

Растительные ресурсы, 2019, T. 55, № 1, стр. 45-55
Онтогенез и онтогенетическая структура ценопопуляций Plantago canescens (Plantaginaceae) в центральной Якутии
В. В. Семенова 1, *, Н. С. Данилова 1
1 Институт биологических проблем криолитозоны СО РАН
г. Якутск, Россия
* E-mail: vvsemenova-8@yandex.ru
Поступила в редакцию 03.04.2018
После доработки 02.07.2018
Принята к публикации 10.09.2018
Аннотация
Изучены 6 ценопопуляций Plantago canescens в Центральной Якутии. Plantago canescens – короткокорневищно-стержнекорневое с полициклическими побегами травянистое растение. В онтогенетическом цикле P. canescens выделены 9 онтогенетических состояний. В природе онтогенез продолжается от 4 до 11 лет. Изученные ценопопуляции нормальные и неполночленные. Из изученных ценопопуляций выявлены 2 типа онтогенетических спектров: левосторонний и центрированный. Центрированный тип онтогенетического спектра не типичный для P. canescens, он складывается в ценопопуляциях с неблагоприятными условиями произрастания под влиянием антропогенного и эколого-фитоценотического фактора. Характерным типом спектра вида является левосторонний тип. По оценке организменных и популяционных показателей наиболее благоприятные условия произрастания складываются в ЦП 2, неблагоприятные – в ЦП 5 и 6.
Plantago canescens Adams. – подорожник седоватый. Восточносибирско-североамериканский вид, заходящий в Арктику. Имеет разорванный ареал, приуроченный к бассейнам крупных рек от Хатанги до Маккензи [1, 2]. В дизъюнктивных фрагментах ареалов популяции находятся в пограничных условиях, определяющих своеобразие их структуры, чувствительность к проявлениям окружающей среды [3]. Описан из окресности г. Якутска. В Якутии встречается во всех районах. Растет по опушкам лесов, на сухих лугах, по степным склонам, на залежах, по обочинам дорог [4, 5].
Вид используется в народной медицине как кровоостанавливающее и ранозаживляющее средство [6]. В официальной медицине используют подорожник большой, его применяют как отхаркивающее, при заживлении ран, болезнях желудочно-кишечного тракта, малокровии и т.д. [7].
Целью статьи является изучение онтогенеза P. canescens, структуры и состояния его ценопопуляций в Центральной Якутии.
МАТЕРИАЛ И МЕТОДЫ
Исследования по изучению структуры ценопопуляций проводились в Центральной Якутии, на территории Якутского ботанического сада (ЯБС) в 2015 и 2016 гг. Исследованиями были охвачены 6 ценопопуляций P. canescens.
Описания растительных сообществ выполнены согласно общепринятым геоботаническим методам [8]. Ниже приводятся описания сообществ с участием P. canescens в окрестности г. Якутска.
Ценопопуляция 1 (ЦП 1) входит в состав остепненного разнотравного луга. В сообществе отмечено 18 видов, доминируют P. canescens, Myosotis arvensis (L.) Hill., Potentilla bifurca L., Onobrychis arenaria (Kit.) DC. Высота травостоя 20–25 см. Общее проективное покрытие (ОПП) – 50–60%, проективное покрытие (ПП) P. canescens – 10%. Антропогенное воздействие на участок минимальное, обусловленное только произрастанием в пределах рекреационной зоны.
Ценопопуляция 2 (ЦП 2) расположена на залежи, примыкающей к коллекционному питомнику и подпадает под зону регулярного полива. Почва рыхлая, богатая остатками перегноя. В сообществе отмечено 24 вида растений, доминируют P. canescens, Potentilla bifurca, Linaria acutiloba Fisch. ex Reichenb. Высота травостоя 30–35 см. ОПП – 60%, ПП P. canescens – 40%. Антропогенное воздействие на участок положительное.
Ценопопуляция 3 (ЦП 3) описана на опушке соснового леса, возле жилого дома. Почва задернованная, уплотненная в результате интенсивного вытаптывания. В сообществе отмечено 32 вида, доминируют Artemisia commutata Bess., Astragalus inopinatus Boriss., Galium verum L., Linum komarovii Juz., Oxytropis candicans (Pall.) DC., P. canescens, Potentilla bifurca. Высота травостоя 60–50 см. ОПП травостоя – 80%, ПП P. canescens – 10%.
Ценопопуляция 4 (ЦП 4) входит в состав смешанного леса, расположенного на правом берегу оз. Ытык-Кюель. В древесном ярусе отмечены Betula pendula Roth., Salix bebbiana Sarg., Crataegus dahurica Koechne et Schneid. Сомкнутость крон составляет 20%. В травянистом ярусе отмечено 11 видов, доминируют P. canescens, Jacobaea vulgaris Gaertn., Astragalus inopinatus, Melilotus albus Medik., Achillea millefolium L. Высота травостоя 60–40 см. ОПП – 60%, ПП P. canescens – 50%. Имеется подстилка из опада листьев мощностью 0.5 см. Антропогенная нагрузка минимальная.
Ценопопуляция 5 (ЦП 5) входит в состав остепненного типчаково-разнотравного луга, произрастающего у подножия коренного берега р. Лена Чучур-Муран. В сообществе произрастают 13 видов, доминируют P. canescens, Potentilla bifurca L., Veronica incana L., Astragalus danicus Retz., Festuca lenensis Drob., Pulsatilla multifida (G.Pritz.) Juz. Высота травостоя 25–30 см. ОПП травостоя 60%, ПП P. canescens – 5–7%. Ценопопуляция испытывает умеренное антропогенное воздействие в виде вытаптывания и нерегламентированного выпаса; кроме того, участок расположен под высоковольтной линией электропередач.
Ценопопуляция 6 (ЦП 6) входит в состав тонконоговой степи, расположенной на территории, близкой к коллекционным участкам и подвергается интенсивному вытаптыванию, воздействию тяжелой сельскохозяйственной техники. Почва очень плотная, сухая. В сообществе отмечено 9 видов, доминируют Carex duriuscula C.A. Mey., P. canescens Veronica incana, Koeleria cristata (L.) Pers. Высота травостоя 35 см. ОПП – 35%, ПП P. canescens – 10%.
Описание онтогенеза P. canescens с выделением онтогенетических состояний и определение онтогенетических спектров их ценопопуляций были выполнены согласно общепринятым методикам [3, 9–13]. Онтогенетическая структура исследуемых ценопопуляций характеризовалась в соответствии с представлениями характерного спектра [14]. Определение типа популяций осуществлялось по классификации “дельта–омега” Л.А. Животовского [15].
Оценка состояния ценопопуляций проводилась с использованием организменных и популяционных признаков [14]. Организменные признаки зрелых генеративных особей: 1 – высота растения, см; 2 – число генеративных побегов, см; 3 – длина соцветий, см. Популяционные признаки: 4 – плотность особей на единицу площади, экз./0.25 м2; 5 – доля молодой фракции (j–g1), %; 6 – доля генеративной фракции (g2–g3), %. Диапазон каждого признака разбивался на 5 классов с одинаковым объемом по равномерной шкале, каждому классу присваивался балл, наименьший балл соответствовал наименьшим показателям.
Жизненная форма Plantago canescens и онтогенез
Plantago canescens – короткокорневищно-стержнекорневое с полициклическими побегами травянистое растение, мезоксерофит. Смена моноподиального нарастания начинается со старого генеративного онтогенетического состояния.
Онтогенез P. canescens продолжается от 4 до 11 лет. В онтогенезе особей P. canescens было выделено 9 онтогенетических состояний, описание которых было проведено в ЦП 5 (рис. 1).
Рис. 1.
Онтогенез P. canescens. j–ss – онтогенетические состояния. Морфологичесие структуры: a – розеточный побег; b – корневище, подземный побег; c – придаточные корни; d – боковые корни; e – придаточные почки; f – главный стержневой корень. Fig. 1. Ontogeny of the P. canescens. j–ss – ontogenetic state. Morphological structures: a – rosette shoot; b – rhizome, underground shoot; c – secondary roots; d – lateral roots; e – adventitious buds; f – main root.

Прорастание семян надземное. Проростки представлены моноподиальным побегом, которые имеют линейные семядольные листья и 1–2 настоящих листа линейно-продолговатой формы длиной 0.3–0.4 см с одной жилкой. Белые характерные волоски появляются, начиная с семядольных листьев. Главный корень удлиняется до 1–3 см с боковыми корнями до 3 шт.
Розеточные однопобеговые ювенильные особи формируют до 2–3 длинночерешковых листьев узколанцетной формы с одной жилкой. Листья 1.0–2.0 см дл., 0.2–0.4 см шир. Длина черешка 1.0–2.0 см. Главный корень удлиняется до 5 см. В этом состоянии особи находятся около 1–2 лет.
Переход в имматурное состояние отмечается с появлением наряду с листьями с 1 жилкой (ювенильный тип) листьев с 3 жилками (взрослый тип). На розеточном побеге насчитывается 2–3 листа. Форма листьев узколанцетная и ланцетная. Размеры листьев 1.7–4.5 см дл., 0.3–0.6 см шир., длина черешка 2.5–5.0 см. Базальная часть побега с помощью контрактильных боковых и придаточных корней начинает углубляться в почву, формируя короткое эпигеогенное корневище. На корневище формируются придаточные спящие почки возобновления. Главный корень удлиняется. Число боковых корней увеличивается, также образуются боковые корни II порядка. Тип корневой системы становится смешанным. В это состояние особи могут перейти в год прорастания и находиться в нем до 1–2 лет.
В виргинильном состоянии на розеточном побеге число длинночерешковых листьев насчитывается до 2–3 (5). В этом состоянии формируется первичный куст, боковые побеги появляются из пазушных и придаточных почек на подземном побеге. Все листья 0.7 см шир., длина черешка 3.0–7.0 см. Число придаточных и боковых корней увеличивается. У некоторых особей, в основном из ЦП 5, на главном корне формируются корневые почки возобновления, которые остаются в спящем состоянии на протяжении всего онтогенеза. Вероятно, эти резервные почки могут раскрыться при механическом повреждении надземных побегов. В благоприятных условиях произрастания с хорошей аэрацией, водоснабжением, отсутствием постоянной сомкнутости травостоя, а также при повреждении главного побега у растений наблюдается корнеотпрысковость. Корнеотпрысковость отмечалась у P. lanceolata L. при гибели его надземных побегов, при котором происходило массовое заложение придаточных почек на главном и боковых корнях [13]. В виргинильное состояние особи могут перейти в год прорастания, поэтому виргинильный период может продолжаться до 1–4 лет.
В молодом генеративном состоянии моноподиальный рост особей P. canescens продолжается, репродуктивные органы развиваются из пазушных почек. В молодом генеративном состоянии особи представлены кустом из 2 розеточных побегов – вегетативного и репродуктивного или без кущения только из одного репродуктивного розеточного побега. Розеточный побег несет до 3–4 листьев и один цветоносный безлистный побег, редко два, длиной 12–26 см. Длинночерешковые листья имеют по 3–5 жилок. Листья могут быть более крупными, чем в виргинильном состоянии, пластинка листа увеличивается до 4.5–7.0 см в дл. и 0.6–1.0 см в шир. Соцветия достигают в длину 1.0–3.5 см. В корневой системе число боковых корней на главном корне и придаточных корней на корневище заметно увеличивается, появляется III порядок боковых корней. Продолжительность молодого генеративного состояния длится от 1 до 2 лет.
В зрелом генеративном состоянии особи достигают максимального развития. Моноподиальный рост побегов продолжается. В кусте содержатся до 3 генеративных розеточных побегов и редко 1 вегетативный розеточный побег. Репродуктивный розеточный побег несет до 3–5 листьев и до 5 цветоносов. Размеры листьев и соцветий как у молодых генеративных особей. На главном корне у некоторых особей число придаточных почек возобновления увеличивается, редко на толстых боковых корнях также появляются придаточные почки. В этом состоянии особи могут находиться до 3 лет.
В старом генеративном состоянии моноподиальное нарастание сменяется на симподиальное. В этом состоянии уменьшается число всех живых структур. В кусте образуются до 2 репродуктивных розеточных побегов и редко 1 вегетативный розеточный. В репродуктивном розеточном побеге содержится до 3 цветоносов. Размеры листьев и генеративных органов в основном не меняются. В кусте наблюдаются отмершие остатки листьев, цветоносов, в подземной части имеются отмершие корневища, число почек заметно сокращается, главный корень сохраняется. Редко при сильном разрастании куста наблюдается процесс партикуляции вдоль стержневого корня. В этом состоянии особи находятся до 1–2 лет и большинство их завершают свой жизненный цикл.
До субсенильного состояния доживают единичные особи P. canescens. В этом состоянии они прекращают формировать генеративные органы, куст состоит из вегетативных розеточных побегов. В кусте имеются отмершие части побегов и листьев. В подземной части скелет главного корня сохраняется и число корней сокращается. В этом состоянии особи могут находиться до 1–2 лет.
Морфогенез P. canescens состоит из следующих фаз: первичный побег (p, j, im) – первичный куст (v, g1, g2, g3, ss). Вид образует моноцентрическую биоморфу.
Онтогенетическая структура ценопопуляций Plantago canescens
Изученные сообщества с участием P. canescens находятся в различных эколого-фитоценотических условиях и различаются по степени увлажнения участков, флористическому составу, общему проективному покрытию травостоя, степени антропогенного воздействия на них и др. Ценопопуляции P. canescens входят в состав степных, луговых, лесных сообществ, а также встречаются на залежных землях. Дефицитом влаги характеризуются ЦП 1, 3, 5, 6, расположенные на открытых, хорошо освещаемых местах. Здесь надо отметить, что по количеству годовых осадков Центральная Якутия соответствует полупустынным районам [16]. ЦП 4 входит в состав смешанного леса на берегу оз. Ытык-Кюель, здесь складываются более комфортные условия: затененность, снижение испарения кронами деревьев и сохранение влаги в почве, благодаря защитному слою листового опада. Благоприятные условия сложились и в ЦП 2, которая прилегает к коллекционному питомнику и подпадает под зону регулярного полива дальноструйной системой. Об этом свидетельствуют биометрические показатели средневозрастных генеративных растений: в ЦП 2 и 4 отмечаются максимальные значения и снижение их в ЦП 1, 3, 5 и 6 (табл. 1).
Таблица 1.
Биометрические показатели средневозрастных растений в ценопопуляциях Plantago canescens Table 1. Biometric indicators of middle-aged plants in Plantago canescens coenopopulations
| ЦП СP |
Число побегов
в кусте, шт. Number of shoots per plant, pcs. |
Длина цветоносного
побега, см Length of flower spike, cm |
Длина соцветия, см Length of inflorescence, cm |
Число цветоносов
в розеточном побеге, шт. Number of flower spikes in rosette, pcs. |
Длина черешка, см Length of petiole, cm |
Длина пластинки
листа, см Leaf blade length, cm |
Ширина пластинки
листа, см Leaf blade width, cm |
|---|---|---|---|---|---|---|---|
| 1 | 1.1 ± 0.1 | 21.5 ± 0.6 | 2.7 ± 0.3 | 2.3 ± 0.2 | 5.0 ± 0.3 | 6.1 ± 0.4 | 0.7 ± 0 |
| 2 | 1.1 ± 0.1 | 29.5 ± 0.8 | 3.8 ± 0.2 | 4.2 ± 0.2 | 7.2 ± 0.7 | 8.4 ± 0.2 | 1.2 ± 0.1 |
| 3 | 2.8 ± 0.4 | 19.4 ± 1.4 | 3.4 ± 0.2 | 1.9 ± 0.3 | 3.9 ± 0.3 | 6.5 ± 0.4 | 1.3 ± 0.1 |
| 4 | 1.2 ± 0.1 | 35.0 ± 1.7 | 4.2 ± 0.4 | 2.1 ± 0.3 | 6.4 ± 1.0 | 12.5 ± 0.6 | 2.9 ± 0.4 |
| 5 | 1.8 ± 0.2 | 25.1 ± 1.0 | 2.7 ± 0.1 | 2.1 ± 0.2 | 6.7 ± 0.7 | 7.2 ± 0.4 | 0.8 ± 0.1 |
| 6 | 1.9 ± 0.1 | 18.1 ± 1.0 | 2.2 ± 0.3 | 3.0 ± 0.3 | 2.9 ± 0.3 | 4.4 ± 0.3 | 0.8 ± 0.1 |
Степень антропогенного воздействия на ЦП 1, 2, 4 минимальная. Умеренную рекреационную нагрузку испытывает ЦП 5; достаточно сильное антропогенное воздействие испытывает ЦП 3, которое проявляется в виде интенсивного вытаптывания; высокое антропогенное влияние ощущается в ЦП 6 (транспортная нагрузка, сельскохозяйственная техника, вытаптывание). На этих участках вытаптывание привело к отсутствию подроста в ценопопуляциях (ЦП 3, 6). Кроме того, у особей, произрастающих в ЦП 3, 5, 6, вследствие вытаптывания рано может отмирать главный побег и моноподиальный рост особей преждевременно сменяется на симподиальный. Это ведет к развитию боковых побегов из пазушных почек возобновления главного побега или придаточных почек на гипокотиле, придаточных корней и росту главного корня, что уводит нижние части побегов вглубь почвы с помощью контрактильности боковых и придаточных корней. На рыхлых и увлажненных почвах на залежи и в лесном сообществе (ЦП 2 и 4) глубина проникновения в почву подземных частей побега достигает 4.0–4.5 см, в ЦП 1, 3 и 5 подземные части побегов составляют в длину 2.5–3.0 см; на сухих и очень плотных почвах в ЦП 6 нижние части побегов особей не углубляются в почву, а располагаются горизонтально по поверхности почвы, удлиняясь всего на 1 см. Отдельно можно отметить, что вытаптывание влечет за собой увеличение числа побегов в кусте; так в ценопопуляциях, в которых достаточно сильно антропогенное воздействие заметно увеличение числа побегов, максимальное число побегов наблюдается в ЦП 3 (2.8 см) и в ЦП 5 и 6 (1.8 и 1.9 см, табл. 1).
Плотность изученных ценопопуляций P. canescens варьирует в среднем от 3.5 до 52.5 особей на единицу площади 0.25 м2 (табл. 2). Высокие показатели плотности особей наблюдаются в ЦП 1 и 2 (66.5 и 52.5 на 0.25 м2), низкие – в ЦП 6 (3.5). Проростки имеются только в ЦП 1, 2 и 4 (1.5–2.6 на 0.25 м2).
Таблица 2.
Демографические показатели ценопопуляций Plantago canescens Table 2. Demographic indicators of Plantago canescens сoenopopulations
| П N |
Ценопопуляция Coenopopulation |
|||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | |
| М | 66.5 ± 0 | 52.5 ± 0.2 | 25.25 ± 0.2 | 17.3 ± 0.1 | 13.5 ± 0.1 | 3.5 ± 0.1 |
| Δ | 0.17 | 0.22 | 0.14 | 0.17 | 0.35 | 0.43 |
| ω | 0.42 | 0.55 | 0.42 | 0.47 | 0.73 | 0.80 |
| * | Молодые Young |
Молодые Young | Молодые Young |
Молодые Young | Зрелые Mature |
Зрелые Mature |
Примечание. Признаки (П): М – плотность особей вида в ценопопуляциях (особей/0.25 м2), Δ – индекс возрастности, ω – индекс эффективности, * – классификация популяций. Note. Indicators (N): M – species density in сoenopopulation (individuals/0.25 m2), Δ – age index, ω – efficiency index, * – classification of populations.
Изученные ценопопуляции характеризуются как нормальные и неполночленные ценопопуляции. По типам характерных спектров Л.Б. Заугольновой [14] онтогенетический спектр P. canescens относится к левостороннему типу. Это выявляется по преобладанию прегенеративной группы особей в ценопопуляциях, а также для видов с простым онтогенезом со стержнекорневой биоморфой и коротким постгенеративным периодом. Изученные ценопопуляции имеют левосторонние (ЦП 1–4) и центрированные (ЦП 5 и 6) типы спектров (табл. 3).
Таблица 3.
Онтогенетический спектр ценопопуляций Plantago canescens, % Table 3. Ontogenetic spectrum of Plantago canescens coenopopulations, %
| ЦП СP |
j | im | v | g1 | g2 | g3 | ss |
|---|---|---|---|---|---|---|---|
| Ценопопуляции с левосторонним спектром Cоenopopulations with a left-sided spectrum |
|||||||
| 1 | 27.4 | 21.1 | 19.5 | 14.7 | 16.5 | 0.4 | 0.4 |
| 2 | 8.1 | 21.4 | 27.1 | 17.4 | 25.3 | 0.7 | – |
| 3 | – | 15.8 | 77.2 | – | 6.5 | – | 0.5 |
| 4 | 4.8 | 25.0 | 47.6 | 2.9 | 19.7 | – | – |
| Ценопопуляции с центрированным спектром Cоenopopulations with a centered spectrum |
|||||||
| 5 | 4.9 | 4.9 | 20.4 | 19.8 | 42.6 | 5.6 | 1.8 |
| 6 | – | 0.8 | 17.2 | 18.7 | 46.9 | 12.5 | 3.9 |
Левосторонние типы спектров ценопопуляций P. canescens складываются на остепненном лугу, залежи, опушке соснового леса и в смешанном лесу (ЦП 1–4). В этих сообществах число видов колеблется от 11 до 32 видов, общее проективное покрытие их довольно высокое – 50–80%. Изученные ценопопуляции двухвершинные, абсолютные максимумы в онтогенетических спектрах приходятся на ювенильные (27.4%) и виргинильные группы (27.1–77.2%), вторые максимумы – на средневозрастные группы особей (16.5–25.3%, табл. 3). По классификации Л.А. Животовского [15] все левосторонние ценопопуляции относятся к молодому типу (табл. 2). В этих ценопопуляциях плотность особей P. canescens выше (17.3–66.5 на 0.25 м2, табл. 2), чем в ценопопуляциях с центрированным типом спектра (3.5–13.5 на 0.25 м2). На увеличение доли виргинильных и средневозрастных особей в онтогенетических спектрах ценопопуляций влияют стрессовые факторы, такие как выпас скота, сенокошение, вытаптывание, приводящие к гибели подроста [17, 18]. Отсутствие подроста в ЦП 3 обусловлено вытаптыванием, а в ЦП 4, входящем в состав лесного сообщества, – слабой освещенностью и развитым слоем листового опада. Вероятно, эти факторы оказывают влияние на периодичность семенного размножения и выпад или низкое прорастание подроста. Из изученных ценопопуляций P. canescens только в ЦП 1 абсолютный максимум приходится на ювенильные группы (27.4) и отмечается самая высокая плотность особей (66.5 на 0.25 м2). Сохранение подроста в ценопопуляции связано с минимальной антропогенной нагрузкой на сообщество.
В центрированных типах спектров в ЦП 5 и 6 наблюдается низкая плотность особей (13.5 и 3.5 на 0.25 м2). Ценопопуляции входят в состав остепненного типчаково-разнотравного луга и тонконоговой степи. Видовой состав насчитывает 13 и 9 видов, общее проективное покрытие 60 и 35%. Абсолютные максимумы приходятся на зрелые генеративные фракции (42.6–46.9%), второй пик в ЦП 5 на виргинильные (20.4%). В ЦП 6, испытывающей очень высокую антропогенную нагрузку, также отсутствует подрост. По классификации “дельта–омега” ценопопуляции относятся к зрелому типу.
Оценку состояния ценопопуляций P. canescens проводили при помощи организменных и популяционных признаков, в качестве организменных показателей служили следующие признаки средневозрастных генеративных особей: высота, число генеративных побегов, высота соцветий (табл. 4). В качестве популяционных признаков выбраны: плотность особей на 0.25 м2, доля особей молодой фракции (j–g1), доля генеративных особей в онтогенетическом спектре ценопопуляции (g2–g3). По сумме баллов наиболее благоприятные условия складываются в ЦП 2 (25, рис. 2).
Таблица 4.
Оценка признаков Planago canescens (в баллах) Table 4. Assessment of signs Plantago canescens (score)
| № | Признак Indicator |
Баллы Scores |
||||
|---|---|---|---|---|---|---|
| I | II | III | IV | V | ||
| 1 | Высота цветоноса, см Height of flower spike, cm |
18.1–21.5 | 21.5–24.9 | 24.9–28.3 | 28.3–31.6 | 31.7–35.0 |
| 2 | Длина соцветия, см Length of inflorescence, cm |
2.2–2.6 | 2.6–3.0 | 3.0–3.4 | 3.4–3.8 | 3.8–4.2 |
| 3 | Число генеративных
побегов, шт. Number of generative shoots, pcs. |
1.9–2.4 | 2.4–2.8 | 2.8–3.3 | 3.3–3.7 | 3.7–4. |
| 4 | Плотность особей,
экз./0.25 м2 Species density, ind./0.25 m2 |
3.6–16.1 | 16.2–28.7 | 28.7–41.3 | 41.3–53.9 | 53.9–66.5 |
| 5 | Доля j–g1, % Share of j–g1, % |
36.7–47.8 | 48.0–59.3 | 59.3–70.5 | 70.5–81.8 | 81.8–93.1 |
| 6 | Доля g2–g3, % Share of g2–g3, % |
6.4–17.0 | 17.0–27.6 | 27.6–38.2 | 38.2–48.8 | 48.8–59.4 |
Рис. 2.
Диаграмма состояния ценопопуляций P. canescens (в баллах). Организменные признаки (средневозрастной генеративной особи): 1 – высота цветоноса, см; 2 – длина соцветия, см; 3 – число генеративных побегов, г; популяционные признаки: 4 – плотность особей, экз./0.25 м2; 5 – доля молодой фракции (j–g1), %; 6 – доля генеративной фракции (g2–g3), %. Fig. 2. Diagram of P. canescens coenopopulations status (score). Characters of organism (mature generative individual): 1 – height of peduncle, sm; 2 – length of inflorescence, sm; 3 – number of generative shoots, pcs.; characters of populations: 4 – density of individuals, ind./0.25 m2; 5 – share j–g1, %; 6 – share g2–g3, %.
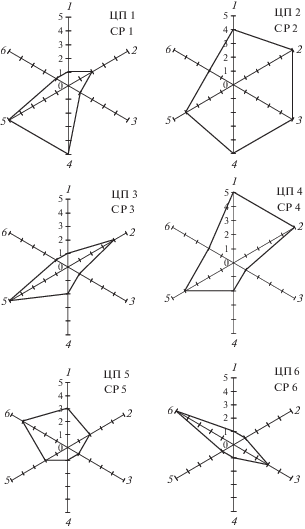
В условиях влажного и легкого субстрата особи чувствуют себя наиболее комфортно, организменные и популяционные показатели в них высокие (14 и 11, табл. 4). Средние показатели по сумме баллов отметили в ЦП 4 (19). В благоприятных лесных условиях также соответственно увеличиваются организменные показатели (11), а популяционные показатели имеют низкие значения (8). Остальные ценопопуляции по сумме баллов имеют низкие показатели. Наиболее низкие значения фиксировались в ЦП 5 и 6 (13 и 12), которые имеют центрированные типы спектров с абсолютным максимумом на средневозрастные растения и с самой низкой плотностью особей. Вероятно, негативные антропогенные и экологические факторы приводят к снижению биометрических показателей и плотности особей. Из общего фона выделяется ЦП 1, в этой ценопопуляции организменные показатели имеют самые низкие баллы (4), популяционные, напротив, высокие (11). На открытом солнцу участке наиболее проявляются засушливые условия района исследований, которые ограничивают рост растений, но отсутствие антропогенного воздействия способствует увеличению плотности особей в ценопопуляции.
ВЫВОДЫ
Анализ полученных данных изучения 6 ценопопуляций Plantago canescens в различных экологических условиях и с антропогенным влиянием на территории ЯБС Центральной Якутии дает следующие выводы.
1. P. canescens короткокорневищное стержнекорневое с полициклическими побегами травянистое растение, мезоксерофит. Смена моноподиального нарастания начинается со старого генеративного онтогенетического состояния.
2. В онтогенезе P. canescens выделены 9 онтогенетических состояний. В природе онтогенез продолжается от 4 до 11 лет. Морфогенез состоит из следующих фаз: первичный побег – первичный куст. Вид образует моноцентрическую биоморфу.
3. Изученные ценопопуляции нормальные и неполночленные. Характерным типом спектра вида является левосторонний. Среди изученных ценопопуляций выявлены 2 типа онтогенетических спектра: левосторонний и центрированный. Центрированный тип онтогенетического спектра не типичный для P. canescens, он складывается в ценопопуляциях с неблагоприятными условиями произрастания под влиянием антропогенного и эколого-фитоценотического фактора.
4. По оценке организменных и популяционных показателей наиболее благоприятные условия произрастания складываются в ЦП 2, на залежи, куда регулярно подается искусственный полив. Наиболее неблагоприятные условия находятся в ЦП 5 и 6, где сообщества испытывают влияние различных факторов.
Таким образом, состояние ценопопуляций P. canescens зависит от эколого-фитоценотических условий произрастания. Вид хорошо реагирует на увлажнение почвы, увеличивая морфометрические показатели, и чувствителен к антропогенному воздействию.
БЛАГОДАРНОСТИ
Работа выполнена в рамках выполнения государственного задания Института биологических проблем криолитозоны СО РАН на 2017–2020 гг. по теме “Фундаментальные и прикладные аспекты изучения разнообразия растительного мира Северной и Центральной Якутии” (№ госрегистрации АААА-А17-117020110056-0).
Список литературы
Юрцев Б.А. 1981. Реликтовые степные комплексы Северо-Восточной Азии. Новосибирск. 168 с.
Арктическая флора СССР. 1983. Вып. VIII. Ч. 2. Семейства Plantaginaceae – Orobanchaceae. Л. 52 с.
Ценопопуляции растений (Основные понятия и структура). 1976. М. 217 с.
Флора Сибири. 1996. Т. 12: Solanaceae–Lobeliaceae. Новосибирск. 208 с.
Кузнецова Л.В., Захарова В.И. 2012. Конспект флоры Якутии. Сосудистые растения. Новосибирск. 272 с.
Савкина З.П., Андреева Т.В., Говорина Т.П., Данилова Н.С., Парилова В.Т., Цареградская А.П. 1981. Дикоpастущие тpавы Якутии в культуpе. Hовосибиpск. 243 с.
Телятьев В.В. 1991. Целебные клады. Иркутск. 400 с.
Корчагин А.А. 1964. Видовой (флористический) состав растительных сообществ и методы его изучения. – В кн.: Полевая геоботаника. Т. 3. С. 39–62.
Работнов Т.А. 1950. Жизненный цикл многолетних травянистых растений в луговых ценозах. – Тр. БИН АН СССР, сер.3. Геоботаника. 3(6): 7–197.
Уранов А.А. 1967. Онтогенез и возрастной состав популяций / Онтогенез и возрастной состав популяций цветковых растений. М. С. 3–8.
Уранов А.А. 1975. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов. – Биол. науки. 2: 7–34.
Заугольнова Л.Б., Жукова Л.А., Комаров А.С., Смирнова О.В. 1988. Ценопопуляции растений: Очерки популяционной биологии. М. 184 с.
Османова Г.О. 2007. Морфологические особенности особей и структура ценопопуляции Plantago lanceolata L. Йошкар-Ола. 184 с.
3аугoльнова Л.Б. 1994. Структура популяций семенных растений и проблемы их мон-иторинга: Автореф. дис. … д-ра биол. наук. СПб. 70 с. http://earthpapers.net/strukt-ura-populyatsiy-s-emennyh-rasteniy-i-problemy-ih-monitoringa
Животовский Л.А. 2001. Онтогенетическое состояние, эффективная плотность и классификация популяций. – Экология. 1: 3–7.
Гаврилова М.К. 1973. Климат Центральной Якутии. Якутск. 96 с.
Семенова В.В., Егорова П.С. 2013. Поливариантность онтогенеза Valeriana alternifolia Ledeb. и структура ее природных ценопопуляций в Якутии. Новосибирск. 111 с.
Семенова В.В., Данилова Н.С. 2017. Онтогенез и структура ценопопуляций Oxytropis candicans (Fabaceae) в Центральной Якутии. – Раст. ресурсы. 53(4): 513–526.
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы