота [35], но в целом хорошо соотносятся с данными, полученными для постпирогенных почв кустарничковой тундры Хибин [8]. Значение SUVA254 зависит от типа экосистемы: ВОВ почвы под кустарничковой тундрой имеет больший индекс SUVA254, следовательно, более высокую степень ароматичности. Такое распределение связано с качеством ежегодно поступающего в почву растительного опада. Для сухоторфяно-подбура характерно ежегодное поступление лигнифицированного опада кустарничков с низким содержанием азота и фосфора, но большим количеством фенольных и других соединений ароматической структуры. В литозем поступает преимущественно опад злаков и других травянистых растений, который содержит меньшее количество лигнина и фенолов и больше азота и фосфора, что ускоряет его трансформацию микроорганизмами. Ранее показано, что для тундровых растений накопление большого количества фенольных соединений в тканях и, соответственно, растительном опаде, является адаптационным механизмом к стрессовым условиям окружающей среды, прежде всего, низкой температуре и низкой доступности питательных элементов [47]. Существенное влияние травянистых растений (люцерна) на снижение показателя SUVA254 ВОВ ранее показано для почв различных севооборотов [56]. Для почв на северо-восточном склоне характерны меньшие значения SUVA254 и меньшая степень ароматичности ВОВ по сравнению с почвами юго-западного склона. Это связано с меньшей температурой и большей влажностью почв на северо-восточном склоне, что определяет меньшую микробиологическую активность почв (см. ниже) и соответственно более продолжительное существование алифатических компонентов в составе ВОВ.
Индекс свежести β/α характеризует долю органического вещества, в которую входят растворимые продукты разложения растительных остатков, корневые экссудаты и компоненты микробной биомассы, в пуле ВОВ почвы. Значения β/α варьируют в пределах 0.3–0.6 и зависят от сезона вегетационного периода, экспозиции склона, типа экосистемы и почвенного горизонта. Низкие значения индекса свежести свидетельствуют о более высокой микробиологической активности по трансформации органического вещества [26], поэтому для почв наблюдается увеличение значений индекса β/α с глубиной. Для ВОВ литозема под злаковым лугом характерны более высокие значения индекса β/α, чем для ВОВ сухоторфяно-подбура. Установлена тенденция роста значения индекса свежести в ходе вегетационного сезона с пиком в сентябре, что связано с поступлением в почву свежего растительного опада. ВОВ литозема на северо-восточном склоне имеет более высокий индекс свежести, чем на юго-западном. Для сухоторфяно-подбура такая закономерность проявляется только в конце вегетационного периода.
Содержание углерода и азота микробной биомассы и биологическая активность почв. Содержание углерода и азота микробной биомассы в почвах варьирует в зачительных пределах (рис. 4) в зависимости от сезона вегетационного периода, экспозиции склона, типа почвы и горизонта. Доля Cмикр в общем пуле углерода почвы не превышает 1%; доля Nмикр в общем пуле азота почвы составляет 1.5–2.0%. По результатам множественного регрессионного анализа установлена взаимосвязь содержания углерода микробной биомассы с температурой почвы, общим содержанием углерода и азота, а также содержанием ВОВ. На содержание азота микробной биомассы в почвах статистически значимо влияют температура почвы, общее содержание углерода в почве, содержание СВОВ, а также соотношение C : N в водораствоимом органическом веществе.
Рис. 4.
Сезонная динамика содержания углерода (А) и азота (Б) микробной биомассы почв: 1 – начало вегетации (июнь); 2 – середина вегетации (июль–август); 3 – окончание вегетации (сентябрь); 4 – северо-восточный склон; 5 – юго-западный склон.
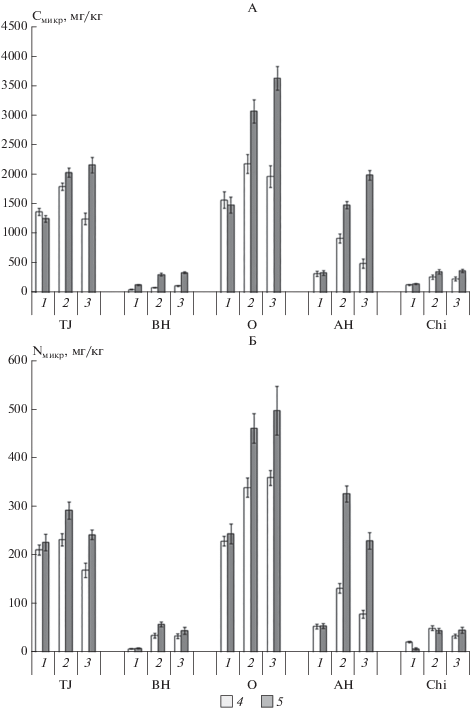
Для содержания углерода и азота микробной биомассы почвы в начале вегетационного сезона (июнь) характерно отсутствие значимых различий между почвами на разных склонах. Однако в последующем содержание элементов микробной биомассы в почвах на юго-западном склоне выше, чем на северо-восточном, что связано с различиями в температурно-влажностном режиме почвы. Для сезонной динамики содержания Cмикр и Nмикр в почвах характерен минимум содержания в начале вегетации, что связано гибелью части микроорганизмов в ходе циклов промораживания–оттаивания [23, 25, 33], характерных для суточного температурного цикла почвы в этот период. В дальнейшем наблюдается увеличение содержания углерода и азота микробной биомассы, что связано как с увеличением температуры почвы, так с активизацией микробиоты за счет разложения органического вещества, в том числе прижизненных выделений растений. Ранее прирост содержания Cмикр и Nмикр в течение вегетационного периода был показан для тундровых почв Канады [18]. В то же время на характер сезонной динамики Cмикр и Nмикр оказывает влияние экспозиция склона: к сентябрю на северо-восточном склоне наблюдается уменьшение содержания элементов микробной биомассы, в то время как на юго-западном склоне наблюдается его максимум. Такие различия связаны с разницей в температурном режиме почв – за счет более высокой температуры почвы на юго-западном склоне происходит пролонгирование периода активности.
Интенсивность БД варьирует в широких пределах от 0.5 до 38 мг С–СО2/(кг ч) (рис. 5) в зависимости от типа горизонта, экосистемы и периода вегетации. Интенсивность БД тесно коррелирует с содержанием углерода и азота микробной биомассы (r = 0.75–0.82, p < 0.05) и углеродом и азотом ВОВ (r = 0.73 мг С–СО2/(кг ч) 0.76, p < 0.05). В то же время множественный регрессионный анализ подтверждает статистически значимую связь этого показателя только с содержанием углерода микробной биомассы и водорастворимых соединений азота. Тесная связь содержания доступного азота с интенсивностью БД объяснима, поскольку тундровые почвы являются одними из наиболее бедных этим элементом [23], что вызывает лимитирование минерализации органического вещества доступностью N [21, 34, 36].
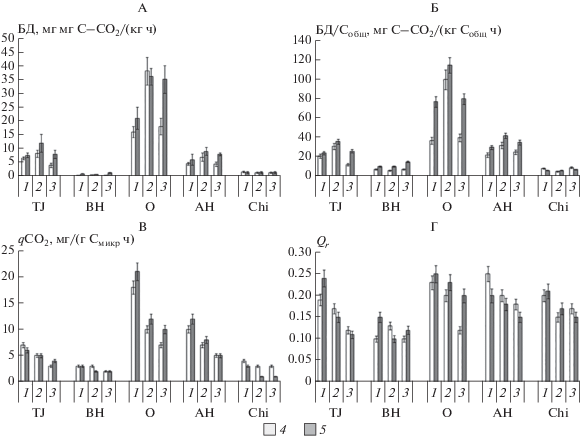
Экспозиция склона не оказывает влияния на интенсивность БД почв, однако выявлено влияние на этот показатель таких факторов, как тип почвенного горизонта, тип экосистемы и сезон вегетационного периода. Наибольшие значения БД характерны для поверхностных органогенных горизонтов почв, что связано не только с содержанием с них наибольших количеств доступного для потребления органического вещества, но с лучшими температурными условиями и аэрацией. Биологическая активность литозема под злаковым лугом выше, чем сухоторфяно-подбура. Это связано, во-первых, с более высокой среднесуточной температурой почвы в течение вегетационного периода, а, во-вторых, с ежегодным поступлением более качественного и минерализуемого растительного опада, включая большое количество опада подземной фитомассы. Сочетание этих факторов приводит к лучшей обеспеченности почвенных микроорганизмов элементами минерального питания, прежде всего, азотом. Сезонная динамика БД в почвах повторяет описанные выше тренды для содержания углерода и азота микробной биомассы и, очевидно, определяется теми же факторами, что и динамика Cмикр и Nмикр.
По сравнению с показателем БД более информативной характеристикой устойчивости органического вещества к разложению является удельная скорость минерализации, отнесенная к единице общего углерода почвы (БД/Собщ, мг/(кг С ч)). При этом, чем меньше значение отношения БД/Собщ, тем более стабильно ОВ почвы. Удельная скорость минерализации ОВ определяется склоном экспозиции, сезоном вегетационного периода и типом почвенного горизонта (табл. 4). Органическое вещество почв на северо-восточном склоне более устойчиво к минерализации, чем в аналогичных почвах на юго-западном склоне. Для поверхностных органогенных горизонтов характерна отчетливая сезонная динамика этого показателя с максимумом в середине вегетационного периода. Для минеральных горизонтов различия, связанные как с влиянием склона, так и с влиянием сезона выражены в меньшей степени, поскольку в целом органическое вещество этих горизонтов в большей степени устойчиво к микробиологическому воздействию по сравнению с поверхностными органогенными горизонтами.
Таблица 4.
Влияние факторов (P-value) на содержание микробной биомассы и показатели биологической активности почв горной тундры (ANOVA)
| Фактор | Cмикр, мг/кг | Nмикр, мг/кг | Cмикр : Nмикр | БД, мг С–CO2/(кг ч) |
БД/Собщ, мг С–CO2/(кг Собщ ч) |
СИД, мг С–CO2/(кг ч) |
qCO2, мг/(г Cмикр ч) |
Qr |
|---|---|---|---|---|---|---|---|---|
| Время отбора (1) | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Экспозиция (2) | <0.001 | <0.001 | <0.001 | 0.058 | 0.589 | <0.001 | 0.029 | 0.589 |
| Экосистема (3) | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.439 | <0.001 |
| Горизонт (4) | <0.001 | <0.001 | 0.148 | <0.001 | 0.049 | <0.001 | <0.001 | 0.049 |
| 1 × 2 | <0.001 | <0.001 | <0.001 | 0.014 | 0.450 | <0.001 | <0.001 | 0.450 |
| 1 × 3 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.077 | 0.003 | <0.001 |
| 2 × 3 | 0.132 | <0.001 | <0.001 | 0.950 | <0.001 | 0.142 | 0.061 | 0.057 |
| 1 × 4 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| 2 × 4 | <0.001 | <0.001 | <0.001 | 0.065 | <0.001 | 0.165 | 0.531 | 0.051 |
| 3 × 4 | <0.001 | <0.001 | <0.001 | <0.001 | 0.051 | <0.001 | <0.001 | 0.035 |
| 1 × 2 × 3 | 0.124 | 0.793 | <0.001 | 0.285 | 0.001 | 0.005 | 0.086 | 0.001 |
| 1 × 2 × 4 | <0.001 | <0.001 | <0.001 | 0.032 | 0.043 | 0.228 | 0.015 | 0.043 |
| 1 × 3 × 4 | <0.001 | <0.001 | <0.001 | <0.001 | 0.939 | <0.001 | <0.001 | 0.226 |
| 2 × 3 × 4 | <0.001 | <0.001 | 0.097 | 0.514 | 0.226 | 0.236 | 0.002 | 0.939 |
| 1 × 2 × 3 × 4 | 0.338 | 0.006 | <0.001 | 0.552 | 0.296 | 0.114 | <0.001 | 0.296 |
Установлено статистически значимое влияние таких факторов, как содержание азота микробной биомассы, водорастворимых соединений азота, а также соотношение C : N в общем органическом веществе почв на интенсивность СИД почвы. Важная роль азота в регулировании интенсивности СИД связана с тем, что в условиях внесения глюкозы и снятия ограничений по доступному органическому субстрату, доступность для потребления азота определяет метаболические возможности микроорганизмов. Для динамики СИД в целом характерны те же закономерности, что и для интенсивности БД.
Величина метаболического коэффициента qCO2 в изученных почвах значительно варьирует в зависимости от сезона и имеет тенденцию к постепенному снижению к концу вегетационного периода (рис. 5), что говорит об увеличении эффективности использования субстрата микроорганизмами в течение вегетационного периода. Высокие значения метаболического коэффициента qCO2 в начале вегетации могут быть объяснены преобладанием на начальных этапах сукцессий микробных популяций быстрорастущих r-стратегов, специализирующихся на утилизации легкоминерализуемых источников углерода, которыми могут быть как растительные остатки, минерализованные в течение зимнего периода, так и остатки микроорганизмов, погибших в ходе весенних циклов промораживания–оттаивания. Популяции r-стратегов очень активны при разложении органического вещества, но малопродуктивны [49, 55] и нуждаются в большом количестве энергии для поддержания своей биомассы.
Величина коэффициента QR, характеризующего соотношение скоростей БД и СИД, для всех почв колеблется в узких пределах 0.10–0.25 (рис. 5), что свидетельствует об отсутствии или слабой степени нарушения функционирования микробных сообществ почв в течение вегетационного периода [1, 2]. Экспозиция склона не оказывает значимого влияния на этот показатель, в то время как остальные изучаемые факторы (сезон вегетационного периода, тип экосистемы и почвенного горизонта) оказывают влияние на этот коэффициент.
ЗАКЛЮЧЕНИЕ
Сопряженное изучение содержания и свойств ВОВ и биологической активности почв в течение вегетационного периода позволило установить, что большинство лабильных показателей почвы имеет выраженную динамику, связанную с фазами активности растений и изменением температуры почвы. Для многих показателей характер сезонной динамики зависит не столько от типа экосистемы, как от экспозиции склона. Это подчеркивает важность проведения систематических исследований в течение всего периода вегетационной активности для определения отклика почв на меняющиеся условия окружающей среды.
Экспозиция склона не влияет на содержание ВОВ в почвах. Однако в условиях естественного дрейфа температурно-влажностных условий, ВОВ почв более теплого и сухого склона юго-западной экспозиции в большей степени трансформируется микроорганизмами. Итоговым результатом этих изменений является снижение содержания общего углерода в почвах юго-западного склона по сравнению с северо-восточным.
Несмотря на то, что в почвах на склонах с разной экспозицией выявлены значимые различия в содержании микробной биомассы, изменение условий увлажнения и обогрева не сказывается на потенциальной минерализационной активности почвенного микробного сообщества ни в почве под кустарничковой, ни под травянистой растительностью. В то же время установлено, что в более теплых и сухих условиях, органическое вещество почв горной тундры менее устойчиво к микробиологической трансформации вне зависимости от его начальных свойств. Изменение температурно-влажностного режима почв существенным образом не сказывается на эффективности использования источников углерода и общем функционировании микробных сообществ. Полученные данные позволяют спрогнозировать адаптацию почвенного микробного сообщества при плавном повышении температуры в Субарктике, что будет выражаться в формировании более гумифицированного органического вещества почвы без резкого увеличения скорости его минерализации микроорганизмами.
Список литературы
Ананьева Н.Д. Микробиологические аспекты самоочищения и устойчивости почв. М.: Наука, 2003. 226 с.
Благодатская Е.В., Ананьева Н.Д., Мякшина Т.Н. Характеристика состояния микробного сообщества почв по величине метаболического коэффициента // Почвоведение. 1995. № 2. С. 205–210.
Бузин И.С., Макаров М.И., Малышева Т.И., Кадулин М.С., Королева Н.Е., Маслов М.Н. Трансформация соединений азота в почвах горно-тундровых экосистем Хибин // Почвоведение. 2019. № 5. С. 570–577. https://doi.org/10.1134/S32180X1903002X
Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
Макаров М.И., Малышева Т.И., Мулюкова О.С., Меняйло О.В. Влияние замораживания горно-луговых альпийских почв на процессы трансформации соединений азота и углерода // Экология. 2015. № 4. С. 257–264.
Маслов М.Н., Маслова О.А., Поздняков Л.А., Копеина Е.И. Биологическая активность почв горно-тундровых экосистем при постпирогенном восстановлении // Почвоведение. 2018. № 6. С. 728–737. https://doi.org/10.1134/S1064229318060108
Маслов М.Н., Маслова О.А., Токарева О.А. Изменение лабильного и микробного пулов углерода и азота в лесной подстилке при разных способах хранения образцов // Почвоведение. 2019. № 7. С. 793–802. https://doi.org/10.1134/S0032180X19070104
Маслов М.Н., Маслова О.А., Копеина Е.И. Биохимическая устойчивость водорастворимого органического вещества почв горной тундры Хибин при постпирогенной сукцессии // Почвоведение. 2021. № 2.
Маслов М.Н., Макаров М.И. Органическое вещество почв горной тундры северной Фенноскандии // Вестник Моск. ун-та. Сер. 17, почвоведение. 2013. № 3. С. 3–7.
Маслов М.Н., Макаров М.И. Трансформация соединений азота в тундровых почвах Северной Фенноскандии // Почвоведение. 2016. № 7. С. 813–821. https://doi.org/10.7868/S0032180X16070078
Переверзев В.Н. Генетические особенности почв природных поясов Хибинских гор (Кольский полуостров) // Почвоведение. 2010. № 5. С. 548–557.
Шамрикова Е.В., Денева С.В., Кубик О.С., Панюков А.Н. Соединения азота в почвах континентальных окраин Европейского сектора Российской Арктики // Почвоведение. 2020. № 7. С. 803–815. https://doi.org/10.31857/S0032180X20070138
Шмакова Н.Ю., Ушакова Г.И., Костюк В.И. Горно-тундровые сообщества Кольской субарктики (эколого-физиологический аспект). Апатиты, 2008. 167 с.
Anderson T.H., Domsch K.H. The metabolic quotient for CO2 (qCO2) as a specific activity parameter to assess the effects of environmental conditions, such as pH, on the microbial biomass of forest soil // Soil Biol. Biochem. 1993. V. 25. P. 393–395.
Anisimov O.A., Vaughan D.G., Callaghan T.V., Furgal C., Marchant H., Prowse T.D., Vilhjalmsson H., Walsh J.E. Polar regions (Arctic and Antarctic // Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, 2007. P. 653–685.
Billings W.D. Carbon balance of Alaskan tundra and taiga ecosystems: past, present and future // Quater. Sci. Rev. 1987. V. 6. P. 165–177.
Brookes P.C., Landman A., Pruden G., Jenkinson D.S. Chloroform fumigation and the release of soil nitrogen: a rapid direct extraction method to measure microbial biomass nitrogen in soil // Soil Biol. Biochem. 1985. V. 17. P. 837–842.
Buckeridge K.M., Banerjee S., Siciliano S.D., Grogan P. The seasonal pattern of soil microbial community structure in mesic low arctic tundra // Soil Biol. Biochem. 2013. V. 65. P. 338–347. https://doi.org/10.1016/j.soilbio.2013.06.012
Callaghan T.V., Bergholm F., Christensen T.R., Jonasson C., Kokfelt U., Johansson M.A. New climate era in the sub-Arctic: Accelerating climate changes and multiple impacts // Geophys. Res. Lett. 2010. V. 37. P. 1–6. https://doi.org/10.1029/2009GL042064
Chapin F.S.III, Johnson D.A., McKendrick J.D. Seasonal movement of nutrients in plants of differing growth form in an Alaskan tundra ecosystem: implications for herbivory // J. Ecology. 1980. V. 11. P. 189–209.
Churchland C., Mayo-Bruinsma L., Ronson A., Grogan P. Soil microbial and plant community responses to single large carbon and nitrogen additions in low arctic tundra // Plant and Soil. 2010. V. 334. P. 409–421. https://doi.org/10.1007/s11104-010-0392-4
Dorrepaal E., Toet S., van Logtestijn R.S.P., Swart E., van de Weg M.J., Callaghan T.V., Aerts R. Carbon respiration from subsurface peat accelerated by climate warming in the subarctic // Nature. 2009. V. 460. P. 616–619. https://doi.org/10.1038/nature08216
Edwards K.A., McCulloch J., Kershaw G.P., Jefferies R.L. Soil microbial and nutrient dynamics in a wet Arctic sedge meadow in late winter and early spring // Soil Biol. Biochem. 2006. V. 38. P. 2843–2851. https://doi.org/10.1016/j.soilbio.2006.04.042
Freeman C., Ostle N., Kang H. An enzymic “latch” on a global carbon store // Nature. 2001. V. 409. P. 149–150. https://doi.org/10.1038/35051650
Grogan P., Michelsen A., Ambus P., Jonasson S. Freeze-thaw effects on carbon and nitrogen dynamics in subarctic heath tundra mesocosms // Soil Biol. Biochem. 2004. V. 36. P. 641–654. https://doi.org/10.1016/j.soilbio.2003.12.007
Huang W., Mcdowell W.H., Zou X., Ruan H., Wang J., Ma Z. Qualitative differences in headwater stream dissolved organic matter and riparian water-extractable soil organic matter under four different vegetation types along an altitudinal gradient in the Wuyi Mountains of China // Appl. Geochem. 2015. V. 52. P. 67–75. https://doi.org/10.1016/j.apgeochem.2014.11.014
Huguet A., Vacher L., Relexans S., Saubusse S., Froidefond J.M., Parlanti E. Properties of fluorescent dissolved organic matter in the gironde estuary // Org. Geochem. 2009. V. 40. P. 706–719. https://doi.org/10.1016/j.orggeochem.2009.03.002
Ilina S.M., Drozdona O.Yu., Lapitskiy S.A., Alekhin Y.V., Demin V.V., Zavgorodnyaya Y.A. et al. Size fractionation and optical properties of dissolved organic matter in the continuum soil solution-bog-river and terminal lake of a boreal watershed // Org. Geochemist. 2017. V. 66. P. 14–24. https://doi.org/10.1016/j.orggeochem.2013.10.008
IUSS Working Group WRB. World Reference Base for Soil Resources 2014, update 2015 International soil classification system for naming soils and creating legends for soil maps. World Soil Resources Reports № 106, Rome.
Kardol P., De Long J.R., Sundqvist M.K. Crossing the threshold: the power of multi-level experiments in identifying global change responses // New Phytologist. 2012. V. 196. P. 323–326. https://doi.org/10.1111/j.1469-8137.2012.04341.x
Kleber M. What is recalcitrant soil organic matter? // Environ. Chem. 2010. V. 7. P. 320–332. https://doi.org/10.1071/en10006
Kuhry P., Ping C.L., Schuur E.A.G., Tarnocai C., Zimov S. Report from the International Permafrost Association: carbon pools in permafrost regions // Permafrost and Periglacial Processes. 2009. V. 20. P. 229–234. https://doi.org/10.1002/ppp.648
Larsen K.S., Jonasson S., Michelsen A. Repeated freeze thaw cycles and their effects on biological processes in two arctic ecosystem types // Appl. Soil Ecol. 2002. V. 21. P. 187–195. https://doi.org/10.1016/s0929-1393(02)00093-8
Lavoie M., Mack M., Schuur E. Effects of elevated nitrogen and temperature on carbon and nitrogen dynamics in Alaskan arctic and boreal soils // J. Geophys. Res. 2011. V. 116. P. 1–14. https://doi.org/10.1029/2010jg001629
Leroy F. Effets des changements de végétation dans les tourbières à sphaignes sur le cycle du carbone. Sciences de la Terre. Université d’Orléans, 2017. Français.
Mack M.C., Schuur E.A.G., Bret-Harte M.S., Shaver G.R., Chapin F.S. Ecosystem carbon storage in arctic tundra reduced by long-term nutrient fertilization // Nature. 2004. V. 431. P. 440–443. https://doi.org/10.1038/nature02887
Moore T., Basiliko N. Decomposition in boreal peatlands // Boreal peatland ecosystems. Springer, Berlin, Heidelberg, 2006. P. 125–143.
Oechel W.C., Billings W.D. Effects of global change on the carbon balance of arctic plants and ecosystems // Arctic ecosystems in a changing climate: an ecophysiological perspective. 1992. P. 139–168.
Ostle N.J., Smith P., Fisher R., Woodward F.I., Fisher J.B., Smith J.U., Galbraith D. et al. Integrating plant-soil interactions into global carbon cycle models // J. Ecol. 2009. V. 97. P. 851–863. https://doi.org/10.1111/j.1365-2745.2009.01547.x
Parlanti E., Wörz K., Geoffroy L., Lamotte M. Dissolved organic matter fluorescence spectroscopy as a tool to estimate biological activity in a coastal zone submitted to anthropogenic inputs // Org. Geochem. 2000. V. 31. P. 1765–1781.
Piirsoo K., Viik M., Koiv T., Kairo K., Laas A., Noges T., Pall P., Selberg A., Toomsalu L., Vilbaste S. Characteristics of dissolved organic matter in the inflows and in the outflow of Lake Vortsjarv, Estonia // J. Hydrol. 2012. V. 475 P. 306–313. https://doi.org/10.1016/j.jhydrol.2012.10.015
Rustad L., Campbell J., Marion G., Norby R., Mitchell M., Hartley A., Cornelissen J., Gurevitch J. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming // Oecologia. 2001. V. 126. P. 543–562. https://doi.org/10.1007/s004420000544
Schimel J.P., Bilbrough C., Welker J.M. Increased snow depth affects microbial activity and nitrogen mineralization in two Arctic tundra communities // Soil Biol. Biochem. 2004. V. 36. P. 217–227. https://doi.org/10.1016/j.soilbio.2003.09.008
Schlesinger W.H. Biogeochemistry: An Analysis of Global Change. San Diego, 1991. 443 p.
Schmidt M.W.I., Torn M.S., Abiven S., Dittmar T., Guggenberger G., Janssens I.A., Kleber M., Kogel-Knabner I. et al. Persistence of soil organic matter as an ecosystem property // Nature. 2011. V. 478. P. 49–56. https://doi.org/10.1038/nature10386
Sundqvist M.K., Sanders N.J., Wardle D.A. Community and ecosystem responses to elevational gradients: processes, mechanisms and insights for global changes // Ann. Rev. Ecology, Evolution, Systematics. 2013. V. 44. P. 261–280. https://doi.org/10.1146/annurev-ecolsys-110512-135750
Sundqvist M.K., Wardle D.A., Olofsson E., Giesler R., Gundale M.J. Chemical properties of plant litter in response to elevation: subarctic vegetation challenges phenolic allocation theories // Functional Ecology. 2012. V. 26. P. 1090–1099. https://doi.org/10.1111/j.1365-2435.2012.02034.x
Tarnocai C., Canadell J.G., Schuur E.A.G., Kuhry P., Mazhitova G., Zimov S. Soil organic carbon pools in the northern circumpolar permafrost region // Global Biogeochemical Cycles. 2009. V. 23. P. 1–11. https://doi.org/10.1029/2008GB003327
Thiffault E., Hannam K.D., Quideau S.A., Paré D., Belanger N., Oh S.W., Munson A.D. Chemical composition of forest floor and consequences for nutrient availability after wildfire and harvesting in the boreal forest // Plant and Soil. 2008. V. 308. P. 37–53. https://doi.org/10.1007/s11104-008-9604-6
Vance E.D., Brookes P.C., Jenkinson D.S. An extraction method for measuring soil microbial biomass C // Soil Biol. Biochem. 1987. V. 19. P. 703–707.
Vergnoux A., Di Rocco R., Domeizel M., Guiliano M., Doumenq P., Theraulaz F. Effects of forest fires on water extractable organic matter and humic substances from Mediterranean soils: UV–vis and fluorescence spectroscopy approaches // Geoderma. 2011. V. 160. P. 434–443. https://doi.org/10.1016/j.geoderma.2010.10.014
Wallenstein M., McMahon S., Schimel J. Seasonal variation in enzyme activities and temperature sensitivities in arctic tundra soils // Global Change Biol. 2009. V. 15. P. 1631–1639. https://doi.org/10.1111/j.1365-2486.2008.01819.x
Wilson H.F., Xenopoulos M.A. Effects of agricultural land use on the composition of fluvial dissolved organic matter // Nature Geosci. 2009. V. 2. P. 37–41. https://doi.org/10.1038/ngeo391
Wolkovich E.M., Cook B.I., Allen J.M., Crimmins T.M., Betancourt J.L., Travers S.E., Pau S., Regetz J. et al. Warming experiments underpredict plant phenological responses to climate change // Nature. 2009 V. 485. P. 494–497. https://doi.org/10.1038/nature11014
Wüthrich C., Schaub D., Weber M., Marxer P., Conedera M. Soil respiration and soil microbial biomass after fire in a sweet chestnut forest in southern Switzerland // Catena. 2002. V. 48. P. 201–215. https://doi.org/10.1016/S0341-8162(01)00191-6
Xu N., Wilson H.F., Saiers J.E., Entz M. Effects of crop rotation and management system on water-extractable organic matter concentration, structure, and bioavailability in a chernozemic agricultural soil // J. Environ. Qual. 2013 V. 42. P. 179–190. https://doi.org/10.2134/jeq2012.0126
Дополнительные материалы отсутствуют.