

Радиационная биология. Радиоэкология, 2019, T. 59, № 5, стр. 527-531
ВЛИЯНИЕ ОБЛУЧЕНИЯ ПРОТОНАМИ НА РЕШЕНИЕ МЫШАМИ КОГНИТИВНОГО ТЕСТА НА ПОИСК ВХОДА В УКРЫТИЕ И НЕЙРОГЕНЕЗ ВЗРОСЛОГО МОЗГА
И. И. Полетаева 1, *, О. В. Перепелкина 1, Н. А. Огиенко 1, А. Ю. Тарасова 1, И. Г. Лильп 1, И. В. Кошлань 3, 4, Г. В. Павлова 2, А. В. Ревищин 2
1 Биологический факультет МГУ им. М.В. Ломоносова
Москва, Россия
2 Институт биологии гена РАН
Москва, Россия
3 Объединенный институт ядерных исследований
Дубна, Россия
4 Государственный университет “Дубна”
Дубна, Россия
* E-mail: ingapoletaeva@mail.ru
Поступила в редакцию 05.12.2018
Аннотация
Облучение мышей самцов F1 (CBA × С57BL/6J) пучком протонов (1, 2 и 4 Гр) вызвало неоднозначные проявления в адаптивном поведении животных. При использовании когнитивного теста на поиск входа в укрытие облученные мыши достоверно хуже, чем контроль, решали те этапы теста, где для попадания в комфортное отделение камеры надо было копать стружку. В то же время те этапы теста, где надо было удалить “пробку”, препятствовавшую переходу, они решали тест достоверно лучше контроля. У мышей облученных групп выявлено ослабление нейрогенеза в двух пролиферативных зонах переднего мозга.
Исследование влияния на ЦНС и поведение заряженных частиц космического происхождения (в частности, протонов) в экспериментах на животных актуально в связи с перспективой полетов человека в дальний космос [1–4]. Поскольку протоны составляют значимую часть космического излучения, то их влияние на организм необходимо исследовать, используя разные биологические модели. Обнаруживаются как негативные эффекты облучения протонами (снижение показателей нейрохимических процессов, усиление тревожности и др.), так и отсутствие значимых изменений [2–5], и эти эффекты отличаются от таковых при облучении заряженными ионами тяжелых металлов [6].
В настоящей статье представлены данные по влиянию облучения протонами на решение лабораторными мышами теста на поиск входа в укрытие [7]. Этот тест позволяет оценить способность животного, стремящегося избежать пребывания в ярко освещенной части экспериментальной камеры, найти замаскированный вход (лаз) в темную часть установки. Способность решить этот тест при замаскированном лазе базируется на понимании животным правила “неисчезаемости”, т.е. понимания, что объект, который ранее был в наличии, но более не видим, продолжает существовать, и его можно отыскать. Эту способность (впервые описанную Ж. Пиаже у маленьких детей, см. [8]) обнаруживают у животных без предварительного научения. Она базируется на наличии у животного соответствующей когнитивной способности. Данный тест на поиск входа в укрытие (в англоязычной литературе он называется puzzle-box) в модификации для мышей был предложен М. Голсуорси относительно недавно [9, 10].
МАТЕРИАЛЫ И МЕТОДИКА
Экспериментальные животные. В работе были использованы самцы мышей-гибридов (CBA × × C57BL/6J)F1 в возрасте 4 мес. Сравнивали поведение животных, облученных протонами (n = 25), и контрольных животных (n = 39). Использовали две контрольные группы. 1-я – группа “контроль-Дубна”, эти животные подвергались тем же манипуляциям, что и облученные, но облучения не получали, 2-я – “контроль-Москва” (интактные мыши, которые подобным манипуляциям не подвергались).
Облучение мышей проводили в лаборатории радиобиологии ОИЯИ на протонном пучке фазотрона Медико-технического комплекса “Лаборатории ядерных проблем ОИЯИ” (г. Дубна). Общее однократное облучение проводили в расширенном пике Брэгга с дозами: 1, 2, 4 Гр – первая серия и 4 Гр – вторая серия (мощность дозы около 1 Гр/мин, энергия протонов на входе пучка составляла 150 МэВ). Животное помещали в жесткий пластиковый контейнер, в котором оно не могло двигаться.
Тест на поиск входа в укрытие. В 1-й серии тест на поиск входа в укрытие проводили трижды. Первый раз – до облучения, при этом различий между выполнением его животными будущих экспериментальной и контрольных группами обнаружено не было, второй раз – через 2 мес., а третий – через 5.5 мес. после облучения. Во 2-й серии тест проводили через 1 мес. после облучения. Результаты выполнения данного теста мышами экспериментальных и контрольных групп в обеих сериях оказались сходными. При тестировании мышь помещают в ярко освещенную часть пластиковой камеры (74 × 30 × 28 см), в которой есть углубленный в пол “лаз” в стенке, разделяющей камеру на освещенную и темную части. Через этот лаз мышь может попасть в темную, более комфортную часть камеры. В течение 2 дней (по четыре пробы в каждый) оценивали быстроту (латентный период) перехода животного в темноту. В пробах 1 и 2 лаз открыт, и животное переходит в темноту беспрепятственно. В пробах 3 и 4 лаз замаскирован чистой древесной стружкой вровень с полом. На 2-й день эксперимента в пробе 5 лаз снова замаскирован стружкой, а в пробах 6 и 7 его закрывают легкой пробкой из пластика и картона, которую мышь может легко вытащить. В 8-й пробе теста вся стенка с лазом засыпана стружкой на высоту 5–7 см. Для решения проб 1–5 и 8 мыши “давалось” 3 мин, для проб 6 и 7 – 4 мин. Успешность выполнения теста оценивали по времени перехода мыши в темную часть камеры, а для группы животных – по доле мышей, решивших “когнитивные” пробы теста (т.е. пробы, когда лаз бывает закрыт и для попадания в темную часть камеры нужно “понять”, что лаз только замаскирован, а не исчез). Если животное не перешло в темную часть камеры за 3 или 4 мин (в зависимости от номера пробы), время решения теста обозначается как 180 или 240 с.
Нейрогенез взрослого мозга оценивали по числу делящихся клеток в зубчатой фасции гиппокампа и в субвентрикулярной зоне переднего мозга. Подсчет клеток проводили на коронарных срезах мозга мышей с последующей окраской на маркер делящихся клеток Ki-67 [11].
Статистическую значимость межгрупповых различий оценивали с помощью 1-факторного ANOVA с LSD post hoc тестом по Фишеру, значимость различий альтернативных долей – с помощью метода φ по Фишеру.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Общее состояние животных оценивали как непосредственно после облучения, так и спустя разные сроки после него (вплоть до нескольких месяцев). Это имеет вполне серьезные основания из-за постепенного формирования последствий облучения, в частности, его воздействий на процессы нейрогенеза [12]. В течение 4 и 10 мес. после облучения не было обнаружено ни резкого снижения массы тела облученных мышей, ни изменений в адекватности их поведения.
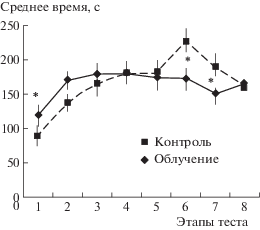
Среднее время решения теста у мышей групп “облучение” и “контроль” (суммарно для животных обеих серий) практически не различалось, пример – на рис. 1. Время решения теста (пробы 6 и 7) у контрольных мышей было большим за счет большей доли животных, не решивших эти “когнитивные” этапы теста, поскольку им “засчитывалось” время – 240 с.
Рис. 1.
Среднее время (± ош. средн.) решения теста на поиск входа в укрытие облученными и контрольными мышами (1-я серия экспериментов с облучением, тестирование, через 1 мес. после облучения). *Достоверно отличается от показателя контроля, p < 0.05 (1 факт. ANOVA, LSD post hoc тест по Фишеру).
Fig. 1. The time (means ± st. error) of puzzle-box “cognitive” trials solution by irradiated (rhombs) and control (squares) mice (the 1st experimental series with irradiation, 1 month after irradiation). *Significantly different from control score, p < 0.05 (1 factorial ANOVA, Fisher LSD post hoc тест).
В то же время доли мышей двух групп, решивших наиболее сложные пробы теста (когда лаз в темную часть камеры замаскирован либо стружкой, либо пробкой), достоверно различались. Среди мышей облученных групп было меньше особей, решивших пробы с лазом, присыпанным стружкой, тогда как пробу с лазом, закрытым пробкой, они решали достоверно более успешно, чем контрольные (результаты теста во 2-й серии представлены на рис. 2). В обеих сериях опытов с облучением результаты были сходными. С целью анализа причин аномалии в реакции на стружку у облученных мышей в 1-й серии опытов с ними был проведен специальный тест на реакцию “копания”. В этом опыте животное помещали в камеру, по углам которой были насыпаны кучки стружки. Оценивали число эпизодов копания, стоек и подходов к кучкам, а также число эпизодов груминга. Данные однофакторного ANOVA (для пяти групп – три группы мышей, облученных разными дозами, и две контрольные) были на грани достоверности, однако post hoc тест LSD (по Фишеру), несмотря на малый размер групп, выявил статистически значимые различия – меньшее число эпизодов “копания” у облученных мышей (3.4 ± 2.04, 3.0 ± 2.63 и 4.5 ± 3.22 для групп, получивших облучение, и 10.8 ± 2.04 и 8.8 ± 1.52 для контрольных групп, р < 0.05). Для числа эпизодов груминга однофакторный ANOVA показал достоверное влияние фактора “группа” (F1–4 = 3.4862, р = 0.0222) с большим числом эпизодов у мышей облученных групп. Таким образом, у мышей, получивших облучение протонами, видоспецифическая реакция “копания” была выражена слабее, чем у контроля, возможно, в связи с более “тревожной” реакцией облученных мышей на стружку. Одним из объяснений этого феномена может быть нарушение обоняния у облученных животных. Тревожность облученных мышей при контакте со стружкой выразилась и в большем числе эпизодов груминга у облученных мышей.
Рис. 2.
Доли мышей (ось ординат), решивших “когнитивные” пробы теста на поиск входа в укрытие во 2-й серии с облучением протонами. а – пробы 3–5 (лаз засыпан стружкой), б – пробы 6 и 7 (лаз закрыт пробкой). 1 – облученные мыши, доза 4 Gy, 2 – “контроль-Дубна”, 3 – “контроль-Москва”. *, **Статистически значимые различия при р < 0.05 и р < 0.01 соответственно (метод φ по Фишеру).
Fig. 2. The proportion of mice (ordinate), which solved the “cognitive” trials of puzzle-box test in the 2nd series with proton irradiation. a – trials 3–5 (the underpass is masked by wood shavings, b – the trials 6 and 7 (the underpass is blocked by the cork). 1 – irradiated mice, dosage 4Gy, 2 – “control Dubna” group, 3 – “control Moscow” group. *, ** Statistically significant differences, p < 0.05 and p < 0.001 respectively (Fisher φ test).
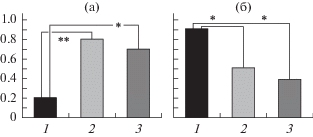
Статистически значимая большая доля мышей облученной группы, решивших “когнитивные” пробы данного теста с лазом, закрытым пробкой, представляется “парадоксальной”, и этот факт показывает, что когнитивные способности этих животных не пострадали после облучения протонами. Это находится в соответствии с отсутствием негативного влияния этого воздействия на поведение животных в целом в течение нескольких месяцев после облучения.
Успешное выполнение мышами теста на ПВУ требует от животного оперирования свойством “неисчезаемости” (по Ж. Пиаже, [8]), т.е. “понимания”, что воспринимавшийся объект, которого более не видно, продолжает существовать, и его можно искать. Заимствованное из психологии понятие “исполнительных функций” (executive functions) включает в себя, наряду с другими способностями, и решение элементарных логических задач, для которых у животного “нет готового решения” (по А.Р. Лурия). В наших экспериментах оказалось, что облучение протонами вызывало у животных статистически значимое улучшение решения задачи на поиск входа в укрытие при сложной маскировке лаза с помощью пробки, выразившееся в более высокой доле мышей, которые смогли решить эти пробы теста. Данный эффект имел долговременный характер, поскольку обнаружился в тесте, который мышам предъявляли через 1, 2 и 5.5 мес. после облучения (в двух независимых сериях опытов). Одним из возможных объяснений этого эффекта может быть предположение о некоторой активации ЦНС мышей, вызванной облучением протонами. Сходный результат – отсутствие негативного влияния облучения протонами и изменение поведения животных в эмоциональном (но не когнитивном) поведении был получен другими авторами ранее [3, 5]. Результаты тестирования исследовательского поведения мышей в тестах “малое открытое поле”, приподнятый крестообразный лабиринт и др. показали некоторое усиление исследовательского поведения облученных мышей (данные не приводятся), что согласуется с предположением о некоторой активации их поведения. В то же время наши данные выявили достоверно менее успешное решение этого же теста на тех его этапах, когда от мыши для перехода в укрытие требовалось раскопать стружку, маскирующую лаз. Очевидно, что причины этого нарушения поведения “раскапывания” нуждаются в специальном анализе.
Данные по количеству новых клеток в зубчатой фасции гиппокампа (FDH) и в субвентрикулярной зоне переднего мозга (SVZ) после облучения и у контрольных мышей того же возраста представлены на рис. 3. У мышей облученной группы (доза 2 и 4 Гр) число клеток иммунопозитивных к Ki-67 (маркер новых клеток) в переднем мозге (FDH и SVZ) было меньше, чем в контроле. Более существенное подавление пролиферации обнаружено в SVZ. Отметим, что межсамцовая агрессия (связанная с обонятельным восприятием) у облученных мышей была нарушена (данные не приводятся).
Рис. 3.
Среднее число (± ош. средн.) новых клеток в пролиферативных зонах переднего мозга (ось ординат) в контроле (1) и после облучения протонами (2 – 1 Гр, 3 – 2 Гр и 4 – 4 Гр, по данным для 4- и 6-месячных сроков после облучения). а – субгранулярная зона (зубчатой фасции) гиппокампа, б – субвентрикулярная зона переднего мозга. * Достоверно (p < 0.05) отличается от показателей для контрольных животных того же возраста.
Fig. 3. The mean number (± st. error, ordinate) of new cells in the forebrain proliferative zones in control mice (1) and after proton irradiation (1 Gy – 2, 2 Gy – 3, 4 Gy – 4) for the 4 m and 6 m periods after the irradiation. a – subgranular area of hippocampal fascia dentata, b – subventricular forebrain area. * Significantly different (p < 0.05) from the scores of control animals of the same age.

Интенсивность нейрогенеза взрослого мозга у лабораторных грызунов и показатели успешности выполнения когнитивных тестов, как показывает сопоставление результатов большого числа исследований [13], коррелируют далеко не во всех случаях, поэтому полученные результаты демонстрируют сложность исследуемых процессов как последствий ионизирующего облучения.
Таким образом, проведенные эксперименты показали отсутствие повреждающего влияния протонного облучения в использованных условиях на поведение мышей в “когнитивном” тесте (даже с превосходством “облученных” мышей в решении “трудного” когнитивного этапа с пробкой в тесте на поиск входа в укрытие), а также постлучевое снижение нейрогенеза в переднем мозге мышей, более четкое при дозах 2 и 4 Гр.
Список литературы
Григорьев А.И., Красавин Е.А., Островский М.А. К вопросу о радиационном барьере при пилотируемых межпланетных полетах // Вест. РАН. 2017. Т. 87. № 1. С. 65–69. [Grigoriev A.I., Krasavin E.A., Ostrovsky M.A. On the problem of radiation barrier during manned space flights // Vestn. Ac. Nauk. V. 87. № 1. P. 65–69. (In Russian)].https://doi.org/10.7868/S0869587317010030
Shukitt-Hale B., Szprengiel A., Pluhar J. et al. The effects of proton exposure on neurochemistry and beha-vior // Adv. Space Res. 2004. V. 33. № 8. P. 1334–1339.
Штемберг А.С., Базян А.С., Лебедева-Георгиевская К.Б. и др. Влияние высокоэнергетических протонов на поведение крыс и его нейрохимические механизмы // Авиакосм. и экол. мед. 2013. Т. 47. № 6. С. 54–60. [Shtemberg A.S., Basian A.I., Lebedeva-Georgievskaya K.B. et al. The influence of high energy protons on rat behavior and its neurochemical mechanisms // Avaicosm. a ecol. med. 2004. V. 47. № 6. P. 54–60 (In Russian)].
Davis C.M., Davis C.M., DeCicco-Skinner K.L., Roma P.G. et al. Individual differences in attentional deficits and dopaminergic protein levels following exposure to proton radiation // Radiat. Res. 2014, 181. № 3. P. 258–271. https://doi.org/10.1667/RR13359.1
Bellone J.A., Rudobeck E., Hartman R.E. et al. A single low dose of proton radiation induces long-term beha-vioral and electrophysiological changes in mice // Radiat. Res. 2015. V. 184. № 2. P. 193–202.
Rabin B.M., Shukitt-Hale B., Joseph J.A. et al. Relative effectiveness of different particles and energies in disrupting behavioral performance // Radiat. Environ. Biophys. 2007. V. 46. № 2. P. 173–177.
Перепелкина О.В., Голибродо В.А., Лильп И. Г. и др. Селекция мышей на высокие показатели решения элементарной логической задачи // Докл. акад. наук. 2015. Т. 460. № 5. С. 617–621. [Perepelkina O.V., Golibrodo V.A., Lilp I.G. et al. Selection of mice for high scores of elementary logic task solution // Dokl. Acad. Nauk. 2015. V. 460. № 5. P. 617–621. (In Russian)].
Zucca P., Milos N., Vallortigara G. Piagetian object permanence and its development in Eurasian jays (Garrulus glandarius) // Anim. Cogn. 2007. V. 10. № 2. P. 243–258.
Galsworthy M.J., Paya-Cano J.L., Liu L. et al. Assessing reliability, heritability and general cognitive ability in a battery of cognitive tasks for laboratory mice // Behav. Genet. 2005. V. 35. № 5. P. 675–692.
Ben Abdallah N.M., Fuss J., Trusel M. et al. The puzzle box as a simple and efficient behavioral test for explo-ring impairments of general cognition and executive functions in mouse models of schizophrenia // Exp. Neurol. 2011. V. 227. № 1. P. 42–52. https://doi.org/10.1016/j.expneurol.2010.09.008
Тимошенко Т.В., Перепелкина О.В., Маркина Н.В. и др. Аудиогенная эпилепсия у крыс двух генотипов после неонатальных воздействий, усиливающих нейрогенез в зубчатой извилине // Бюл. эксперим. биологии и медицины. 2009. Т. 147. № 4. С. 441–444. [Timoshenko T.V., Perepelkina O.V., Markina N.V., et al. Audiogenic epilepsy in rats of two genotypes after neonatal treatments, which increase the neurogenesis in the dentate gyrus // Bull. exper. biol. med. 2009. V. 147. № 4. P. 441–444. (In Russian)]
Sweet T.B., Panda N., Hein A.M. et al. Central nervous system effects of whole-body proton irradiation // Radiat. Res. 2014. V. 182. № 1. P. 18–34. https://doi.org/10.1667/RR13699
Lipp H.-P. Evolutionary shaping of adult hippocampal neurogenesis in mammals–cognitive gain or developmental priming of personality traits? // Front. Neuro-sci. 2017. V. 21. https://doi.org/10.3389/fnins.2017.00420
Дополнительные материалы отсутствуют.
Инструменты
Радиационная биология. Радиоэкология