

Почвоведение, 2023, № 11, стр. 1418-1436
Изменение микробиологических и физико-химических свойств почв после пожаров в сосновых и березовых лесах центральных районов Забайкальского края
А. В. Богородская a, *, Е. А. Кукавская a, О. П. Каленская b, Л. В. Буряк a, b, c
a Институт леса им. В.Н. Сукачева ФИЦ КНЦ СО РАН
660036 Красноярск, Академгородок, 50/28, Россия
b Сибирский государственный университет науки и технологий им. М.Ф. Решетнева
660049 Красноярск, пр. Мира, 82, Россия
c Филиал ФБУ ВНИИЛМ “Центр лесной пирологии”
660062 Красноярск, ул. Крупской, 42, Россия
* E-mail: anbog@ksc.krasn.ru
Поступила в редакцию 14.02.2023
После доработки 19.06.2023
Принята к публикации 25.06.2023
- EDN: YIJSXP
- DOI: 10.31857/S0032180X23600245
Аннотация
Исследованы микробиологические и некоторые физико-химические свойства дерново-подбуров иллювиально-железистых (Entic Rustic Podzols) сосновых и серогумусовых типичных легкосуглинистых почв (Umbrisols) производных березовых лесов центральных районов Забайкальского края. После пожаров в сосновых насаждениях в дерново-подбурах отмечается уменьшение содержания суммы обменных оснований, валового азота, подвижных соединений калия и фосфора, увеличение соотношения C : N; для березняков характерно увеличение названных показателей и сужение соотношения C : N в профиле серогумусовых типичных почв. Содержание гумуса в верхнем почвенном горизонте снижается только на свежей гари в сосняке после высокоинтенсивного пожара, тогда как на других участках оно увеличилось. Уменьшение кислотности почв отмечено на всех пройденных пожарами участках. Высокоинтенсивные пожары приводят к достоверному снижению содержания микробной биомассы и увеличению базального дыхания, а также к изменению структуры эколого-трофических групп микроорганизмов в профиле почв до глубины 10 см минерального горизонта. Низкоинтенсивные пожары затрагивают в основном подстилку. Коэффициент qCO2 увеличивался в 2–5 раз после пожаров в подстилке и в 1.5–2 раза в гумусовом горизонте только после высокоинтенсивных пожаров. На свежих гарях в сосняках значительно сокращались запас микробной биомассы и микробная продукция CO2 до глубины 10 см минеральной части почвенных профилей. На остепненном участке, сформировавшемся после воздействия пожаров в сосняке, и в березняке после пожара высокой интенсивности в гумусовом горизонте запас углерода микробной биомассы сокращался на 15–20%, а микробная продукция СО2 увеличивалась на 10–20%. Рассматриваемая послепожарная трансформация структурно-функциональных параметров микробоценоза почв и снижение на 20–40% суммарных запасов углерода микробной биомассы на всех участках предопределяют длительный период восстановления почв после пожаров в светлохвойных и лиственных насаждениях центральных районов Забайкальского края.
ВВЕДЕНИЕ
Лесные экосистемы Забайкальского края характеризуются одним из самых высоких показателей горимости на территории России. Согласно данным дистанционного зондирования, за период с 1996 по 2015 гг. ежегодная площадь лесных пожаров в регионе варьировала от 0.04 до 5.6 млн га. При этом наибольшая горимость наблюдается в центральных и южных районах края [49]. Отмечено, что древостои Забайкалья подвергаются регулярному воздействию пожаров [19]. В последнее время для лесных земель Забайкальского края характерны повторные пожары с периодичностью ниже полного цикла развития коренных фитоценозов, что приводит к трансформации лесных экосистем в нелесные [40, 49].
Географическое положение Забайкальского края, суровые климатические условия, горный рельеф с выраженной поясностью и разнообразие растительности определяют многообразие почв, которые характеризуются различной мощностью, степенью развития, плодородием. Многими исследователями отмечается, что пирогенный фактор является ключевым в современной динамике лесов и почв региона [9, 19, 22, 37].
Интерес к изучению послепожарной трансформации микробоценозов почв определяется их главенствующей ролью в циклах биогенных элементов и продукционно-деструкционном звене почвенного органического вещества [36, 42, 51]. Непосредственное воздействие пожара на почвенную биоту связано с пиковыми температурами, развивающимися при горении, их продолжительностью, а также со свойствами почвы и содержанием в ней воды [42, 52]. Косвенные эффекты пожара часто превосходят прямые. Пожар изменяет такие свойства почв, как кислотность, количество и качество органического вещества, доступность питательных веществ и удержание влаги, которые определяют скорость восстановления микробных популяций [42, 51, 53]. Влияние пожаров на почвенные микроорганизмы сохраняется в течение десятилетий, а интенсивность пожара является основным предиктором длительных нарушений в микробном сообществе почв [43, 45, 46, 54].
Ранее исследовалось влияние пожаров на агрохимические свойства почв и микробоценозов в зависимости от характеристик пожара и давности пирогенного воздействия в насаждениях европейской части России [12, 15, 18, 20, 25, 26, 30, 35], Средней Сибири [5, 6, 8, 16, 27, 31, 50], Дальнего Востока [17, 22, 37]. Для территории Забайкалья имеются немногочисленные данные по микробиологической активности почв степных и луговых фитоценозов [10, 24]. Комплексные исследования изменений свойств почв и микробоценозов после пожаров разной интенсивности в лесах Забайкальского края ранее не проводились.
Цель работы – оценка влияния пожаров разной интенсивности на микробиологические и некоторые физико-химические свойства почв сосновых и березовых лесов центральных районов Забайкальского края.
ОБЪЕКТЫ И МЕТОДЫ
Почвенно-микробиологические исследования проводили в сосновых и березовых насаждениях в Читинском и Карымском районах Забайкальского края. В исследованных районах доминируют горные морфоструктуры. Климат – резко континентальный, среднегодовая температура составляет –0.5…–6°C. Среднегодовое количество осадков варьирует от 250–300 мм в лесостепной зоне до 500–700 мм на хребтах [13]. Для региона характерно малое количество снега зимой, раннее снеготаяние весной и жаркое лето [14]. Вследствие этого складывается высокая пожарная опасность по погодным условиям, особенно в первую половину пожароопасных сезонов [9].
В ходе полевых исследований в 2016 г. было заложено 6 пробных площадей (ПП) в сосновых (51°54′ N, 113°18′ E) и производных березовых (51°16′ N, 113°41′ E) насаждениях, пройденных пожарами различной интенсивности и находящихся на различных стадиях послепожарной сукцессии, а также в контрольных к ним длительно не горевших насаждениях (табл. 1). Исследуемые насаждения до пирогенного воздействия представлены чистыми сосновыми (Pinus sylvestris L., ПП 1–4) и березовыми (Betula pubescens Ehrh., ПП 5–6) высокополнотными древостоями III класса бонитета (рис. 1). Пройденные пожарами участки расположены в одинаковых контрольными лесорастительных условиях, а древостои изначально имели схожие лесоводственно-таксационные характеристики.
Таблица 1.
Лесоводственно-таксационное описание насаждений и характеристика пожаров на пробных площадях
| № пробной площади | ВНУМ* – местоположение, экспозиция и крутизна склона |
Характеристика пожара | Характеристика насаждения | ||||
|---|---|---|---|---|---|---|---|
| период после пожара, лет | вид, сила | тип леса или категория участка | полнота | проективное покрытие и преобладающие виды живого напочвенного покрова | мощность напочвенного покрова (опад и подстилка), см | ||
| Сосняки | |||||||
| ПП 1, контроль | 694 м, ровное | – | – | Сосняк рододендроновый | 0.8 | 55% – Iris uniflora, Lathyrus humilis, Pulsatilla orientali-sibirica, Carex sp., Sanguisorba officinalis, Trifolium lupinaster, Agrimonia pilosa, Astragalus adsurgens, Artemisia tanacetifolia, Potentilla fragarioides, Equisetum sylvaticum, Fragaria orientalis | 6.0 |
| ПП 2 | 723 м, ЮЗ 10° | 3 | Низовой, от слабой до средней | Сосняк мертвопокровный | 0.6 | 15% – Artemisia tanacetifolia, Astragalus adsurgens, Sanguisorba officinalis, Iris uniflora 10% – Dicranum sp. |
1.2 |
| ПП 3 | 735 м, ровное | 1 | Верховой, сильный | Гарь мертвопокровная | 0 | 5% – Artemisia tanacetifolia, Iris uniflora, Trifolium lupinaster | 0.2 |
| ПП 4 | 712 м, СВ 12° | 16 8 3 |
Верховой,
сильный Низовой, сильный Низовой, сильный |
Гарь разнотравно-вейниковая, остепненный участок | 0 | 80% – Calamagrostis epigeios, Artemisia tanacetifolia, A. integrifolia, A. commutate, Pulsatilla orientali-sibirica, Sanguisorba officinalis, Trifolium lupinaster, Astragalus adsurgens, Chamaenerion angustifolium, Polygonatum odoratum, Oxytropis grandiflora, Dendranthema zawadskii | 0.1 |
| Березняки | |||||||
| ПП 5, контроль | 717 м, С 3° | – | – | Березняк разнотравно-вейниковый | 0.8 | 40% – Calamagrostis epigeios, Equisetum sylvaticum, Sanguisorba officinalis, Pyrola rotundifolia, Adenophora tricuspidata, Rubus saxatilis, Lathyrus humilis, Galium boreale, Filipendula ulmaria | 10.5 |
| ПП 6 | 757 м, ровное | 1 | Низовой, сильный | Гарь разнотравно-вейниковая | 0 | 100% – Calamagrostis epigeios, Galium boreale, Chamaenerion angustifolium, Thalictrum minus, Astragalus adsurgens, Filipendula ulmaria, Vicia sepium, Aquilegia sibirica, Veronica longifolia, Geranium sibiricum | 0.2 |
Рис. 1.
Исследуемые участки лесных земель в сосновых (а–d) и березовых (e–f) насаждениях: ПП 1 (а), ПП 2 (b), ПП 3 (c), ПП 4 (d), ПП 5 (e), ПП 6 (f).

Сосновые насаждения представлены чистыми группово-разновозрастными древостоями, включающими 2 поколения. Возраст старшего поколения древостоев составил 100–120 лет, средний диаметр – 26 см, средняя высота – 24 м. Возраст младшего элемента леса 30–40 лет, средний диаметр – 13 см, средняя высота – 14 м. Березовые насаждения – спелые одновозрастные (60–70 лет), средний диаметр – 24 см, средняя высота – 20 м. В сосняках изучены контрольное не горевшее насаждение (ПП 1) и насаждение, пройденное 3 года назад низовым пожаром от слабой до средней силы с сохранением древесного яруса (ПП 2), а также насаждение, пройденное 1 год назад верховым сильным пожаром с полной гибелью деревьев (ПП 3). Обследована разнотравно-вейниковая гарь, в настоящее время представляющая собой остепненный участок лесных земель (ПП 4). Данная гарь образовалась после воздействия верхового пожара на сосновый древостой 16 лет назад. Погибший древостой на гари был вырублен, а вырубленный участок гари два раза (8 и 3 года назад до проведения исследований) был пройден повторными сильными низовыми пожарами. В производных березняках, сформировавшихся на месте коренных сосновых насаждений, исследовано контрольное длительно не горевшее насаждение (ПП 5) и насаждение, пройденное 1 год назад сильным низовым пожаром с полной гибелью древостоя (ПП 6). Описание пробных площадей проводили с использованием общепринятых в практике лесоведения, лесной таксации, геоботаники и пирологии методик [3, 23, 33].
Не горевшее сосновое насаждение (ПП 1) представлено рододендроновым типом леса, пройденное низовым пожаром 3 года назад сосновое насаждение (ПП 2) и свежая гарь, пройденная верховым сильным пожаром 1 год назад, (ПП 3) – мертвопокровным, а остепненный участок вырубленной и неоднократно горевшей гари (ПП 4) – разнотравно-вейниковым типом. В не горевшем насаждении (ПП 1) подлесок средней густоты представлен рододендроном даурским (Rhododendron dauricum). Степень проективного покрытия живым напочвенным покровом составляет 50–60%. В составе травяного покрова преобладает разнотравье, в котором встречаются осока (Carex sp.), полынь пижмолистная (Artemisia tanacetifolia) и хвощ лесной (Equisetum sylvaticum). В насаждении, пройденном огнем (ПП 2), подлесок редкий, он представлен рододендроном даурским. Степень проективного покрытия травяным покровом составляет 10–20%, в его составе встречаются астрагал приподнимающийся (Astragalus adsurgens), кровохлебка лекарственная (Sanguisorba officinalis), касатик одноцветковый (Iris uniflora), на сильно горевших участках напочвенного покрова отмечается разрастание послепожарного мха – дикранума (Dicranum sp.). На свежей гари, пройденной верховым пожаром 1 год назад, (ПП 3) травяной покров только начал отрастать и его проективное покрытие не превышало 5%.
На остепненном участке (ПП 4) отмечается разрастание трав, степень их проективного покрытия составляет более 80%, а в составе преобладает (до 50%) вейник наземный (Calamagrostis epigeios), значительна доля иван-чая узколистного (Chamaenerion angustifolium), встречаются несколько видов полыней (Artemisia sp.), дендрантема Завадского (Dendranthema zawadskii), клевер люпинолистный (Trifolium lupinaster).
Длительно не горевшее березовое насаждение (ПП 5) и участок гари (ПП 6) представлены разнотравно-вейниковым типом леса и гари. В подлеске встречаются спирея средняя (Spiraea media), береза кустарниковая (Betula fruticosa), ива Бебба (Salix bebbiana), курильский чай (Dasiphora fruticosa). Степень проективного покрытия живого напочвенного покрова в насаждении (ПП 5) составляет от 30 до 50%, в его составе преобладает вейник наземный. Также на участке распространены другие представители разнотравья: хвощ лесной, грушанка круглолистная (Pyrola rotundifolia), лабазник вязолистный (Filipendula ulmaria). Под пологом березового насаждения доминирует 1.2 тыс. шт./га подрост сосны (9С1Б). Данное обстоятельство и присутствие угля в почвенном горизонте указывают, что березняк сформировался на гари, образовавшейся после высокоинтенсивного пожара в сосновом насаждении. На гари (ПП 6) степень проективного покрытия травяного покрова достигает 100%, травяной покров двухъярусный, а в его составе отмечаются иван-чай узколистный, горошек заборный (Vicia sepium), встречаются вероника длиннолистная (Veronica longifolia), водосбор сибирский (Aquilegia sibirica) и другие виды разнотравья.
На пробных площадях в 2016 г. закладывали почвенные разрезы для определения генетической принадлежности почв и отбирали репрезентативные почвенные образцы по генетическим горизонтам для проведения агрохимического [11] и микробиологического [29] анализов. Почвенные исследования проводили через 1–3 года после последнего пожара, когда пирогенные изменения свойств почв проявляются наиболее выражено [5, 7, 22, 25]. Для оценки динамики микробиологических параметров дополнительно провели исследования в 2019 г.
При отборе почвенных образцов определяли влажность почвы на момент взятия образцов (термовесовым методом), температуры почвенных слоев (портативным термометром Checktemp) и плотности при помощи бура Качинского. Физико-химические свойства почв исследовали согласно [4, 11].
Численность и структуру эколого-трофических групп микроорганизмов (ЭКТГМ) изучали по соотношению количества колониеобразующих единиц (КОЕ) в 1 г абсолютно-сухой почвы, выросших на разных диагностических средах [29]. Гидролитические микроорганизмы учитывали на мясопептонном агаре (МПА), копиотрофы – на крахмало-аммиачном агаре (КАА), олиготрофы – на почвенном агаре (ПА). Рассчитывали коэффициент микробиологической минерализации (Kмин = КАА : МПА), иллюстрирующий разложение и накопление органических веществ, и коэффициент олиготрофности (Kолиг = ПА : МПА), показывающий способность микроорганизмов к аккумуляции питательных элементов из “рассеянного” состояния [28].
Для изучения экофизиологических параметров функциональной активности определяли содержание микробной биомассы (Смик) методом субстрат-индуцированного дыхания [1, 38] и базальное дыхание микроорганизмов (БД). Подробное описание методики и рабочих параметров хроматографа описано ранее [6]. Рассчитывали микробный метаболический коэффициент qСО2 (мкг С–СО2/(мг Смик ч)), иллюстрирующий удельное дыхание микробной биомассы и характеризующий экологический статус микробного сообщества почвы [39].
Запасы углерода микробной биомассы Смик (г С/м3) и микробное продуцирование СО2 (мг С–СО2/(м3 ч) в исследуемых горизонтах почв рассчитывали как произведение содержания Смик (г/г почвы) или интенсивности БД (мг/г почвы), плотности почвы (ρ, г/см3) и объема почвы в данном слое (V, м3). Суммарные запасы Смик и общее микробное продуцирование СО2 в профиле почв получали суммированием полученных данных для каждого исследуемого горизонта [32, 34].
Все микробиологические анализы выполняли в трех повторностях и рассчитывали стандартное отклонение от среднего. Корреляционную связь между ЭКТГМ, Смик, БД и физико-химическими свойствами почвы оценивали коэффициентами (r) Пирсона или Спирмена при доверительной вероятности 95%.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Послепожарная трансформация физико-химических свойств почв. Установлено, что исследованные почвы сосновых насаждений относятся к отделу альфегумусовых и представлены дерново-подбурами иллювиально-железистыми (Entic Rustic Podzols) [21, 48]. Профиль типа: O–AY–{AY + + BF}–BF–BС–C. Органогенный горизонт (О) вниз по профилю почв сменяется темно-бурым, либо буро-коричневым гумусово-аккумулятивным горизонтом AY, мощность которого составляет около 5 см. Далее располагается переходный горизонт, состоящий из части горизонта AY и иллювиального горизонта BF, имеющий буроватые или буровато-охристые тона окраски. Ниже горизонт BF растянут и переходит в почвообразующую породу. Для минеральной части профиля характерно наличие примазок окислов железа. Для насаждений, ранее подвергавшихся пожарам, в почвенных горизонтах отмечаются включения угля.
В березовых насаждениях почвы относятся к отделу органо-аккумулятивных и диагностируются как серогумусовые типичные легкосуглинистые на бескарбонатных породах (Umbrisols) [21, 48]. Профиль типа: O–AY–С1–C2. Морфологический профиль состоит из лесной подстилки (О–4 см), ниже сменяется маломощным серогумусовым горизонтом (AY, 4–5 см), сразу переходящим к материнской породе.
Почвы имеют различный гранулометрический состав (супесчаный и песчаный – в сосновых насаждениях и легкосуглинистый – в березовых) и степень увлажнения (в сосняках почвы сухие, в березняках – свежие).
В сосновом длительно не горевшем контрольном насаждении в составе мелкозема преобладают фракции среднего и мелкого песка, доля которых составляет 46%. Содержание средней и мелкой пыли незначительно. После пожаров содержание среднего и мелкого песка увеличивается до 52%. Доля крупного песка также возрастает до 32%. Доля илистой фракции в ненарушенном сосняке составляет 15% и увеличивается с глубиной. После воздействия пожаров доля ила снижается по мере увеличения интенсивности пожара – до 10%.
Проведенный анализ показал, что почва в длительно не горевшем березовом насаждении отличается меньшей кислотностью и большим содержанием гумуса, по сравнению с почвой контрольного соснового насаждения (табл. 2). Отмечено, что пирогенная трансформация свойств почв выражается в усилении минерализации органического вещества, резком увеличении количества водорастворимых соединений щелочных и щелочноземельных элементов, что снижает кислотность почв. В то же время изменения агрохимических показателей происходят не только в органогенном, но и в минеральных горизонтах почв, что обусловлено поступлением в них легкорастворимых щелочных компонентов золы вместе с осадками [22, 37, 41].
Таблица 2.
Некоторые физико-химические показатели почв исследуемых участков
| Горизонт, глубина, см | рН H2O | Обменные катионы Са2+ + Mg2+, смоль(экв)/кг | Валовые % | С/N | Подвижные, мг/кг | Содержание фракций, %; размер частиц, мм | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| гумус | N | N–NH4 | К2О | Р2О5 | <0.001 | <0.01 | ||||
| ПП 1. Сосняк рододендроновый | ||||||||||
| AY, 2–7 | 5.4 | 18.0 | 3.0 | 0.21 | 8.3 | 6.8 | 181.0 | 56.8 | 10.2 | 6.4 |
| {AY + BF}, | 5.5 | 8.0 | 1.5 | 0.08 | 10.9 | 4.6 | 81.8 | 70.8 | 14.3 | 2.9 |
| 7–12 BF, 12–26 |
6.0 | 7.5 | 0.8 | 0.04 | 11.6 | 1.8 | 73.6 | 81.9 | – | – |
| ПП 2. Сосняк мертвопокровный | ||||||||||
| AY, 1–2 | 5.6 | 7.2 | 3.4 | 0.17 | 11.6 | 7.7 | 130.0 | 92.1 | 0.4 | 5.7 |
| {AY + BF}, | 5.9 | 4.0 | 1.4 | 0.06 | 13.5 | 3.1 | 77.0 | 33.3 | 10.7 | 3.6 |
| 2–5 BF, 5–26 |
5.2 | 4.0 | 0.6 | 0.04 | 8.7 | 2.0 | 62.2 | 68.9 | – | – |
| ПП 3. Гарь мертвопокровная | ||||||||||
| AY, 0–5 | 6.0 | 4.0 | 1.3 | 0.06 | 12.6 | 10.4 | 121.0 | 22.0 | 11.9 | 2.8 |
| {AY + BF}, | 6.0 | 3.5 | 1.5 | 0.05 | 17.4 | 2.90 | 74.6 | 20.1 | 8.8 | 0.3 |
| 5–11 BF, 11–26 |
7.0 | 3.0 | 0.6 | 0.04 | 8.7 | 1.6 | 66.4 | 35.0 | – | – |
| ПП 4. Гарь разнотравно-вейниковая | ||||||||||
| AY, 0–5 | 6.3 | 11.5 | 3.2 | 0.08 | 23.2 | 2.7 | 216.0 | 85.0 | 8.0 | 1.1 |
| {AY + BF}, | 6.2 | 7.0 | 1.2 | 0.05 | 13.9 | 1.7 | 119.0 | 26.9 | 11.1 | 1.1 |
| 5–11 BF, 11–26 |
6.2 | 7.0 | 0.3 | 0.04 | 4.4 | 1.8 | 94.9 | 38.2 | – | – |
| ПП 5. Березняк разнотравно-вейниковый | ||||||||||
| AY, 4–8 | 5.6 | 7.7 | 5.6 | 0.14 | 23.2 | 3.7 | 64.5 | 54.0 | – | – |
| С1, 8–13 | 5.7 | 8.0 | 0.6 | 0.06 | 5.8 | 2.5 | 45.3 | 29.0 | – | – |
| С2, 13–26 | 5.8 | 7.5 | 0.5 | 0.06 | 4.8 | 3.1 | 44.8 | 37.0 | – | – |
| ПП 6. Гарь разнотравно-вейниковая | ||||||||||
| AY, 0–5 | 6.5 | 32.6 | 5.8 | 0.23 | 14.6 | 2.7 | 149.0 | 71.0 | – | – |
| С1, 5–12 | 6.9 | 20.4 | 0.8 | 0.05 | 9.3 | 1.6 | 66.3 | 70.0 | – | – |
| C2, 12–26 | 6.4 | 9.3 | 0.5 | 0.03 | 9.7 | 2.2 | 72.7 | 43.0 | – | – |
Выявлено, что в профиле почв насаждений, пройденных огнем, отмечены изменения актуальной кислотности почвенных горизонтов, которая варьировала в зависимости от интенсивности пожара и характеристик послепожарной растительности. Значения рН водной вытяжки в гумусовом горизонте на всех участках лесных земель, пройденных пожарами, изменились в сторону нейтральной. Максимальные изменения отмечены в горизонте AY остепненного участка повторно горевшей гари разнотравно-вейниковой (с 5.4 на контроле – до 6.3 на участке гари) и в пройденном сильным низовым пожаром березняке (с 5.6 до 6.5). После пожара слабой силы, в сосновом насаждении (ПП 2) кислотность почв снижалась в меньшей степени. Такая закономерность обусловлена тем, что на уменьшение кислотности почв оказывает воздействие не только подщелачивание почвы за счет вмывания золы, но и разрастание на гарях травяного покрова, способствующего образованию гуминовых кислот. В связи с этим большее снижение кислотности верхнего горизонта почв характерно для остепненного участка гари в сосняке (ПП 4) и гари в березняке (ПП 6), где проективное покрытие травяным покровом превышает 80%, а запасы трав увеличились соответственно в 4.5 и 2.7 раз относительно допожарных значений. Вниз по почвенному профилю значения рН, как правило, увеличиваются.
Содержание гумуса в сосновых насаждениях, в том числе пройденных пожарами, закономерно ниже, чем в березняках (табл. 2). В длительно не горевшем сосняке рододендроновом на ПП 1 в горизонте AY дерново-подбура содержание гумуса составляет 3%, а максимальное его значение (3.4%) отмечено в сосновом насаждении, пройденном низовым пожаром слабой силы (ПП 2). В березовом насаждении на ПП 5 содержание гумуса в верхнем горизонте серогумусовой типичной почвы составляет 5.6%, а после воздействия пожаров отмечается увеличение содержания гумуса до 5.8%. Это обусловлено разрастанием на участке гари густого 2-ярусного травяного покрова, запасы которого достигали 170 г/м2, превышая контроль в 2.7 раз. Разрастание травяного покрова способствовало более активному дерновому процессу почвообразования и накоплению гуминовых кислот. Минимальное содержание гумуса в верхнем горизонте почвы (1.3%) отмечено на свежей гари соснового насаждения после воздействия верхового пожара (ПП 3). Это вызвано полным выгоранием напочвенного покрова (опада и подстилки), а также водной и ветровой эрозией обнаженной легкой песчаной почвы в год воздействия пожара. Через несколько лет после воздействия пожаров, в сосняках содержание гумуса в верхнем горизонте почвы достигает допожарных показателей и даже несколько превышает их. Так, на участке остепненной разнотравно-вейниковой гари (ПП 4) содержание гумуса в горизонте AY дерново-подбура через 3 года после воздействия последнего пожара составляет 3.2%. Увеличение содержания гумуса на гарях обусловлено разрастанием травяного покрова, прежде всего злаков, в первую очередь – вейника наземного.
Сумма обменных оснований в сосновых насаждениях после воздействия пожара слабой силы во всех горизонтах профиля дерново-подбура (ПП 2) снизилась в 2 раза по сравнению с контрольным показателем (ПП 1), а после верхового высокоинтенсивного пожара – в 4.5 раза в горизонте AY и в 2.5 раза в нижележащих горизонтах почвы (ПП 3). На остепненном участке гари, повторно пройденном пожарами (ПП 4), сумма обменных оснований уменьшилась относительно значения на контрольном участке в меньшей степени (в 1.6 раза) и только в верхнем горизонте почв. Напротив, в березняке, через год после высокоинтенсивного пожара, сумма обменных оснований в горизонте AY серогумусовой типичной почвы на гари составляет 32.6 смоль(экв)/кг (ПП 6), что в 2.5–4.5 раза выше данного показателя в не горевшем насаждении. Увеличение суммы обменных оснований после воздействия пожаров в березняках, вероятно, связано с тем, что при сгорании травянистой растительности и лиственного опада зольных элементов образуется значительно больше, чем при сгорании подстилки в хвойном насаждении.
Соотношение C : N в сосновых насаждениях, произрастающих на песчаных и супесчаных почвах, после воздействия пожаров несколько расширяется. Причинами вышеизложенного могут быть снижение активности протекающих микробиологических процессов на бедных сухих почвах в результате трансформации органического вещества, а также послепожарное иссушение почвы [22, 41]. В березняках, приуроченных к более богатым и влажным почвам, после пожара в верхнем гумусовом горизонте отмечается сужение соотношения C : N (табл. 2). Это обусловлено улучшением физико-химических показателей органического вещества и активизацией процессов минерализации [12, 18, 22, 35, 50].
Термическое воздействие на органогенные горизонты почв приводит к значительным изменениям в содержании доступных соединений азота, фосфора и калия. Степень этих изменений различна и определяется интенсивностью огня [41]. Выявлено, что максимальное количество питательных элементов отмечается в первые месяцы после пожара, а затем их содержание постепенно уменьшается [15, 22].
Проведенные исследования показали, что содержание валового азота в гумусовом горизонте дерново-подбура контрольного соснового насаждения составляет 0.21%, при этом его доля закономерно уменьшается вниз по профилю. После низкоинтенсивного пожара содержание валового азота в горизонте AY снижается до 0.17% (ПП 2), а после высокоинтенсивного – до 0.06% (ПП 3). На остепненном участке разнотравно-вейниковой гари (ПП 4) через 3 года после пожара доля азота в верхних горизонтах почвы в 2–2.5 раза ниже, чем в почвенных горизонтах контрольного соснового насаждения, что обусловлено неоднократным прогоранием напочвенного покрова и послепожарной эрозией почвы легкого гранулометрического состава. После высокоинтенсивного пожара в березняке (ПП 6), напротив, содержание азота в горизонте AY серогумусовой типичной почвы выше контрольного показателя. Возможно, это обусловлено отмиранием корней травянистых растений, разложение которых увеличивает содержание аммонийного азота [15, 17, 37, 55].
Ранее отмечалось, что повышение температуры поверхностных горизонтов почв после прохождения огня вызывает резкое увеличение в них содержания аммиачного азота [15, 22]. Выявлено, что на свежих гарях в сосняках после низко- и высокоинтенсивных пожаров (ПП 2 и ПП 3) в горизонте AY наблюдается увеличение содержания N–NH4. Максимальное значение N–NH4 (10.4 мг/кг) отмечено в горизонте AY, в сосняке, через год после воздействия верхового пожара (ПП 3). Однако ниже по почвенному профилю в других горизонтах, значение N–NH4 после пожаров снижается и создается его дефицит. Следует отметить, что на участке гари, повторно пройденной пожарами (ПП 4), содержание N–NH4 в горизонте AY в 2–3 раза ниже контроля, а в березняке после высокоинтенсивного пожара (ПП 6) отмечается снижение содержания N–NH4 во всех почвенных горизонтах. Уменьшение содержания аммонийного азота, вероятно, связано с активным его потреблением разросшейся травяной растительностью и почвенными микроорганизмами.
Увеличение содержания Р2O5 и K2О в пирогенных почвах происходит из-за значительного их количества в золе, образовавшейся при пожарах. В качестве положительного влияния пожара на почвы большинство исследователей отмечают увеличение количества как валовых, так и подвижных соединений калия [22, 41]. Данная закономерность отмечена в дерново-подбуре на остепненной разнотравно-вейниковой гари (ПП 4), где содержание K2О на 20–45% выше контрольного и в почвенном профиле березняка, пройденного огнем, где в гумусовом горизонте серогумусовой типичной почвы данный показатель выше контрольного в 2 раза. Однако на свежих гарях в сосняках содержание доступных соединений калия ниже контрольных показателей. Вымывание зольных элементов и снижение доли их содержания в почвенных горизонтах в сосновых насаждениях после пожаров обусловлено легким гранулометрическим составом почв и отсутствием травяной растительности на участках свежих гарей в данных условиях местопроизрастания.
После воздействия низовых пожаров в сосновых и березовых насаждениях и на повторно прогоревшей гари отмечается увеличение содержания подвижного фосфора в горизонте AY. Однако через год после высокоинтенсивного верхового пожара в сосняке (ПП 3) этот показатель в 2.5 раза ниже контроля. Вероятно, это также связано с послепожарной эрозией почвы вследствие полного сгорания подстилки и с вымыванием зольных элементов. Следует отметить, что в нижележащих горизонтах дерново-подбуров сосняков, пройденных огнем, содержание подвижного фосфора в 2–3.5 раза ниже контроля, что на легких песчаных и супесчаных почвах обусловлено вымыванием данного элемента.
Таким образом, сравнительный анализ свойств почв выявил различия в физико-химических параметрах их горизонтов в сосновых и березовых насаждениях. Серогумусовая типичная почва в длительно не горевшем березовом насаждении отличается меньшей кислотностью и большим содержанием гумуса по сравнению с дерново-подбуром соснового контрольного насаждения. Степень и направленность послепожарной трансформации свойств почв определяются их исходными параметрами, в том числе различиями гранулометрического состава, увлажненностью, интенсивностью огневого воздействия и восстановительными сукцессиями растительности.
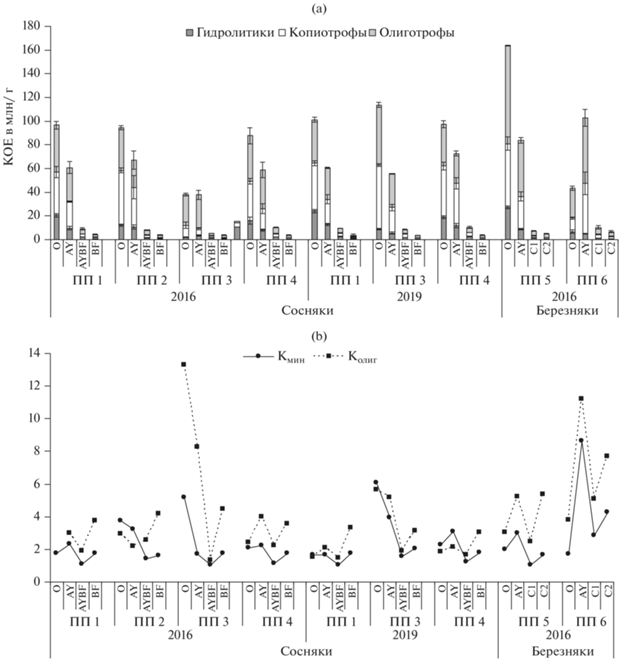
Послепожарная трансформация микробных комплексов почв. Анализ распределения ЭКТГМ по почвенным профилям в 2016 г. показал, что максимальная численность гетеротрофных микроорганизмов приурочена к органогенным горизонтам дерново-подбура иллювиально-железистого соснового (ПП 1) и серогумусовой типичной почвы березового (ПП 2) контрольных насаждений (рис. 2а). Количество гидролитиков в подстилке сосняка рододендронового (ПП 1) несколько ниже (20.56–24.2 млн КОЕ/г), чем в березняке разнотравно-вейниковом (ПП 5) (27 млн КОЕ/г), тогда как в гумусовых (9.7–12.8 млн КОЕ/г) и нижележащих горизонтах почв их количество сравнимо в сосновых и березовых контрольных насаждениях. Численность копиотрофов всегда выше, чем гидролитиков, особенно в органогенных горизонтах почвы березового насаждения на ПП 5 (Kмин = 1.98–3), тогда как олиготрофная группировка доминирует в профиле почв длительно не горевших сосняков и березняков, где Kолиг достигает 1.9–5.4 (рис. 2b). Значение Kолиг выше Kмин, особенно в нижней части профиля контрольных насаждений, что демонстрирует ослабление минерализационных процессов. При переходе от органогенных к минеральным горизонтам наблюдается резкое снижение численности КОЕ разных групп микроорганизмов, что характерно для почв таежных экосистем [8, 32]. Численность всех ЭКТГМ зависела от температуры и влажности почв (r = 0.93–0.99), содержания гумуса и валового азота (r = 0.98–0.99) и отрицательно коррелировала с рН (r = –0.66–0.75).
Рис. 2.
Численность эколого-трофических групп микроорганизмов (ЭКТГМ) (а) и коэффициенты минерализации и олиготрофности (b) в профиле почв на исследуемых участках.
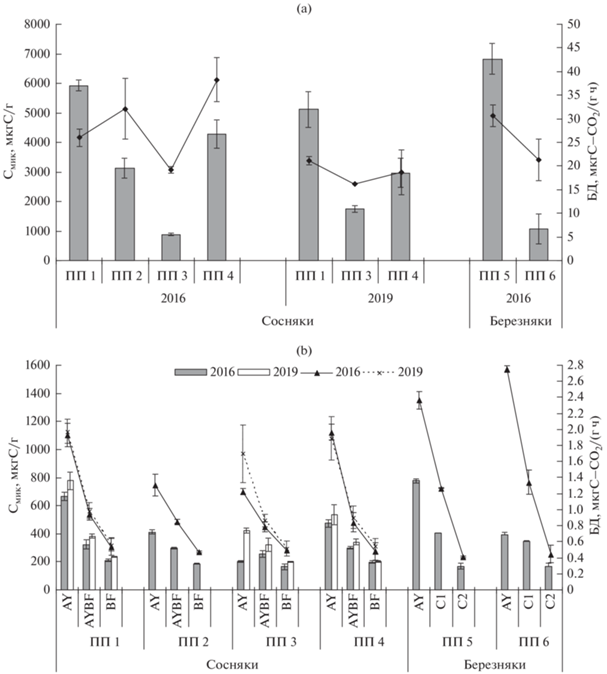
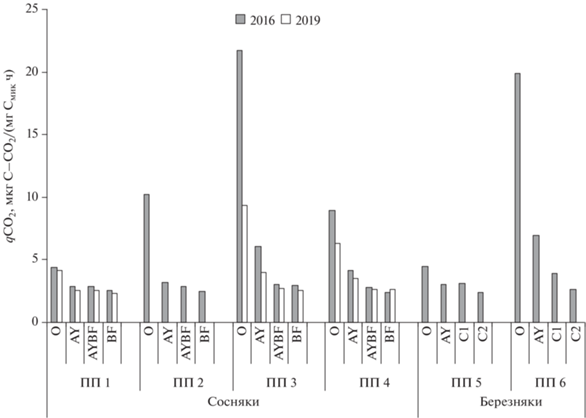
Содержание Смик и интенсивность БД почв сосновых и березовых контрольных насаждений приурочены к содержанию Сорг и N, которые с глубиной закономерно убывают (рис. 3). Данные показатели максимальны в подстилке березового разнотравно-вейникового насаждения (ПП 5), где достигают 6825 мкг С/г и 30.6 мкг С–СО2/(г ч), соответственно (рис. 3а), снижаясь до 777 мкг С/г и 2.36 мкг С–СО2/(г ч) в гумусовом горизонте (рис. 3b). В подстилке и горизонте AY контрольного сосняка рододендронового (ПП 1) значения рассматриваемых показателей несколько ниже (5920 и 665.5 мкг С/г и 26 и 1.93 мкг С–СО2/(г ч)). Микробиологические процессы в почвах сосновых и березовых насаждений на контрольных участках хорошо сбалансированы (qCO2 = 4.4–4.45 и 3.1–2.4 мкг С–СО2/(мг Смик ч) для органогенного и минеральных горизонтов соответственно) (рис. 4). Рассматриваемые показатели постепенно снижаются с глубиной почвенного профиля и имеют значимые связи с температурой (r = 0.93, 0.92), влажностью (r = 0.82, 0.81), рН (r = –0.80, –0.82), содержанием гумуса (r = 0.99, 0.98) и валового азота (r = 0.99, 0.99) для Смик и БД соответственно, что отражает устойчивый характер распределения микробного компонента в профиле почв контрольных насаждений.
Рис. 3.
Содержание углерода микробной биомассы (Смик) и интенсивность базального дыхания (БД) в подстилках (а) и минеральной части профиля (b) почв на исследуемых участках.
Пожары в сосновых и березовых насаждениях приводят к снижению численности и изменению структуры ЭКТГМ почв (рис. 2а). Максимальное сокращение численности ЭКТГМ дерново-подбура отмечено в сосняке через год после высокоинтенсивного пожара (ПП 3): гидролитиков – в 10.5 и 3 раза, копиотрофов – в 3.6 и 3.8 раза, по сравнению с контролем для подстилки и гумусового горизонта соответственно. Численность олиготрофов меньше в 1.5 раза только в подстилке. В нижележащем горизонте {AY + BF} количество микроорганизмов рассматриваемых групп также ниже контроля в 1.5–2 раза, в горизонте BF на глубине 11–25 см различия недостоверны. В верхних органогенных горизонтах почвы доминируют олиготрофные микроорганизмы (Kолиг = 13.28 и 8.27). В подстилке отмечено резкое преобладание микроорганизмов, использующих минеральные формы азота, над утилизаторами органических форм (Kмин = 5.16), что в совокупности с высокой олиготрофностью может свидетельствовать об обедненности озоленных подстилок после высокоинтенсивных пожаров доступными формами органических веществ. Зола содержит минеральные формы азота, что дает преимущество соответствующим микроорганизмам. Количество гидролитиков зависело от содержания гумуса и азота (r = 0.63, 0.99) и имело отрицательную связь с рН (r = –0.77), тогда как с другими физико-химическими свойствами почв достоверной корреляции не обнаружено. Численность копиотрофов и олиготрофов положительно коррелировала с температурой (r = 0.99, 0.85), влажностью почвы (r = 0.93, 0.63) и содержанием валового азота (r = = 0.89, 0.95).
Содержание Смик в подстилке и гумусовом горизонте дерново-подбура сосняка после высокоинтенсивного пожара (ПП 3) меньше аналогичных показателей на контрольном участке (ПП 1) в 6.7 и 3.5 раза соответственно, тогда как интенсивность БД снижалась в 1.3–1.6 раза (рис. 3). Значения qCO2 в подстилке и горизонте AY максимальны (21.7 и 3 мкг С–СО2/(мг Смик ч)) и выше контроля в 5–2 раза соответственно (рис. 4), что иллюстрирует большие затраты микроорганизмов на поддержание жизнедеятельности [47]. Высокие значения qCO2 связаны с большей скоростью отмирания микробной биомассы после высокоинтенсивных пожаров, что может указывать на потерю углерода почвой [39]. Отмечена тесная взаимосвязь Смик и БД с температурой (r = 0.87, 0.88), влажностью (r = 0.98, 0.99), рН (r = –0.79, –0.80), содержанием гумуса (r = 0.90, 0.66) и валового азота для БД (r = 0.99) после пожара высокой интенсивности.
Через 3 года после низкоинтенсивного пожара в сосняке (ПП 2) в подстилке количество гидролитиков было в 2 раза ниже контроля, тогда как численность других ЭКТГМ достоверно не изменялась как в подстилке, так и в минеральных почвенных горизонтах (рис. 2а). Коэффициенты минерализации и олиготрофности повышены в подстилке, тогда как в нижележащих горизонтах почвы не изменялись (рис. 2b). Содержание Смик в подстилке снижено почти в 2 раза и составляло 3138 мкг С/г, тогда как БД увеличилось на 20% от контроля (32.1 мкг С–СО2/(г ч)). В горизонте AY дерново-подбура значения рассматриваемых показателей на 35–40% ниже контрольных значений, тогда как в нижележащих горизонтах почвы снижены незначительно (рис. 3). В подстилке на ПП 2 значение qCO2 достигало 10.2 мкг С–СО2/мг Смик ч, что в 2.3 раза выше контроля, тогда как в минеральных почвенных горизонтах сравнимо с ним (рис. 4). Отмечена тесная взаимосвязь микробиологических параметров с температурой, влажностью и содержанием гумуса и азота (r = 0.89–0.99), значимой связи с рН не обнаружено.
На остепненном участке, образовавшемся после воздействия пожаров в сосняке (ПП 4), достоверных изменений численности ЭКТГМ в профиле дерново-подбура не обнаружено. Несколько увеличивается олиготрофность в подстилке и горизонте AY. Содержание Смик составляло 4290 мкг С/г в подстилке, что на 30% ниже контрольных значений, а интенсивность БД при этом достигала 38.25 мкг С–СО2/(г ч), превышая данный показатель на контроле на 50%. В горизонте AY Смик и БД также ниже контроля. Значение qCO2 в подстилке выше контроля в 2, а горизонте AY – в 1.5 раза и достигает 8.9 и 4.2 мкг С–СО2/(мг Смик ч) соответственно (рис. 4). Тесная взаимосвязь микробиологических параметров обнаружена с температурой (r = 0.88–0.99), рН (r = 0.91–0.99), содержанием гумуса и азота (r = 0.89–0.99), связь с влажностью слабее (r = 0.41–0.77).
Через год после высокоинтенсивного пожара в березняке разнотравно-вейниковом (ПП 6) в подстилке серогумусовой типичной почвы регистрировалось максимальное снижение численности микроорганизмов в 3–5 раз, содержания Смик – более, чем в 6 раз (1070 мкг С/г) и интенсивности БД – на 40% (21.32 мкг С–СО2/(г ч)). В гумусовом горизонте почти в 2 раза снижено количество гидролитиков, преимущество их получают олиготрофы и копиотрофы (Kмин = 8.6 и Kолиг = 11.2). Содержание Смик в 2 раза ниже, чем в горизонте AY контрольного березового насаждения (ПП 5) (394 мкг С/г), при этом БД повышалось на 20% от контроля и составляло 2.75 мкг С–СО2/(г ч). Значение qCO2 в подстилке превышало контроль (ПП 5) в 4.5 раза, достигая 19.92 мкг С–СО2/(мг Смик ч), в горизонте AY – выше контроля в 2 раза (6.98 мкг С–СО2/(мг Смик ч)). В нижележащих почвенных горизонтах, пройденного высокоинтенсивным пожаром березняка, значимых изменений численности и экофизиологических параметров микробного комплекса не выявлено. Количество гидролитиков после пожара в березняке зависело от содержания гумуса и азота (r = 0.99), влажности почвы (r = 0.95) и температуры (r = 0.87), тогда как численность копиотрофов и гидролитиков имела тесную корреляцию только с содержанием гумуса и азота (r = 0.99). Смик и БД коррелировали с температурой почвы (r = 0.98–0.99), влажностью (r = 0.71–0.79), содержанием гумуса (r = 0.73–0.94) и азота (r = = 0.76–0.96).
Динамика численности ЭКТГМ и экофизиологических параметров в профиле дерново-подбуров в пройденных пожарами сосняках в 2019 г. демонстрирует восстановительные тенденции в микробных комплексах почв (рис. 2–4). На четвертый год после высокоинтенсивного пожара (ПП 3) в подстилке и гумусовом горизонте в 2–3 раза ниже контрольных значений только количество гидролитиков, значительно повышаются Kмин и Kолиг (в 4 раза в подстилке и 2.5 раза в горизонте AY), что свидетельствует об усилении минерализации и изменении качества органического вещества [46, 55, 56]. Смик в подстилке достигает 1740 мкг С/г, в горизонте AY – 423 мкг С/г, что соответственно в 3 и 2 раза ниже аналогичных показателей на контрольном участке (ПП 1), интенсивность БД снижена на 15–25%. Величины qCO2 в органогенных горизонтах заметно снизились за 3 года после пожара, но превышают контрольные значения в 1.6–2.3 раза. Через 3 года в подстилке остепненного участка (ПП 4) количество гидролитиков снижено на 20%, в гумусовом горизонте – сравнимо с контролем. В органогенных горизонтах снижается олиготрофность (Kолиг = 1.84–2.15), интенсивно идут минерализационные процессы (Kмин = 2.25–3.1). Разрастание травяного покрова на остепненных безлесых участках предопределяет переход бактериального сообщества от олиготрофной к копитрофной стратегии жизнеобеспечения [55, 56] и, наряду со сменой гидротермических показателей, предполагает улучшение условий роста микробных популяций в течение 20–30 лет после пожаров [43]. Содержание Смик снижено в 1.7–1.8 раза в верхних горизонтах дерново-подбура остепненного участка гари (ПП 4), тогда как интенсивность БД на 10–15% ниже контроля. Величины qCO2 по сравнению с 2016 г. снижаются до 6.28 и 3.51 мкг С–СО2/мг Смик ч соответственно для подстилки и горизонта AY, но при этом превышают контроль на 40–50%.
Таким образом, пожары высокой интенсивности на исследуемых участках приводят к трансформации структурно-функциональных параметров микробоценоза в профиле почв до глубины 10 см минерального горизонта, тогда как низкоинтенсивные – затрагивают в основном подстилку. Коэффициент qCO2, как важный индикатор эффективности использования и доступности органических соединений [47], увеличивался в 2–5 раз после пожаров в подстилке и в 1.5–2 раза в гумусовом горизонте только после высокоинтенсивных пожаров. Увеличение значений qCO2 после пожаров за счет высокой интенсивности базального дыхания отражает активные минерализационные процессы и может объясняться преобладанием быстрорастущих r-стратегов, нуждающихся в большом количестве энергии для поддержания своей биомассы [44, 46]. Послепожарные восстановительные сукцессии инициируют развитие медленно растущих К-стратегов и значение qCO2 уменьшается.
Следовательно, микробоценозы почв после высокоинтенсивных пожаров в сосняках и березняках центральных районов Забайкальского края имеют длительный период восстановления и проявляют схожие тенденции с изменениями микробных комплексов после пожаров высокой интенсивности в южнотаежных сосняках лишайниково-зеленомошных [6, 8]. В то же время микробоценозы почв сосново-лиственничных насаждений Нижнего Приангарья после пожаров высокой интенсивности быстрее восстанавливали свою функциональную активность [8], что связано как с большей трофностью почвы, так и быстрым послепожарным развитием травянистого покрова, запасы которого через год после пожара возрастали в 6 раз. Степень трансформации микробных комплексов почв через 3 года после низкоинтенсивного пожара в сосняке, выраженная в снижении содержания Смик в подстилке и гумусовом горизонте дерново-подбура, изменении структуры ЭКТГМ и высоких коэффициентах qCO2, выше, чем после беглых пожаров низкой интенсивности в сосновых и сосново-лиственничных насаждениях Нижнего Приангарья [8]. Вероятно, это связано с засушливыми климатическими условиями районов исследования и большей степенью прогорания напочвенного покрова в Забайкальском крае. Беглые низкоинтенсивные пожары в насаждениях Нижнего Приангарья не приводили к значительному сгоранию подстилки, и в течение 1–3 послепожарных лет экофизиологическое состояние микробоценозов почв демонстрировало тенденцию к восстановлению к контрольным показателям.
Запасы углерода микробной биомассы и продуцирование СО2 в профиле почв после пожаров. Распределение запасов микробной биомассы и продукции СО2 по разным генетическим горизонтам дерново-подбура в сосновых и серогумусовой типичной почвы в березовых контрольных насаждениях не одинаково. Максимальные значения зарегистрированы в горизонте BF в сосновых (49–51 г Смик/м3 и 120–125 мг С–СО2/(м3 ч)) и в горизонте С2 березовых (38 г Смик/м3 и 92 мг С–СО2/(м3 ч)) контрольных насаждений (ПП 1 и ПП 5 соответственно) (рис. 5). Запасы Смик в гумусовом горизонте дерново-подбура и серогумусовой типичной почвы исследуемых насаждений не велики (30–37 г/м3) и составляют 29–31% от общих запасов Смик, что в 4 раза ниже, чем запасы Смик в горизонте AY серой почвы осинника южной тайги Калужской области, где мощность гумусового горизонта достигала 19 см [34]. Микробная продукция СО2 гумусовым горизонтом исследуемых почв достигала 92–95 мг С–СО2/(м3 ч), что составляет 30% от общей продукции данного профиля.
Рис. 5.
Запасы углерода микробной биомассы (а) и продуцирование СО2 (b) в профилях почв на исследуемых участках после пожаров разной интенсивности.
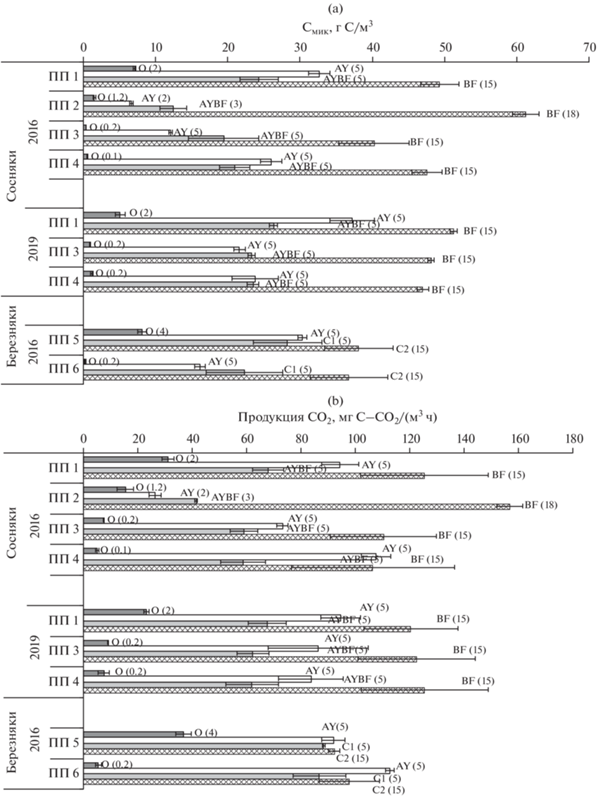
Несмотря на максимальное содержание углерода микробной биомассы в подстилке лесных ценозов, подстилка вносила наименьший вклад (4–12%) в профильные запасы Смик и продукции СО2 исследуемых почв, что связано как с ее низкой плотностью, так и малой мощностью в исследованных насаждениях Забайкальского края.
В профилях дерново-подбура в сосновом и серогумусовой типичной почвы в березовом контрольных насаждений общие запасы Смик сравнимы и достигали 113–120 г С/м3, при этом общая потенциальная микробная продукция СО2 составила 305–320 мг С–СО2/(м3 ч) (рис. 6), что несколько ниже, чем в дерново-подзолистых и серых почвах светлохвойных и подтаежных лесов Средней Сибири [6, 7], и в 1.5–8 раз меньше, чем в дерново-подзолистых и серых лесных почвах в разных типах южно-, средне- и подтаежных лесов европейской части России [2, 32, 34].
Рис. 6.
Суммарные запасы углерода микробной биомассы (1) и общее микробное продуцирование СО2 (2) в профилях почв на исследуемых участках после пожаров разной интенсивности.
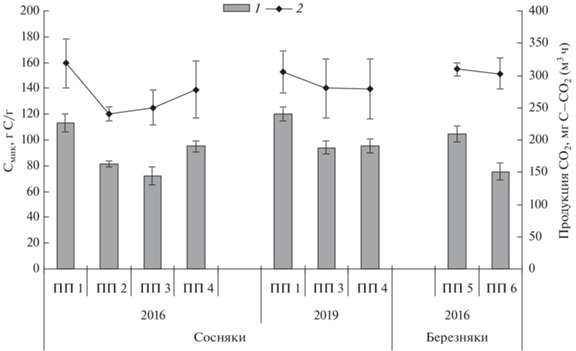
После пожаров в изучаемых лесных насаждениях происходит трансформация профильного распределения запасов Смик и микробной продукции углекислого газа. Самое значительное сокращение запаса Смик отмечено в подстилке: в 30 раз – в березняке через год после высокоинтенсивного пожара (ПП 6), более чем в 20 раз – в сосняке после высокоинтенсивного пожара (ПП 3), в 12 раз – на остепненном участке гари (ПП 4) и почти в 5 раз – в сосняке после низкоинтенсивного пожара (ПП 2). Микробное продуцирование СО2 подстилкой также меньше в 2–6.6 раза, но не так заметно, как запас Смик. Отмеченные тенденции связаны с полным прогоранием подстилки. После высокоинтенсивных пожаров показатели длительно восстанавливаются. Через 3 года в сгоревшем сосняке (ПП 3) и на остепненном участке гари (ПП 4) запас Смик и продуцирование СО2 подстилкой меньше в 2.5–5 раз по сравнению с контрольными значениями на ПП 1 и ПП 5.
Значительная трансформация в профильном распределении рассматриваемых величин отмечена в дерново-подбуре сосняка после низкоинтенсивного пожара (ПП 2): запасы Смик и микробная продукция СО2 снижены в 1.6–5 раз в горизонтах AY и {AY + BF} (6.7 и 12.4 г С/м3 и 26.4 и 41.5 мг С–СО2/(м3 ч)) и увеличиваются на 25% в горизонте BF (62 г С/м3 и 157 мг С–СО2/(м3 ч)). Это связано как с воздействием пожара, так и с морфологическими особенностями почвы данного участка, где мощность горизонтов AY и {AY + BF} меньше, а горизонта BF больше, чем в контрольном сосновом насаждении на ПП 1 (рис. 5).
Через год после пожара высокой интенсивности в сосняке (ПП 3) запас Смик сокращается почти в 3 раза в горизонте AY (12 г С/м3) и на 20% в нижележащих горизонтах. При этом микробная продукция СО2 сокращается на 10–20% в разных горизонтах дерново-подбура. На остепненном участке, образовавшемся после воздействия пожаров в сосняке (ПП 4), запас Смик снижен на 15–20% только в горизонтах AY и {AY + BF}, тогда как продуцирование углекислого газа гумусовым горизонтом было больше на 15%, а нижележащими горизонтами почвы – меньше на 15%.
Через год после пожара высокой интенсивности в березняке (ПП 6) запас Смик снижен в 2 раза в горизонте AY серогумусовой типичной почвы (16 г С/м3), на 20% – в горизонте С1 (22 г С/м3) и сравним с контролем в горизонте С2, тогда как продукция СО2 гумусовым горизонтом увеличилась на 20% (113 мг С–СО2/(м3 ч)) и также сравнима с контролем в нижней части.
Через 3 года (в 2019 г.) на участках в сосняках после высокоинтенсивных пожаров (ПП 3 и ПП 4) выявлено, что запасы Смик уменьшились на 40% в гумусовом горизонте (22–24 г С/м3), на 10% – в {AY + BF} и не отличались ниже по профилю. Микробная продукция СО2 сократилась на 10% только в гумусовом горизонте дерново-подбура, относительно контрольных показателей (ПП 1).
Суммарные запасы углерода микробной биомассы в профиле почв достоверно ниже контроля на 30% в сосняке после низкоинтенсивного пожара (ПП 2) и березняке после пожара высокой интенсивности (ПП 6), на 40% – в сосняке после пожара высокой интенсивности (ПП 3) и на 20% – на остепненном участке гари (ПП 4). Общая микробная продукция СО2 ниже контроля на 20–25% на участках прогоревших сосняков вне зависимости от интенсивности (245–280 мг С–СО2/(м3 ч)) и сравнима с контролем в почве березняка после высокоинтенсивного пожара (303 мг С–СО2/(м3 ч)). Через 3 года в 2019 г. в профиле дерново-подбуров в сосняках на гарях суммарный запас Смик стал меньше контроля на 20%, но достоверных снижений общей микробной продукции СО2 не выявлено.
Таким образом, послепожарная трансформация профильного распределения запасов Смик и микробной продукции углекислого газа выражается в значительном их сокращении в подстилках, гумусовом и нижележащем горизонтах до 10 см минерального профиля почв. Однако ниже 10 см достоверное уменьшение рассматриваемых величин отмечено только после высокоинтенсивных пожаров в сосняках.
ЗАКЛЮЧЕНИЕ
Исследования почв сосновых и березовых лесов центральных районов Забайкальского края показали, что пожары разной интенсивности оказывают значительное влияние на физико-химические и микробиологические свойства почв. Степень и направленность послепожарной трансформации свойств почв определяются их исходными параметрами, в том числе различиями гранулометрического состава и увлажненностью, а также интенсивностью огневого воздействия и восстановительными сукцессиями растительности. Показано, что в дерново-подбурах, характерных для сосновых насаждений, произрастающих на почвах легкого гранулометрического состава, после пожаров отмечается снижение показателей суммы обменных оснований, валового азота, подвижных соединений калия и фосфора, увеличение соотношения C : N. В серогумусовых типичных почвах в производных березовых насаждениях, приуроченных к более богатым и увлажненным почвам, отмечается увеличение данных показателей и сужение соотношения C : N. Содержание гумуса в верхнем почвенном горизонте уменьшается только на свежей гари в сосняке после высокоинтенсивного пожара, тогда как на других участках оно увеличивается. Уменьшение кислотности в сторону нейтральных значений характерно для всех участков.
Пожары высокой интенсивности в сосновых и березовых насаждениях приводят к достоверному снижению содержания Смик и интенсивности БД, численности и изменению структуры ЭКТГМ в профиле почв до глубины 10 см минеральных горизонтов. Максимальные изменения наблюдаются в подстилке и гумусовом горизонте через год после воздействия пожаров. Низкоинтенсивные пожары в сосновом насаждении приводят к трансформации структурно-функциональных параметров микробоценоза в основном в подстилке. Коэффициент qCO2 выше контроля в 2–5 раз в подстилке на всех участках и в 1.5–2 раза в гумусовом горизонте только после высокоинтенсивных пожаров. Послепожарная динамика микробиологических параметров в дерново-подбурах сосняков демонстрирует восстановительные тенденции: через 4 года после высокоинтенсивного пожара в сосняке и на старой остепненной гари в 1.5–3 раза уменьшилась численность гидролитиков, в 2–3 раза – содержание Смик, на 10–25% – интенсивность БД, величины qCO2 заметно уменьшаются за 3 послепожарных года, но в 1.5–2.3 раза превышают контроль.
На свежих гарях в сосняках значительно сокращаются запасы Смик и микробная продукция углекислого газа в минеральной части почвенного профиля дерново-подбуров до глубины 10 см, тогда как на остепненном участке, образовавшемся после воздействия пожаров в сосняке, и в березняке после пожара высокой интенсивности в гумусовом горизонте почв запасы Смик уменьшаются на 15–20%, а микробная продукция углекислого газа увеличивается на 10–20%. Это связано с активизацией минерализационных процессов при разложении травянистого опада [7], запасы которого увеличивались по сравнению с контролем в 2.7–4.5 раза, обеспечивая более полное и быстрое его разложение и вовлечение в биологический круговорот.
Суммарные запасы углерода микробной биомассы в профиле почв на 20–40% ниже контрольных значений на всех участках, пройденных пожарами, и составляют 72–95 г/м3. При этом общая микробная продукция СО2 ниже контроля на 15–20% на участках сосняков, пройденных пожарами, вне зависимости от интенсивности (240–280 мг С–СО2/(м3 ч)) и сравнима с контролем в почве березняка после высокоинтенсивного пожара (303 мг С–СО2/(м3 ч)).
Таким образом, физико-химические и микробиологические свойства почв светлохвойных и лиственных насаждений Забайкальского края имеют длительный период восстановления после воздействия пожаров, что обусловливает необходимость проведения комплексных долговременных исследований почв этого региона.
Список литературы
Ананьева Н.Д., Сусьян Е.А., Гавриленко Е.Г. Особенности определения углерода микробной биомассы почвы методом субстрат-индуцированного дыхания // Почвоведение. 2011. № 11. С. 1327–1333.
Ананьева Н.Д., Сусьян Е.А., Рыжова И.М., Бочарникова Е.О., Стольникова Е.В. Углерод микробной биомассы и микробное продуцирование двуокиси углерода дерново-подзолистыми почвами постагрогенных биогеоценозов и коренных ельников южной тайги (Костромская область) // Почвоведение. 2009. № 9. С. 1108–1116.
Анучин Н.П. Лесная таксация. М.: Лесная промышленность, 1982. 552 с.
Аринушкина Е.В. Руководство по химическому анализу почв. М.: Изд-во МГУ, 1970. 489 с.
Безкоровайная И.Н., Иванова Г.А., Тарасов П.А., Сорокин Н.Д., Богородская А.В., Иванов В.А., Конард С.Г., Макрае Д.Дж. Пирогенная трансформация почв сосняков средней тайги Красноярского края // Cибирский экологический журн. 2005. № 1. С. 143–152.
Богородская А.В., Кукавская Е.А. Состояние микробных сообществ в почвах лиственных и светлохвойных лесов Средней Сибири после рубок и пожаров // Лесоведение. 2016. № 5. С. 383–396.
Богородская А.В., Пономарева Т.В., Ефимов Д.Ю., Шишикин А.С. Трансформация эколого-функциональных параметров микробоценозов почв на просеках линий электропередач в условиях Средней Сибири // Почвоведение. 2017. № 6. С. 731–743.
Богородская А.В. Структурно-функциональные параметры микробоценозов почв после пожаров в светлохвойных насаждениях // Воздействие пожаров на светлохвойные леса Нижнего Приангарья. Новосибирск: Наука, 2022. С. 147–167.
Буряк Л.В., Кукавская Е.А, Каленская О.П., Малых О.Ф., Бакшеева Е.О. Последствия лесных пожаров в южных и центральных районах Забайкальского края // Сибирский лесной журн. 2016. № 6. С. 94–102. https://doi.org/10.15372/SJFS20160609
Буянтуева Л.Б., Никитина Е.П. Микробиологические исследования каштановых почв юго-западного Забайкалья // Самарский научный вестник 2015. № 2. С 38–40.
Воробьева Л.А. Химический анализ почв. М.: Изд-во Моск. ун-та, 1998. 272 с.
Габбасова И.М., Гарипов Т.Т., Сулейманов Р.Р., Комиссаров М.А., Хабиров И.К., Сидорова Л.В., Назырова Ф.И., Простякова З.Г., Котлугалямова Э.Ю. Влияние низовых пожаров на свойства и эрозию лесных почв Южного Урала (Башкирский государственный природный заповедник) // Почвоведение. 2019. № 4. С. 412–421.
Гениатулин Р.Ф. Энциклопедия Забайкалья. Т. 1. Читинская область. Новосибирск: Наука, 2000. 102 с.
Герасимов И.П. Предбайкалье и Забайкалье. М.: Наука, 1965. 492 с.
Горбунова Ю.С., Девятова Т.А., Григорьевская А.Я. Влияние пожаров на почвенный и растительный покров лесов центра русской равнины // Вестник Воронежского гос. ун-та. Сер. Химия, биология, фармация. 2014. № 4. С.52–56.
Гродницкая И.Д., Карпенко Л.В., Пашкеева О.Э., Гончарова Н.Н., Старцев В.В., Батурина О.А., Дымов А.А. Влияние лесных пожаров на микробиологические свойства торфяных олиготрофных почв и торфяно-подзолов глеевых в болотах северной части Сым-Дубчесского междуречья (Красноярский край) // Почвоведение. 2022. № 4. С. 454–468.
Гынинова А.Б., Дыржинов Ж.Д., Гончиков Б.-М.Н. Особенности трансформации почв под влиянием пожаров в сосновых лесах Прибайкалья // Вестник Бурятского гос. ун-та. 2018. № 1. С. 44–53. https://doi.org/10.18101/2587-7143-2018-1-44-53
Дымов А.А., Дубровский Ю.А., Габов Д.Н. Пирогенные изменения подзолов иллювиально-железистых (средняя тайга, Республика Коми) // Почвоведение. 2014. № 2. С. 144–154.
Евдокименко М.Д. Пирогенные нарушения лесорастительной среды в сосняках Забайкалья и их лесоводственные последствия // Лесоведение. 2014. № 1. С. 3–12.
Казеев К.Ш., Одабашян М.Ю., Трушков А.В., Колесников С.И. Оценка влияния разных факторов пирогенного воздействия на биологические свойства чернозема // Почвоведение. 2020. № 11. С. 1372–1382.
Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
Краснощеков Ю.Н., Чередникова Ю.С. Постпирогенная изменчивость лесных почв в горном Прибайкалье. Новосибирск: СО РАН, 2022. 164 с.
Курбатский Н.П. Исследование количества и свойств лесных горючих материалов. Вопросы лесной пирологии. Красноярск, 1970. С. 5–58.
Макарова А.П., Напрасникова Е.В. Микробиологическая и биохимическая характеристика мерзлотных глеевых почв Северного Забайкалья // Известия Иркутского гос. ун-та. Сер.Биология. Экология. 2011. № 1. С. 25–32.
Максимова Е.Ю., Кудинова А.Г., Абакумов Е.В. Функциональная активность почвенных микробных сообществ постпирогенных островных сосновых лесов г. Тольятти Самарской области // Почвоведение. 2017. № 2. С. 249–255.
Маслов М.Н., Маслова О.А., Поздняков Л.А., Копеина Е.И. Биологическая активность почв горнотундровых экосистем при постпирогенном восстановлении // Почвоведение. 2018. № 6. С. 728–737.
Масягина О.В., Евграфова С.Ю., Титов С.В., Прокушкин А.С. Динамика дыхания почвы на разных стадиях послепожарной восстановительной сукцессии на примере разновозрастных гарей Эвенкии // Экология. 2015. № 1. С. 23–32.
Мишустин Е.Н., Емцев В.Т. Микробиология. М.: Агропромиздат, 1987. 368 с.
Практикум по микробиологии / Под ред. Нетрусова А.И. М.: Academia, 2005. 603 с.
Ставрова Н.И., Калимова И.Б., Горшков В.В., Дроздова И.В., Алексеева-Попова Н.В., Баккал И.Ю. Долговременные послепожарные изменения характеристик почв в темнохвойных лесах Европейского Севера // Почвоведение. 2019. № 2. С. 246–256.
Старцев В.В., Дымов А.А., Прокушкин А.С. Почвы постпирогенных лиственничников Средней Сибири: морфология, физико-химические свойства и особенности почвенного органического вещества // Почвоведение. 2017. № 8. С. 912–925.
Стольникова Е.В., Ананьева Н.Д., Чернова О.В. Микробная биомасса, ее активность и структура в почвах старовозрастных лесов Европейской территории России // Почвоведение. 2011. № 4. С. 479–494.
Сукачев В.Н., Зонн С.В. Методические указания по изучению типов леса. М.: Изд-во АН СССР, 1961. 144 с.
Сусьян Е.А., Ананьева Н.Д., Гавриленко Е.Г., Чернова О.В., Бобровский М.В. Углерод микробной биомассы в профиле лесных почв южной тайги // Почвоведение. 2009. № 10. С. 1233–1240.
Цибарт А. С., Геннадиев А. Н. Влияние пожаров на свойства лесных почв Приамурья (Норский заповедник) // Почвоведение. 2008. № 7. С. 783–792.
Чернов Т.И., Семенов М.В. Управление почвенными микробными сообществами: возможности и перспективы (обзор) // Почвоведение. 2021. № 12. С. 1506–1522.
Шахматова Е.Ю. Изменение свойств подстилки и почв на гарях в сосновых лесах Западного Забайкалья // Наука и образование. 2017. № 3. С. 101–106.
Anderson J.P.E., Domsch K.H. A physiological method for the quantitative measurement of microbial biomass in soils // Soil Biol. Biochem. 1978. V. 10. P. 314–322. https://doi.org/10.1016/0038-0717(78)90099-8
Anderson T.-H., Domsch K.H. Application of eco-physiological quotients (qCO2 and qD) on microbial biomasses from soils of different cropping histories // Soil Biol. Biochem. 1990. V. 22. P. 251–255.https://doi.org/10.1016/0038-0717(90)90094-G
Barrett K., Baxter R., Kukavskaya E., Balzter H., Shvetsov E., Buryak L. Postfire recruitment failure in Scots pine forests of southern Siberia // Remote Sensing of Environment. 2020. V. 237. https://doi.org/10.1016/j.rse.2019.111539
Certini G. Fire as a soil-forming factor. 2014. Ambio 43. P. 191–195. https://doi.org/10.1007/s13280-013-0418-2
Certini G., Moya D., Lucas-Borja M.E., Mastrolonardo G. The impact of fire on soil-dwelling biota: A review // Forest Ecology and Management. 2021. V. 488. P. 118989. https://doi.org/10.1016/j.foreco.2021.118989
Cutler N.A., Arróniz-Crespo M., Street L.E., Jones D.L., L. Chaput D.L., DeLuca T.H. Long-term recovery of microbial communities in the boreal bryosphere following fire disturbance // Microb. Ecol. 2017. V. 73. P. 75–90. https://doi.org/10.1007/s00248-016-0832-7
Dicen G.P., Rallos R.V., Labides J.L.R., Navarrete I.A. Vulnerability of soil organic matter to microbial decomposition as a consequence of burning // Biogeochemistry. 2020. V. 150. P. 123–137. https://doi.org/10.1007/s10533-020-00688-1
Dooley S.R., Treseder K.K. The effect of fire on microbial biomass: a meta-analysis of field studies // Biogeochemistry. 2012. V. 109. P. 49–61. https://doi.org/10.1007/s10533-011-9633-8
Fritze H., Pennanen T., Pietikainen J. Recovery of soil microbial biomass and activity from prescribed burning // Can. J. Forest Research. 1993. V. 23. P. 1286–1290. https://doi.org/10.1139/x93-164
Insam H., Haselwandter K. Metabolic quotient of the soil microflora in relation to plant succession // Oecologia. 1989. V. 79. P. 174–178. https://doi.org/10.1007/bf00388474
IUSS Working Group WRB, 2015. World Reference Base for Soil Resources 2014, update 2015. Int. soil Classif. Syst. naming soils creating legends soil maps.
Kukavskaya E.A., Buryak L.V., Shvetsov E.G., Conard S.G., Kalenskaya O.P. The impact of increasing fire frequency on forest transformations in southern Siberia // Forest Ecology Management. 2016. V. 382. P. 225–235. https://doi.org/10.1016/j.foreco.2016.10.015
Ludwig S.M., Alexander H.D., Kielland K., Mann P. J., Natali S.M., Ruess R.W. Fire severity effects on soil carbon and nutrients and microbial processes in a Siberian larch forest // Global Change Biology. 2018. V. 24. P. 5841–5852. https://doi.org/10.1111/gcb.14455
Mataix-Solera J., Guerrero C., García-Orenes F., Bárcenas G.M., Torres M. P. Forest fire effects on soil microbiology // Fire Effects on Soils and Restoration Strategies. Enfield: Cerdà Science Publishers, 2009. P. 133–175.https://doi.org/10.1201/9781439843338n
Pingree M.R.A., Kobziar L.N., 2019. The myth of the biological threshold: A review of biological responses to soil heating associated with wildland fire // Forest Ecology and Management. V. 432. P. 1022–1029. https://doi.org/10.1016/j.foreco.2018.10.032
Prendergast-Miller M.T., DeMenezes A.B., Macdonald L.M., Toscas P., Bissett A., Baker G., Farrell M., Richardson A.E., Wark T., Thrall P.H. Wildfire impact: Natural experiment reveals differential short-term changes in soil microbial communities // Soil Biol. Biochem. 2017. V. 109. P. 1–13. https://doi.org/10.1016/j.soilbio.2017.01.027
Pressler Y., Moore J.C., Cotrufo M.F. Belowground community responses to fire: meta-analysis reveals contrasting responses of soil microorganisms and mesofauna // Oikos. 2019. V. 128. P. 309–327. https://doi.org/10.1111/oik.05738
Sharma U., Garima, Sharma J.C., Devi M. Effect of forest fire on soil nitrogen mineralization and microbial biomass: A review // J. Pharmacognosy Phytochemistry. 2017. V. 6. P. 682–685.
Yeager C.M., Northup D.E., Grow C.C., Barns S.M., Kuske C.R. Changes in nitrogen-fixing and ammonia-oxidizing bacterial communities in soil of a mixed conifer forest after wildfire // Appl. Environ. Microbiol. 2005. V. 71. P. 2713–2722. https://doi.org/10.1128/AEM.71.5.2713-2722.2005
Дополнительные материалы отсутствуют.