

Почвоведение, 2021, № 5, стр. 582-591
Азотминерализующая способность серой лесной почвы Новосибирского Приобья при компостировании и паровании растительных остатков
Г. П. Гамзиков a, *, С. З. Сулейменов a
a Новосибирский государственный аграрный университет
630039 Новосибирск, ул. Добролюбова, 160, Россия
* E-mail: gamolgen@rambler.ru
Поступила в редакцию 29.06.2020
После доработки 14.08.2020
Принята к публикации 24.10.2020
Аннотация
Азотминерализующая способность почв характеризует интенсивность разложения органических азотсодержащих соединений и отражает способность их к накоплению минерального подвижного азота – объективного показателя обеспеченности растений основным элементом питания. Объект исследования – агросерая лесная почва (Luvic Greyzemic Phaeozem), широко используемая в сибирском полеводстве. Применяя лабораторный (компостирование при оптимальных условиях) и полевой (парование в природных условиях) методы исследований, удалось получить новую агрохимическую информацию о количественных и качественных изменениях соединений почвенного азота и азотсодержащих органических удобрений. Установлена кинетика разложения и высвобождения доступного азота из навоза и растительных остатков (донника, рапса, озимой ржи, соломы) при взаимодействии с серой лесной почвой. Проведены режимные наблюдения за выделением СО2, содержанием азота микробной биомассы, активностью целлюлозолитических бактерий и накоплением минеральных форм азота. Экспериментально показано значение растительной биомассы в поддержании азотного статуса в полевых агроценозах.
ВВЕДЕНИЕ
Серые лесные почвы Западной Сибири в основном распространены в лесостепи, подзоне подтайги и, менее, в подзоне южной тайги [1]. В Сибирском федеральном округе в пахотном земельном фонде на долю этих почв приходится более 2.7 млн га (14.2% пашни) [14]. Их значение, наряду с сопутствующими черноземами и лугово-черноземными почвами, весьма велико для производства продукции растениеводства. Этому способствуют и благоприятные климатические условия подзоны: сравнительно высокий тип увлажнения (годовое количество осадков более 350 мм) и тепловой режим (коэффициент теплообеспеченности – 0.8–0.9) [1]. Для почвенного покрова северной лесостепи характерен непродолжительный теплый период (135–140 сут), переход среднесуточных температур почвы через 10°С на глубине 20 см в конце мая весной и в третьей декаде сентября осенью. После оживления микрофлоры в почве происходит активная минерализующая деятельность микроорганизмов по разложению органических азотсодержащих веществ. Этот период, как правило, и обуславливает количественные параметры накопления в почве доступных для растений соединений азота.
В серой лесной почве содержание гумуса находится в пределах 2.8–5.6%, общего азота 0.12–0.30%, отношение С : N = 10–12, Сгк : Сфк около 1.8 [7]. Почвенный азот серой лесной почвы в основном представлен сложными органическими соединениями, среди которых преобладают негидролизуемые (77–79% от Nобщ) и трудногидролизуемые (12–13%) формы. Содержание легкогидролизуемых органических соединений элемента в этих почвах, как правило, не превышает 10% от Nобщ, тем не менее, азот этой фракции служит основным источником минеральных форм, доступных для питания растений (обменного аммония и нитратов). Суммарная доля подвижного минерального азота (N-NH4 и N-NO3) не превышает 0.5–1.5% от общего его содержания в почве.
Многолетними исследованиями установлено [6], что от накопления подвижных минеральных форм азота в почвах, обусловленных погодными условиями и агротехническими приемами, зависит обеспеченность вегетирующих растений элементом. Величина будущего урожая в значительной степени ограничивается исходным содержанием нитратов в почве перед посевом культур и количеством их накопления за счет интенсивности текущей минерализации органических соединений и нитрификации аммония под культурами.
Анализ современного состояния применения удобрений в нашей стране, проведенный Кудеяровым [11], свидетельствует, что современное российское земледелие “основывается практически на естественном плодородии почв”, а “истощительный баланс азота ведет к питательной деградации пахотных почв”. Это подтверждается крайне недостаточным уровнем применения азотных удобрений в стране (около 14 кг N/га за последние 25 лет), что приводит к потере плодородия большей части пахотных почв. Еще более низкий уровень применения удобрений в сибирском земледелии (около 9 кг/га д. в. NPK) [14]. В настоящее время без систематического применения органических и минеральных удобрений только сидераты, корневые и послеуборочные остатки полевых культур севооборота могут служить дополнительными источниками элементов минерального питания [3, 8, 21, 27]. В связи с этим в большинстве фермерских и крестьянских хозяйств получают урожаи зерновых на крайне низком уровне – 10–12 ц/га. Перспектива просматривается лишь в ряде стабильно работающих хозяйств, где применение органических и промышленных удобрений позволяет переходить к освоению интенсивных технологий земледелия с устойчивыми урожаями зерновых на уровне 25–30 ц/га [7, 21, 25].
Исследования убеждают, что послеуборочные остатки полевых культур, запахиваемая послеуборочная биомасса многолетних трав и сидератов не только способствуют пополнению в почвах органического вещества и элементов минерального питания, но и приводят к увеличению биологической их активности за счет усиления процессов иммобилизации–минерализации [9, 12, 19].
Цель работы – дать теоретическое обоснование практическому использованию процессов минерализации азота органических соединений для поддержания плодородия почв и повышения продуктивности полевых культур за счет растительной биомассы послеуборочных остатков и сидератов, а также оценить количественных параметры азотмобилизующей способности почвы и растительных остатков.
ОБЪЕКТЫ И МЕТОДЫ
Оценку азотного фонда серой лесной почвы Новосибирского Приобья и ее азотмобилизующей способности проводили в лабораторных исследованиях при моделировании почвенных процессов в оптимальных условиях температуры и влажности, а также в полевом опыте при паровании почвы [23]. Экспериментальным объектом явилась серая лесная среднесуглинистая почва опытного участка НГАУ. Агрохимическая характеристика пахотного слоя почвы: рН – 6.3, содержание гумуса – 4.7%, общего азота – 0.19%, нитратного – 6.7 мг/кг, обменного аммонийного – 1.2 мг/кг, подвижных форм (по Чирикову) фосфора – 98–128 мг/кг и калия – 62–64 мг/кг.
С целью определения интенсивности минерализации органического вещества серой лесной почвы и вносимых минеральных и биологических источников азота на азотминерализующую ее способность был заложен лабораторный опыт по 60-дневному компостированию. Почву пахотного слоя отбирали с замыкающего поля зернопарового севооборота после ячменя. Компостирование почвы проводили в оптимальных условиях (температура – 28–30°C, влажность – 60% от полевой влагоемкости). Схема опыта: 1 – без удобрений, 2 – аммоний сернокислый (Na), 3 – навоз, 4 – биомасса донника, 5 – солома пшеничная. Норма внесения сульфата аммония, навоза и растительной массы выровнена по содержанию в них азота, из расчета 30 мг N на 100 г почвы. Опыт закладывали в пластиковых сосудах (объем 1 л) с массой почвы 500 г в пятикратной повторности. К просеянной и отобранной от растительных остатков почве добавляли, перемешивая, измельченные органические (навоз, донник, солома) и минеральное удобрение (Na). Почвенные образцы для анализа на содержание минеральных форм азота (N-NO3 и N-NH4) и азота микробной биомассы отбирали через 7, 15, 30 и 60 дней.
Полевые наблюдения за азотминерализующей способностью серой лесной почвы проводили в течение всего лета в парующихся делянках на том же поле, где отбирали почву для компостирования. Мелкоделяночный опыт закладывали в семикратной повторности, площадь делянки 1 м2, размещение рендомизированное. Схема опыта: 1 – контроль; 2 – навоз; растительная масса: 3 – донника, 4 – рапса, 5 – озимой ржи; 6 – солома пшеничная. Норма внесения растительной массы и навоза выровнена по количеству в них азота (из расчета 90 кг/га). Воздушно-сухую массу растений, тщательно перемешивая, вносили в размолотом виде в пахотный слой почвы. Для равномерного контакта вносимых удобрений почву вынимали на полиэтиленовую пленку и перемешивая, вносили растительную массу, затем удобренную почву возвращали на прежнее место. Каждую делянку с боков на глубину 0–20 см ограничивали пленкой.
Отбор почвенных образцов с делянок для анализа проводили через 15 дней на глубину 0–20 см и через 30 дней на глубину 0–20 и 20–40 см. В свежих образцах определяли подвижный минеральный азот (N-NO3 и N-NH4), а также азот биомассы микроорганизмов, в воздушно-сухих – углерод общий и лабильного органического вещества, азот общий и легкогидролизуемый. Определение продуцирования СО2 при компостировании и при паровании в течение теплого периода проводили при суточной экспозиции 2 раза в неделю.
Агрохимические анализы почв проведены в лаборатории “Современные проблемы экспериментальной агрохимии” НГАУ – испытательном центре по агрохимии (аккредитация Госстандартом России № РОСС RU.0001.514898) общепринятыми методами [2, 17]. Азот биомассы микроорганизмов почвы определяли по Brookes et al. [28], эмиссию СО2 – абсорбционным методом [26], углерод лабильного органического вещества по Тюрину в модификации [10], интенсивность разложения целлюлозы – методом аппликации [16]. Полученные результаты исследований статистически обработаны с помощью компьютерной программы SNEDECOR [22]. В тексте использованы условные обозначения: Собщ – углерод общий; Слов – углерод лабильного органического вещества, Nобщ – общий азот; Nлг – легкогидролизуемый; Nтг – трудногидролизуемый; Nнг – негидролизуемый; N-NО3 – нитратный; N-NН4 – обменно-поглощенный аммонийный; Nб. м. – азот биомассы микроорганизмов.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Азотмобилизующая способность серой лесной почвы при компостировании. Потенциал азотмобилизующей способности почвы зависит от ее биологических и агрохимических свойств, качественного состава органических и минеральных удобрений и условий, при которых проходят процессы минерализации. Двухмесячное компостирование серой лесной почвы с органической биомассой практически во всех вариантах не влияло на увеличение содержания в ней общего углерода и азота (табл. 1). Исключением служил вариант с соломой, где при преобладании углеродистой части, значительно возрастала (в 1.4–1.6 раза), в сравнении с контролем, доля общего углерода в почве, что изменяло отношение С : N.
Таблица 1.
Изменение содержания углерода, азота и их соединений в серой лесной почве под влиянием удобрений при компостировании, %
| Вариант | Собщ | Nобщ | С : N | Cлов, % | Nлг, мг/кг | |
|---|---|---|---|---|---|---|
| от массы почвы | от Собщ | |||||
| Исходное | 2.7 | 0.192 | 14 | 0.30 | 12.9 | 115 |
| 30 суток | ||||||
| Контроль | 2.7 | 0.192 | 14 | 0.33 | 14.2 | 118 |
| (NH4)2SO4 | 2.5 | 0.183 | 13 | 0.32 | 13.5 | 180 |
| Навоз | 2.9 | 0.218 | 13 | 0.37 | 12.8 | 155 |
| Донник | 2.9 | 0.208 | 13 | 0.36 | 15.0 | 176 |
| Солома | 3.9 | 0.198 | 20 | 0.61 | 15.5 | 188 |
| НСР05 | 0.3 | 0.022 | – | 0.02 | – | 21 |
| 60 суток | ||||||
| Контроль | 2.6 | 0.182 | 14 | 0.33 | 12.6 | 111 |
| (NH4)2SO4 | 2.7 | 0.185 | 15 | 0.37 | 13.7 | 136 |
| Навоз | 2.7 | 0.180 | 15 | 0.41 | 15.2 | 152 |
| Донник | 2.7 | 0.175 | 15 | 0.38 | 14.1 | 164 |
| Солома | 4.2 | 0.173 | 24 | 0.54 | 12.8 | 189 |
| НСР05 | 0.2 | 0.046 | – | 0.03 | – | 20 |
Растительные остатки с низким содержанием азота и преобладанием углерода в биомассе способствуют, по данным Будажапова [5], увеличению гетеротрофной микрофлоры в почве, что приводит к существенному потреблению и закреплению минерального азота микроорганизмами. Возросшие в варианте с соломой величины содержания легкогидролизуемых форм углерода и азота в составе органического вещества почвы служат подтверждением этого процесса. Расчеты показывают, что содержание Cлов в серой лесной почве находится в тесной зависимости от суммарного количества общего углерода в почве и во внесенной органической биомассе.
Содержание легкогидролизуемого азота в период компостирования почвы варьировало в значительных пределах и зависело от вносимых удобрений и срока отбора образцов. После месячной инкубации доля мобильных органических соединений азота заметно повышается во всех вариантах опыта в сравнении с исходным количеством. При этом максимум легкогидролизуемых соединений азота возрастает как за счет иммобилизации, так и частичного разложения трудногидролизуемой фракции [6].
Наибольшее накопление Nлг наблюдается в вариантах с внесения соломы и донника. Отмеченные изменения в накоплении Слов и Nлг находятся в тесной зависимости от микробиологической активности почвы. В процессе разложения биомассы растений микроорганизмы трансформируют эти формы углерода и азота в потенциально минерализуемое состояние [18, 29].
Наблюдения за динамикой накопления азота в биомассе микроорганизмов позволяют судить о высокой их активности при благоприятных условиях компостирования и наличии минерального и органических источников азота (рис. 1). На контроле темпы развития микробиологической жизни, судя по низкому исходному содержанию Nб. м. (4.8 мг/кг), были весьма сдержанны на протяжении всего периода инкубации. Высокая активность аммонифицирующих и нитрифицирующих микроорганизмов в первые же 7 дней компостирования, судя по количеству закрепленного азота в их биомассе, наблюдалась во всех вариантах опыта, как при внесении минерального азота, так и органических соединений азота. Количество Nб. м. при внесении сульфата аммония к 60‑му дню компостирования постепенно уменьшается до уровня на контроле, что свидетельствует о полном завершении нитрификации аммония минерального удобрения.
Рис. 1.
Влияние удобрений на изменение содержания азота микробной биомассы в серой лесной почве при компостировании, мг/кг.
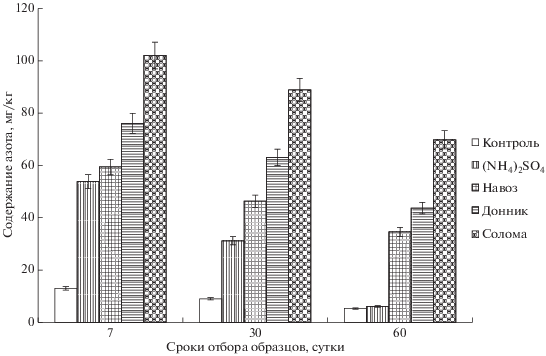
В процессе разложения навоза, донника и соломы в первые 7 дней отмечены высокие темпы иммобилизации минерального азота в состав биомассы микроорганизмов. Несмотря на постепенное количественное снижение Nб. м. в вариантах с внесением навоза и массы донника, содержание его оставалась на достаточно высоком уровне. Солома на протяжении всего периода компостирования способствовала высокой иммобилизации азота микроорганизмами. Установлено [13, 18], что интенсивность минерализационно-иммобилизационных процессов при разложении растительных остатков во многом обусловлена отношением С : N в них. Статистически достоверно доказано, что между количеством внесенного углерода с удобрениями и азотом микробной массы существует прямая корреляционная зависимость (r = 0.72 ± 0.06).
Важным показателем биологической активности почвы служит продуцирование ею диоксида углерода в атмосферу (табл. 2). В первые 10 дней компостирования во всех вариантах опыта отмечена наибольшая активность выделения СО2 (40–45% от всего количества, учтенного за весь период инкубации). Экспериментальные данные свидетельствуют, что на контроле и при внесении минерального азота выделение двуокиси углерода было ниже по сравнению с вариантами внесения органических веществ. При компостировании растительных остатков интенсивность выделения СО2 обусловлена составом внесенных растительных остатков. При этом характерной особенностью для всех вариантов, включая навоз, оказалось интенсивное выделение двуокиси углерода в атмосферу в первые две декады (около 60–70%).
Таблица 2.
Динамика выделения СО2 серой лесной почвой под влиянием удобрений при компостировании, мг/сосуд
| Вариант (В) | Периоды экспозиции, сутки (А) | Сумма за 60 суток, кг/га | |||||
|---|---|---|---|---|---|---|---|
| 1–10 | 11–20 | 21–30 | 31–40 | 41–50 | 51–60 | ||
| Контроль | 227.6 | 66.6 | 49.3 | 63.6 | 48.4 | 51.2 | 506.7 |
| (NH4)2SO4 | 223.6 | 64.4 | 60.7 | 67.8 | 44.6 | 42.6 | 503.7 |
| Навоз | 273.4 | 127.9 | 83.2 | 92.7 | 61.1 | 54.0 | 658.3 |
| Донник | 981.0 | 229.3 | 113.1 | 106.0 | 71.8 | 57.4 | 1558.6 |
| Солома | 988.5 | 450.9 | 299.3 | 263.1 | 226.5 | 179.7 | 2407.4 |
| Среднее по фактору А (НСР05 = 4.8); по фактору В (НСР05 = 2.7) | |||||||
Во второй половине инкубационного периода при преобладании нитрификационных процессов темпы выделения СО2 стабилизируются на уровне 40–60 мг/сосуд. Наибольшая активность эмиссии двуокиси углерода свойственна для массы донника и соломы, при этом активное разложение первого из них заканчивается лишь к последней декаде опыта, а второй продолжается в течение всего периода компостирования, не прекращая активного выделения диоксида.
Наблюдения за режимом обменно-поглощенного аммония показали, что нативное содержание этой формы азота в серой лесной почве на протяжении всего срока компостирования небольшое – 1.5–5.4 мг/кг (табл. 3). Максимальное количество восстановленной формы минерального азота отмечалось на седьмые сутки после закладки опыта, в последующие сроки определения содержание аммонийного азота, как правило, не превышало 3 мг/кг, что свидетельствует об одновременно высоких темпах двух процессов при компостировании – минерализации и нитрификации. Высокая активность нитрификации аммонийного азота прослеживалась при внесении сульфата аммония – за 60 сут инкубации N-NH4 почти полностью трансформировался в нитраты. При внесении органических источников азота в исследуемую почву, в связи с высокой активностью нитрификационных процессов, во всех вариантах содержание обменного аммонийного азота оставалось на низком уровне.
Таблица 3.
Динамика содержания подвижных форм минерального азота в серой лесной почве при длительном компостировании по суткам, мг/кг
| Вариант | N-NH4 | N-NO3* | |||||||
|---|---|---|---|---|---|---|---|---|---|
| исходное | 7 | 15 | 30 | 60 | 7 | 15 | 30 | 60 | |
| Контроль | 2.0 | 5.4 | 1.5 | 1.4 | 3.0 | 6.9 | 18.5 | 35.4 | 75.5 |
| (NH4)2SO4 | 301.3 | 222.5 | 172.5 | 85.4 | 6.3 | 6.3 | 25.5 | 126.7 | 259.5 |
| Навоз | 2.0 | 5.0 | 1.6 | 1.6 | 3.6 | 8.7 | 34.9 | 52.2 | 91.4 |
| Донник | 4.5 | 14.0 | 3.3 | 3.3 | 3.1 | Cл. | 18.6 | 68.7 | 132.0 |
| Солома | 1.7 | 3.9 | 1.9 | 0.6 | 2.9 | Cл. | Cл. | Cл. | Cл. |
| НСР05 | – | 9.6 | 8.5 | 3.1 | 1.1 | 3.0 | 1.8 | 3.8 | 1.7 |
Одновременные наблюдения за режимом нитратов подтвердили высокую нитрифицирующую способность серой лесной почвы. Перед закладкой опыта в ней содержались лишь следы N-NO3, в ходе инкубации почвы в оптимальных условиях количество нитратного азота на контроле увеличивалось высокими темпами на протяжении всего срока наблюдений и достигло максимального значения (75.5 мг/кг). Достаточно энергично и полностью (в течение 60-дневного компостирования) прошла трансформация аммония в нитраты при внесении минерального удобрения. Под влиянием навоза с первых дней после внесения, также отмечено активное накопление N-NO3, существенно (в 1.2–1.8 раза) превышающее контроль на протяжении компостирования. При внесении биомассы донника в первую неделю опыта преобладал процесс аммонификации, затем активизировалась нитрификация. Пшеничная солома способствовало практически полной иммобилизации минерального азота почвы на протяжении всего опыта.
Таким образом, двухмесячное компостирование серой лесной почвы при оптимальных условиях с добавлением минерального и органических источников азота позволило выявить ряд особенностей их азотмобилизующей способности. Высокая активность оживления микробиологической деятельности почвы в первой декаде инкубации, практически во всех вариантах опыта, способствует быстрому включению части азота и углерода в состав биомассы микроорганизмов. При этом активность процессов минерализации внесенного органического вещества определяется его составом. Скорость процессов: минерализация – аммонификация – нитрификация, как правило, одинаково высокая, о чем можно судить по конечному продукту – накоплению нитратов. В течение инкубации успешно трансформируются аммоний минерального удобрения, навоза и высокобелковой биомассы донника в нитраты. Весьма медленно этот процесс осуществляется при разложении и минерализации соломы в связи с широким отношением в биомассе С : N.
Азотминерализующая способность серой лесной почвы при паровании в природных условиях. В сибирском земледелии в условиях короткого теплого периода на процессы мобилизации почвенного органического вещества и растительных остатков оказывают комплексное влияние, как погодные условия (температурный режим и увлажнение), так и агротехнические приемы (предшественник, обработка, удобрения). От них в основном зависит активность биоты почвы и накопление мобильных азотсодержащих соединений [5, 7, 25]. Результаты исследований подтверждают, что наиболее благоприятные условия для минерализации соединений азота в регионе создаются в паровом поле при отвальной вспашке (табл. 4).
Таблица 4.
Динамика содержания минерального азота в серой лесной почве при паровании и внесении органических удобрений, мг/кг
| Вариант (В) | Срок отбора образцов, дней (А) | |||||||
|---|---|---|---|---|---|---|---|---|
| 15 | 30 | 45 | 60 | 75 | 90 | 105 | 120 | |
| Обменный аммонийный азот (N-NH4) (исходное содержание – 1.2 мг/кг) | ||||||||
| Контроль | 1.2 | 1.3 | 2.2 | 2.0/1.3* | 2.5 | 1.8 | 1.8 | 2.4/1.4* |
| Навоз | 1.5 | 1.7 | 2.1 | 1.9/1.4 | 2.5 | 2.1 | 2.1 | 2.6/1.3 |
| Донник | 2.3 | 1.7 | 2.4 | 2.9/1.4 | 2.8 | 2.0 | 2.0 | 2.2/1.2 |
| Рапс | 2.4 | 2.1 | 2.6 | 2.2/1.7 | 2.8 | 2.1 | 2.1 | 1.9/1.1 |
| Рожь | 2.4 | 1.5 | 2.6 | 2.2/1.4 | 2.6 | 2.2 | 2.2 | 2.3/1.2 |
| Солома | 2.2 | 1.6 | 2.5 | 2.2/2.0 | 2.6 | 2.2 | 2.2 | 1.5/1.4 |
| Средние по фактору А (НСР05 = 0.2); средние по фактору В (НСР05 = 0.2) | ||||||||
| Нитратный азот (N-NO3) (исходное содержание – 6.7 мг/кг) | ||||||||
| Контроль | 7.2 | 8.7 | 9.7 | 19.7/7.0 | 16.2 | 18.4 | 15.6 | 16.9/8.9 |
| Навоз | 7.2 | 9.2 | 8.4 | 19.5/7.4 | 20.5 | 23.8 | 18.3 | 19.3/16.6 |
| Донник | 6.1 | 8.9 | 9.2 | 22.6/6.9 | 22.2 | 34.1 | 27.1 | 24.4/10.1 |
| Рапс | 7.5 | 12.0 | 12.7 | 21.8/8.3 | 24.9 | 32.0 | 30.9 | 26.1/19.5 |
| Рожь | Сл. | 1.6 | 8.9 | 15.0/5.4 | 22.1 | 22.3 | 18.5 | 13.6/15.1 |
| Солома | 1.9 | Сл. | 7.3 | 14.2/3.1 | 12.2 | 12.7 | 2.7 | 7.6/11.5 |
| Средние по фактору А (НСР05 = 2.5), средние по фактору В (НСР05 = 2.2) | ||||||||
Содержание аммонийного азота почвы, в связи с иммобилизацией и одновременно нитрификацией на протяжении всего периода парования, не превышало 3 мг/кг, как в пахотном, так и подпахотном слоях. При этом, если в первую половину наблюдений отмечены достоверные положительные различия в содержании N-NH4 по вариантам внесения органического вещества в сравнении с контролем, то во вторую – практически все выравнивается на низком уровне в связи с динамичной его нитрификацией. Недостаточная скорость накопления нитратов в почве в течение первых 15 дней связана вначале со слабыми темпами минерализации мобильного органического вещества из-за недостатка тепла, а затем в последующие 30 дней – в связи с высокой иммобилизацией минерального азота [7]. В дальнейшем (45–60 дней) при активной нитрификации содержание N-NO3 существенно возрасталo до максимума дифференцированно по вариантам опыта.
Динамика образования нитратов в неудобренной серой лесной почве при паровании свидетельствует о среднем уровне ее азотмобилизующей способности: на 60–70 день парования в почве накопилось 18.7 мг/кг N-NO3. Следовательно, потенциальные возможности исследуемой почвы при компостировании (75.5 мг/кг) в полевых условиях реализуется только на 25%.
Применение навоза и растительной массы в большинстве вариантов позволяло увеличивать азотминерализующую способность. При минерализации массы рапса и донника получено 32–34 мг/кг N-NO3, что обусловлено в основном узким отношением C : N в их биомассе (16 и 19 соответственно). Большое количество нитратного азота (22–24 мг/кг) получено при разложении навоза и массы озимой ржи (С : N – 22 и 42 соответственно). Внесение же пшеничной соломы с очень широким отношением углерода к азоту (74) значительно сдерживало процессы ее минерализации на протяжении всего парования почвы, в связи с чем, накопление N-NO3 ограничивалось на уровне 12–14 мг/кг.
Значительные изменения в содержании нитратного азота наблюдались в подпахотном слое серой лесной почвы. Если до 60-х сут парования существенных различий по вариантам в слое 20–40 см не было выявлено, то во второй половине лета, за счет в основном миграционного перераспределения на вариантах внесения рапса, навоза и озимой ржи, обнаруживается большое количество N-NO3 в подпахотном слое. Прежние исследования с 15N [6] показали, что в сибирских почвах миграционных потерь нитратов, как правило, не бывает. Нитраты пахотного и подпахотного слоев под влиянием нисходящего тока влаги за счет летних дождей могут опускаться во вторую половину полуметра, но после их прекращения поднимаются с восходящей миграцией влаги и остаются доступными не только для первой, но и для двух последующих культур севооборота. Оценивая растительную массу, используемую в опытах, по доступности к разложению по классификации [29], которую приводят в обзоре Семенов и Ходжаева [20], биомасса донника относится к быстроразлагаемой, рапса и навоза – к умеренно, озимой ржи – к медленно- и соломы – к слаборазлагаемой источникам доступного азота. Особенности разложения растительной биомассы и количественные параметры накопления продуктов минерализации в опытах подтверждают возможность использования этой шкалы для характеристики темпов минерализации азота растительных остатков в сибирских условиях.
Азот микробной биомассы является ближайшим резервом и постоянном соучастником процессов минерализации азота органических соединений [5, 25]. Определено, что в среднем за сезон в пахотном слое серой лесной почвы может содержаться до 150–200 кг/га микробного азота или 6–10% от общего его содержания [4]. Количественная оценка азота, включаемого в биомассу микроорганизмов, показывает, что на удобренных вариантах в первый период парования серой лесной почвы наблюдаются активные темпы закрепления азота в состав микроорганизмов (табл. 5). Активное закрепление азота в составе микробной биомассы с максимумом на 30-й день (более 20 мг/кг) свойственно для навоза и всех видов растительных остатков, с максимумом в донниковой массе. К 60-му дню парования доля азота в биомассе микроорганизмов постепенно уменьшается за счет текущей минерализации. На контроле при дефиците свежего органического вещества в течение всего периода наблюдений отмечено пониженное содержания иммобилизованного азота.
Таблица 5.
Влияние органических удобрений на содержание азота биомассы микроорганизмов в серой лесной почве при паровании, мг/кг
| Вариант | Сроки отбора образцов, сутки | ||
|---|---|---|---|
| 15 | 30 | 60 | |
| Контроль* | 7.2 | 15.4 | 10.8 |
| Навоз | 11.2 | 20.8 | 12.3 |
| Донник | 17.4 | 35.5 | 23.4 |
| Рапс | 15.1 | 28.3 | 22.4 |
| Озимая рожь | 18.3 | 21.7 | 15.3 |
| Солома | 11.4 | 23.5 | 25.6 |
| НСР05 | 3.3 | 4.7 | 4.2 |
Полевая оценка выделения СО2 показала, что в серой лесной почве в начале наблюдений на всех вариантах внесения энергетического материала отмечена низкая способность к его продуцированию (рис. 2). Лишь на 30-е сутки активность его выделения постепенно и неоднозначно возрастала по вариантам опыта, в большинстве их максимальная интенсивность продуцирования СО2 (до 70%) приходилось на период 90–120 дней. Внесение органической биомассы существенно увеличивало эмиссию СО2 на протяжении всех сроков наблюдений. Максимальное его выделение свойственно для биомассы озимой ржи, далее по убывающей достаточно продуктивными были растительные остатки донника, рапса и пшеничной соломы. Интенсивность выделения диоксида в варианте с навозом в первый период парования была минимальной, затем увеличивалась, но значительно уступала таковой при внесении растительных остатков. Суммарная эмиссия углерода в серой лесной почве за сезон парования под влиянием органических удобрений составила (т/га): на контроле – 1.6, внесении навоза – 1.9, донника – 2.7, рапса – 2.6, озимой ржи – 3.3 и пшеничной соломы – 2.4 т/га. Следовательно, интенсивность и суммарное количество выделения СО2 из почв в основном зависит от содержания и вида органического вещества.
Рис. 2.
Интенсивность продуцирования СО2 серой лесной почвой при паровании с внесением навоза и растительной массы, кг/га.

Использование остатков растительного происхождения в качестве органических удобрений и заделка их в почву активизирует рост и развитие почвенной биоты, тем самым вызывая интенсивное дыхание и выделение СО2 из почвы в атмосферу. При этом, чем больше внесено растительного материала с высоким содержанием углерода в почву, тем менее интенсивно проходят процессы минерализации [10, 24, 27].
Известно, что поступающие в почву растительные остатки наполовину представлены клетчаткой, являющейся существенным источником энергии для функционирования микробиоценозов почвы. В опыте проведены определения интенсивности разложения целлюлозы льняной ткани в серой лесной почве при внесении разных видов органического вещества (% от исходного веса):
| Экспозиция | Контроль | Навоз | Донник | Рапс | Рожь | Солома | НСР05 |
| 30 дней | 21.8 | 33.6 | 32.6 | 38.0 | 23.8 | 26.1 | 4.0 |
| 60 дней | 58.0 | 80.5 | 82.7 | 86.4 | 66.5 | 57.4 | 6.9 |
Как свидетельствуют данные, наиболее интенсивное разложение целлюлозы наблюдалось в середине лета при 60-дневной экспозиции – в период лучшего прогревания и увлажнения почвы. Биомасса рапса, донника и навоза позволила в 1.4–1.7 раза активизировать целлюлозолитическую способность почвы в сравнении с контролем. Биомасса ржи в меньшей степени, а солома пшеницы практически не оказывала влияния на активность разложения полотна в сравнении с контролем. Уменьшение веса льняного полотна достигало максимальных значений через 60 дней, при этом активное разрушение целлюлозы отмечено как и в предыдущий срок наблюдений, при внесении в почву навоза, растительной массы рапса и донника (в 1.5–1.7 раза больше контроля).
Наблюдения за азотминерализующей способностью серой лесной почвы, навоза и растительных остатков (донника, рапса), как свидетельствуют полевые исследования, реализуются при паровании на 25–26% от возможного потенциала, полученного при компостировании. Несмотря на частичную реализацию накопления нитратного азота в неудобренной почве (18.7 мг/кг) и при внесении навоза, массы донника и рапса (23.8–34.1 мг/кг), следует признать, что этого количества доступного азота в соответствии градациями обеспеченности [7] достаточно для получения полноценного урожая полевых культур в первый год после пара. Реализация потенциального запаса легкоминерализуемых органических соединений и нитратов после удобренных паров, как показывают многолетние исследования сибирских агрохимиков [3, 7, 27], бывает достаточно для двух–трех последующих культур.
ЗАКЛЮЧЕНИЕ
Длительное компостирование почвы и вносимых удобрений при оптимальных условиях позволило оценить их возможности по накоплению доступного азота. Определен высокий потенциал биологической мобилизации азота при 60-суточном компостировании для серой лесной почвы – 75 мг/кг, навоза – 91 мг/кг и массы донника – 132 мг/кг. При компостировании пшеничной соломы, в связи с широким отношением С : N и преобладанием процессов иммобилизации, выявлено почти полное закрепление минерального азота.
В полевых условиях потенциальная азотмобилизующая способность серой лесной почвы реализуется на 25% от компостируемой. Органические удобрения (навоз, биомасса донника, рапса, озимой ржи) оказывают положительное влияние на биологическую активность серой лесной почвы, повышая количество азота биомассы микроорганизмов и эмиссию СО2, усиливая целлюло-золитическую активность и минерализационные процессы, что способствует накоплению минерального подвижного азота. Под влиянием растительных остатков в почве при паровании накапливается до 32–34 мг/кг нитратного азота, что в 3–4 раза больше исходного содержания N-NO3 и в 1.2–1.7 раза в неудобренном фоне. Применение соломы на протяжении первого года парования сдерживает минерализационные процессы в почве за счет иммобилизации почвенного азота.
Скорость и полнота разложения органического вещества внесенных удобрений (навоз, биомасса донника, рапса, озимой ржи и пшеничной соломы) как при компостировании, так и при паровании в основном зависит от их качественного состава. Способность органической массы растений к микробному разложению обусловлена, прежде всего, отношением С : N в них: масса донника (15) относится к быстроразлагаемой, рапса (19) и навоза (22) – к умеренно-, озимой ржи (40) – к медленно- и соломы (74) – к слаборазлагаемым источникам доступного азота. Следовательно, в практическом земледелии с целью эффективного использовании растительных остатков и сидеральной массы необходимо, учитывая их азотмобилизующий потенциал, чередовать в севообороте посевы зерновых с бобовыми и капустными культурами, а при запашке больших доз злаковой соломы – добавлять азотные удобрения.
Для сибирского земледелия широкое использование растительных источников элементов питания полевых культур может служить перспективным и эффективным приемом поддержания плодородия почв и существенного повышения продуктивности полевых культур не только в действии, но и в последействии. Широкое комплексное применение растительной остатков всех видов возделываемых культур, сидератов и органических отходов должно быть неотъемлемой составной частью обязательных приемов в технологиях современного адаптивно-ландшафтного земледелия.
Список литературы
Агрохимическая характеристика почв СССР. Районы Западной Сибири. М.: Наука, 1968. 383 с.
Агрохимические методы исследования почв. М.: Наука, 1975. 656 с.
Берзин А.М. Зеленые удобрения в Средней Сибири. Красноярск: Красноярский ГАУ, 2002. 395 с.
Благодатский С.А., Паников Н.С., Самойлов Т.И. Влияние агротехнических приемов на динамику запасов микробного азота в серой лесной почве // Почвоведение. 1989. № 2. С. 52–59.
Будажапов Л.В. Биокинетический цикл азота и оборот азотных пулов. М.: ВНИИА, 2019. 288 с.
Гамзиков Г.П. Агрохимия азота в агроценозах. Новосибирск: РАСХН НГАУ, 2013. 790 с.
Гамзиков Г.П. Азот в земледелии Западной Сибири. М.: Наука, 1981. 266 с.
Гамзиков Г.П., Кулагина М.Н. Изменение содержания гумуса в почвах в результате сельскохозяйственного использования: обзорная информация. М.: ВНИИТЭИагропром, 1992. 48 с.
Завалин А.А., Соколов О.А. Потоки азота в агроэкосистеме: от идей Д.Н. Прянишникова до наших дней. М.: ВНИИА, 2016. 591 с.
Когут Б.М., Булкина Л.Ю. Сравнительная оценка воспроизводимости методов определения лабильных форм гумуса в почвах // Почвоведение. 1987. № 4. С. 143–145.
Кудеяров В.Н. Агрогеохимические циклы углерода и азота в современном земледелии // Агрохимия. 2019. № 12. С. 3–15. https://doi.org/10.1134/S000218811912007X
Кудеяров В.Н. Азотно-углеродный баланс в почве // Почвоведение. 1999. № 1. С. 73–82.
Кудеяров В.Н., Заварзин Г.А., Благодатский С.А. и др. Пулы и потоки углерода в наземных экосистемах России. М.: Наука, 2007. 315 с.
Межрегиональная схема размещения и специализации сельскохозяйственного производства в субъектах Российской Федерации Сибирского федерального округа: рекомендации. Новосибирск: ФГБУ СО АН, 2016. 255 с.
Методическое руководство по проектированию применения удобрений в технологиях адаптивно-ландшафтного земледелия. М.: Минсельхоз РФ, РАСХН, 2008. 392 с.
Методы почвенной микробиологии и биохимии / Под ред. Д.Г. Звягинцева М.: Изд-во Моск. ун-та, 1991. 304 с.
Практикум по агрохимии / Под ред. В.Г. Минеева. М.: Изд-во Моск. ун-та, 2001. 689 с.
Семенов В.М., Когут В.М. Почвенное органическое вещество. М.: ГЕОС, 2015. 233 с.
Семенов В.М., Семенов А.М., Ван Бругген А.Х.К., Феррис Х., Кузнецова Т.В. Трансформация азота почвы и растительных остатков сообществом микроорганизмов и микроскопических животных // Агрохимия. 2002. № 1. С. 5–11.
Семенов В.М., Ходжаева А.К. Агроэкологические функции растительных остатков в почве // Агрохимия. 2006. № 7. С. 63–81.
Сорокин И.Б. Органическое вещество в адаптивно-ландшафтных системах земледелия Томской области. Томск, 2007. 304 с.
Сорокин О.Д. Прикладная статистика на компьютере. Новосибирск: СО РАСХН, 2004. 162 с.
Сулейменов С.З. Азотмобилизующая способность почв Западной Сибири и Северного Казахстана. Автореф. дис. … канд. с.-х. наук. Новосибирск: НГАУ, 2009. 20 с.
Тулина А.С. Влияние температуры, влажности и внесения соломы на динамику минерализации органического вещества и почвенные пулы углерода и азота // Агрохимия. 2019. № 3. С. 3–18. https://doi.org/10.1134/S0002188119030141
Чупрова В.В Углерод и азот в агроэкосистемах Средней Сибири. Красноярск: Красноярский госуниверситет, 1997. 166 с.
Шарков И.Н. Абсорбционный метод определения эмиссии СО2 из почв // Методы исследования органического вещества почв. М.: Россельхозакадемия. 2005. С. 401–407.
Шарков И.Н. Роль растительных остатков зерновых культур в регулировании плодородия почв Сибири // Биологические источники элементов минерального питания растений. Новосибирск: РАСХН. Сиб. отд-ние, 2006. С. 69–77.
Brookes O.C., Landman A., Pruden G., Jenkinson D.S. Chloroform fumigation and the release of soil nitrogen: a rapid direct extraction method to measure microbial biomass nitrogen in the soil // Soil Biol. Biochem. 1985. № 17. P. 837–842.
Praveen-Kumar, Jagadish C. Tarafdar, Jitendra Panwar, Shyam Kathju. A rapid method for assessment of plant residue quality // J. Plant Nutrition Soil Sci. 2003. V. 166. P. 662–666.
Дополнительные материалы отсутствуют.