

Растительные ресурсы, 2019, T. 55, № 1, стр. 56-71
Поливариантность развития особей Picea obovata (Pinaceae) в средневозрастных сосново-еловых лесах Кольского полуострова
А. Е. Мишко 1, *, Н. И. Ставрова 2, В. В. Горшков 2, 3
1 Северо-Кавказский федеральный научный центр садоводства, виноградарства, виноделия
г. Краснодар, Россия
2 Ботанический институт им. В.Л. Комарова РАН
г. Санкт-Петербург, Россия
3 Санкт-Петербургский государственный лесотехнический университет им. С.М. Кирова
г. Санкт-Петербург, Россия
* E-mail: mishko-alisa@mail.ru
Поступила в редакцию 14.08.2018
После доработки 17.09.2018
Принята к публикации 25.09.2018
Аннотация
Проанализировано размерное, онтогенетическое и виталитетное разнообразие особей первого послепожарного поколения Picea obovata L. в северотаежном елово-сосново-березовом кустарничково-зеленомошном сообществе с давностью пожара 85 лет (Кольский п-ов). Выявлена повышенная степень вариабельности большинства исследованных параметров (CV = 40–55%). Показано, что крайние значения высоты, диаметра ствола, параметров кроны, текущего прироста в пределах всего поколения различаются в 8–10 раз, у одновозрастных особей – в 3–3.5 раза. Поколение отличается высоким онтогенетическим разнообразием. Особи ели сибирской могут находиться на пяти разных стадиях развития – от im3 до g1; большинство из них (80%) относится к категориям умеренно и сильно ослабленных. Установлено, что возрастными различиями обусловлено в среднем около 50% варьирования высоты и диаметра, около 35% варьирования текущего прироста, примерно 30% онтогенетического разнообразия и не более 15% виталитетных различий особей. Сделан вывод, что дифференциация особей ели сибирской в составе первого послепожарного поколения в наибольшей степени обусловлена высокой контрастностью локальных участков сообщества по степени напряженности корневой конкуренции.
Полный жизненный цикл древесного растения в зависимости от условий произрастания претерпевает различные изменения. В одних случаях, обусловленных как внешними, так и внутренними факторами, он ускоряется или замедляется; в других приобретает какие-либо новые качественные характеристики. Данное явление получило название поливариатности онтогенеза [1]. Выделяют несколько основных типов поливариатности: структурную, динамическую, функциональную и экологическую [2–5]. Чаще всего в работах, посвященных вопросам изучения поливариантности онтогенеза растений, затрагивают структурный и динамический типы. Первый включает размерную, морфологическую, виталитетную дифференциацию особей, а также разделение растений по способам размножения. Второй подразделяется на временной подтип и ритмологический.
Как показали результаты наших предыдущих исследований, для ценопопуляций ели сибирской в северотаежных сосново-еловых лесах характерно существенное расслоение особей одного возраста по размерным показателям, скорости роста, жизненному состоянию и стадиям развития [6].
Цель настоящей работы заключается в изучении поливариантности роста и развития особей ели сибирской первого послепожарного поколения на одной из начальных стадий восстановления северотаежных кустарничково-зеленомошных сосново-еловых лесов после катастрофических пожаров.
МАТЕРИАЛ И МЕТОДЫ
Исследования проведены на территории Кольского п-ва в районе среднего течения р. Лива (67°49′26″ с.ш., 31°19′12″ в.д.) в елово-сосново-березовом сообществе, сформировавшемся через 85 лет после пожара в условиях местообитания, характерных для кустарничково-зеленомошных сосново-еловых лесов (Pineto–Piceetum empetroso–myrtilloso–hylocomiosum), которые большинством исследователей рассматриваются как коренной тип леса, характерный для европейской северной тайги [7–10].
В изученном сообществе древесный ярус сложен тремя видами – елью сибирской Picea obovata Ledeb., сосной обыкновенной Pinus sylvestris L. и березой пушистой Betula pubescens Ehrh. На рассматриваемой стадии послепожарной сукцессии в древесном ярусе по числу особей доминирует береза пушистая (1120 экз./га), участие ели и сосны составляет соответственно 640 и 128 экз./га. По сумме площадей сечения около 42% составляет участие березы, 36% – сосны и 22% – ели. Наиболее высокими значениями средней высоты и диаметра на высоте 1.3 м отличается сосновый компонент древесного яруса – соответственно 13.9 м и 22 см, высота и диаметр деревьев березы пушистой составляют соответственно 7.9 м и 8.0 см, наиболее низкими параметрами высоты и диаметра характеризуется еловый древостой – 6.8 м и 7.7 см. Средний возраст деревьев березы пушистой, сосны обыкновенной и ели сибирской в древесном ярусе составляет соответственно 80, 76 и 62 г. Сквозистость древесного яруса составляет в среднем 60% (14–94%). Общая численность живых особей в ценопопуляции ели – 2096 экз./га, более половины из них входит в полог подроста (крупный подрост – 448 экз./га, мелкий – 1008 экз./га).
Доминантами травяно-кустарничкового яруса являются Vaccinium myrtillus L., V. vitis-idaea L., Empetrum hermaphroditum Hagerup. при общем проективном покрытии 30–40%. В мохово-лишайниковом ярусе преобладает Pleurozium schreberi (Brid.) Mitt, проективное покрытие которого составляет 80%.
Интенсивность последнего пожара в исследованном сообществе была высокой, о чем свидетельствует отсутствие на этом участке допожарного компонента (т.е. деревьев, переживших пожар) в составе сосновой части древостоя. Всходы ели, появлявшиеся в первые годы после пожара, по-видимому, погибали из-за высоких перепадов температур на открытой гари. Наиболее благоприятные условия для прорастания семян, выживания всходов и роста сеянцев ели сложились после образования полога из крупного подроста березы (~10–15 лет после пожара). Этот период продолжался до формирования относительно сомкнутого напочвенного покрова (~40–60 лет после пожара), который приводит к “зависанию” семян и угнетает развитие всходов. Еловый компонент древостоя сформировался из особей, заселившихся в течение 22-летнего интервала в период от 13 до 35 лет после пожара.
В работе проанализированы параметры первого послепожарного поколения ели сибирской, особи которого имели на момент исследования возраст от 50 до 72 лет. Мы рассматриваем объект исследования как подсистему ценопопуляции этого вида [11]. В выборку включены 92 модельные особи. У каждой особи ели устанавливали возраст у основания ствола (у 55% особей – по кернам, у остальных особей – по калибровочной кривой, связывающей возраст с величиной диаметра ствола). Стадии онтогенеза определялись в соответствии с общепринятой классификацией [1, 12–14] и результатами наших исследований онтогенеза ели сибирской [15].
Жизненное состояние особей ели определяли на основе интегрального параметра – плотности кроны [16–19] с учетом совокупности признаков, характеризующих ее форму и относительные размеры, степень разветвленности, долю сухих ветвей, состояние (наличие и степень проявления хлороза и некроза) и продолжительность жизни хвои. Живые особи подразделялись на четыре группы: I – здоровые (неугнетенные), II – умеренно ослабленные (умеренно угнетенные), III – сильно ослабленные (сильно угнетенные), IV – усыхающие.
Комплекс определяемых морфометрических показателей включал общую высоту, диаметр основания ствола, диаметр ствола на высоте 1.3 м, диаметр и протяженность кроны, верхушечный прирост главной и боковой оси (за последние 5 лет).
Обработка данных проводилась на основе методов описательной статистики. Оценка значений коэффициента вариации (CV) параметров осуществлялась по модифицированной шкале С.А. Мамаева [20]: значения, близкие к величине коэффициента вариации нормального распределения (33%), рассматривались как средние (25–39%), величины от 40 до 63% – как повышенные, более 63% – как высокие и очень высокие. Для оценки связи между средними значениями параметров особей использован корреляционный и линейный регрессионный анализ. Аппроксимация эмпирических распределений теоретическими проводилась на основе нормального распределения, гамма-распределения и распределения Вейбулла с 2 и 3 параметрами с оценкой значимости по критерию хи-квадрат.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
В ходе изучения параметров особей ели сибирской первого послепожарного поколения было выявлено, что они находятся на пяти различных онтогенетических стадиях. Большинство из них (70%) относится к прегенеративному периоду. Распределение особей ели по онтогенетическим стадиям является бимодальным: максимальная встречаемость (30 и 29%) характерна для ${{\text{v}}_{1}}$ и g1, встречаемость im3 и v3 (10%) является наиболее низкой (рис. 1).
Рис. 1.
Онтогенетическая структура первого послепожарного поколения Picea obovata в северотаежном кустарничково-зеленомошном елово-сосново-березовом сообществе с давностью пожара 85 лет. По горизонтали – стадии онтогенеза: im3 – поздняя стадия имматурного состояния, ${{v}_{1}}$ – начальная стадия виргинильного состояния, ${{v}_{2}}$ – промежуточная стадия виргинильного состояния, ${{v}_{3}}$ – поздняя стадия виргинильного состояния, g1 – начальная стадия генеративного состояния; по вертикали – доля участия, %. Fig. 1. Ontogenetic structure of the first post-fire generation of Picea obovata in the northern taiga Siberian spruce-Scots pine-white birch dwarf shrub-green moss community 85 years post-fire. X-axis – ontogenetic stages: im3 – late sub-stage of the immature stage, ${{v}_{1}}$ – initial sub-stage of the virginile stage, ${{v}_{2}}$ – intermediate sub-stage of the virginile stage, ${{v}_{3}}$ – late sub-stage of the virginile stage, g1 – initial sub-stage of the generative stage; y-axis – participation, %.

Анализ виталитетной структуры показал, что только 3% особей ели сибирской первого послепожарного поколения относятся к категории здоровых; преобладают умеренно и сильно ослабленные (33 и 50%); доля усыхающих составляет 14% (рис. 2).
Рис. 2.
Виталитетная структура первого послепожарного поколения Picea obovata в северотаежном кустарничково-зеленомошном елово-сосново-березовом сообществе с давностью пожара 85 лет. По горизонтали – категории жизненного состояния: I – здоровые (неугнетенные) особи, II – умеренно ослабленные (умеренно угнетенные), III – сильно ослабленные (сильно угнетенные), IV – усыхающие; по вертикали – доля участия, %. Fig. 2. Vitality structure of the first post-fire generation of Picea obovata in the northern taiga Siberian spruce-Scots pine-white birch dwarf shrub-green moss community 85 years post-fire. X-axis – vitality classes: I – healthy (not oppressed) individuals, II – moderately weakened (moderately oppressed), III – severely weakened (severely oppressed), IV – dying out; y-axis – participation, %.

Величина диаметра основания ствола особей ели варьирует от 1.7 до 14.5 см (табл. 1). Коэффициент вариации отличается повышенным значением (CV = 41%). Распределение симметричное, плосковершинное (коэффициент асимметрии примерно равен 0, коэффициент эксцесса имеет значительную отрицательную величину) и имеет два довольно хорошо выраженных максимума частот в области значений 4–5 и 8–10 см (рис. 3а). Аппроксимация этого распределения теоретическими дает достоверные результаты, но точность ее невысока и примерно одинакова для нормального распределения и распределения Вейбулла (табл. 2).
Таблица 1.
Параметры распределений морфометрических характеристик особей Picea obovata (первое послепожарное поколение) в северотаежном елово-сосново-березовом сообществе с давностью пожара 85 лет Table 1. Distribution of morphometric parameters of Picea obovata individuals (first post-fire generation) in the northern taiga Siberian spruce-Scots pine-white birch community 85 years post-fire
| Параметр Parameter |
N | M | S | CV | Min | Max | A | E |
|---|---|---|---|---|---|---|---|---|
| Высота, м Height, m |
92 | 4.4 | 1.9 | 44.2 | 1 | 8.4 | 0.02 | –0.99 |
| Диаметр основания ствола, см Basal diameter, cm |
92 | 7.7 | 3.1 | 40.8 | 1.7 | 14.5 | 0.02 | –0.81 |
| Диаметр ствола на высоте 1.3 м, см Breath height diameter, cm |
86 | 5.3 | 2.3 | 43.7 | 0.8 | 10.2 | –0.05 | –0.91 |
| Протяженность кроны, м Crown length, m |
91 | 3.4 | 1.8 | 53.3 | 0.3 | 7.0 | 0.14 | –1.04 |
| Диаметр кроны, см Crown diameter, cm |
91 | 1.5 | 0.5 | 32.7 | 0.3 | 2.9 | –0.14 | 0.0 |
| Текущий линейный прирост
ствола, см/год Current linear trunk increment, cm/year |
42 | 12.6 | 6.9 | 55.3 | 2.7 | 30.5 | 0.39 | –0.53 |
| Текущий линейный прирост
боковых ветвей, см/год Current linear increment of the lateral branches, cm/year |
37 | 6.0 | 2.6 | 43.4 | 1.7 | 11.7 | 0.49 | –0.49 |
Примечание. N – объем выборки, M – среднее значение, S – стандартное отклонение, CV – коэффициент вариации, Min – минимальное значение, Max – максимальное значение, A – коэффициент асимметрии, E – коэффициент эксцесса. Note. N – sample size, M – average, S – standard deviation, CV – coefficient of variation, Min – minimum, Max – maximum, A – skewness, E – kurtosis.
Рис. 3.
Распределение особей первого послепожарного поколения Picea obovata по диаметру основания ствола (а), диаметру на высоте 1.3 м (b) и высоте (c) в северотаежном кустарничково-зеленомошном елово-сосново-березовом сообществе с давностью пожара 85 лет. По горизонтали – градации диаметра, см (а, b) и высоты, м (c); по вертикали – частота, %. Fig. 3. Distribution of the first post-fire generation of Picea obovata by basal diameter (a), breath height diameter (b) and height (c) in the northern taiga Siberian spruce-Scots pine-white birch dwarf shrub-green moss community 85 years post-fire. X-axis – gradation of diameter, cm (a, b) and height, m (c); y-axis – frequency, %.

Таблица 2.
Результаты аппроксимации распределений параметров особей Picea obovata теоретическими распределениями Table 2. The results of approximation of distributions parameters of Picea obovata individuals by theoretical distributions
| Параметр Parameter |
Нормальное
распределение Normal distribution |
Гамма-распределение Gamma distribution |
Распределение Вейбулла Weibull distribution |
Распределение Вейбулла с 3 параметрами 3-parameter Weibull distribution |
|---|---|---|---|---|
| Диаметр основания ствола, см Basal diameter, cm |
$\frac{{7.7;\,\,3.13*}}{{0.09}}$ | $\frac{{5.03;\,\,0.66}}{{0.08}}$ | $\frac{{2.71;\,\,8.64}}{{0.20}}$ | $\frac{{2.80;\,\,8.87;\,\, - 0.22}}{{0.16}}$ |
| Диаметр ствола на высоте 1.3 м, см Breath height diameter, cm |
$\frac{{5.3;\,\,2.30}}{{0.15}}$ | – | $\frac{{2.50;\,\,5.94}}{{0.28}}$ | $\frac{{2.96;\,\,6.80;\,\, - 0.79}}{{0.20}}$ |
| Высота ствола, м Height, m |
$\frac{{4.4;\,\,1.94}}{{0.11}}$ | – | $\frac{{2.47;\,\,4.96}}{{0.11}}$ | $\frac{{2.50;\,\,5.01;\,\, - 0.05}}{{0.06}}$ |
| Протяженность кроны, м Crown length, cm |
– | – | – | – |
| Диаметр кроны, см Crown diameter, m |
$\frac{{1.48;\,\,0.48}}{{0.67}}$ | $\frac{{7.58;\,\,5.12}}{{0.06}}$ | $\frac{{3.43;\,\,1.65}}{{0.63}}$ | $\frac{{3.99;\,\,1.89;\,\, - 0.23}}{{0.37}}$ |
| Текущий линейный прирост ствола, см/год Current linear trunk increment, cm/year |
– | – | – | – |
| Текущий линейный прирост боковых
ветвей, см/год Current linear increment of lateral branches, cm/year |
$\frac{{5.96;\,\,2.59}}{{0.14}}$ | $\frac{{5.20;\,\,0.87}}{{0.30}}$ | $\frac{{2.52;\,\,6.74}}{{0.21}}$ | $\frac{{1.87;\,\,5.22;\,\,1.33}}{{0.18}}$ |
Примечание. *Над чертой – коэффициенты распределений: для нормального распределения – среднее значение и стандартное отклонение; для распределений гамма и Вейбулла – коэффициент формы и коэффициент масштаба; для распределения Вейбулла с тремя параметрами – коэффициент формы, коэффициент масштаба, сдвиг. Под чертой – уровень значимости (при P > 0.05 подтверждается гипотеза о соответствии распределения теоретическому распределению на 95%-ном доверительном уровне). Note. *Above the line – distribution coefficients: for the normal distribution – the mean and standard deviation; for gamma and Weibull distributions – shape and scale parameters; for the 3-parameter Weibull distribution – the shape, scale and threshold parameters. Below the line – significance level (at P > 0.05 hypothesis that the distribution is consistent with the theoretical distribution is confirmed at 95% confidence level).
При анализе диаметра ствола на высоте 1.3 м установлено, что его диапазон изменяется от 0.7 до 10.2 см при среднем значении равном 5.3 см (табл. 1). Этот параметр, так же как диаметр основания ствола, имеет повышенную вариабельность (CV = 44%). Распределение симметричное, выраженно плосковершинное (табл. 1; рис. 3б): в основной части диапазона значений (от 2 до 9 см) частоты практически одинаковы – 8–13%. Рассматриваемое распределение аппроксимируется нормальным распределением и распределением Вейбулла (табл. 2).
Высота ели сибирской в составе первого послепожарного поколения находится в пределах от 1 до 8 м (табл. 1) и характеризуется повышенной вариабельностью (CV = 44%). Распределение особей по градациям высоты отличается симметричностью и плосковершинностью (табл. 1); особи высотой от 2 до 7 м имеют очень близкую долю участия, составляющую от 12 до 17% (рис. 3c). Распределение высот может быть достоверно аппроксимировано нормальным распределением и распределением Вейбулла (табл. 2).
Протяженность кроны особей ели первого послепожарного поколения составляет от 0.3 до 7.0 м и отличается повышенным уровнем изменчивости (CV = 53%). Распределение близко к симметричному, плосковершинное (табл. 1): примерно половина всех особей имеет протяженность кроны от 1 до 4 м (рис. 4а). Распределение не аппроксимируется ни одним из теоретических распределений (табл. 2).
Рис. 4.
Распределение особей первого послепожарного поколения Picea obovata по протяженности (а) и диаметру (b) кроны в северотаежном кустарничково-зеленомошном елово-сосново-березовом сообществе с давностью пожара 85 лет. По горизонтали – градации протяженности и диаметра кроны, м; по вертикали – частота, %. Fig. 4. Distribution of the first post-fire generation of Picea obovata by crown length (a) and diameter (b) in the northern taiga Siberian spruce-Scots pine-white birch dwarf shrub-green moss community 85 years post-fire. X-axis – gradation of crown length and diameter, m; y-axis – frequency, %.

У такого параметра как диаметр кроны диапазон значений составляет от 0.3 до 2.9 м при среднем значении 1.5 м (табл. 1) и среднем уровне варьирования (CV = 33%). Кривая распределения (рис. 4b) относительно симметричная и имеет близкую к колоколообразной форму (коэффициент эксцесса равен 0). Распределение хорошо аппроксимируется нормальным распределением и распределениями Вейбулла с двумя и тремя параметрами (табл. 2).
Текущий прирост главной оси у особей ели сибирской первого послепожарного поколения в среднем составляет 12.6 см/год при диапазоне от 2.7 до 30.5 см/год (табл. 1) и отличается повышенным уровнем вариабельности (CV = 55%). Распределение значений этого показателя имеет умеренную положительную асимметрию и хорошо выраженную бимодальность (рис. 5а). Два максимума частот приходятся на величины прироста 6–9 и 15–18 см/год. Распределение не аппроксимируется ни одним из теоретических распределений (табл. 2).
Рис. 5.
Распределение особей первого послепожарного поколения Picea obovata по величине текущего прироста ствола (а) и боковых ветвей (b) в северотаежном кустарничково-зеленомошном елово-сосново-березовом сообществе с давностью пожара 85 лет. По горизонтали – градации прироста, см; по вертикали – частота, %. Fig. 5. Distribution of the first post-fire generation of Picea obovata by the value of current increment of the trunk (а) and lateral branches (b) in the northern taiga Siberian spruce-Scots pine-white birch dwarf shrub-green moss community 85 years post-fire. X-axis – gradation of increment, cm; y-axis – frequency, %.
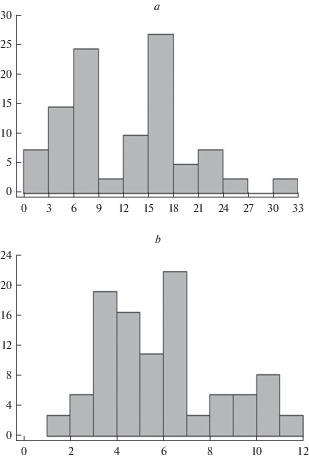
Текущий прирост боковой оси II порядка у особей первого послепожарного поколения ели составляет от 1.7 до 11.7 см/год при среднем значении 6 см (табл. 1), т.е. он примерно в 2 раза ниже прироста главной оси и имеет более низкую вариабельность (43%). Так же как распределение величин прироста главной оси, распределение этого параметра является слабо положительно асимметричным (рис. 5b). Наиболее часто встречающиеся значения прироста лежат в области от 3 до 7 см/год (70% всех особей). Распределение с примерно одинаковой точностью может быть аппроксимировано всеми использованными для анализа теоретическими распределениями (табл. 2).
Таким образом, в изученном сообществе распределения особей ели сибирской первого послепожарного поколения по основным морфометрическим и ростовым параметрам симметричны или слабо положительно асимметричны, часто плосковершинны и в большинстве случаев могут быть с удовлетворительной точностью описаны теоретическими двускатными кривыми, среди которых наибольшей близостью к эмпирическим данным и наибольшей универсальностью отличаются кривые распределения Вейбулла.
Морфометрические параметры особей характеризуются довольно тесной линейной связью. Наиболее высокая степень взаимной положительной корреляции (r = 0.93–0.97; R2 = 0.86–0.94; P < 0.001) характерна для общей высоты, диаметра основания ствола, диаметра на высоте 1.3 м и протяженности кроны. Морфометрические параметры примерно в равной мере связаны с текущим приростом (r = 0.72–0.82; R2 = 0.51–0.66; P < 0.001) и возрастом особей (r = 0.69–0.74; R2 = 0.47–0.55; P < 0.001). Заметно более слабой является их связь с уровнем жизненного состояния (r = –0.37–0.52; R2 = 0.13–0.27; P < 0.05) (отрицательное значение коэффициента обусловлено тем, что более высокий уровень жизненного состояния соответствует более низкому ранговому значению). Связь текущего прироста с возрастом (r = 0.56–0.60; R2 = 0.32–0.36; P < 0.001) и уровнем жизненного состояния особи (r = –0.37–0.41; R2 = 0.13–0.16; P < 0.05) заметно слабее, чем с морфометрическими параметрами. Уровень жизненного состояния особей ели достоверно, но очень слабо связан с календарным возрастом (r = 0.34; R2 = 0.11; P = 0.001) и стадией развития (r = –0.38; R2 = 0.16; P < 0.001).
Варьирование особей по размерным параметрам и скорости роста зависит от двух основных факторов; возраста (времени заселения) и условий произрастания. П.В. Воропановым [21] было отмечено разнообразие вариантов начальной и конечной скорости роста особей в составе северотаежных древостоев. Основной причиной этих различий на ранних этапах послепожарного восстановления является разнообразие микроместообитаний на гари, обусловленное неравномерным выгоранием напочвенного покрова и подстилки, что приводит в свою очередь к высокой вариабельности толщины, водоудерживающих и химических свойств субстрата [22, 23]. В дальнейшем на первый план выступает формирующаяся неоднородность ценотической среды и уровень напряженности корневой конкуренции [24–26]. Освещенность в условиях района исследований не является лимитирующим фактором. Доступность света создает возможность длительного выживания особей не только в благоприятных для роста условиях, но и при высокой напряженности корневой конкуренции и низкой скорости роста. Это приводит к формированию размерной структуры, отличающейся примерно одинаковой долей участия особей с разными морфометрическими параметрами. В то же время следует отметить и проявление тенденции к упорядочению размерной структуры, проявляющейся в бимодальности распределения текущего прироста. В дальнейшем это должно привести к формированию в составе поколения размерно и функционально обособленных групп особей. Такого рода процессы характерны как для отдельных поколений древесных растений, так и для древостоев в целом и являются результатом конкуренции на ресурсы среды [27–31].
Для исследованного поколения ели сибирской характерно не только размерное, но и онтогенетическое разнообразие: особи одного календарного возраста могут находиться на четырех-пяти разных стадиях развития: от im3 до g1. Судя по онтогенетическому спектру высокая поливариантность развития особей, имеющих узкий диапазон календарного возраста (не более 20–25 лет), отмечалась в разных регионах для разных видов хвойных: ели аянской [32], пихты белокорой [33], кедра сибирского [34]. Факт отсутствия прямой (точнее тесной) связи между календарным возрастом и стадией развития конкретных особей неоднократно отмечался в литературе [2, 13]. Результаты выполненного анализа показывают, что в среднем, даже в пределах 20-летнего поколения, чем больше возраст особи (т.е. чем раньше она заселилась после пожара), тем выше вероятность ее более быстрого развития. Однако связь онтогенетического состояния и календарного возраста является относительно слабой: в исследованном сообществе стадия развития особей первого послепожарного поколения лишь на ~30% обусловлена их возрастом.
При высоком онтогенетическом разнообразии первое послепожарное поколение ели относительно однородно по уровню жизненного состояния особей: абсолютное большинство из них (~80%) относится к категориям умеренно и сильно ослабленных, причем половину составляют сильно ослабленные, здоровые особи встречаются редко. Этот результат согласуется с данными многих исследователей, согласно которым особи высокой жизненности развиваются в основном на открытых или полуоткрытых пространствах в условиях высокого уровня освещенности и низкого уровня конкуренции [33, 35, 36]. По данным С.А. Николаевой с соавторами [34], в составе первого послепожарного поколения хвойных, даже при одновременном с лиственными заселением, представлены особи как высокой, так и пониженной и низкой жизненности. В изученном сообществе уровень жизненного состояния особей ели на 10–15% определяется их календарным возрастом и положением в онтогенетическом ряду: те из них, которые появились раньше и достигли более высокой стадии развития, имеют в среднем несколько лучшее жизненное состояние.
Следует отметить, что, по данным И.Д. Махаткова [35], которые подтверждают сделанный ранее вывод Г.Г. Левина [37], оптимальным путем развития особей древесных растений является развитие на среднем уровне жизненности (примерно соответствует умеренно ослабленным особям и, возможно, части сильно ослабленных особей в настоящем исследовании). Они формируются в условиях “умеренного фитоценотического пресса”, при котором растение способно пройти полный жизненный цикл, активно мобилизуя при этом свой биологический потенциал. Эти особи наиболее жизнестойки и составляют основу господствующего компонента ценопопуляций. Трудно не согласиться с этой точкой зрения, учитывая результаты исследований по оценке виталитетной структуры древостоев лесообразующих видов в малонарушенных темнохвойных лесах [38–40].
ЗАКЛЮЧЕНИЕ
В северотаежном елово-сосново-березовом сообществе с давностью пожара 85 лет первое послепожарное поколение ели сибирской Picea obovata Ledeb., состоящее из особей в возрасте 50–70 лет, отличается значительной гетерогенностью морфометрических параметров и скорости роста. Крайние значения высоты, диаметра ствола, параметров кроны, текущего прироста в пределах всего поколения различаются в 8–10 раз, у одновозрастных особей – в 3–3.5 раза. На рассматриваемой стадии сукцессии для изученного поколения характерна, с одной стороны, относительная выравненность большинства размерных распределений, т.е. близкая доля участия особей с разными размерными параметрами, с другой – начало процесса упорядочения структуры, формирования размерно (а, следовательно, и функционально) обособленных групп. Как показало выполненное исследование, возрастными различиями обусловлено в среднем около 50% варьирования высоты и диаметра и только 35% варьирования текущего линейного прироста. Это свидетельствует о том, что роль возрастного фактора в размерной дифференциации особей, еще довольно существенная на рассматриваемой стадии сукцессии, с течением времени будет снижаться.
Параметры роста и развития тесно связаны между собой, поэтому различиям в скорости роста сопутствует выраженная поливариантность онтогенеза особей первого послепожарного поколения: на рассматриваемом этапе сукцессии они находятся на пяти разных стадиях развития – от поздней имматурной до начальной генеративной. По доле участия выделяются ${{\text{v}}_{1}}$ и g1 особи (60% всех особей). Выявленное онтогенетическое разнообразие лишь на 30% определяется возрастными различиями.
При значительном размерном и онтогенетическом разнообразии исследованному поколению присуща относительная упорядоченность виталитетной структуры. В его составе представлены особи всех категорий жизненного состояния – от здоровых (неугнетенных) до усыхающих, однако подавляющее большинство из них (80%) относится к двум категориям – умеренно и сильно ослабленных. Варьирование жизненного состояния особей ели в очень слабой степени (на 10–15%) обусловлено их возрастом и стадией развития.
Результаты выполненного исследования позволяют заключить, что роль возрастных различий (20 лет) особей первого послепожарного поколения ели сибирской в их размерной, онтогенетической и виталитетной дифференциации в процессе сукцессии снижается. Правомерно предположить, что основным фактором становится неоднородность ценотической среды и прежде всего различие локальных участков сообщества по степени напряженности корневой конкуренции. Через 85 лет после пожара медленно изменяющиеся параметры (размерные) еще сохраняют относительно высокую степень зависимости от фактора возраста, в то время как более лабильные (текущий прирост, стадия развития, жизненное состояние) в значительной мере ее утрачивают.
Список литературы
Ценопопуляции растений (основные понятия и структура). 1976. М. 216 с.
Smirnova O.V., Chistyakova A.A., Zaugolnova L.B., Evstigneev O.I., Popadiouk R.V., Romanovsky A.M. 1999. Ontogeny of a tree. – Бот. журн. 84 (12): 8–19.
Поливариантность развития организмов, популяций и сообществ. 2006. Йошкар-Ола. 326 с.
Жукова Л.А. 2008. Роль популяционно-онтогенетического направления в сохранении биоразнообразия растений. – В сб.: Принципы и способы сохранения биоразнообразия: Материалы III Всерос. науч. конф. Марийский гос. ун-т. Йошкар-Ола; Пущино. С. 22–23. https://istina.msu.ru/media/publications/article/a5f/067/26761635/Zhukova_LA_2008_01.pdf
Жукова Л.А., Османова Г.О., Ведерникова О.П., Козырева С.В. 2011. Развитие идей А.А. Уранова в работе популяционно-онтогенетического музея Марийского государственного университета. – В сб.: Современные проблемы популяционной экологии, геоботаники, систематики и флористики: Материалы междунар. науч. конф., посвящ. 110-летию А.А. Уранова. Кострома. Т. 1. С. 99–103. http://csl.isc.irk.ru/BD/Books/Пробл%20экология%20т1.pdf
Мишко А.Е., Ставрова Н.И., Горшков В.В. 2018. Онтогенетическая структура ценопопуляций Picea obovata на разных стадиях послепожарных сукцессий в северотаежных сосново-еловых лесах. – Бот. журн. 103(9.): 50–78.
Некрасова Т.П. 1961. Взаимоотношения сосны и ели в лесах Кольского полуострова. – В кн.: Леса Кольского полуострова и их возобновление. М. С. 63–70.
Никольский П.Н., Изотов И.И. 1936. Очерк растительности полосы вдоль Парандово-Ругозерского тракта (Карелия). – Тр. АН СССР. Сер. 3. Геоботаника. 3: 345–393.
Steijlen I., Zackrisson O. 1987. Long-term regeneration dynamics and successional trends in Northern Swedish coniferous forest stand. – Can. J. Bot. 65: 839–848. https://doi.org/10.1139/b87-114
Nicklasson M., Hörnberg G., Zackrisson O. 1998. Age structure and disturbance history in a boreal Pi-nus-Picea forest. – Acta Universitatis Agriculturae Sueciae. Silvestria. 52: 23 p.
Дыренков С.А.1984. Структура и динамика таежных ельников. Л. 174 с.
Работнов Т.А. 1950. Вопросы изучения состава популяций для целей фитоценологии. – Проблемы ботаники. 1: 466–483.
Уранов А.А. 1975. Возрастной спектр ценопопуляции как функции времени и энергетических волновых процессов. – Науч. докл. высш. шк. Биол. науки. 2: 7–34.
Диагнозы и ключи возрастных состояний лесных растений. Деревья и кустарники. 1989. М. 102 с.
Ставрова Н.И., Горшков В.В., Мишко А.Е. 2017. Онтогенез ели сибирской (Picea obovata Ledeb.) в малонарушенных северотаежных кустарничково-зеленомошных сосново-еловых лесах. – Бот. журн. 102(2): 163–185.
Санитарные правила в лесах СССР. 1970. М. 16 с.
Алексеев В.А. 1989. Диагностика жизненного состояния деревьев и древостоев. – Лесоведение. 4: 51–57.
Алексеев В.А. 1990. Некоторые вопросы диагностики и классификации поврежденных загрязнением лесных экосистем. – В кн.: Лесные экосистемы и атмосферное загрязнение. Л. С. 33–53.
Ярмишко В.Т., Горшков В.В., Ставрова Н.И. 2003. Виталитетная структура Pinus sylvestris L. в лесных сообществах с разной степенью и типом антропогенной нарушенности. – Растит. ресурсы. 39 (4): 1–19.
Мамаев С.А. Формы внутривидовой изменчивости древесных растений: (на примере семейства Pinaceae на Урале). М. 1973. 282 с.
Воропанов П.В. 1950. Ельники Севера. М.; Л. 179 с.
Попова Э.П. 1979. О продолжительности пирогенного воздействия на свойства лесных почв. – В кн.: Горение и пожары в лесу. Ч. 3. Красноярск. С. 110–116.
Санников С.Н., Санникова Н.С. 1985. Экология естественного возобновления сосны под пологом леса. М. 152 с.
Санников С.Н., Санникова Н.С. 2014. Лес как подземно-сомкнутая дендроценоэкосистема. – Сиб. лесн. журн. 1: 25–34. http://сибирскийлеснойжурнал.рф/upload/iblock/b62/b62af78f6a11a64fea970b13df821c49.pdf
Санникова Н.С., Санников С.Н., Петрова И.В., Мищихина Ю.Д., Черепанова О.Е. 2012. Факторы конкуренции древостоя-эдификатора: количественный анализ и синтез. – Экология. 6: 403–409.
Pothier D. 2017. Relationships between patterns of stand growth dominance and tree competition mode for species of various shade tolerances. – Forest Ecology and Management. 406: 155–162. https://doi.org/10.1016/j.foreco.2017.09.066
Ипатов В.С. 1968. Дифференциация древостоя I. – Вестн. ЛГУ. Сер. Биология. 21: 59–68.
Ипатов В.С. 1969. Дифференциация древостоя II. – Вестн. ЛГУ. Сер. Биология. 3: 43–54.
Ипатов В.С. 2010. Дифференциация деревьев в лесных сообществах. – Междисциплинарный научный и прикладной журнал “Биосфера”. 2 (4): 544–553.
Fraver S., Jonsson B.G., Jönsson M., Esseen P.-A. 2008. Demographics and disturbance; history of a boreal old-growth Picea abies forest. – J. Veg. Sci. 19(6): 789–798. https://doi.org/10.3170/2008-8-18449
Василенко Н.А. 2009. Динамика строения древостоев смешанных лесов юга Дальнего Востока. – Бюл. Бот. сада-ин-та ДВО РАН. 3: 78–86. http://botsad.ru/media/oldfiles/journal/number3/number3_5.pdf
Ухваткина О.Н., Комарова Т.А., Трофимова А.Д. 2010. Онтоморфогенез ели аянской (Picea ajanensis (Lindl. et Gord.) Fisch. ex Carr.) в условиях среднегорного пояса Южного Сихотэ-Алиня. – Бюл. Бот. сада-ин-та ДВО РАН. 2010. 5: 150–158. http://botsad.ru/media/oldfiles/journal/number5/25.pdf
Комарова Т.А. 2011. Рост и развитие Abies nephrolepis (Pinaceae) в Южном Сихотэ-Алине. – Раст. ресурсы. 47(4): 19–33.
Николаева С.А., Велисевич С.Н., Савчук Д.А. 2011. Онтогенез Pinus sibirica на юго-востоке З-ападно-Сибирской равнины. – Журн. Сиб. федерального ун-та. Биология. 4(1): 3–22. http://elib.sfu-kras.ru/bitstream/handle/2311/2375/Nikolaeva.pdf;jsessionid=7ADA96590603CECD6690B8AAD9193067?sequence=1
Махатков И.Д. 1991. Поливариантность онтогенеза пихты сибирской. – Бюл. МОИП. Отд. биол. 96 (4): 79–88.
Евстигнеев О.И. 2014. Поливариантность сосны обыкновенной в Брянском полесье. – Лесоведение. 2: 69–77.
Левин Г.Г. 1966. Возрастные изменения у растений (анализ некоторых понятий и представлений). – Бот. журн. 51(12): 1774–1795.
Алексеев В.А., Ярмишко В.Т. 1990. Влияние атмосферного загрязнения двуокисью серы с примесью тяжелых металлов на строение и продуктивность северотаежных древостоев. – В кн.: Лесные экосистемы и атмосферное загрязнение. Л. С. 105–114. https://www.booksite.ru/fulltext/rusles/Les_eco/text.pdf
Бебия С.М. 2000. Дифференциация деревьев в лесу, их классификация и определение жизненного состояния древостоев. – Лесоведение. 4: 35–43.
Ставрова Н.И., Горшков В.В., Катютин П.Н. 2010. Динамика виталитетной структуры ценопопуляций Picea obovata Ledeb. и Betula pubescens Ehrh. в процессе послепожарных сукцессий северотаежных еловых лесов. – Бот. журн. 95 (11): 1550–1566.
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы