

Почвоведение, 2021, № 5, стр. 592-605
Использование особенностей структурно-функциональной организации подстилок для оценки интенсивности круговорота в городских насаждениях (на примере Москвы)
О. В. Семенюк a, *, В. М. Телеснина a, Л. Г. Богатырев a, А. И. Бенедиктова a
a МГУ им. М.В. Ломоносова
119991 Москва, Ленинские горы, 1, Россия
* E-mail: olgatour@rambler.ru
Поступила в редакцию 14.07.2020
После доработки 21.10.2020
Принята к публикации 24.10.2020
Аннотация
Изучены годовая динамика, компонентный состав, общие запасы органического вещества и зольных элементов в подстилках насаждений г. Москвы. В исследованных фитоценозах преобладают подстилки деструктивного типа; высока доля легкореализуемого органического вещества (62–90%), что позволяет оценить скорость круговорота как высокую и типичную для аналогичных естественных насаждений. Для оценки трансформации органического вещества в годовом цикле в рамках методики изучения биологического круговорота введено понятие “реализация подстилок”, под которым подразумевается совокупность абиотических и биотических процессов преобразования подстилок, приводящих к изменению состава и свойств после листопада в годичном цикле функционирования в период между окончанием листопада в текущем году и началом листопада в следующем. Предложено 3 метода оценки вовлечения органического вещества подстилок в круговорот. Первый метод основан на использовании типологии лесных подстилок. Второй заключатся в сопоставлении запасов подстилки в послелистопадный период и долистопадный следующего года, третий – в сопоставлении общих запасов легкоразлагаемых и устойчивых составляющих в сочетании с анализом компонентного состава подстилок и фракционного состава детрита. Показано, что при однотипной направленности процессов преобразования органического вещества подстилок в условно эталонных насаждениях ботанического сада и парковых насаждениях характеристики их сезонной динамики детерминированы в большей степени видом дерева-эдификатора, нежели характером антропогенного воздействия. Максимальные запасы подстилки выявлены для кленовника (800 г/м2) ботанического сада, а минимальные для березняка (250 г/м2) парковой территории, что соответствует закономерности, установленной для общей мощности подстилок. Подстилки липовых насаждений характеризуются наибольшей долей фракции листьев и легкоразлагаемых компонентов (около 60%).
ВВЕДЕНИЕ
Зеленые насаждения – это рефугиум биоразнообразия для урбанизированных территорий, выполняющий множество экологических функций [33, 48], поэтому одной из важных задач при решении проблем оптимального функционирования городских экосистем является формирование и мониторинг функционирования древесных насаждений. Функционирование озелененных территорий основывается на биологическом круговороте и определяет устойчивость городских экосистем.
Важнейшим звеном биологического круговорота признана подстилка как наиболее динамичный компонент экосистемы. Подстилка – особое природное образование, которое не всеми почвоведами признается как почвенный горизонт [4]. Тем не менее, подстилка характеризуется мощностью, запасом, составом и строением. Она играет несомненную роль в поддержании нормального функционирования экосистем и круговороте питательных веществ в них, депонировании углерода. Она же является базисом первичного почвообразования, влагозащитным напочвенным слоем, а также индикатором лесовозобновительного и лесовосстановительного потенциала лесных экосистем [10, 12, 28, 39, 41]. Карпачевский [12] отмечал, что подстилки, как и почвы, имеют горизонтное строение в соответствии со степенью разложения материала. Существует несколько различных подходов к классификации подстилок, большая часть из них фокусируется на их конкретных свойствах [3, 18].
В городских условиях подстилка, как и кроны деревьев, выполняет функции биогеохимического барьера на пути загрязняющих веществ из атмосферы. Деревья способны задерживать до 40 кг атмосферной пыли [29], а подстилки соответственно аккумулируют пылевые частицы, что приводит к увеличению зольности их компонентов [13, 50].
В пределах города почвенный покров представлен спектром самых разных почв – от малонарушенных почв садов и парков до так называемых техногенных поверхностных образований [15] – в зависимости от положения в элементарных городских ландшафтах – от парково-рекреационных до промышленных [23]. Уникальными городскими озелененными территориями являются ботанические сады, которые можно рассматривать в качестве условных экологических эталонов [22], где активно проводятся фундаментальные научные исследования [6, 25, 40, 43]. Городские почвы существенно отличаются от естественных как строением профиля, так и физико-химическими и химическими свойствами [11, 16, 22–25, 28, 45–47, 54], поскольку не только являются в той или иной степени искусственно созданными образованиями, но и подвержены как рекреации, так и атмосферному загрязнению. Как правило, естественная последовательность горизонтов в почвах города нарушена, и они содержат большую долю так называемых артефактов (кирпичей и других включений). Поскольку часто почвы бывают насыпными, то есть их создают с помощью искусственных грунтов, многие химические свойства почв могут слабо отражать современные факторы почвообразования. Таким образом, ведущим фактором почвообразования становится антропогенная деятельность [23].
Биологический круговорот в экосистемах, в том числе и городских, включает две группы процессов: синтез и разложение органического вещества. Разложение органического вещества детерминировано как биотическими, так и абиотическими факторами (промораживание–оттаивание опада с последующим выщелачиванием важнейших биофильных элементов, процессы иссушения–увлажнения, влияние интенсивности солнечной радиации) [9, 31, 34, 36, 51, 55]. Важнейшую роль в круговороте органического вещества играет биота. В разложении детрита участвуют как микробные сукцессии – от грибов к бактериям [32, 35, 49, 56], так и почвенная фауна [37, 42, 52]. Интегральная роль биоты в отечественном классическом почвоведении получила отражение в показателях гумусного состояния почв, где кроме характеристик подстилки приводится интенсивность дыхания [8].
Система диагностики функционирования древесных насаждений в городских условиях на основе методик классического изучения биологического круговорота в настоящее время не выработана. Одним из принятых методов определения интенсивности круговорота в наземных фитоценозах продолжает оставаться отношение запасов подстилки к опаду, получившее свое место в классификации биологического круговорота, до сих пор являющейся примером долговременной научной концепции [17, 44]. На этой основе оценивается ежегодное количество органического вещества и зольных элементов, которое поступает в циклы круговорота в результате сложных процессов, обусловливающих преобразование годичного опада. При проведении стационарных исследований об этом судят на основе многолетних наблюдений за динамикой поступления опада, последующего слежения за процессами минерализации органического вещества и сопоставления запасов подстилки и ежегодного опада посезонно, что теоретически основано на интенсивности потоков [2, 7]. Однако проведение такого мониторинга далеко не всегда осуществимо в условиях городских ландшафтов, особенно слежение за динамикой опада. В значительно большей степени доступно изучение подстилок.
В настоящее время работ, посвященных особенностям лесных подстилок, гораздо меньше, чем посвященных собственно почвам. Несмотря на наличие довольно обширных данных по особенностям почв в разных функциональных зонах города, подстилки таких почв изучены крайне мало.
Цель исследования – оценка биологического круговорота в городских экосистемах на основе сравнительного анализа типологии и структурно-функциональной организации подстилок древесных насаждений. Задачи работы включали разработку комплексного метода для исследования биологического круговорота с использованием показателей подстилок, оценку направленности и скорости биологического круговорота в древесных насаждениях, изучение влияния дерева-эдификатора на показатели подстилок, оценку влияния антропогенного воздействия на свойства подстилок городских насаждений.
ОБЪЕКТЫ И МЕТОДЫ
Территория Москвы относится к зоне хвойно-широколиственных лесов. Озелененные территории МГУ формировались при строительстве научно-учебного комплекса в 50-х годах ХХ в., древесные насаждения ботанического сада и парковых зон имеют примерно одинаковый возраст.
Объектами исследования выбраны наиболее распространенные в Москве типы насаждений: кленовые (Acer platanoides), березовые (Betula pendula) и липовые (Tilia cordata). Были изучены 3 типа насаждений в двух вариантах – условно-эталонный (ботанический сад) и с периодическим уходом (парковая территория МГУ). Периодический уход за городскими насаждениями включает удаление подроста и подлеска, сбор и вывоз подстилки, выкашивание травостоя, которые применяются один раз в несколько лет. В условно эталонных биогеоценозах сада такой уход не проводят. Территории, не относящиеся к саду, несомненно, испытывают и рекреационное воздействие.
Для решения поставленных задач детально исследовалась структурно-функциональная организация подстилок. Первая задача сводится к определению классификационной принадлежности подстилок, что дает основание для оценки сравнительной скорости преобразования органического вещества с использованием таксономии подстилок [3]. Вторая задача – раскрыть особенности годичного цикла преобразования растительного опада в условиях древесных насаждений на основе определения запасов подстилок дважды за сезон. Первое определение проводят в осенний период, который приурочен ко времени максимального поступления опада на почву (конец октября–начало ноября). Второе определение осуществляют в конце летнего периода (последние числа августа), когда отмечается максимальная степень разложения подстилок и, соответственно, минимальные их запасы в годовом цикле, но массовое поступление опада текущего года еще не началось. Разница запасов органического вещества подстилок (г/м2), определенных в летний и осенний периоды, дает основание для оценки той части органического вещества, которая включается в круговорот в результате интегральных процессов разложения и гумификации растительных остатков. В рамках методики изучения круговорота для удобства оценки трансформации органического вещества в годовом цикле введено понятие “реализация подстилок”, под которым подразумевается совокупность абиотических и биотических процессов преобразования подстилок, приводящих к изменению состава и свойств после листопада в годичном цикле функционирования. Реализация выражается в уменьшении запасов подстилки в течение определенного периода времени в результате разложения органического вещества. Период реализации подстилки составляет в данном случае 1 год за вычетом времени интенсивного листопада, а именно от окончания листопада (примерно конец октября) до начала периода листопада в следующем году (конец августа–начало сентября). Таким образом, величина реализации измеряется путем вычисления разности запасов подстилок, определенных в осенний период текущего года и летний период следующего и измеряется в г/(м2 год), причем под “годом”, естественно, подразумевается вышеназванный период функционирования подстилки в отсутствие интенсивного поступления опада. Третья задача исследований – оценить относительную скорость разложения подстилки с использованием комплексного анализа соотношения содержания трудноразлагаемых и легкоутилизируемых ее составляющих и данных фракционного состава.
В фитоценозах образцы подстилок отбирались с площадки 50 × 50 см в пятикратной повторности до и после листопада – в конце лета и начале осени. Морфологическая характеристика подстилок, включая мощность и строение, определялась в полевых условиях, типовая принадлежность подстилок устанавливалась на основе морфогенетической классификации [3, 5]. Верхние горизонты подстилок, представляющие собой опад текущего года, разбирались на следующие компоненты: ветви, листья, кора, ветошь, семена, детрит и т. д. Под ветошью понимают опад травянистых растений. К детриту были отнесены растительные остатки, по морфологическим признакам которых было невозможно определить их принадлежность к какой-либо фракции подстилки. Фракционный состав детрита определяли на основе ситового анализа с разделением фракций по размерности: больше 10, 10-7, 7-5, 5-3, 3-2, 2-1, 1-0.5 и меньше 0.5 мм. Определение содержания золы в компонентах подстилок проводилось методом сухого озоления при температуре 450°С [1].
Рассчитывали следующие показатели, связанные с биологическим круговоротом: 1 – общие запасы подстилки (г/м2); 2 – запас легкоразлагаемых компонентов (сумма фракций ветоши и листьев, г/м2) и его доля от общего запаса горизонта (%); 3 – запас фракции детрита (г/м2) и его доля от общего запаса горизонта (%); 4 – общие запасы зольных элементов в подстилках (г/м2); 5 – средневзвешенная зольность горизонта L подстилки (%); 6 – запас золы в легкоразлагаемых компонентах (г/м2) и его доля в запасе зольных элементов горизонта L (%).
Количество ежегодно реализуемого органического вещества (РОВ) в подстилке рассчитывали по разности запасов легкоразлагаемой части подстилки (ЛРП) после листопада и перед следующим периодом листопада, а его доля – от запасов после листопада:
Аналогичный расчет был проведен для трудноразлагаемой части подстилки (ветвей и детрита). В результате был получен баланс как для легкоразлагаемых, так и для устойчивых компонентов – со знаком (–), если в процессе реализации запас компонентов уменьшался, и со знаком (+), если увеличивался.
Для статистической обработки полученных результатов использовали программы Excel и Statistica.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Состояние насаждений, почвы. Результаты полевых исследований показали, что основное отличие зеленых насаждений с регулярным уходом от насаждений ботанического сада состоит в полном отсутствии в них подлеска, а подрост отмечен в виде единичных всходов, что повышает роль ветрового переноса в латеральной миграции наземного опада. Анализ условий, в которых функционируют фитоценозы, показывает, что парковые насаждения являются более открытым пространством по сравнению с ботаническим садом [20, 26]. Фитоценозы ботанического сада, напротив, существенно продуктивнее и разнообразнее в отношении подроста, что особенно характерно для березняка. Спецификой мелколиственных березовых фитоценозов является форма и размер ажурной кроны, которая определяет их относительно невысокую сомкнутость (0.5) по отношению к широколиственным породам, где эта величина составляет до 0.8. В силу этого обстоятельства различия в световом режиме между мелколиственными и широколиственными фитоценозами весьма существенны. Это приводит к тому, что проективное покрытие травяного яруса в березовых насаждениях достигает 90%, тогда как в широколиственных не превышает 40%.
Почвы исследуемых объектов парковой территории и ботанического сада (Technic Anthrosols) сформированы на насыпных грунтах, привнесенных при строительстве и организации территории МГУ, и представляют собой целенаправленно созданные техногенные поверхностные образования [6, 25]. Почвенный профиль характеризуется залеганием органо-минерального плодородного горизонта мощностью 6–16 см легкого и среднего гранулометрического состава на серии насыпных техногенных горизонтов. В пределах профиля встречаются многочисленные антропогенные включения, в том числе и карбонатного состава. На глубине 15–20 см начинает резко увеличиваться плотность почвы и пропадает выраженная структура. Все это позволяет отнести эти почвы к группе квазиземов и подгруппе урбиквазиземов [14].
Типология и мощность подстилок. Исследуемые подстилки древесных насаждений по классификации Богатырева [3] соответствуют двум типам: деструктивному или ферментативному. Большая часть подстилок представлено одним горизонтом – опадом прошлых лет, и относится к деструктивному типу. Ферментативная подстилка была определена только под березовыми насаждениями в ботаническом саду, для нее характерно наличие двух горизонтов – деструктивного и ферментативного. Формирование второго горизонта происходит из-за пониженной скорости разложения опада.
Мощность подстилок после листопада колеблется в широких пределах и составляет от 0.7 до 5.2 см, что соответствует литературным данным и позволяет отнести их к очень маломощным и маломощным [3]. Полученные значения мощности исследуемых подстилок в целом совпадают с другими данными по лиственным насаждениям [19]. На парковой территории и в ботаническом саду максимальная мощность отмечена у подстилки под насаждениями клена (соответственно 4.2 и 5.2 см), что связано с размером и формой листовой пластины, определяющей рыхлое сложение данного горизонта. После периода реализации опада мощность подстилки всех насаждений уменьшается. Максимальное уменьшение мощности – в 2.5–3 раза – отмечено в подстилках кленовых насаждений.
Запасы подстилок. Как и мощности подстилки, их запасы характеризуются значительной пространственной и временной изменчивостью. После листопада запасы подстилки составляют от 250 до 800 г/м2, что соответствует полученным данным по подстилкам лиственных парковых насаждений [19]. Максимальные запасы подстилки выявлены для кленовника ботанического сада (рис. 1), а минимальные – для березняка парковой территории, что не противоречит закономерности, установленной для общей мощности подстилок. Запасы подстилок остальных насаждений значимо не различаются. После периода реализации наблюдается уменьшение запасов подстилок до 120–640 г/м2, связанное с их разложением. Результаты исследований показывают, что в ботаническом саду обнаруживается тенденция к сокращению общих запасов подстилок в ряду березовые–липовые–кленовые насаждения при отсутствии значимых различий – в среднем от 570 до 450 г/м2. Иное распределение обнаруживается в том же ряду древостоев для парковых насаждений.
Рис. 1.
Запасы подстилки, г/м2. Обозначения здесь и далее в рисунках 2 и 4: А – после листопада, Б – до листопада на следующий год; Ботанический сад: 1 ‒ березовые насаждения; 2 — липовые насаждения; 3 — кленовые насаждения; парковые насаждения: 4 — березовые насаждения; 5 — липовые насаждения, 6 – кленовые насаждения. Здесь и на рис. 4, 5: I – среднее, II – среднее + стандартная ошибка, III – среднее + стандартное отклонение.
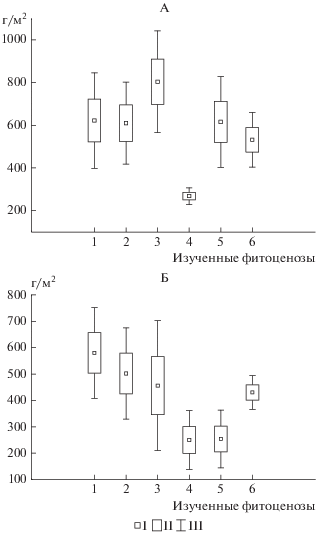
Пространственное варьирование запасов подстилок условно эталонных насаждений ботанического сада после периода реализации больше, чем после листопада, также оно превышает варьирование запасов подстилки на парковой территории. Упрощение структуры паркового фитоценоза уменьшает разнообразие условий разложения поступившего опада с точки зрения его биохимического состава, а также в связи с разнообразием экологических ниш, в которых происходит его разложение. Причем именно после периода реализации опада эта неоднородность становится очевидной. По этой же причине варьирование морфологических свойств подстилок (мощности, строения, компонентного состава) данных насаждений для всех сроков отбора больше, чем на парковой территории. В отличие от подстилок мелколиственных березовых насаждений, для подстилок широколиственных пород установлены значимые различия в запасах двух сроков отборов до и после листопада.
Легкоразлагаемые компоненты подстилок. Легкореализуемую часть, представленную ветошью и листвой, не случайно относят к так называемым “горячим точкам”, так как именно опад листьев и трав является первым компонентом, заселяемым микроорганизмами, и обусловливает основную массу образующегося углекислого газа при разложении [38]. Установлено, что запасы легкоразлагаемых компонентов (ЛРК) после листопада составляют от 100 до 380 г/м2. Распределение этих компонентов подстилок после листопада в парковых насаждениях и в ботаническом саду схожи с распределением общих запасов подстилок. Максимальные запасы листьев и ветоши выявлены для кленовых фитоценозов в ботаническом саду. После периода реализации этот показатель уменьшается за счет разложения органического вещества и составляют 15–141 г/м2, что соответствует данным, приводимым в литературе для лиственных фитоценозов [19, 26]. Доля разлагаемого за год органического вещества в подстилке довольно высока – в подстилках ботанического сада она варьирует от 72 до 85%. В подстилках березовых насаждений запасы ЛРК невелики (100 г/м2), однако скорость их разложения достаточно высока: доля ежегодно реализуемого органического вещества составляет около 85% (табл. 1). В пределах парковых насаждений доля легкореализуемого органического вещества подстилки колеблется в большем диапазоне – от 62 до 90%. Значительное варьирование запасов ЛРК и их доли в подстилках парковых насаждений связано, по-видимому, с особенностями ветрового перераспределения ЛРК подстилок в условиях более открытых пространств. Полученные отрицательные значения баланса ЛРК подстилки говорят о выраженном процессе разложения органического вещества. Исходя из полученных данных, в березняках интенсивность разложения органического вещества самая низкая. В пределах насаждений ботанического сада отмечается важнейшая тенденция, заключающаяся в возрастании интенсивности круговорота от мелколиственных насаждений к широколиственным. На результаты расчетов запасов подстилки в сторону как увеличения, так и уменьшения, может влиять ветровой перенос опада [27, 50], перераспределение ЛРК подстилки в пределах биогеоценоза и вынос за его границы. Тот факт, что запасы трудноразлагаемых компонентов подстилок увеличиваются за период наблюдения, может также объясняться латеральным ветровым переносом, а также тем, что поступление таких компонентов, как ветви и кора, происходит круглогодично.
Таблица 1.
Оценка интенсивности трансформации различных компонентов подстилок после листопада в 2016 г. и до него – в 2017 г.
| Эдификатор | Запас компонентов подстилки, г/м2 | Доля компонентов подстилки, % | Ежегодный баланс | |||
|---|---|---|---|---|---|---|
| после листопада | до листопада | после листопада | до листопада | запас, г/м2 | доля, % | |
| Легкоразлагаемые компоненты | ||||||
| Ботанический сад | ||||||
| Береза | 181.5 ± 5.3* | 28.9 ± 5.0 | 29.0 ± 3.2 | 4.9 ± 0.5 | –152.6 | –84.0 |
| Липа | 264.7 ± 6.3 | 43.5 ± 3.9 | 43.4 ± 3.8 | 10.0 ± 0.4 | –221.2 | –83.5 |
| Клен | 357.4 ± 10 | 90 ± 9.3 | 44.4 ± 5.3 | 19.8 ± 1.2 | –267.4 | –74.8 |
| Зеленые насаждения территории МГУ | ||||||
| Береза | 101.9 ± 7.0 | 14.8 ± 2.4 | 38.0 ± 2.5 | 6.0 ± 0.9 | –87.1 | –85.5 |
| Липа | 378.3 ± 12.2 | 140.8 ± 7.7 | 61.4 ± 8.1 | 55.6 ± 5.2 | –237.5 | –62.7 |
| Клен | 259.7 ± 10.1 | 26.5 ± 3.8 | 48.8 ± 4.7 | 6.8 ± 1.2 | –233.2 | –89.8 |
| Трудноразлагаемые компоненты | ||||||
| Ботанический сад | ||||||
| Береза | 441 ± 10.2 | 551 ± 21.2 | 71 ± 3.8 | 95.1 ± 2.9 | +110 | +24.9 |
| Липа | 345 ± 9.5 | 382 ± 10.2 | 56.6 ± 6.3 | 90.0 ± 5.8 | +37 | +10.7 |
| Клен | 447 ± 12.4 | 366 ± 12.3 | 55.6 ± 4.1 | 80.2 ± 6.8 | –81 | –18 |
| Зеленые насаждения территории МГУ | ||||||
| Береза | 166 ± 7.6 | 235 ± 8.9 | 62.0 ± 2.9 | 94 ± 5.8 | +69 | +41.5 |
| Липа | 238 ± 7.9 | 113 ± 5.7 | 38.6 ± 3.6 | 44.4 ± 2.3 | –125 | –52.5 |
| Клен | 272 ± 11.6 | 376 ± 12.3 | 51.1 ± 5.2 | 93.2 ± 5.6 | +104 | +38 |
Анализ полученных данных показал, что комплексный подход с использованием таких показателей, как общие запасы подстилок, запасы и доля ЛРК и устойчивых компонентов информативен для изучения функционирования подстилок.
Компонентный состав подстилок не только отражает влияние видов деревьев-эдификаторов, но и интересен в динамическом отношении, так как он показывает изменения соотношений устойчивых и легкореализуемых составляющих в годичном цикле [53]. В подстилках широколиственных насаждений после листопада на парковой территории и в ботаническом саду доля листьев составляет от 34 до 66% и является преобладающей (рис. 2). Особенностью подстилок березовых насаждений независимо от типа ухода и времени отбора является преобладание такого компонента, как тонкие ветви (до 66%). Долевое содержание детрита небольшое: от 7% в липняке до 12% в березняке ботанического сада.

Анализ годовой динамики компонентного состава подстилок показывает, что в конце периода реализации долевое участие компонентов подстилки изменяется – доля листьев уменьшается в 2–9 раз и составляет от 4 до 34%. В компонентном составе подстилок на парковой территории и в ботаническом саду под всеми насаждениями после периода реализации преобладают ветви – от 38 до 66%. Долевое содержание детрита остается неизменным или увеличивается и составляет от 2 до 20%, что связано с поступлением мелких фракций в детрит после разложения опада, а в парковых насаждениях за счет добавления мелких частиц после механического воздействия в процессе его уборки. Увеличение доли детрита в подстилках парковых насаждений с уходом, а также в результате рекреации, отмечалось в литературе [19, 45, 46]. В ботаническом саду для условно эталонных подстилок в ряду береза – липа – клен наблюдается увеличение доли листьев и фракции плодов и семян по причине ботанических особенностей этих видов в связи с величиной листьев и плодов. Кроме того, в том же ряду выявлено уменьшение доли детрита. Максимальная доля детрита в подстилке березы связана с замедленным круговоротом веществ по отношению к другим биогеоценозам, что проявляется в наличии ферментативного горизонта в данных подстилках.
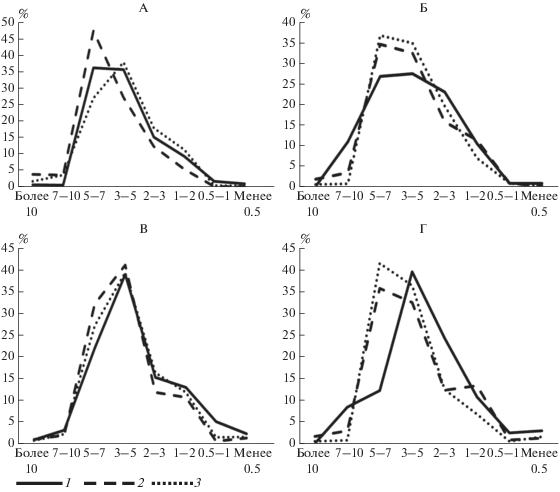
Фракционный состав детрита. Изучение фракционного состава детрита показало, что основная доля в подстилках фитоценозов ботанического сада и в пределах парковых насаждений представлена фракциями размерностью 7–5 и 5–3 мм. Минимальное содержание приходится на самые крупные фракции (7–5 мм) и мелкие – менее 1 мм (рис. 3), что как особенность распределения фракций детрита было отмечено в более ранних исследованиях [19]. Фракционный состав детрита, как и компонентный состав подстилок, отражает влияние деревьев-эдификаторов и их роль в формировании подстилок. В пределах парковых насаждений и в ботаническом саду в детрите подстилок березовых фитоценозов после периода реализации увеличивается доля крупных фракций 7–10 мм, что является источником поступления слаборазложившихся растительных остатков и материалом для формирования второго ферментативного горизонта. В ботаническом саду для подстилок широколиственных пород, которые приняты как условно эталонные, по мере разложения опада наблюдается иная тенденция – увеличение доли мелких фракций (2–3 мм) и уменьшение доли фракций 5–7 мм. На парковых территориях отмечено, напротив, увеличение доли крупных фракций (5–7 мм), что, по-видимому, связано с поступлением частиц, образующихся при механическом разрушении опада при его уборке. В то же время увеличение содержания крупных фракций происходит при одновременном уменьшении содержания мелких (менее 1–2 мм), что может указывать на усиление процесса разложения. Расчеты показали, что для всех насаждений в любые сроки отбора средневзвешенные размеры частиц составляют 3–5 мм. Исключением является детрит подстилки в липняке ботанического сада, где в процессе годовой реализации средневзвешенный размер частиц уменьшается с 5 до 2 мм. Уменьшение размера частиц свидетельствует о том, что процессы деструкции детрита подстилок липняка ботанического сада происходят интенсивнее по сравнению с другими фитоценозами.
Рис. 3.
Фракционный состав детрита после листопада (А – сад, Б – парковая территория) и до листопада (В – сад, Г – парковая территория). Обозначения насаждений здесь и на рис. 6: 1 – березовые, 2 – липовые, 3 – кленовые.
Зольность. Установлено, что зольность для разных компонентов подстилки колеблется в широких пределах, а наибольшая зольность соответствует такому компоненту, как детрит (до 55%), что согласуется с литературными данными [19]. Как после листопада, так и конце периода реализации, зольность листьев и детрита подстилки для всех типов насаждений больше на парковой территории, чем в ботаническом саду, за исключением подстилки березы, где зольность этих компонентов существенно не различается.
О локальном увеличении зольности подстилок также свидетельствуют данные по средневзвешенной зольности (рис. 4). Подстилки ботанического сада до и после листопада характеризуются зольностью около 10–20%. Для подстилок парковых насаждений зольность до листопада составляет около 30%, а после листопада увеличивается до 55%. В отличие от ботанического сада, парковые насаждения испытывают влияние транспортных объектов, а увеличение зольности компонентов подстилки, вероятно, связано с осевшими на них частичками пыли от дорог и латерального поступления с открытых поверхностей. Аналогичное увеличение зольности листьев в городских условиях ранее было отмечено в литературе [13]. По мере разложения листьев осевшая пыль, по-видимому, концентрируется в детрите: зольность детрита подстилок парковых насаждений практически во всех случаях в 1.5 раза больше зольности детрита в ботаническом саду.
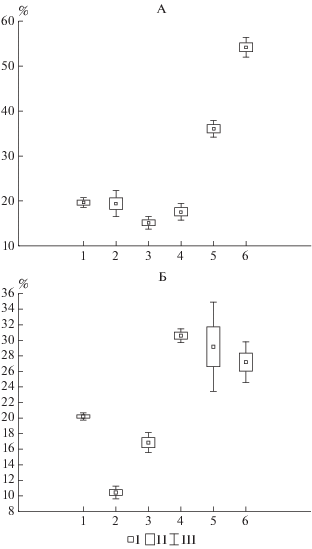
Повышенная зольность в сочетании со значительными запасами в подстилках широколиственных насаждений на парковой территории определяет то, что запасы зольных элементов в 1.5–2 раза превышают запасы золы в подстилках мелколиственного березняка. Запасы зольных элементов в подстилке после листопада довольно широко варьируют – от 50 до 270 г/м2, а в конце вегетационного периода уменьшаются в 2 раза. Наименьшие изменения в запасах зольных элементов до и после листопада наблюдаются в подстилке березняка, что определяется низкой зольностью.
Количество поступающих в почву зольных элементов за период реализации опада из подстилок под насаждениями березы, липы и клена разное и определяется скоростью их разложения. Более замедленный круговорот веществ наблюдается в подстилке березняка; количество поступающих зольных элементов составляет до 25 г/м2 за год. В условно эталонных насаждениях ботанического сада максимальным поступлением зольных элементов в почву характеризуются липняк – 65 г/м2 за год. Существенные различия между количеством поступающих зольных элементов из подстилок широколиственных пород в ботаническом саду и в парковых зеленых насаждениях, связано с повышенной зольностью компонентов в последних (рис. 5). Мелкие листья березы не так активно собирают пылевые частицы, как листья широколиственных пород, которые имеют значительную площадь листовой пластинки. Перечисленные особенности способствуют аккумуляции пыли на листьях липовых и кленовых насаждений с последующим их поступлением с опадом, что ранее отмечалось в литературе [29].
Рис. 5.
Зольность разных компонентов подстилки в насаждениях липы. Ботанический сад: А – после листопада, В – до листопада. Парковая территория: Б – после листопада, Г – до листопада; 1 – листья, 2 – ветви, 3 – детрит, 4 – семена, 5 – плоды, 6 – ветошь, 7 – кора.
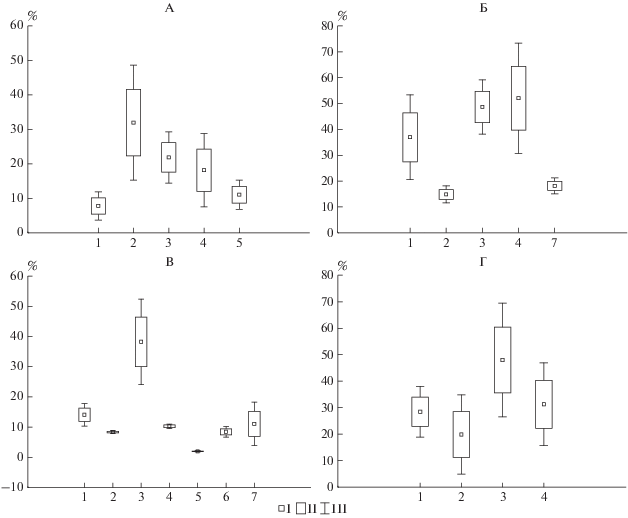
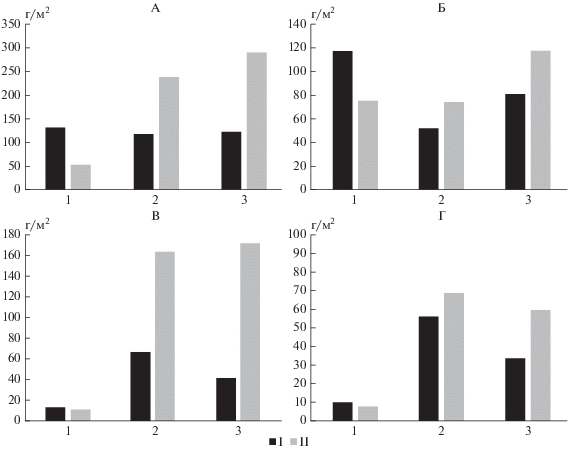
Закономерности распределения доли ежегодно поступающих из подстилки в почву зольных элементов от их исходного количества после листопада в целом соответствуют распределению общих запасов (рис. 6). Исключением являются подстилки кленовников, в которых доля поступающих в почву зольных элементов в условиях парковых насаждений превышает этот же показатель для ботанического сада почти вдвое. По-видимому, в кленовых насаждениях захват пылевых частиц компонентами опада наиболее интенсивен. Полученные данные показывают, что в городских условиях поступление в подстилки зольных элементов с пылью может в 4 раза превышать их поступление за счет естественных процессов реализации и составляет 97 и 130 г/м2 в год для липы и клена соответственно.
Рис. 6.
Запасы зольных элементов в подстилке после листопада (А), до листопада (Б), запасы (В) и доля (Г) зольных элементов, поступающих в почву. I – ботанический сад, II – парковая территория.
На основе проведенных комплексных исследований установлено, что роль видовой принадлежности деревьев-эдификаторов в формировании свойств подстилки и объеме поступающего в почву количества зольных элементов в городских насаждениях проявляется прежде всего в компонентном составе подстилок. Для березовых насаждений характерен менее интенсивный биологический круговорот, что адекватно отражается морфологией, запасами и компонентным составом подстилок, а также их сезонной динамикой. Для парковых насаждений в силу большей открытости пространства и повышенной подверженности ветровому воздействию происходит увеличение варьирования запасов подстилок и их отдельных фракций. Вместе с тем аэральные поступления пылевых частиц в парковых насаждениях увеличивают зольность ряда компонентов подстилок, особенно детрита и ветвей, что приводит к увеличению ежегодного поступления зольных элементов в почву и, как следствие, вовлечение их в круговорот – в большей степени это относится к широколиственным насаждениям. Функционирование подстилок отражается в изменении их характеристик в годовом цикле. Так, мощность может меняться в 1.5–2 раза, запасы подстилки и зольных элементов в подстилке – в 1.5–3 раза. Характеристики сезонной динамики детерминированы в большей степени видом дерева-эдификатора, нежели характером антропогенного воздействия. Ежегодное поступление зольных элементов из подстилки в почву в насаждениях широколиственных пород в 2–14 раз превышает их поступление при разложении годичного опада в подстилках березовых насаждений как сада, так и территории МГУ.
ВЫВОДЫ
1. Предложена и показана целесообразность оценки интенсивности круговорота на основе трехступенчатой системы исследования подстилок, включающей в себя определение типологии подстилок, запасов подстилки в послелистопадный период предыдущего года и долистопадный период следующего года, оценку структурной организации подстилок, подразумевающую соотношение легкоразлагаемых и устойчивых составляющих, а также компонентный и фракционный состав. Используется новое понятие – реализация подстилки – под которым понимается совокупность абиотических и биотических процессов преобразования подстилок, приводящих к изменению состава и свойств после листопада в годичном цикле функционирования.
2. Выявлено, что в городских экосистемах наблюдается интенсивный биологический круговорот веществ, о чем свидетельствует преобладание маломощных подстилок деструктивного типа и высокая доля ежегодно реализуемого органического вещества подстилок.
3. Среди исследованных насаждений подстилки мелколиственных березняков характеризуются минимальной скоростью биологического круговорота, что проявляется в относительном накоплении детрита и формировании ферментативного горизонта, а также минимальном количестве зольных элементов, высвобождающимся при реализации подстилок. Высокой скоростью биологического круговорота отличаются подстилки условно эталонных липовых насаждений ботанического сада, для которых определены максимальный объем и доля зольных элементов, поступающих в почву после периода реализации, что соответственно в 4–10 и 3–5 раз превышает соответствующие показатели подстилок березняка.
4. На парковой территории в насаждениях широколиственных пород установлено локальное увеличение зольности подстилок в целом и зольности таких их компонентов, как листья и детрит за счет антропогенного влияния.
5. Применение системы периодического ухода не приводит к существенному изменению фракционного состава и уменьшению запасов подстилки, так как в случае одногоризонтных деструктивных подстилок происходит их ежегодное восполнение за счет поступления опада. Однако изъятие части запасов подстилки при уборке и вывозе опада выводит значительное количество органического вещества из экосистем, уменьшая степень замкнутости круговорота в городских условиях.
6. Подстилки широколиственных насаждений менее устойчивы к применению системы ухода в сравнении с подстилками мелколиственных насаждений.
Список литературы
Аринушкина Е.В. Руководство по химическому анализу почв. М.: Изд-во Моск. ун-та, 1970. 392 с.
Базилевич Н.И., Титлянова А.А., Смирнов В.В. Методы изучения биологического круговорота в различных природных зонах. М.: Мысль, 1978.182 с.
Богатырев Л.Г. О классификации лесных подстилок // Почвоведение. 1990. № 3. С. 118–127.
Богатырев Л.Г. Является ли подстилка самостоятельным биогеоценотическим телом природы? // Экология. 1990. № 6. С. 3–7.
Богатырев Л.Г., Демин В.В., Матышак Г.В., Сапожникова В.А. О некоторых теоретических аспектах исследования лесных подстилок // Лесоведение. 2004. № 4. С. 17–29.
Гончарова О.Ю., Семенюк О.В., Матышак Г.В., Бобрик А.А. Сезонная динамика продукции СО2 почвами дендрария Ботанического сада МГУ им. М.В. Ломоносова // Вестник Моск. ун-та. Сер. 17. Почвоведение. 2016. № 2. С. 3–10.
Горчаковский А.Л., Андреяшкина К.Н. Изучение первичной продуктивности сообществ лесотундры на стационаре Харп // Ресурсы биосферы. 1975. Вып. 1. С. 25–33.
Гришина Л.А. Гумусообразование и гумусное состояние почв. М.: Изд-во Моск. ун-та, 1986. 244 с.
Злотин Р.И., Ходашова К.С. Роль животных в биологическом круговороте лесостепных экосистем. М.: Наука, 1974. 220 с.
Ильина Т.М., Сапожников А.П. Лесные подстилки как компонент лесного биогеоценоза // Вестник Красноярского гос. аграрного ун-та. 2007. № 5. С 45–47.
Ильяшенко М.А. Почвы и растительный покров структурно-функциональных компонентов парка музея-усадьбы “Архангельское”. Автореф. дис. … канд. биол. н. М., 2014. 164 с.
Карпачевский Л.О. Лес и лесные почвы М.: Лесная промышленность, 1981. С. 76–199.
Кавеленова Л.М., Здетоветский А.Г., Огневенко А.Я. К специфике содержания зольных веществ в листьях древесных растений в городской среде в условиях лесостепи (на примере Самары) // Химия растительного сырья. 2001. № 3. С. 85–90.
Классификация и диагностика почв России. Смоленск: Ойкумена, 2004.
Прокофьева Т.В., Герасимова М.И., Безуглова О.С. Введение почв и почвоподобных образований городских территорий в классификацию почв России // Почвоведение. 2014. № 10. С. 1155–1164.
Прокофьева Т.В., Розанова М.С., Попутников В.О. Некоторые особенности органического вещества почв на территориях парков и прилегающих жилых кварталов Москвы // Почвоведение. 2013. № 3. С. 302–315. https://doi.org/10.7868/S0032180X13030076
Родин Л.Е., Базилевич Н.И. Динамика органического вещества и биологический круговорот зольных элементов и азота в основных типах растительности Земного шара. М.: Наука, 1965. 264 с.
Сапожников А.П. Лесная подстилка – номенклатура, классификация, индексация // Почвоведение. 1984. № 5. С. 96–105.
Семенюк О.В., Богатырев Л.Г., Ваганова М.А. Характеристика подстилок парковых насаждений исторических ландшафтов на примере музея-усадьбы “Архангельское” // Бюл. Моск. о-ва испытателей природы. Отд. биол. 2017. Т. 122. Вып. 5. С. 37–49.
Семенюк О.В., Ваганова М.А. Характеристика растительного покрова как важнейшая составляющая комплексных почвенно-экологических исследований исторических парков // Бюл. Моск. о-ва испытателей природы. Отд. биол. 2016. Т. 121. Вып. 4. С. 32–42.
Семенюк О.В., Минин А.А. Экологические функции исторических природных территорий // Градостроительство. 2017. № 1(47). С. 63–68.
Смагин А.В., Азовцева Н.А., Смагина М.В., Степанов А.Л., Мягкова А.Д., Курбатова А.С. Некоторые критерии и методы оценки экологического состояния почв в связи с озеленением городских территорий // Почвоведение. 2006. № 5. С. 603–615.
Стома Г.В., Манучарова Н.А., Белокопытова Н.А. Биологическая активность микробных сообществ в почвах некоторых городов России // Почвоведение. 2004. № 6. С. 1–13.
Строганова М.Н., Мартыненко И.А., Прокофьева Т.В., Рахлеева А.А. Физико-химические и физико-механические свойства урбанизированных лесных почв // Лесные экосистемы и урбанизация. М., 2008. С. 90–124.
Строганова М.Н., Раппопорт А.В. Антропогенные почвы ботанических садов крупных городов южной тайги // Почвоведение. 2005. № 9. С. 1094–1101.
Телеснина В.М., Семенюк О.В., Богатырев Л.Г., Бенедиктова А.И. Особенности напочвенного покрова и лесных подстилок в искусственных липовых насаждениях в зависимости от характера ухода // Вестник Моск. ун-та. Сер. 17. Почвоведение. 2018. № 2. С. 3–11.
Телеснина В.М., Семенюк О.В., Богатырев Л.Г. Свойства лесных подстилок во взаимосвязи с напочвенным покровом в лесных экосистемах Подмосковья (на примере УОПЭЦ “Чашниково”) // Вестник Моск. ун-та. Сер. 17. Почвоведение. 2017. № 4. С. 11–20.
Чупина В.И. Антропогенные почвы ботанических садов (обзор) // Почвоведение. 2020. № 4. С. 495–506. https://doi.org/10.31857/S0032180X20040048
Экология города. М.: Научный мир, 2004. 624 с.
Abdullah M., AlHarun Y., Johnson J., Nazim M., Randall U., Robinson W. The effects of temperature on decomposition and allelopathic phytotoxicity of boneseed litter // J. Environ. Sci. 2015. V. 33. № 1. P. 1–11. https://doi.org/10.1016/j.jes.2014.12.017
Balla B.A., Christmanb M.P., Sharon S.J. Nutrient dynamics during photodegradation of plant litter in the Sonoran Desert // J. Arid Environ. 2019. V. 160. P. 1–10. https://doi.org/10.1016/j.jaridenv.2018.09.004
Bani A., Pioli S., Ventura M., Panzacchi P., Borruso L., Tognetti R., Tonon G., Brusetti L. The role of microbial community in the decomposition of leaf litter and deadwood // Applied Soil Ecology. 2018. V. 126. P. 75–84. https://doi.org/10.1016/j.apsoil.2018.02.017
Barrico L., Castro H., Pereira A., Coutinho M., Gonçalves M., Freitas H., Castro P. Plant and microbial biodiversity in urban forests and public gardens: Insights for cities’ sustainable development // Urban Forestry & Urban Greening. 2018. V. 29. P. 19–27. https://doi.org/10.1016/j.ufug.2017.10.012
Barry R., Parkinson D. Does repeated wetting and drying accelerate decay of leaf litter? // Soil Biol. Biochem. 1988. V. 20. № 5. P. 647–656. https://doi.org/10.1016/0038-0717(88)90149-6
Berg B., Söderström B. Fungal biomass and nitrogen in decomposing scots pine needle litter // Soil Biol. Bi-ochem. 1979. V. 11. № 4. P. 339–341. https://doi.org/10.1016/0038-0717(79)90045-2
Bogatyrev L., Berg B., Staaf H. Leaching of plant nutrients and total phenolic substances from some foliage litters – a laboratory study // Swedish Coniferous Forest Project Barrskogs landskapets Ekologi. Technical Report 33. Department of Ecology and Environmental Research, Swedish University of Agricultural Sciences. Upsala.1983. V. 33. P. 1–57.
Coq S., Nahmani J., Kazakou E., Fromin N., David J.F. Do litter-feeding macroarthropods disrupt cascading effects of land use on microbial decomposer activity? // Basic and Applied Ecology. 2020. V. 46. P. 24–34. https://doi.org/10.1016/j.baae.2020.03.004
Curiel J., Yuste D.D., Flores-Rentería D., García-Angulo D., Heres A.M., García-Angul C., Petritan M., Petritan I.C. Cascading effect associated with climate-change-induced conifer mortality in mountain temperate forests result in hot-spots of soil CO2 emissions // Soil Biol. Biochem. 2019. V. 133. P. 50–59. https://doi.org/10.1016/j.soilbio.2019.02.017
Das P.K., Nath S., Mykhopadhyay N., Banerjee S.K. Decomposition of litter and their effect on physicochemical and microbial properties of soil // Proc. Indian. Nat. Sci. Acad. B. 1993. V. 59. № 5. P. 517–524.
Galbraitha D.A., Rapleyb W.A. Museum Management and Curatorship Research at Canadian zoos and botanical gardens // Museum Management and Curatorship. 2005. V. 20. № 4. P. 313–331.
Hairiah K., Sulistyani H., Suprayogo D., Widianto L., Purnomosidh P., Widodo R.H., Van Noordwijk M. Litter layer residence time in forest and coffee agroforestry systems in Sumberjaya, West Lampung // Forest Ecology and Management. 2006. V. 224. № 1. P. 45–57.
Hassall M., Adl S., Berg M., Griffiths B., Scheu S. Soil fauna-microbe interactions: towards a conceptual framework for research // European J. Soil Biol. 2006. V. 42(1). P. 54–60.
Jackson P.W., Sutherland L.A. Encyclopedia of Biodiversity. Role of Botanic Gardens // Encyclopedia of Biodiversity. 2013. V. 6. P. 504–521.
Jenny H., Gessel S.P., Bingham F.T. Comparative study of decomposition rates of organic matter in temperate and tropical region // Soil Sci. 1949. V. 68. P. 419–432.
Kuznetsov V.A., Ryzhova I.M., Stoma G.V. Changes in the properties of soils of Moscow forest parks under the impact of high recreation loads // Eurasian Soil Science. 2017. V. 50. № 10. P. 1225–1235. https://doi.org/10.1134/S1064229317100052
Kuznetsov V.A., Ryzhova I.M., Stoma G.V. Transformation of forest ecosystems in Moscow megapolis under recreation impacts // Eurasian Soil Science. 2019. V. 52. № 5. P. 584–592. https://doi.org/10.1134/S1064229319050065
Matinyan N.N., Bakhmatova K.A., Korentsvit V.A. Soil of the Summer Garden (Saint Petersburg) // Eurasian Soil Science. 2017. V. 50. № 6. P. 637–645. https://doi.org/10.1134/S1064229317060060
Milano V., Maisto G., Baldantoni D., Bellino A., Bernard C., Croce A., Dubs F., Strumia S., Cortet S. The effect of urban park landscapes on soil Collembola diversity: A Mediterranean case study // Landscape and Urban Planning. 2018. V. 180. P. 135–147. https://doi.org/10.1016/j.landurbplan.2018.08.008
Otaki M., Tsuyuzaki S. Succession of litter-decomposing microbial organisms in deciduous birch and oak forests, northern Japan // Acta Oecologica. 2019. V. 101. 103485. https://doi.org/10.1016/j.actao.2019.103485
Semenyuk O.V., Telesnina V.M., Bogatyrev L.G., Benediktova A.I., Kuznetsova Ya.D. Assesment of intra-biogeocenotic variability of forest litters and dwarfshrub-herbaceous vegetation in spruce sand // Eurasian Soil Science. 2020. V. 53. № 1. P. 31–43. https://doi.org/10.1134/S1064229320010135
Shurong L., Ronggui H., Gaochao C. The role of UV-B radiation and precipitation on straw decomposition and topsoil C turnover // Soil Biol. Biochem. 2014. V. 77. P. 197–202. https://doi.org/10.1016/j.soilbio.2014.06.009
Sofo A., Mininni A.N., Ricciuti P. Comparing the effects of soil fauna on litter decomposition and organic matter turnover in sustainably and conventionally managed olive orchards // Geoderma. 2020. V. 372. 114393. https://doi.org/10.1016/j.geoderma.2020.114393
Titlyanova A.A. Universal nature of the processes of biotic turnover // Eurasian Soil Science. 2014. № 7. P. 631–640. https://doi.org/10.1134/S1064229314050226
Trifonova T.A., Zabelina O.N. Changes in the biological activity of heavy-metal- and oil-polluted soils in urban recreation territories // Eurasian Soil Science. 2017. № 4. P. 483–490. https://doi.org/10.1134/S1064229317040147
Wu F., Yang W., Zhang J. Litter decomposition in two subalpine forests during the freeze–thaw season // Acta Oecologica. 2010. V. 36. № 1. P. 135–140. https://doi.org/10.1016/j.actao.2009.11.002
Yan J., Wang L., Hu Y, Tsang Y. F., Zhang Y., Wu J, Fu X., Sun Y. Plant litter composition selects different soil microbial structures and in turn drives different litter decomposition pattern and soil carbon sequestration capability // Geoderma. 2018. V. 319(1). P. 194–203. https://doi.org/10.1016/j.geoderma.2018.01.009
Дополнительные материалы отсутствуют.