

Почвоведение, 2021, № 8, стр. 935-945
Выделение СО2 из образцов почв различных природных зон России при промерзании–оттаивании в лабораторных условиях
Д. В. Сапронов *
Институт физико-химических и биологических проблем почвоведения РАН, Федеральный исследовательский центр “Пущинский научный центр биологических исследований РАН”
142290 Московская область, Пущино, ул. Институтская, 2, Россия
* E-mail: sadmvas@gmail.com
Поступила в редакцию 17.12.2020
После доработки 01.03.2021
Принята к публикации 02.03.2021
Аннотация
Кратковременное промерзание поверхности почвы может происходить практически на всей территории России. Это явление стимулирует всплеск эмиссии почвенного СО2. В связи с этим в лабораторных условиях изучали динамику выделения СО2 из почв различных природных зон при периодически повторяющихся краткосрочных циклах промерзания–оттаивания. Исследовали образцы из верхних горизонтов следующих почв: криозема надмерзлотно-глееватого, глеезема криотурбированного, подбура оподзоленного, криозема грубогумусированного, подзола типичного, серой лесной, чернозема типичного, дерново-карбонатной, чернозема южного, каштановой, солонца, луговой слитизированной, бурой полупустынной. Установлено, что динамика выделения СО2 в течение циклов промерзания–оттаивания имела сходные черты для всех представленных почв. Достоверно зафиксировано выделение СО2 из мерзлых образцов. Оттаивание почвы приводило к импульсному увеличению скорости эмиссии СО2 независимо от типа почвы по сравнению с потоком углекислого газа до и после цикла промерзания–оттаивания. На широком ряде почв впервые показано, что значительный отклик эмиссии СО2 на промерзание–оттаивание поверхности почвы может происходить в любой природной зоне. Величина отклика зависит от свойств почв в первую очередь от содержания в ней органического вещества. Влияние единичных циклов промерзания–оттаивания почвы на годовой поток СО2 незначительно. Частые циклы промерзания–оттаивания могут оказывать существенное влияние на поток СО2 как в рамках отдельных сезонов, так и за год.
ВВЕДЕНИЕ
Заморозки, продолжительностью от нескольких часов до нескольких суток, ведущие к кратковременному промерзанию верхних 1–5 см почвы, могут происходить практически на всей территории России. В регионах с умеренным климатом циклы промерзания–оттаивания поверхности почвы обычно происходят в течение осеннего и весеннего периодов, а в арктической и субарктической зонах эти процессы вероятны на протяжении всего вегетационного сезона [35, 37]. Климатические изменения, ведущие к повышению температур в течение холодного сезона, увеличению продолжительности бесснежного периода, а также уменьшению мощности снежного покрова в зимний период, способствуют увеличению количества циклов промерзания–оттаивания [2, 9, 41]. Территорий, на которых эти процессы возможны, становится больше.
В ряде лабораторных исследований, а также in situ, показано, что краткосрочное промерзание–оттаивание почвы стимулирует импульсное увеличение почвенной эмиссии СО2 [12, 13, 16, 21, 26, 29, 35, 37]. Кроме того, циклы промерзания–оттаивания ускоряют выделение других парниковых газов, особенно закиси азота [22, 23, 28, 30, 31, 39]. В то же время имеются данные, что промерзание–оттаивание почвы не изменяет скорость выделения CO2 [42].
При оттаивании почвы происходит повышение ее температуры и, как следствие, увеличиваются физиологическая активность почвенных организмов, ферментативная и дыхательная активность, ускоряется деструкция субстратов. [15, 24, 27]. Однако считается, что промерзание–оттаивание не является фактором, значительно влияющим на выделение углерода из почвы. Это утверждение базируется на том, что подавляющее количество CO2 выделяется в теплый период за счет дыхания корней и почвенных микроорганизмов [14]. В то же время Коксон и Паркинсон [11] показали, что величина зимнего потока увеличивалась на 11% при учете эмиссии во время циклов промерзания–оттаиваниям. Следовательно, циклы промерзания–оттаивания влияют на годовой поток СО2 и могут менять соотношение между теплым и холодным периодами, а также между сезонами года. Степень влияния всплесков эмиссии, вызванных промерзанием–оттаиванием почвы на годовой поток СО2 зависит от периодичности полевых измерений скорости эмиссии СО2, методики определения годового потока, а также климатических особенностей конкретного региона и почвенных свойств.
Вышесказанное важно для России, поскольку связано с огромным разнообразием почв и климатических условий на ее территории. Многие российские почвы обладают большими запасами активного (потенциально-минерализуемого) органического вещества [6], что предопределяет их высокий эмиссионный потенциал [3]. Кроме того, значительная часть территории России характеризуется холодным климатом. В связи с этим циклы промерзания–оттаивания необходимо учитывать при расчетах почвенной эмиссии углекислого газа и составлении углеродного баланса, как отдельных экосистем, так территории в целом. Предположительно, этот вклад будет иметь особое значение в зонах тундры и тайги, для которых характерно неоднократное промерзание–оттаивание поверхности почвы в теплый период.
Цель работы – в лабораторных условиях изучить закономерности эмиссии СО2 из почв различных природных зон России в ходе краткосрочных, периодически повторяющихся циклов промерзания–оттаивания и получить удельные величины эмиссии для исследуемого ряда почв. Основная гипотеза исследования состояла в том, что почвы различных природных зон будут по-разному реагировать на промерзание–оттаивание.
ОБЪЕКТЫ И МЕТОДЫ
Исследования проводили в лабораторных условиях. Объектами послужили образцы верхних горизонтов почв различных природных зон (табл. 1). Тундровая зона представлена криоземом надмерзлотно-глееватым (Reductaquic Turbic Cryosol (Loamic)), глееземом криотурбированным (Reductaquic Turbic Cryosol (Loamic)) и подбуром оподзоленным (Spodic Turbic Cryosol (Arenic)). В подзоне северной тайги отобрали криозем грубогумусированный (Histic Cryosol (Glacic)), в подзоне южной тайги – подзол иллювиально-гумусовый (Albic Rustic Podzol (Arenic)), в зоне широколиственных лесов – серую лесную почву (Haplic Luvisol (Loamic, Aric)). Лесостепная подзона представлена черноземом типичным (Haplic Chernozem (Loamic, Pachic)) и дерново-карбонатной почвой (рендзиной) (Rendzic Leptosol (Humic)), степная – черноземом южным (Haplic Chernozem (Siltic)), светло-каштановой почвой (Haplic Kastanozem (Siltic)), а также солонцом луговым (Gypsic Solonetz (Loamic, Columnic)) и луговой гипсовой cлитизированной почвой (Pellic Gypsic Protosalic Vertisol (Hypereutric)). В полупустыне изучали бурую полупустынную почву (Haplic Calcisol (Loamic)).
Таблица 1.
Объекты исследования
| Тип почвы | Природная зона, растительные ассоциации | Место отбора, географические координаты |
|---|---|---|
| Глеезем криотурбированный Reductaquic Turbic Cryosol (Loamic) |
Тундра, мохово-осоково-кустарничковая ассоциация | Нижнеколымский район Республика Саха (Якутия) 69°25′25.8″ N, 161°46′57.6″ E |
| Криозем надмерзлотно-глееватый Reductaquic Turbic Cryosol (Loamic) |
Тундра, кустарничково-моховая и кочкарно-пушицевая ассоциации | Там же; 69°22′01.7″ N, 161°48′58.3″ E |
| Подбур оподзоленный Spodic Turbic Cryosol (Arenic) |
Тундра, кустарничково-лишайниковая ассоциация | Там же; 68°49′55.8″ N, 161°01′30″ E |
| Криозем грубогумусированный Histic Cryosol (Glacic) |
Северная тайга, лиственничное редколесье | Билибинский район, Чукотский автономный округ, обнажение Молотковский Камень 68°12′33.2″ N, 161°58′04.5″ E |
| Подзол иллювиально-гумусовый Albic Rustic Podzol (Arenic) |
Южная тайга, чернично-зеленомошный елово-сосновый лес с примесью осины и березы | Дмитровский район, Московская обл. 56°30′20.0″ N, 37°39′20.9″ E |
| Серая лесная почва Haplic Luvisol (Loamic, Aric) |
Широколиственные леса, липово-кленово-осиновый лес | Московская область, Пущино, 54°50′06.5″ N, 37°34′21.8″ E |
| Чернозем типичный Haplic Chernozem (Loamic, Pachic) |
Лесостепь, разнотравно-злаковая степь | Воронежская область, Каменная степь 51°01′44.2″ N, 40°43′38.5″ E |
| Дерново-карбонатная почва (Рендзина) Rendzic Leptosol (Humic) |
Лесостепь, “Докучаевская” лесополоса | Воронежская область, Дивногорье, 50°58′21.4″ N, 39°18′13.2″ E |
| Чернозем южный Haplic Chernozem (Siltic) |
Степь, разнотравно-типчаково-ковыльная | Волгоградская область, Родниковский, 50°29′01.3″ N, 42°36′33.2″ E |
| Солонец луговой Gypsic Solonetz (Loamic, Columnic) |
Степь, полынно-злаковая | Волгоградская область, 50°28′54.6″ N, 42°36′14.0″ E |
| Луговая гипсовая cлитизированная почва Pellic Gypsic Protosalic Vertisol (Hypereutric) |
Степь, разнотравно-злаково-осоковая | Волгоградская область, Новоаннинский район, Волгоградская область, Веселый, 50°30′20.8″ N, 42°34′43.6″ E |
| Светло-каштановая почва Haplic Kastanozem (Siltic) |
Степь, ковыльно-типчаковая | Волгоградская область, Качалино, 49°06′12.9″ N, 44°03′05.8″ E |
| Бурая полупустынная почва Haplic Calcisol (Loamic) |
Полупустыня, эфемерово-полынная |
Астраханская область, Ахтубинск 48°16′01.2″ N, 46°27′29.4″ E |
Смешанные образцы отбирали из верхних горизонтов почв методом конверта. Из подзола брали два образца: из слоя 0–10 см – подстилочно-торфяный горизонт (О) и 10–15 см – органо-минеральный, перегнойный (АО). Для остальных почв глубина отбора составила 0–10 см. Почву, высушенную до воздушно-сухого состояния при +20°С, просеивали через сито (3 мм) и отбирали крупные растительные остатки. Характеристика объектов исследования представлена в табл. 2. Определения физико-химических характеристик выполняли в Центре коллективного пользования ИФХ и БПП РАН с применением стандартных методик [1]. Содержание общего углерода и азота в образцах определяли сжиганием навески в кислороде при температуре >1000°C на анализаторе LECO CHNS-932. рН определяли потенциометрически, для оценки обменной кислотности использовали титрование солевой вытяжки щелочью по Соколову (соотношение почва : 1 M KСl 1 : 2.5). SO4 гипса извлекали 0.2 M НСl и определяли гравиметрическим методом. Для определения СО2 карбонатов применяли ацидометрический метод, суммы обменных оснований расcчитывали по Каппену–Гильковицу. Грануломет-рический состав определяли методом пипетки с обработкой пирофосфатом натрия.
Таблица 2.
Характеристика почвенных образцов (глубина отбора 0–10 см)
| Тип почвы | N, % | рНKCl/${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$ | Ʃ обменных оснований | Обменная кислотность | CaCO3 | SO4 гипса | Сухой остаток | <0.01 мм |
|---|---|---|---|---|---|---|---|---|
| смоль(экв)/кг | % | |||||||
| Глеезем криотурбированный | 0.11 | 4.5/5.7 | 14.38 | 0.32 | 0 | 0 | 0.04 | 31.0 |
| Криозем надмерзлотно-глееватый | 0.18 | 3.8/5.0 | 12.13 | 1.70 | 0 | 0 | 0.07 | 24.2 |
| Подбур оподзоленный | 0.20 | 3.5/4.9 | 2.63 | 6.34 | 0 | 0 | 0.05 | 13.2 |
| Криозем грубогумусированный | 0.10 | 4.8/5.9 | 20.38 | 0.10 | 0 | 0 | 0.06 | 38.0 |
| Подзол иллювиально-гумусовый, гор. О | 2.81 | 2.7/3.9 | 3.75 | 8.60 | 0 | 0 | 0.19 | 8.2 |
| Подзол иллювиально-гумусовый, гор. АО | 0.16 | 2.7/3.8 | 2.85 | 3.24 | 0 | 0 | 0.03 | 7.3 |
| Серая лесная почва | 0.22 | 5.1/5.7 | 18.63 | 0.10 | 0 | 0 | 0.12 | 23.9 |
| Чернозем типичный | 0.43 | 6.9/7.6 | 53.13 | 0.10 | 5.7 | 0.01 | 0.15 | 37.2 |
| Дерново-карбонатная почва |
1.02 | 7.5/8.0 | 53.88 | 0.10 | 54.1 | 0.05 | 0.13 | 14.0 |
| Чернозем южный | 0.13 | 6.2/7.2 | 25.88 | 0.06 | 2.0 | 0.01 | 0.05 | 31.8 |
| Солонец луговой | 0.18 | 7.1/8.7 | 39.13 | 0.00 | 3.6 | 0.03 | 0.28 | 46.6 |
| Луговая гипсовая cлитизированная почва | 0.46 | 7.4/7.9 | 54.13 | 0.00 | 7.0 | 4.02 | 2.04 | 50.8 |
| Светло-каштановая почва | 0.17 | 6.2/7.4 | 25.63 | 0.06 | 2.0 | 0.01 | 0.06 | 32.6 |
| Бурая полупустынная почва | 0.37 | 7.3/8.2 | 53.88 | 0.00 | 10.0 | 0.01 | 0.13 | 36.6 |
Динамику выделения СО2 при промерзании–оттаивании почвы оценивали в эксперименте общей продолжительностью 59 сут. Для инкубации почвенных образцов использовали стеклянные флаконы объемом 100 см3. Масса навески сухой почвы составляла 10 г. Образцы инкубировали при постоянной влажности (80% от предельной полевой влагоемкости) в автоматическом термостате с охлаждением Panasonic MIR-254. В ходе эксперимента последовательно проводили два цикла промерзания–оттаивания. Все почвенные варианты инкубировали в четырехкратной повторности.
В начале эксперимента проводили предварительную инкубацию почв, в течение которой температуру ступенчато понижали. Сначала флаконы выдерживали 14 дней при температуре +21°С, затем 10 дней при +10°С и 7 дней при +5°С. Цикл промерзания–оттаивания длился две недели. Замораживание образцов проводили при –5°С в течение 7 дней. Затем температуру повышали до +5°С (период оттаивания) и выдерживали при этой температуре еще 7 дней. Повторный цикл промерзания–оттаивания проводили по той же схеме.
Интенсивность выделения углекислого газа измеряли с различными интервалами в зависимости от температурного режима. Между измерениями флаконы были закрыты полиэтиленовой пленкой, что позволяло избежать быстрого пересыхания образцов и формирования во флаконе анаэробных условий за счет свободного газообмена. Перед каждым измерением концентрации СО2 с флаконов снимали пленку, проветривали и герметично закрывали резиновыми мембранами для накопления СО2. Время накопления углекислого газа длилось от нескольких часов до нескольких суток в зависимости от скорости его выделения. Концентрацию СО2 в газовых пробах определяли на газовом хроматографе “Кристалл люкс-4000М”. Скорость выделения СО2 из почвы рассчитывали по формуле:
Расчеты проводили в программе Microsoft Excel. Для статистического анализа применяли программу Statistica. Рассчитывали средние величины, стандартные отклонения (STD) и стандартные ошибки (SE) измерений эмиссии. При анализе использовали группировку данных в соответствии с разными периодами эксперимента.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В начале инкубации регистрировали интенсивное выделение углекислого газа, которое варьировало от 0.83 до 15.9 мкг C–CО2/(г ч). Наименьшее и наибольшее значения отмечены в вариантах с горизонтами АО и О подзола соответственно. Остальные варианты стартовали в диапазоне от 1 до 5 мкг C–CО2/(г ч). Начальный уровень эмиссии СО2 был обусловлен откликом микроорганизмов на увлажнение образцов. По мере формирования равновесного состояния во флаконах интенсивность выделения СО2 уменьшалась. Коэффициент зависимости уровня эмиссии СО2 от температуры во время циклов промерзания–оттаивания изменялся в широких пределах, но оставался значимым для всех почв, кроме дерново-карбонатной почвы и слитозема (R2 от 0.19 до 0.68; P < 0.05).
Снижение температуры уменьшало диапазон варьирования и нивелировало различия дыхания исследуемых почв. Эмиссия СО2 в период предварительной инкубации при температуре +5°С менялась от 0.01 до 0.16 мкг С/(г ч), в среднем составляя 0.06 мкг С/(г ч). В варианте с торфяным горизонтом подзола эмиссия СО2 была на 1–2 порядка больше, чем в вариантах с органо-минеральными горизонтами других почв и достигала 1.26 мкг С/(г ч) (рис. 1). Вероятно, это связано с различным содержанием органического вещества в образцах [4]. Торфяный горизонт подзола характеризовался наибольшими значениями скорости выделения СО2 при всех температурных режимах.
Рис. 1.
Динамика выделения СО2 из таежного подзола во время циклов промерзания–оттаивания: 1 – горизонт О, 2 – горизонт АО, ±STD.
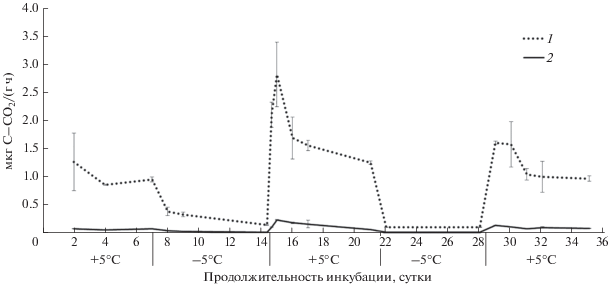
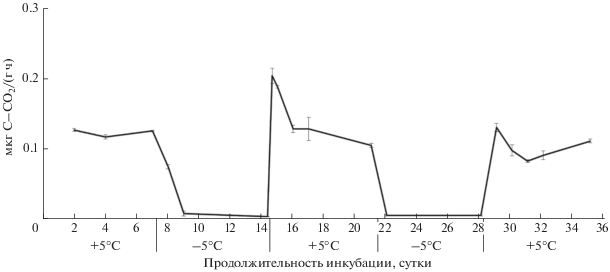
Промерзание почвенных образцов из всех представленных природных зон приводило к уменьшению интенсивности выделения СО2 до значений, близких к 0 (рис. 1–6). Как правило, такое снижение эмиссии происходило за 1–2 дня. В ряде случаев в период промерзания образцов фиксировали отрицательные величины потока СО2. Это может свидетельствовать о низкой продукции СО2, затруднении газообмена между почвой и атмосферой и, вероятно, о поглощении СО2 почвенными компонентами. Такая динамика эмиссии СО2 была характерна для почвенных образцов, отобранных в лесостепной и степной зонах (рис. 4, 5). Эти почвы характеризуются присутствием карбоната кальция и, как следствие, нейтральным или щелочным рН. В таких условиях, возможно, происходит образование гидрокарбоната кальция (Ca(HCO3)2) и поглощение СО2 в результате хемосорбции. Тем не менее, бурая полупустынная почва, также содержащая карбонаты, не продемонстрировала при промерзании снижения уровня эмиссии углекислого газа (рис. 6). Проведенный дисперсионный анализ выявил недостоверное различие между суммарными величинами потоков СО2 за оба цикла промерзания–оттаивания в связи с наличием или отсутствием карбонатов в почвах.
Рис. 2.
Динамика выделения СО2 из различных тундровых почв во время циклов промерзания–оттаивания: 1 – подбур оподзоленный, 2 – криозем надмерзлотно-глееватый, 3 – глеезем криотурбированный, ±STD.
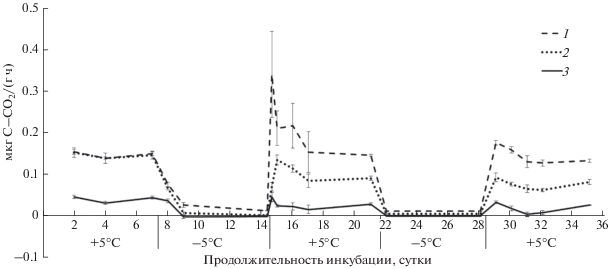
Рис. 3.
Динамика выделения СО2 из почв лесной зоны во время циклов промерзания–оттаивания: 1 – криозем грубогумусированный, 2 – подзол (горизонт АО), 3 – серая лесная почва, ±STD.
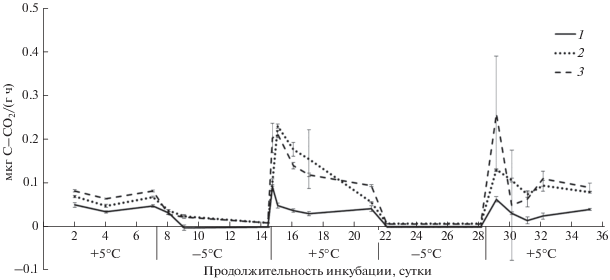
Рис. 4.
Динамика выделения СО2 из почв лесостепной зоны во время циклов промерзания–оттаивания: 1 – чернозем типичный, 2 – дерново-карбонатная почва, ±STD.
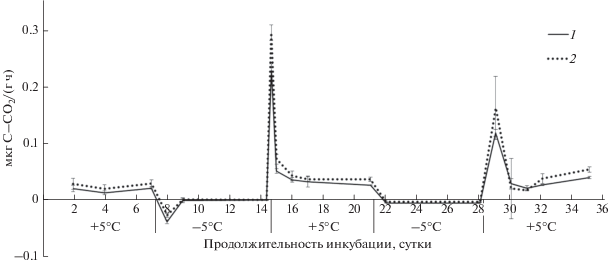
Рис. 5.
Динамика выделения СО2 из почв степной зоны в ходе циклов промерзания–оттаивания: 1 – чернозем южный, 2 – светло-каштановая почва, 3 – солонец, 4 – луговая слитизированная, ±STD.
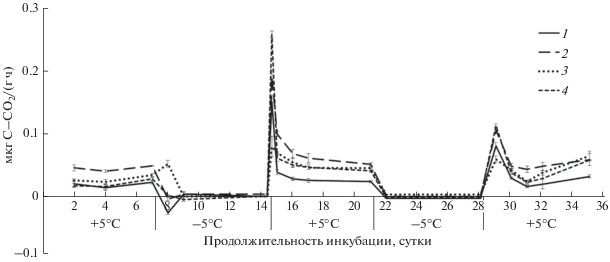
Рис. 6.
Динамика выделения СО2 из бурой полупустынной почвы в течение циклов промерзания–оттаивания, ±STD.
Некоторые исследователи при изучении циклов промерзания–оттаивания почвы с началом промерзания почвенных образцов наблюдали увеличение скорости выделения СО2, что по их мнению связано с дегазацией почвенного раствора [5, 21]. Показано, что при промерзании–оттаивании почвы поглощение и выделение СО2 зависит от рН почвенного раствора [4]. В нашей работе только в варианте с солонцом (рис. 5) при промерзании почвенного образца и переходе температуры от +5 до –5°С отмечено значимое увеличение выделения углекислого газа. При повторном промерзании солонца такой эффект не наблюдался.
В ходе эксперимента, при отрицательных температурах фиксировалось выделение СО2. Полученные результаты подтверждают то, что выделение СО2 из промерзших почв возможно, но находится на низком уровне. Это хорошо согласуется с результатами исследований в естественных условиях, свидетельствующих о существовании зимних потоков СО2 из почв [18, 20]. Низкая интенсивность выделения углекислого газа при отрицательных температурах обусловлена затрудненной диффузией вследствие высокой влажности и монолитного замерзания образцов [21]. Известно, что микроорганизмы в почве остаются активными при температуре грунта до –7°С [16, 20, 25, 27, 39]. Следовательно, при температуре ‒5°С, которая обеспечивает промерзание поверхностного горизонта, микроорганизмы способны потреблять почвенное органическое вещество и продуцировать СО2. Замечено, что при температурах, близких к 0°C, микроорганизмы смещают потребление с детрита на растворенные соединения, в основном микробного происхождения [36]. Во время оттаивания микробный субстрат, богатый азотом, постепенно замещается бедным азотом детритом [36]. Также выявлено, что в результате промерзания происходит физическая деградация почвенных агрегатов и органических молекул, увеличивая доступность органического вещества для потребления микроорганизмами [10, 38].
Предыдущими исследованиями установлено, что промерзание и последующее оттаивание почвы стимулирует резкое увеличение скорости выделения СО2 по сравнению с потоком СО2 до и после цикла промерзания–оттаивания [12, 21, 35]. В нашем исследовании проведено два цикла промерзания–оттаивания один за другим. При каждом оттаивании всех исследуемых почв зарегистрировано импульсное увеличение скорости выделения СО2. Увеличение скорости эмиссии СО2 происходило одновременно с началом оттаивания почвы, достигая пика в тот же, а иногда на следующий день, что хорошо согласуется с результатами Кургановой и Лопес де Гереню [4]. Усиленное выделение углекислого газа длилось 1–2 дня. В течение первого цикла промерзания–оттаивания максимальная скорость выделения СО2 варьировала в зависимости от типа почвы от 0.05 до 0.34 мкг С/(г ч), превысив средние значения скорости выделения СО2 при температуре +5°С в 1.4–3.3 раза.
Скорость выделения СО2 во время первого цикла промерзания–оттаивания была примерно в 1.5–2 раза больше, чем во время второго. Исключение составила серая лесная почва, в которой оба всплеска эмиссии были сопоставимы (рис. 3). Суммарная эмиссия СО2 во время второго периода оттаивания почв была в среднем на 14% меньше, чем в первый период (парный t-тест, n = 13, p = 0.003). Считается, что при промерзании часть микроорганизмов гибнет, а при оттаивании происходит лизис погибших клеток, и их биомасса обеспечивает усиление выделения СО2 [30, 35, 37]. При повторных циклах промерзания–оттаивания количество активных микроорганизмов уменьшается, как и их воздействие на органическое вещество почвы [21, 35]. В результате расходования доступного микроорганизмам субстрата эффект импульсного увеличения выделения СО2 от промерзания–оттаивания снижается [7]. Наши результаты укладываются в вышеизложенную теорию, которая в первую очередь охватывает лабораторные исследования с образцами небольшой массы. В то же время есть работы, показывающие, что промерзание и последующее оттаивание почв может не влиять на функционирование и структуру микробоценоза почвы [19].
При сравнении реакции исследуемых почвенных образцов на промерзание–оттаивание установлено, что динамика выделения СО2 имеет общие черты для всех исследованных типов почв, хотя различается количественно. Проведенный дисперсионный анализ не выявил достоверных различий величин всплесков, обусловленных зональностью или типовой принадлежностью почв. Полученные результаты свидетельствуют о том, что, по-видимому, в любой природной зоне может происходить усиление выделения СО2 из почвы в результате процессов промерзания–оттаивания.
Интенсивность выделения СО2 и других парниковых газов при оттаивании почвы зависит от ряда факторов, например, от влажности почвы [8, 32], ее температуры [17], глубины активного слоя [33] и др. По результатам корреляционного анализа установлена значимая отрицательная связь величины отклика с рН (R2 = –0.50, P < 0.05), с суммой обменных оснований (R2 = –0.44, P < 0.05) и положительная связь с обменной кислотностью (R2 = 0.75, P < 0.05). Отмечена достоверная отрицательная связь с содержанием глинистой фракции (R2 = –0.51, P < 0.05), следовательно легкий гранулометрический состав также положительно сказывается на увеличении отклика эмиссии.
Наибольший прирост эмиссии при оттаивании исследуемых образцов, как правило, отмечался у почв с большим содержанием органического вещества. Тем не менее корреляционный анализ не выявил достоверной связи между величиной отклика и содержанием углерода в образцах. Существует мнение, что различная величина эмиссионного отклика при промерзании–оттаивании почв обусловлена неодинаковой доступностью, количеством и характером органического субстрата [34]. Примером может служить различная реакция двух горизонтов подзола с разным содержанием, и возможно, с разной доступностью органического вещества в них (рис. 1).
Доступность для почвенных микроорганизмов содержащегося в почве органического вещества может быть оценена отношением эмиссии C–CО2 к запасам углерода органического вещества в почве (C–CО2/Сорг) (табл. 3). Видно, что некоторые почвы, имея большие запасы углерода, характеризуются относительно небольшой продукцией СО2. Это свидетельствует о стабильности и малодоступности их органического вещества. Все эти свойства почвы прямо или косвенно влияют на почвенные микроорганизмы. Вероятно, интенсивность всплеска эмиссии зависит не только от количества углерода в почве, но также определяется величиной и структурой микробного сообщества [35, 40].
Таблица 3.
Содержание углерода в почвах и его суммарная эмиссия
| Тип почвы | Собщ, % | C–CО2, мкг/г почвы |
мкг C–CО2/г Собщ | |
|---|---|---|---|---|
| за 2 цикла | за 2 цикла | за весь эксперимент | ||
| Глеезем криотурбированный | 1.2 | 9 | 679 | 14.3 |
| Криозем надмерзлотно-глееватый | 3 | 31 | 995 | 15.4 |
| Подбур оподзоленный | 2.8 | 57 | 1983 | 18.6 |
| Криозем грубогумусированный | 1.3 | 14 | 1032 | 15.9 |
| Подзол, гор. О | 40.5 | 480 | 1163 | 9.8 |
| Подзол, гор. АО | 4 | 36 | 872 | 6.0 |
| Серая лесная | 2.6 | 41 | 1532 | 12.8 |
| Чернозем типичный | 5.1 | 12 | 248 | 4.0 |
| Дерново-карбонатная | 15 | 16 | 114 | 1.8 |
| Чернозем южный | 1.5 | 10 | 679 | 8.2 |
| Солонец луговой | 1.9 | 18 | 884 | 13.9 |
| Луговая cлитизированная | 5.1 | 17 | 335 | 6.9 |
| Светло-каштановая | 1.9 | 21 | 1087 | 12.7 |
| Бурая полупустынная | 4.1 | 40 | 923 | 12.9 |
Таким образом, величина усиления эмиссии СО2 в результате промерзания определяется не природной зоной, а свойствами самих почв. При этом решающую роль играет содержание и доступность органического вещества в почве. Обобщая результаты нашего и других лабораторных исследований, можно заключить, что циклы промерзания–оттаивания почв безусловно влияют на скорость выделения из них СО2. При этом отдельные циклы промерзания–оттаивания незначительно влияют на величину годовой эмиссии СО2 из почвы. Тем не менее значительное количество повторяющихся циклов промерзания–оттаивания может существенно отразиться на эмиссионном потоке СО2 за год, особенно в ходе отдельных сезонов и в пределах определенных природных зон.
Для более полного понимания влияния циклов промерзания–оттаивания почвы на баланс углерода в экосистемах необходимы детальные исследования в естественных условиях, которые позволят установить количество таких циклов и реальные величины эмиссии СО2 в природе. Лабораторные исследования проводят с небольшими образцами почвенного материала, не содержащими живых корней растений, что ограничивает оценку влияния этих циклов только на микроорганизмы. В лабораторных экспериментах не учитывается, что краткосрочное промерзание–оттаивание затрагивает лишь верхние 1–2 см почвы, а СО2 продуцирует вся почвенная толща. Образцы малого объема в отличие от почвенной толщи промерзают полностью. В связи с этим остается неясным вклад термодиффузии и других важных физических процессов. Очевидно, что механизмы, стоящие за резким приростом эмиссии СО2 при оттаивании мерзлой поверхности почвы, намного сложнее, и не могут быть сведены лишь к гибели микробных клеток и их лизису.
ЗАКЛЮЧЕНИЕ
Характер эмиссии углекислого газа во время циклов промерзания–оттаивания демонстрирует общие черты для всех представленных зональных типов почв. В проведенном эксперименте зарегистрирован поток СО2 из почв, находящихся в мерзлом состоянии. Оттаивание почв способствовало резкому непродолжительному увеличению выделения СО2. Следовательно, промерзание–оттаивание одинаково влияет на различные типы почв, и значимый эмиссионный отклик может наблюдаться в любой природной зоне. Различия между величинами почвенной эмиссии СО2 даже в пределах одной зоны обусловлены физико-химическими свойствами почв, в первую очередь разным содержанием и доступностью органического вещества. Так как в эксперименте использовали почвенные образцы без корневых систем растений, можно утверждать, что отклик микробного сообщества почвы на краткосрочное промерзание–оттаивание будет одинаков, несмотря на типовую принадлежность почвы. Установлено, что влияние единичных циклов промерзания–оттаивания почвы на годовой поток СО2 незначительно. Напротив, частые циклы промерзания–оттаивания могут оказывать существенное влияние на годовую эмиссию СО2.
Список литературы
Воробьева Л.А. Теория и практика химического анализа почв. М.: ГЕОС, 2006. 400 с.
Гулев С.К., Катцов В.М., Соломина О.Н. Глобальное потепление климата продолжается // Вестн. PAH. 2008. Т. 78. № 1. С. 20–27.
Пулы и потоки углерода в наземных экосистемах России / Под ред. Г.А. Заварзина. М.: Наука, 2007. 315 с.
Курганова И.Н., Лопес де Гереню В.О. Вклад абиотических факторов в усиление эмиссии СО2 из почв при процессах замораживания–оттаивания // Почвоведение. 2015. № 9. С. 1145–1152. https://doi.org/10.7868/S0032180X15090099
Лопес де Гереню В.О., Курганова И.Н., Типе Р., Лофтфильд Н. Влияние процессов замораживания–оттаивания на эмиссию парниковых газов из пахотной буроземной почвы // Агрохимия. 2004. № 2. С. 23–30.
Семенов В.М., Когут Б.М., Зинякова Н.Б., Масютенко Н.П., Малюкова Л.С., Лебедева Т.Н., Тулина А.С. Биологически активное органическое вещество в почвах европейской части России // Почвоведение. 2018. № 4. С. 457–472.
Семенов В.М., Когут Б.М., Лукин С.М. Влияние повторяющихся циклов высушивания–увлажнения–замораживания–оттаивания на активный пул органического вещества почвы // Почвоведение. 2014. № 4. С. 443–454.
Blanc-Betes E., Welker J.M., Sturchio N.C., Chanton J.P., Gonzalez-Meler M.A. Increases in winter precipitation and snow accumulation transform Arctic tundra from a sink to a source of methane // Global Change Biol. 2016. V. 22. P. 2818–2833. https://doi.org/10.1111/gcb.13242
Campbell J.L., Ollinger S.V., Flerchinger G.N., Wicklein H., Hayhoe K., Bailey A.S. Past and projected future changes in snowpack and soil frost at the Hubbard Brook Experimental Forest, New Hampshire, USA // Hydrological Processes. 2010. V. 24. P. 2465–2480.
Christensen S., Christensen B.T. Organic matter available for denitrification in different soil fractions: effect of freeze/thaw cycles and straw disposal // J. Soil Sci. 1991. V. 42. P. 637–647.
Coxson D.S., Parkinson D. Winter respiratory activity in aspen woodland forest floor litter and soils // Soil Biol. Biochem. 1987. V. 19. P. 49–59.
DeLuca T.H., Keeney D.R., Mc Carty G.W. Effect of freeze-thaw events on mineralization of soil nitrogen // Biol. Ferti. Soils. 1992. V. 14. P. 116–120.
Feng X., Nielsen L.L., Simpson M. Responses of soil organic matter and microorganisms to freeze – thaw cycles // Soil Biol. Biochem. 2007. V. 39. P. 2027–2037.
Groffman P.M., Hardy J.P., Driscoll C.T., Fahey T.J. Snow depth, soil freezing, and fluxes of carbon dioxide, nitrous oxide and methane in a northern hardwood forest // Global Change Biol. 2006. V. 12. P. 1748–1760.
Grogan P., Michelsen A., Ambus P., Jonasson S. Freeze-thaw regime effects on carbon and nitrogen dynamics in sub-Arctic heath tundra mesocosms // Soil Biol. Biochem. 2004. V. 36. P. 641–654.
Herrmann A., Witter E. Sources of C and N contributing to the flush in mineralization upon freeze-thaw cycles in soils // Soil Biol. Biochem. 2002. V. 34. P. 1495–1505.
Hicks Pries C.E., Schuur E.A.G., Crummer K.G. Thawing permafrost increases old soil and autotrophic respiration in tundra: Partitioning ecosystem respiration using δ13C and Δ14C // Global Change Biol. 2013. V. 19. P. 649–661.
Holst J., Liu C., Yao Z., Brüggemann N., Zheng X., Giese M., Butterbach-Bahl K. Fluxes of nitrous oxide, methane and carbon dioxide during freezing–thawing cycles in an Inner Mongolian steppe // Plant Soil. 2008. V. 308. P. 105–117.
Koponen H.T., Jaakkola T., Keinanen-Toivola M.M., Kaipainen S., Tuomainen J., Servomaa K. et al. Microbial communities, biomass, and activities in soils as affected by freeze thaw cycles // Soil Biol. Biochem. 2006. V. 38. P. 1861–1871.
Kurganova I., Lopes de Gerenyu V., Rozanova L., Sapronov D., Myakshina T., Kudeyarov V. Annual and seasonal CO2 fluxes from Russian southern taiga soils // Tellus. 2003. V. 55B. P. 338–344.
Kurganova I., Teepe R., Loftfield N. Influence of freeze thaw events on carbon dioxide emission from soil at different moisture and land use // Carbon Balance and Management. 2007. V. 2. P. 1–9.
Kurganova I.N., Lopes de Gerenyu V.O., Khoroshaev D.A., Myakshina T.N., Sapronov D.V., Zhmurin V.A., Kudeyarov V.N. Analysis of the Long-Term Soil Respiration Dynamics in the Forest and Meadow Cenoses of the Prioksko-Terrasny Biosphere Reserve in the Perspective of Current Climate Trends // Euras. Soil Sci. 2020. V. 53. № 10. P. 1421–1437. https://doi.org/10.1134/S1064229320100117
Kurganova I.N., Lopes de Gerenyu V.O. Effect of the Temperature and Moisture on the N2O Emission from Some Arable Soils // Euras. Soil Sci. 2010. V. 43. № 8. P. 919–928. https://doi.org/10.1134/S1064229310080090
Kurganova I.N., Rozanova L.N., Myakshina T.N., Kudeyarov V.N. Monitoring of CO2 emission from soils of different ecosystems in Southern part of Moscow region: data base analyses of long-term field observations // Euras. Soil Sci. 2004. V. 37. Sup. 1. P. 74–78.
Kurganova I.N., Teepe R. The effects of freezing-thawing processes on soil respiration activity // Euras. Soil Sci. 2003. V. 36. P. 976–985.
Larsen K.S., Jonasson S., Michelsen A. Repeated freeze-thaw cycles and their effect on biological processes in two arctic ecosystem types // Appl. Soil Ecol. 2002. V. 21. P. 187–195.
Lopes de Gerenyu V.O., Kurganova I.N., Rozanova L.N., Kudeyarov V.N. Annual emission of carbon dioxide from soils of the Southern Taiga soils of Russia // Euras. Soil Sci. 2001. V. 34. P. 931–944.
Lopes de Gerenyu V.O., Kurganova I.N., Teepe R., Loftfild N., Flessa H. Effect of Contrasting Changes in Hydrothermic Conditions on the N2O Emission from Forest and Tundra Soils // Euras. Soil Sci. 2007. V. 40. № 7. P. 795–799. https://doi.org/10.1134/S1064229307070137
Ludwig B., Teepe R., Lopes de Gerenyu V., Flessa H. N2O and CO2 emissions from gleyic soils in the Russian tundra and a German forest during freeze thaw periods – a microcosm study // Soil Biol. Biochem. 2006. V. 38. P. 3815–3819.
Matzner E., Borken W. Do freeze-thaw events enhance C and N losses from soils of different ecosystems? A review // Eur. J. Soil Sci. 2008. V. 59. P. 274–284.
Morkved P.T., Dörsch P., Henriksen T.M., Bakken L.R. N2O emissions and product ratios of nitrification and denitrification as affected by freezing and thawing // Soil Biol. Biochem. 2006. V. 38. P. 3411–3420.
Natali S.M., Schuur E.A.G., Mauritz M., Schade J.D., Celis G., Crummer K.G., Johnston C., Krapek J., Pegoraro E., Salmon V.G., Webb E. Permafrost thaw and soil moisture driving CO2 and methane release from upland tundra // J. Geophys. Res.: Biogeosci. 2015. V. 120. P. 525–537. https://doi.org/10.1002/2014JG002872
O’Donnell J.A., Harden J.W., McGuire A.D., Kanevskiy M.Z., Joregenson M.T., Xu X. The effect of fire and permafrost interactions on soil carbon accumulation in an upland black spruce ecosystem of interior Alaska: Implications for post-thaw carbon loss // Global Change Biol. 2011. V. 17. P. 1461–1474. https://doi.org/10.1111/j.1365-2486.2010.02358.x
Priemé A., Christensen S. Natural perturbations, drying–wetting and freezing–thawing cycles, and the emission of nitrous oxide, carbon dioxide and methane from farmed organic soils // Soil Biol. Biochem. 2001. V. 33. P. 2083–2091.
Schimel J.P., Clein J.S. Microbial response to freeze-thaw cycles in tundra and taiga soils // Soil Biol. Biochem. 1996. V. 28. P. 1061–1066.
Schimel J.P., Mikan C. Changing microbial substrate use in Arctic tundra soils through a freeze–thaw cycle // Soil Biol. Biochem. 2005. V. 37. P. 1411–1418.
Skogland T., Lomeland S., Goksoyr J. Respiratory burst after freezing and thawing of soil: Experiments with soil bacteria // Soil Biol. Biochem. 1988. V. 20. P. 851–856.
Soulides D.A., Allison F.E. Effects of drying and freezing soils on carbon dioxide production, available mineral nutrients, aggregation, and bacterial population // Soil Sci. 1961. V. 91. P. 291–298.
Teepe R., Brumme R., Beese F. Nitrous oxide emission from soil during freezing and thawing periods // Soil Biol. Biochem. 2001. V. 33. P. 1269–1275.
Treat C.C., Wollheim W.M., Varner R.K., Grandy A.S., Talbot J., Frolking S. Temperature and peat type control CO2 and CH4 production in Alaskan permafrost peats // Global Change Biol. 2014. V. 20. P. 2674–2686. https://doi.org/10.1111/gcb.12572
Venäläinen A., Tuomenvirta H., Heikinheimo M., Kellomaki S., Peltola H., Strandman H., Vaisanen H. Impact of climate change on soil frost under snow cover in a forested landscape // Climate Res. 2001. V. 17. P. 63–72.
Wang F.L., Bettany J.R. Influence of freeze-thaw and flooding on the loss of soluble organic carbon and carbon-dioxide from soil // J. Environ. Quality. 1993. № 22. P. 709–714.
Дополнительные материалы отсутствуют.