

Растительные ресурсы, 2019, T. 55, № 2, стр. 151-169
Динамические модели наземных экосистем для количественной оценки продуктивности растительности
О. Г. Чертов 1, *, П. Я. Грабарник 2, В. Н. Шанин 2, 3, С. С. Быховец 2, Б. С. Петропавловский 4, И. В. Припутина 2, П. В. Фролов 2, Е. В. Зубкова 2
1 Бингенский технический университет
г. Бинген, Германия
2 Институт физико-химических и биологических проблем почвоведения РАН
г. Пущино, Россия
3 Центр по проблемам экологии и продуктивности лесов РАН
г. Москва, Россия
4 Ботанический сад-институт ДВО РАН
г. Владивосток, Россия
* E-mail: oleg_chertov@hotmail.com
Поступила в редакцию 25.11.2018
После доработки 12.12.2018
Принята к публикации 16.01.2019
Аннотация
Рассмотрены существующие подходы к оценке роста и продуктивности растительных сообществ. Подчеркивается необходимость создания наукоемких методов оценки и прогноза продуктивности растительных сообществ и наземных экосистем. Такими инструментами становятся математические (имитационные, численные, динамические) модели. Модели продуктивности травяных экосистем имеют средне- или долгосрочный уровень прогнозирования продуктивности различных по площади растительных сообществ. Модели агроэкосистем прогнозируют динамику роста посевов в течение одного вегетационного сезона и, как правило, с дневным шагом по времени. Лесные модели чаще используют для оценки долгосрочной динамики продуктивности насаждений. Рост растений в моделях травяных сообществ и посевах вычисляется по функциям фотосинтеза и дыхания, а разложение растительных остатков – в почвенных моделях. Лесные модели рассчитывают динамику продуктивности насаждений различными способами: по таблицам хода роста, по эмпирическим функциям нетто биологической продуктивности и по моделям фотосинтеза и дыхания, также в сочетании с вычислением разложения лесного опада. Первоначально модели динамики наземных экосистем были инструментом теоретических исследований. В настоящее время они используются для количественной оценки влияния на продуктивность растительных сообществ разных режимов хозяйственного использования, пожаров, загрязнения среды и для оценки последствий изменения климата. В создающихся в агрономии и лесоводстве системах поддержки принятия решений эти модели служат основным компонентом. Делается заключение о перспективности развития динамического моделирования для оценки продуктивности и динамики наземных экосистем в декларированной наукоемкой биоэкономике.
ВВЕДЕНИЕ
Запросы современного общества привели к формулировке новых подходов в природопользовании, в том числе в отношении возобновляемых растительных ресурсов. Парадигма устойчивого развития [1] в настоящее время дополнена активно развивающейся концепцией биоэкономики [2]. Но если в отношении устойчивого развития в природопользовании уже выработаны вполне определенные критерии оценки экологических функций наземных экосистем, в частности, “экосистемные услуги” [3], то биоэкономика играет роль перспективной стратегической цели природопользования будущего.
Как известно, с начала ХХ в. основными инструментами оценки продуктивности растительности в мире были бонитировка почв и земель в сельском хозяйстве [4–6] и таблицы хода роста (ТХР) древостоев в лесоводстве [7, 8]. Эти два инструмента оценки и прогноза продуктивности, основанные на обобщении эмпирических данных того времени, были вполне удовлетворительными в условиях экстенсивного природопользования при неконтролируемой эксплуатации почвенно-растительных ресурсов Земли.
Однако в XXI в. возникла потребность в более точных количественных оценках динамических параметров и экологических характеристик культурной и лесной растительности, в новых методах расчета продуктивности и структурной динамики и других компонентов лесных, травяных и агроэкосистем для их научно обоснованного использования в качестве “экосистемных услуг” [3], что в более точном переводе означает “экосистемные активы”.
Таким методом служит математическое моделирование динамики и продуктивности растительности и наземных экосистем [9, 10, 33]. Эта ветвь экологии начала интенсивно развиваться с середины ХХ в. в связи с реализацией Международной биологической программы (МБП) и программы “Человек и биосфера” (ЧиБ), в рамках которых был получен большой экспериментальный материал, до сих пор использующийся при моделировании экосистем. В настоящее время в системной экологии существует уже несколько направлений математического имитационного моделирования экосистем [11, 12].
Цель данной работы – рассмотрение существующих подходов к оценке продуктивности разных типов растительности и ее динамики в наземных экосистемах на основе математических методов для анализа ресурсного потенциала фитоценозов, а также как инструмент для экологического обоснования и разработки новых подходов в биоэкономике – основе природопользования ХХI в.
СУЩЕСТВУЮЩИЕ МЕТОДЫ ОЦЕНКИ ПРОДУКТИВНОСТИ И ДИНАМИКИ РАСТИТЕЛЬНОСТИ
На качественном уровне прямое отношение к оценке продуктивности растений имеют экологические шкалы, в которых по градациям богатства и влажности почвы описывается эдафический ареал существования растений [13], но без количественных оценок их продуктивности.
Предшественниками динамической оценки продуктивности растительности можно считать “экологию земель” Л.Г. Раменского [14] и существующие в лесоведении классификации типов леса и местообитаний, в которых каждая классификационная единица имеет характеристику продуктивности насаждений [15–17], однако с достаточно широким интервалом ее значений.
Существующая бонитировка почв в сельском и лесном хозяйстве является практически ориентированным подходом в оценке продуктивности растительности. В сельском хозяйстве бонитировка почв представляет собой ранжирование урожайности культурных растений в зависимости от свойств почв, таких как гранулометрический состав, кислотность, содержание гумуса и подвижных элементов питания. Это ранжирование может проводиться как в физических единицах урожайности, так и в условных баллах [5, 18]. При этом оценивается не общая биомасса растений, а только ее используемая часть. Такая оценка является, как правило, региональной, но может быть интегрирована в агроклиматическое районирование [6]. Также предпринимались успешные попытки бонитировки лесных почв по такой же методике [19, 20]. Однако они не получили распространения в связи с отсутствием почвенной информации на землях лесного фонда.
В лесоводстве методами оценки продуктивности леса, фактически прироста древесины, остаются таблицы хода роста (ТХР). Первые ТХР были созданы еще в XIX в., а современные ТХР до сих пор используются в большинстве стран мира. ТХР были основаны на представлении о неизменности почвенных условий произрастания [21]. Изначально они задумывались только как таблицы относительной производительности древостоев по классам бонитета, и в этом своем качестве ТХР необходимы и не потеряли своего значения и в XXI в. Однако в настоящее время ТХР придан статус именно таблиц “хода роста”. Это означает, что без экспериментальной проверки было принято, что темпы роста древостоев строго соответствуют начальному классу бонитета, т.е. исходному плодородию почвы [7]. ТХР совершенствуются и используются для оценки темпов роста древостоев в лесохозяйственном планировании и в ХХI в. [8].
В настоящее время ТХР подвергаются серьезной критике, поскольку данные долговременных наблюдений на постоянных пробных площадях показывают, что реальные кривые хода роста лесных сообществ свидетельствуют об увеличении продуктивности леса на 2–3 класса бонитета в период интенсивного роста [22–27] со снижением продуктивности в старых лесах [28]. Принимая также во внимание то, что в ТХР приводятся только высота, диаметр и запасы древесины в чистых (монодоминантных) насаждениях без рассмотрения всей биомассы и других компонентов лесных экосистем, можно заключить о неудовлетворительности этого инструмента для решения новых экологических задач в лесоводстве. Таким образом, существовавшие ранее методы оценки и прогноза продуктивности растительности уже не могут в полной мере удовлетворить современные запросы природопользования в рамках декларированной концепции биоэкономики.
МЕТОДЫ ДИНАМИЧЕСКОЙ ОЦЕНКИ ПРОДУКТИВНОСТИ РАСТИТЕЛЬНОСТИ
Математическое моделирование в естественных науках можно определить как наиболее точное количественное описание функционирования природного объекта или явления. Фактически это запись в математических функциях существующих знаний об объекте с возможностью вычислить его изменения при различных сочетаниях как свойств самого объекта, так и влияющих на него разнородных факторов. При этом появляется возможность получения дополнительных знаний о поведении объекта на основе уже имеющегося арсенала экспериментальной науки [11, 29].
Теоретической основой математического описания продуктивности и динамики растительности является существующий в экологии закон толерантности Шелфорда, описывающий интервал значений факторов, при которых возможно существование и рост организма [30, 31]. При этом сам рост описывается сигмоидальной кривой в области дефицита фактора/ресурса для роста растений с выходом на плато в оптимуме и снижением продуктивности при избыточности фактора роста.
В современный период происходит энергичное развитие динамического (численного, имитационного) моделирования (simulation modelling) травяных, агро- и лесных экосистем [9, 32–36] и количество моделей уже исчисляется сотнями. Ранее они использовались для теоретического анализа роста и динамики растительности; теперь они применяются и для решения экологических задач, а также для создания систем сельского и лесного хозяйства.
Существует много различных классификаций моделей растительности и наземных экосистем, однако многие модели имеют характеристики нескольких классификационных подходов. Поэтому ниже они рассматриваются по четырем крупным обобщенным категориям: модели травяных и агроэкосистем (модели урожайности, crop models), лесные модели масштаба растительного сообщества, ландшафтные (региональные) модели, глобальные модели.
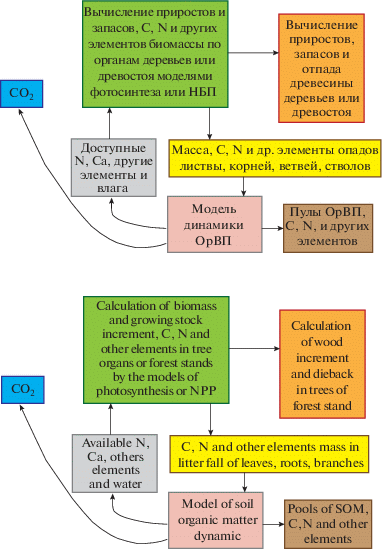
Ядром всех моделей продуктивности растительности и наземных экосистем является модель роста отдельного растения, либо популяции или сообщества растений. В последнем случае рассматриваются горизонтально распределенные по площади и вертикальным ярусам компартменты листьев, ветвей, стеблей/стволов, плодов/семян и корней. Эти модели могут сильно различаться по своей структуре и сложности: от эмпирических регрессионных уравнений, учитывающих ограниченное количество факторов роста [20], до процессных моделей фотосинтеза и дыхания, в которых учтено влияние большого количества факторов и ресурсов роста и развития растений [37]. Факторы и ресурсы роста могут учитываться с помощью константных величин (например, климат и эдафические условия), внешними сценариями (метеорологические ряды наблюдений) либо сложными процедурами и моделями (локальная освещенность, водно-тепловой режим почв, динамика органического вещества почв для вычисления элементов питания и эмиссии СО2, конкуренция за ресурсы, режимы нарушений и использования). Общая структура таких “процессных моделей” (на примере лесного сообщества) представлена на рис. 1. Как правило, фотосинтез вычисляется по принципу редукции теоретически максимально возможной в данном климате для данного вида величины с помощью функций отклика на влияние внешних факторов и наличных ресурсов. Разложение растительных остатков и гумуса оценивается почвенными моделями минерализации и гумификации органического вещества.
Модели травяных и агроэкосистем для многолетних растительных сообществ лугов и пастбищ и однолетних посевов культурных растений являются практически-ориентированными инструментами прогноза и планирования в природопользовании и в сельском хозяйстве [32, 38]. Модели продуктивности травяных экосистем имеют средне- или долгосрочный уровень прогнозирования продуктивности различных по площади растительных сообществ. Модели агроэкосистем прогнозируют динамику роста посевов чаще всего с дневным шагом в течение одного вегетационного периода в пределах отдельного поля.
Примером современной имитационной модели травяных растительных сообществ может служить модель Century [39]. Она вычисляет продуктивность и динамику растительного покрова (а не отдельных растений) с применением модели фотосинтеза и оценкой пулов и потоков углерода, азота, фосфора и серы в биомассе растений и в почве. В модели учитываются метеорологические и эдафические факторы роста и влияние хозяйственной деятельности. Ядром этой модели являются процедуры расчета продукции биомассы растений (с имитацией фотосинтеза и дыхания) и одноименная модель минерализации растительных остатков и формирования пулов доступных элементов питания в почве. Эта модель была калибрована для широкого использования в различных природных зонах Америки и других континентов. Были также созданы отдельная лесная и детальная сельскохозяйственная версия Daycent [40].
Развитие динамического моделирования продуктивности агроэкосистем началось во второй половине ХХ в. [41–43]. Российская модель продуктивности агроэкосистем AGROTOOL [32, 44] является экофизиологической моделью формирования урожая с модулями фотосинтеза, дыхания и разложения органического вещества почвы. Модель оценивает баланс органического вещества биомассы и урожая растений и динамику органического вещества почвы в течение вегетационного периода в зависимости от свойств почвы, метеорологических условий, применения удобрений и агротехники.
Модель продуктивности агроэкосистем DNDC [45, 46] представляет собою подход к оценке продуктивности с особым вниманием к динамике азота. В этой модели в дополнение к процедурам фотосинтеза и деструкции органического вещества имеется детальный модуль денитрификации и эмиссии окислов азота из почвы, что сделало ее особенно востребованной в начале XXI в. в связи с проблемами охраны окружающей среды. Была создана отдельная версия этой модели, UK-DNDC, калиброванная для Великобритании [47]. Для территории России верификация модели DNDC выполнена О.Э. Суховеевой [48, 49] с целью оценки параметров углеродного и азотного обмена в пахотных почвах Нечерноземья.
Практическая направленность моделей продуктивности агроэкосистем нашла выражение в создании моделей уровня фермы или сельскохозяйственного предприятия с экономическими и природоохранными модулями для принятия оперативных агротехнических решений. Так, модель OVERSEER [50] имитирует баланс большого количества элементов питания для максимизации продуктивности. Модель PASIM [51] создана для количественной оценки продуктивности пастбищ с расчетом не только продуктивности, но и влияния пастбищной нагрузки на эмиссию СО2 и изменение климата.
Общая тенденция в развитии моделей агроэкосистем – это переход от собственно моделей к “агросистемам” (Agro-Systems) управления продуктивностью (урожаем). Это стало актуальным в связи с развитием концепции и методологии “точного земледелия” [52] на основе математического моделирования и интеллектуальных систем принятия решений. Успешным примером такого подхода является австралийская система APSIM [38, 53]. Эта модель агроэкосистемы постоянно развивается за счет ассимиляции новых научных данных и обратной связи с пользователями в большом количестве стран. Система имеет возможность вычисления динамики культурной травяной и древесной растительности, фауны, почвы с дневным шагом при различных хозяйственных (managerial) сценариях с широким охватом природных и экономических условий. Стоит отметить, что тенденция к формированию такой системы прослеживается и у российских ученых [32].
Лесные модели, в отличие от агрономических, где рост вычисляется уравнениями фотосинтеза и дыхания, рассчитывают рост растений различными способами: с помощью традиционных ТХР, эмпирических уравнений прироста биомассы (нетто биологической продуктивности, NPP) и функций фотосинтеза и дыхания. Фактически ТХР и кривые хода роста с рассчитанными по ним функциями были первыми моделями продуктивности растительности. Современные модели также могут использовать ТХР как обобщенную модель прироста высоты, диаметра и древесины ствола в насаждениях. Это так называемые growth and yield models для вычисления прироста древесины [54, 55]. Более сложная процедура используется в “индивидуально-ориентированных” (individual-based) моделях с определением прироста каждого дерева в сообществе. В них базовый прирост дерева по ТХР модифицируется по индексам конкуренции с соседними деревьями [54, 56] либо в зависимости от локальной освещенности дерева [57, 58]. Такие модели используются в среднесрочной и долгосрочной перспективе при различных лесохозяйственных сценариях [54, 59–61]. Особо следует отметить известную канадскую модель CBM-CFS [62, 63], в которой присутствует подмодель динамики почвы. Эта модель была использована в Канаде на национальном уровне с прогонами модели (model runs) по 70 000 выделов канадской инвентаризации лесов для оценки динамики продуктивности и компонентов углеродного баланса под воздействием рубок, пожаров и изменения климата.
Процессные модели лесных экосистем, в отличие от рассмотренных выше моделей древостоев, имеют в своем ядре эколого-физиологические модели фотосинтеза и дыхания или экологические модели прироста биомассы (нетто биологической продуктивности, net primary productivity – NPP) популяции древесных растений или каждого дерева. Они начали создаваться в 70-х годах ХХ в., когда появились эколого-физиологические и “гэп” модели (gap models) [9, 34]. В лесных эколого-физиологических моделях прирост биомассы вычислялся, как и в травяных экосистемах, с помощью достаточно простых моделей фотосинтеза и дыхания на основе видоспецифичных параметров максимального фотосинтеза с его редукцией по конкретным условиям [37]. В гэп-моделях воспроизводилась восстановительная “гэп-динамика” в разновозрастных лесах как смена поколений после отмирания доминирующего дерева и формирования окон возобновления (gap), в которых вычислялся рост деревьев по эмпирическим функциям прироста биомассы (NPP) в зависимости от затенения и трофности почвы [64]. В настоящее время оба подхода слились в “индивидуально-ориентированные пространственно-распределенные” модели (individual-based spatially explicit models) [12], в которых рассматривается рост каждого дерева с точным заданием его местоположения в древостое.
Для компиляции моделей этого класса потребовалось создание процедур и отдельных моделей компонентов экосистем на основе имеющихся экспериментальных данных. Прежде всего это модели роста биомассы дерева, в которых общий прирост биомассы вычисляется а) по эколого-физиологическим функциям фотосинтеза и дыхания [37, 65–68]; б) по экологическим функциям расчета NPP, например по удельной биологической продуктивности единицы биомассы листвы/хвои при полной освещенности [9, 35, 69, 70].
Значительно меньше внимания уделяется моделированию растений травяно-кустарничкового яруса в лесных сообществах, несмотря на существование моделей травяных сообществ. Перспективными являются клеточно-автоматные модели, которые могут воспроизводить рост биомассы и пространственную динамику отдельных растений напочвенного покрова [71, 72, 100]. Отдельное направление в моделировании растений травяно-кустарничкового яруса занимает определение компонентов биоразнообразия растительности по эколого-ценотическим группам растений в зависимости от трофности почв и освещенности под пологом леса. Это система BioCalc [73, 74], которая работает в связке с моделью EFIMOD [34, 70].
Процедуры и модели динамики органического вещества почв (ОрВП) необходимы при моделировании наземных экосистем, во-первых, при расчете баланса углерода в системе “растительность–почва” с оценкой эмиссии CO2 в атмосферу [9, 63, 75]. Во-вторых, они нужны в процессных моделях с обратной связью от почвы к растительности для определения количества доступных для роста растений элементов питания, например, как в модели ROMUL [76]; ее последняя версия Romul_Hum [77] интегрирована в процессную модель EFIMOD [34, 70].
Процессные индивидуально-ориентированные пространственно-распределенные модели представляют собою наиболее “продвинутую” разновидность всех моделей, которые позволяют количественно описывать как рост каждого растения, так и динамику продуктивности всей растительности. К этому типу относится модель авторов IMPACT [69] и система моделей EFIMOD 2 [9], включающая в себя подмодель роста дерева, почвенную модель ROMUL [76] с генератором почвенного климата SCLISS [78], обобщающий популяционный модуль и блок имитации хозяйственных воздействий и нарушений (рубки, пожары и инвазии насекомых). Эти модели использовались для решения экологических задач (естественная динамика, лесопользование, лесные пожары, изменение климата) от России до Канады [9, 79–81].
Все рассмотренные выше модели относятся к масштабу растительного сообщества. Ландшафтные модели следующего пространственного уровня включают в себя мозаику сообществ, и в их ядре всегда присутствуют модели растительного сообщества [12, 68, 82, 83]. В основе методологии существующих ландшафтных моделей лежит разделение территории по пространственной решетке, что превращает естественные ландшафтные контуры разной конфигурации в набор клеток, принимаемых за гомогенные единицы по всем своим параметрам. Далее по ГИС методологии производится накладка на эту пространственную решетку тематических карт и баз данных с загрузкой параметров этих карт в характеристику каждой клетки. В этих пространственных единицах производится агрегация в одну систему моделей климата, рельефа, водного режима, почв, растительности, режима хозяйственного использования и нарушений с учетом взаимодействия между отдельными клетками. Теоретический фундамент научного ландшафтоведения в существующих ландшафтных моделях никак не используется, хотя эти модели и могут воспроизводить сукцессионные процессы в ландшафтах [84]. В методологическом плане различие между ландшафтными и региональными моделями заключается лишь в масштабах генерализации данных при увеличении моделируемой территории и размера клеток. В категорию ландшафтных и региональных моделей входит также математико-картографическое моделирование больших лесных территорий с оценкой потенциальной продуктивности лесных экосистем и ресурсного потенциала для оптимизации лесо- и природопользования на Дальнем Востоке [85, 86].
Существующие модели продуктивности и динамики лесной растительности используются в основном в исследовательских целях, например при параметризации сукцессий [12]. Они оказались эффективными при количественной оценке долгосрочных изменений растительности и почв в лесном хозяйстве [9, 11, 58, 61, 87]. Кроме того, лесные модели часто используются для оценки баланса углерода в лесных экосистемах при изменении климата [62, 63, 88, 89], загрязнении окружающей среды [79, 90] и пожарах [80, 81, 91, 92].
Как и при моделировании агроэкосистем, в лесном хозяйстве стали появляться системы поддержки принятия лесохозяйственных решений на основе лесных моделей. Примером может служить финская обширная система MELA [93], в ядро которой встроена лесная модель MOTTI [60, 61] с вычислением динамики продуктивности лесных экосистем при различных режимах ведения лесного хозяйства и экономических сценариях. Элементы такой системы присутствуют также в Канадской модели FORECAST [94] и Российской FORRUS [95]. Более того, процессная модель IMPACT была интегрирована в систему экологической сертификации лесных продуктов Финляндии для количественной оценки изъятия из экосистем углерода и азота с лесными продуктами [69].
Существует также значительное количество глобальных моделей практически всех типов наземных экосистем. Структура глобальных моделей растительности (dynamic global vegetation models, DGVMs) совпадает с таковой экофизиологических моделей растительного сообщества. Эти модели строятся с генерализацией исходных параметров по пространственной решетке относительно больших размеров (например, 2° × 2°) для глобальных оценок динамики растительности и почв преимущественно при изменении климата [96–98]. Такая генерализация не является оптимальной, поскольку она, безусловно, нивелирует сложную ландшафтную структуру и мозаику растительных сообществ и почв внутри каждой достаточно крупной клетки, определяемую не только естественными, но и антропогенными факторами. Поэтому глобальные оценки на основе этих моделей являются приблизительными. Так, одна из последних глобальных моделей, созданая в Академии наук Китая в рамках общей глобальной модели Земли (Earth System Model, CAS-ESM), вдвое переоценивает продуктивность растительности [99].
Рис. 1.
Структура процессной модели растительных сообществ на примере лесной модели (модифицировано из Моделирование динамики…, 2007) [9]. С – углерод, N – азот, Са – кальций, НБП – нетто биологическая продуктивность (NPP), ОрВП – органическое вещество почв, СО2 – углекислый газ. Fig. 1. Flow chart of the vegetation community model structure, by the example of forest model (adopted from Modelirovanie dinamiki…, 2007) [9]. C – carbon, N – nitrogen, Ca – calcium, NPP – net primary productivity, SOM – soil organic matter; СО2 – carbon dioxide.
ЗАКЛЮЧЕНИЕ
В настоящее время разработан достаточно обширный арсенал математических моделей для количественной оценки продуктивности разных типов растительности. Индивидуально- и пространственно-ориентированные модели (individual-based spatially explicit models) представляются наиболее перспективным инструментом, который может успешно применяться для создания локальных, ландшафтных и региональных систем поддержки принятия решений в лесоводстве. Процессные (process-based) модели являются основой точного земледелия и позволяют принимать управленческие решения, ориентируясь на объективные характеристики состояния почвы, ее механического и химического состава, биофизические характеристики урожая и прогноз изменения метеоусловий.
Большим достоинством динамического моделирования является возможность модернизации моделей по мере накопления новых научных данных с соответствующим увеличением их точности и расширением областей применения. Тенденция к формированию систем поддержки принятия решений в агрономии и лесном хозяйстве, в которых важнейшим компонентом являются динамические модели продуктивности наземных экосистем, свидетельствует о переходе динамического моделирования на уровень непосредственного практического использования.
Можно с уверенностью заключить о перспективности развития и применения динамических моделей продуктивности наземных экосистем для решения теоретических проблем динамики растительности в качестве современного инструмента для количественного прогноза в экологических и ресурсных оценках при принятии стратегических решений и реализации концепции наукоемкой биоэкономики.
Список литературы
Somogyi Z. 2016. A framework for quantifying environmental sustainability – Ecol. Indicators. 61: 338–345. https://doi.org/https://doi.org/10.1016/j.ecolind.2015.09.034
Бобылев С.Н., Михайлова С.Ю., Кирюшин П.А. 2014. Биоэкономика: проблемы становления. – Экономика. Налоги. Право. 6: 20–24. http://www.fa.ru/org/div/edition/enp/journals/2014%20%E2%84%966.pdf
Bukvareva E.N., Grunewald K., Bobylev S.N., Zamolodchikov D.G., Zimenko A.V., Bastian O. 2015. The current state of knowledge of ecosystems and ecosystem services in Russia: A status report. – AMBIO. 44(6): 491–507. https://doi.org/https://doi.org/10.1007/s13280-015-0674-4
Благовидов Н.Л. 1962. Качественная оценка земель и их рациональное использование. Л. 165 с.
Семенов В.А. 1970. Качественная оценка сельскохозяйственных земель. Л. 160 с.
Карманов И.И., Булгаков Д.С. 2012. Методика почвенно-агроклиматической оценки пахотных земель для кадастра. М. 120 с.
Орлов М.М. 1927. Лесоустройство. Элементы лесного хозяйства. Л. 428 с.
Швиденко А.З., Щепащенко Д.Г., Нильссон С., Булуй Ю.И. 2008. Таблицы и модели хода роста и продуктивности насаждений основных лесообразующих пород Северной Евразии. М. 886 с. http://webarchive.iiasa.ac.at/Research/FOR/forest_cdrom/Articles/THR.pdf
Моделирование динамики органического вещества в лесных экосистемах. 2007. М. 380 с.
Ågren G., Bosatta E. 1996. Theoretical ecosystem ecology: Understanding element cycling. Cambridge. 234 p.
Larocque G.R., Komarov A., Chertov O., Shanin V., Liu J., Bhatti J.S., Wang W., Peng C., Shugart H.H., Xi W., Holm J.A. 2016a. Process-based models: A synthesis of models and applications to address environmental and management issues. – In: G. R. Larocque (Ed.). Ecological Forest Management Handbook. Boca Raton, FL, USA. P. 223–266. https://www.taylorfrancis.com/books/9780429188787/chapters/10.1201/b19150-14
Larocque G.R., Shugart H.H., Xi W., Holm J.A. 2016b. Forest succession models. – In: G.R. Larocque (Ed.). Ecological Forest Management Handbook. Boca Raton, FL, USA. P. 179–221. https://www.taylorfrancis.com/books/9780429188787/chapters/10.1201/b19150-13
Жукова Л.А., Дорогова Ю.А., Турмухаметова Н.В., Гаврилова М.Н., Полянская Т.А. 2010. Экологические шкалы и методы анализа экологического разнообразия растений. Йошкар-Ола. 368 с.
Раменский Л.Г. 1938. Введение в комплексное почвенно-геоботаническое исследование земель. М. 620 с.
Сукачёв В.Н. 1972. Основы лесной типологии и биогеоценологии. Л. 419 с.
Погребняк П.С. 1955. Основы лесной типологии. Киев. 456 с.
Федорчук В.Н., Нешатаев В.Ю., Кузнецова М.Л. 2005. Лесные экосистемы северо-западных районов России: Типология, динамика, хозяйственные особенности. СПб. 382 с. http://www.twirpx.com/file/1445527/
Шишов Л.Л., Дурманов Д.Н., Карманов И.И., Ефремов В.В. 1991. Теоретические основы и пути регулирования плодородия почв. М. 305 с.
Зеликов В.Д. 1971. Почвы и бонитет насаждений. М. 119 с.
Чертов О.Г. 1981. Экология лесных земель. Почвенно-экологическое исследование лесных местообитаний. Л. 192 с.
Cotta H. 1828. Anweisung zum Waldbau. Dresden und Leipzig. (1. Auflage in 1817). 431 p. http://www.mdz-nbn-resolving.de/urn/resolver.pl?urn=urn:nbn:de:bvb:12-bsb10294676-2
Сеннов С.Н. 1996. Тенденции роста южно-таежных лесов северо-запада. – Лесоведение. 4: 68–71.
Сеннов С Н. 1999. Итоги 60-летних наблюдений за естественной динамикой леса. СПб. 98 с.
Сеннов С.Н. 2008. Результаты длительных наблюдений за естественным ростом древостоев и влиянием на него рубок ухода. СПб. 29 с.
Кузмичев В.В. 1977. Закономерности роста древостоев. Новосибирск. 160 с.
Рогозин М.В., Разин Г.С. 2015. Модели динамики и моделирование развития древостоев. – Сиб. лесн. журн. 2: 55–70. https://doi.org/https://doi.org/10.15372/SJFS20150205
Rogozin M.V., Razin G.S. 2015. Development of forest stands. Models, laws, hypotheses. Raleigh, NC, USA.196 p.
Итоги экспериментальных работ в лесной опытной даче ТСХА за 1862–1962 годы. 1964. М. 562 с.
Галицкий В.В., Глотов Н.В., Комаров А.С., Тюрюканов А.Н. 1975. Естественно-исторические, генетические и математические аспекты биопродуктивности. В сб. Имитационное моделирование и экология. М. 18–25.
Одум Ю. 1975. Основы экологии. М. 742 с.
Риклефс Р. 1979. Основы общей экологии. М. 424 с.
Полуэктов Р.А., Смоляр Э.И., Терлеев В.В., Топаж А.Г. 2006. Модели продукционного процесса сельскохозяйственных культур. СПб. 396 с.
Korzukhin M.D., Ter-Mikaelian M.T., Wagner R.G. 1996. Process versus empirical models: Which approach for forest ecosystem management? – Can. J. Forest Res. 26(5): 879–887. https://doi.org/https://doi.org/10.1139/x26-096
Chertov O.G., Komarov A.S., Tsiplianovsky A.V. 1999. A combined simulation model of Scots pine, Norway spruce and Silver birch ecosystems in European boreal zone. – Forest Ecol. Manag. 116(1–3): 189–206. https://doi.org/https://doi.org/10.1016/S0378-1127(98)00456-3
Porté A., Bartelink H.H. 2002. Modelling mixed forest growth: A review of models for forest management. – Ecol. Model. 150(1–2): 141–188. https://doi.org/https://doi.org/10.1016/S0304-3800(01)00476-8
Keating B.A., Carberry P.S., Hammer G.L., et al. 2003. An overview of APSIM, a model designed for farming systems simulation. – European J. Agronomy. 18(3–4): 267–288. https://doi.org/https://doi.org/10.1016/S1161-0301(02)00108-9
Корзухин М.Д., Цельникер Ю.Л., Семенов С.М. 2008. Экофизиологическая модель первичной продуктивности древесных растений и оценки климатических пределов их произрастания. – Метеорология и гидрология. 12: 56–69.
Holzworth D.P., Huth N.I., deVoil P.G. et al., totally 46 authors. 2014. APSIM – Evolution towards a new generation of agricultural systems simulation. – Environmental Modelling & Software. 62: 327–350. https://doi.org/https://doi.org/10.1016/j.envsoft.2014.07.009
Parton W.J., Stewart J.W.B., Cole C.V. 1988. Dynamics of C, N, P and S in grasslands soils: A model. – Biogeochemistry. 5(1): 109–131. https://doi.org/https://doi.org/10.1007/BF02180320
Parton W.J., Hartman M., Ojima D., Schimel D. 1998. DAYCENT and its land surface submodel: Description and testing. Glob. – Planet. Change. 19(1–4): 35–48. https://doi.org/https://doi.org/10.1016/S0921-8181(98)00040-X
Сиротенко О.Д. 1981. Математическое моделирование водно-теплового режима и продуктивности агроэкосистем. Л. 167 с.
Бондаренко Н.Ф., Жуковский Е.Е., Мушкин И.Г., Нерпин С.В., Полуэктов Р.А., Усков И.Б. 1982. Моделирование продуктивности агроэкосистем. Л. 347 с.
Полуэктов Р.А., Пых Ю.А., Швытов И.А. 1982. Динамические модели экологических систем Л. 288 с.
Poluektov R.A., Fintushal S.M., Oparina I.V., Shatskikh D.V., Terleev V.V., Zakharova E.T. 2002. AGROTOOL – a system for crop simulation. – Archives of Agronomy and Soil Science. 48(6): 609–635. https://doi.org/https://doi.org/10.1080/0365034021000041597. URL: http://agrotool.ru/
Zhang Y., Li C.S., Zhou X.J., Moore B. 2002. A simulation model linking crop growth and soil biogeochemistry for sustainable agriculture. – Ecol. Model. 151(1): 75–108. https://doi.org/https://doi.org/10.1016/S0304-3800(01)00527-0
Li S., Jin X., Fan X., Huang W., Cao Z. 2010. Ruminant production and carbon emission reduction measures. – Chin. J. Anim. Nutr. 22: 2–9.
Brown L., Syed B., Jarvis S.C., Sneath R.W., Phillips V.R., Goulding K.W.T., Li C. 2002. Development and application of a mechanistic model to estimate emission of nitrous oxide from UK agriculture. – Atmos. Environ. 36(6): 917–928. https://doi.org/https://doi.org/10.1016/S1352-2310(01)00512-X
Суховеева О.Э. 2018а. Приложение модели DNDC к оценке параметров углеродного и азотного обмена в пахотных почвах Нечерноземья. – Изв. РАН. Серия географическая. 2: 74–85. https://doi.org/https://doi.org/10.7868/S2587556618020073
Суховеева О.Э. 2018б. Верификация модели DNDC для оценки параметров углеродного и азотного обмена в пахотных почвах Нечерноземья. – Изв. РАН. Серия географическая. 4: 89–95.
Wheeler D.M., Ledgard S.F., DeKlein C.A.M. 2008. Using the OVERSEER nutrient budget model to estimate on-farm greenhouse gas emissions. – Aust. J. Exp. Agric. 48: 99–103. https://doi.org/https://doi.org/10.1071/EA07250
Riedo M., Gyalistras D., Fuhrer J. 2000. Net primary production and carbon stocks in differently managed grasslands: Simulation of site-specific sensitivity to an increase in atmospheric CO2 and to climate change. – Ecol. Model. 134(2–3): 207–227. https://doi.org/https://doi.org/10.1016/S0304-3800(00)00356-2
Srinivasan A. 2006. Handbook of Precision Agriculture: Principles and Applications. N.Y. 568 p. https://doi.org/https://doi.org/10.1201/9781482277968
APSIM Initiative. 2018. URL http://www.apsim.info//
Weiskittel A.R., Hann D.W., Kershaw J.A.Jr., Vanclay J.K. 2011. Forest growth and yield modeling. Blackwell. 430 p.
Schneider R., Franceschini T., Fortin M., Martin-Ducup O., Gauthray-Guyénet V., Larocque G.R., Marshall P., Bérubé-Deschênes A. 2016. Growth and yield models for predicting tree and stand productivity. – In: G. R. Larocque (Ed.). Ecological Forest Management Handbook. Boca Raton, FL, USA. P. 141–178. https://doi.org/https://doi.org/10.1201/b19150
Daniels R.F., Burkhart H.E., Clason T.R. 1986. A comparison of competition measures for predicting growth of loblolly pine trees. – Canad. J. For. Research. 16: 1230–1237. https://doi.org/https://doi.org/10.1139/x86-218
Чумаченко С.И. 1993. Базовая модель динамики многовидового разновозрастного лесного ценоза. – В кн.: Вопросы экологии и моделирования лесных экосистем. Науч. труды МЛТИ. М. 248: 147–180.
Chumachenko S.I., Korotkov V.N., Palenova M.M., Politov D.V. 2003. Simulation modeling of long-term stand dynamics at different scenarios of forest management for coniferous-broad-leaved forests. – Ecol. Model. 170(2–3): 345–362. https://doi.org/https://doi.org/10.1016/S0304-3800(03)00238-2
Gobakken L.R., Mattson J., Alfredsen G. 2008. In-service performance of wood depends upon the critical in-situ conditions. IRG/WP 08-20382. The Intern. Res. Group Wood Protection. Stockholm, Sweden. 13 p.
Hynynen J., Ojansuu R., Hökkä H., Siipilehto J., Salminen H., Haapala P. 2002. Models for predicting stand development in MELA system. – Finnish Forest Res. Inst. Res. Papers. 835: 1–16. http://www.metla.fi/julkaisut/mt/2002/835.htm
Hynynen J., Ahtikoski A., Siitonen J., Sievänen R., Liski J. 2005. Applying the MOTTI simulator to analyse the effects of alternative management schedules on timber and non-timber production. – For. Ecol. Manag. 207(1–2): 5–18. https://doi.org/https://doi.org/10.1016/j.foreco.2004.10.015
Kurz W.A., Apps M.J. 1999. A 70-year retrospective analysis of carbon fluxes in the Canadian forest sector. – Ecol. Appl. 9(2): 526–547. http://www.cfs.nrcan.gc.ca/bookstore_pdfs/18664cannotpostonline.pdf
Kurz W.A., Dymond C.C., White T.M., Stinson G., Shaw C.H., Rampley G.J., Smyth C., Simpson B.N., Neilson E.T., Trofymow J.A., Metsaranta J., Apps M.J. 2009. CBM-CFS3: A model of carbon-dynamics in forestry and land-use change implementing IPCC standards. – Ecol. Model. 220(4): 480–504. https://doi.org/https://doi.org/10.1016/j.ecolmodel.2008.10.018
Shugart H.H. 1984. Theory of Forest dynamics. Berlin. 278 p.
Бугровский В.В., Дудин Е.В., Меллина Е.Г., Цельникер Ю.Л. 1982. Моделирование продукционных процессов в чистых древостоях. – Журн. общ. биологии. 43(4): 480–488.
Березовская Ф.С., Карев Г.П., Швиденко А.З. 1991. Моделирование динамики древостоев: эколого-физиологический подход. М. 84 с.
Perttunen J. 2009. The LIGNUM Functional-Structural Tree Model, Doctoral thesis. Helsinki. 52 p.
Seidl R., Rammer W., Scheller R.M., Spies T.A. 2012. An individual-based process model to simulate landscape-scale forest ecosystem dynamics. – Ecol. Model. 231: 87–100. https://doi.org/https://doi.org/10.1016/j.ecolmodel.2012.02.015
Chertov O., Komarov A., Kolström M., Pitkänen S., Strandman H., Zudin S., Kellomäki S. 2003. Modelling the long-term dynamics of populations and communities of trees in boreal forests based on competition on light and nitrogen. – For. Ecol. Manag. 176(1): 355–369. https://doi.org/https://doi.org/10.1016/S0378-1127(02)00284-0
Komarov A., Chertov O., Zudin S., Nadporozhskaya M., Mikhailov A., Bykhovets S., Zudina E., Zoubkova E. 2003. EFIMOD 2 – A model of growth and cycling of elements in boreal forest ecosystems. – Ecol. Model. 170(2–3): 373–392. https://doi.org/https://doi.org/10.1016/S0304-3800(03)00240-0
Комаров A.C. 1988. Математическое моделирование ценопопуляций растений. – В кн.: Ценопопуляции растений. Очерки популяционной биологии растений. М. С. 137–155.
Frolov P.V., Zubkova E.V., Komarov A.S. 2015. A cellular automata model for a community comprising two plant species of different growth forms. – Biology Bulletin. 42(4): 279–286. https://doi.org/https://doi.org/10.1134/S1062359015040044
Ханина Л.Г., Бобровский М.В., Комаров А.С., Михайлов А.В., Быховец С.С., Лукьянов А.М. 2006. Моделирование динамики разнообразия лесного напочвенного покрова. – Лесоведение. 1: 70–80.
Khanina L., Bobrovsky M., Komarov A., Mikhailov A. 2007. Modeling dynamics of forest ground vegetation diversity under different forest management regimes. – For. Ecol. Manag. 248(1-2): 80–94. https://doi.org/https://doi.org/10.1016/j.foreco.2007.03.021
Schelhaas M.J., Eggers J., Lindner M., Nabuurs G.J., Pussinen A., Päivinen R., Schuck A., Verkerk P.J., van der Werf D.C., Zudin S. 2007. Model documentation for the European Forest Information Scenario model (EFISCEN 3.1.3). – EFI Technical Report. 26: 1–118 p. http://www.fefr.org/files/attachments/publications/alterrarapport1559.pdf
Chertov O.G., Komarov A.S., Nadporozhskaya M.A., Bykhovets S.A., Zudin S.L. 2001. ROMUL – a model of forest soil organic matter dynamics as a substantial tool for forest ecosystem modelling. – Ecol. Model. 138: 289–308. https://doi.org/https://doi.org/10.1016/S0304-3800(00)00409-9
Komarov A., Chertov O., Bykhovets S., Shaw C., Nadporozhskaya M., Frolov P., Shashkov M., Shanin V., Grabarnik P., Priputina I., Zubkova E. 2017. Romul_Hum model of soil organic matter formation coupled with soil biota activity. I. Problem formulation, model description, and testing. – Ecol. Model. 345: 113–124. https://doi.org/https://doi.org/10.1016/j.ecolmodel.2016.08.007
Быховец С.С., Комаров А.С. 2002. Простой статистический имитатор климата почвы с месячным шагом – Почвоведение. 4: 443–452.
Kahle H.-P., Karjalainen T., Schuck A., Ågren G., Kellomäki S., Mellert K., Prietzel J., Rehfuess K.E., Spiecker H. 2008. Causes and Consequences of Forest Growth Trends in Europe – Results of the RECOGNITION Project. European Forest Institute Res. Rep. 21. Leiden; Boston; Köln. 261 p.
Bhatti J., Chertov O., Komarov A. 2009. Influence of climate change, fire, insect and harvest on C dynamics for jack pine in central Canada: simulation approach with the EFIMOD model. – Intern. J. Climate Change. Impacts and Responses. 1: 43–61. https://doi.org/https://doi.org/10.18848/1835-7156/CGP/v01i03/37265
Shanin V.N., Komarov A.S., Mikhailov A.V., Bykhovets S.S. 2011. Modelling carbon and nitrogen dynamics in forest ecosystems of Central Russia under different climate change scenarios and forest management regimes. – Ecol. Model. 222(14): 2262–2275. https://doi.org/https://doi.org/10.1016/j.ecolmodel.2010.11.009
He H.S., Mladenoff D.J., Boeder J. 1999. An object-oriented forest landscape model and its representation of tree species. – Ecol. Model. 119(1): 1–19. https://doi.org/https://doi.org/10.1016/S0304-3800(99)00041-1
Lexer M.J., Hönninger K. 2001. A modified 3D-patch model for spatially explicit simulations of vegetation composition in heterogeneous landscapes. – For. Ecol. Manag. 144(1–3): 43–65. https://doi.org/https://doi.org/10.1016/S0378-1127(00)00386-8
Scheller R.M., Domingo J.B., Sturtevant B.R., Williams J.S., Rudy A., Gustafson E.J., Mladenoff D.J. 2007. Design, development, and application of LANDIS-II, a spatial landscape simulation model with flexible temporal and spatial resolution. – Ecol. Model. 201(3-4): 409–419. https://doi.org/https://doi.org/10.1016/j.ecolmodel.2006.10.009
Петропавловский Б.С. 2011. Математико-картографическое моделирование оптимальных мест произрастания лесообразующих пород (на примере Приморского края). – Сиб. экол. журн. 6: 767–772. http://www.sibran.ru/upload/iblock/74a/74a396a3d3c0972d583199d286d53aec.pdf
Петропавловский Б.С. 2014. Использование информационной статистики для задач картографического моделирования лесной растительности (на примере Приморского края) – В сб.: Проблемы устойчивого управления лесами Сибири и Дальнего Востока: Материалы Всерос. конф. Хабаровск. С. 75–77. http://dalniilh.ru/wp-content/uploads/2014/10/ПРОБЛЕМЫ-УСТОЙЧИВОГО-УПРАВЛЕНИЯ-ЛЕСАМИ-СИБИРИ-И-ДАЛЬНЕГО-ВОСТОКА-2014.pdf
Припутина И.В., Фролова Г.Г., Быховец С.С., Шанин В.Н., Лебедев В.Г., Шестибратов К.А. 2016. Моделирование продуктивности лесных плантаций при разных схемах пространственного размещения деревьев. – Математическая биология и биоинформатика. 11(2): 245–262. https://doi.org/https://doi.org/10.17537/2016.11.245
Замолодчиков Д.Г., Грабовский В.И., Краев Г.Н. 2011. Динамика бюджета углерода лесов России за два последних десятилетия. – Лесоведение. 6: 16–28.
Schelhaas M.J., van Esch P.W., Groen T.A., de Jong B.H.J., Kanninen M., Liski J., Masera O., Mohren G.M.J., Nabuurs G.J., Palosuo T., Pedroni L., Vallejo A., Vilén T. 2004. CO2FIX V 3.1 – A modelling framework for quantifying carbon sequestration in forest ecosystems. – Alterrarapport. 1068: 1–122. http://dataservices.efi.int/casfor/downloads/co2fix3_1_description.pdf
Priputina I., Zubkova E., Shanin V., Smirnov V., Komarov A. 2014. Evidence of plant biodiversity changes as a result of nitrogen deposition in permanent pine forest plots in central Russia. – Ecoscience. 21 (3–4): 286–300. https://doi.org/https://doi.org/10.2980/21-(3-4)-3681
Chertov O.G., Komarov A.S., Gryazkin A.V., Smirnov A.P., Bhatti D.S. 2013. Simulation modeling of the impact of forest fire on the carbon pool in coniferous forests of European Russia and Central Canada. – Contemp. Probl. Ecol. 6(7): 727–733. https://doi.org/https://doi.org/10.1134/S1995425513070032
Nadporozhskaya M.A., Chertov O.G., Bykhovets S.S., Shaw C.H., Maksimova E.Y., Abakumov E.V. 2018. Recurring surface fires cause soil degradation of forest land: A simulation experiment with the EFIMOD model. – Land Degradation Development. 29(7): 2222–2232. https://doi.org/https://doi.org/10.1002/ldr.3021
Hirvelä H., Härkönen K., Lempinen R., Salminen O. 2017. MELA2016 Reference Manual. Helsinki. 547 p. http://jukuri.luke.fi/handle/10024/538149
Kimmins J.P., Mailly D., Seely B. 1999. Modelling forest ecosystem net primary production: the hybrid simulation approach used in FORECAST. – Ecol. Model. 122(3): 195–224. https://doi.org/https://doi.org/10.1016/S0304-3800(99)00138-6
Чумаченко С.И., Паленова М.М., Коротков В.Н. 2001. Прогноз динамики таксационных показателей лесных насаждений при разных сценариях ведения лесного хозяйства: модель динамики лесных насаждений FORRUS-S – Экология, мониторинг и рациональное природопользование. Науч. тр. МГУЛ. 314: 128–146.
Sitch S., Smith B., Prentice I.C., Arneth A., Bondeau A., Cramer W., Kaplan J.O., Levis S., Lucht W., Sykes M.T., Thonicke K., Venevsky S. 2003. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ Dynamic Global Vegetation Model. – Global Change Biol. 9(2): 161–185. https://doi.org/https://doi.org/10.1046/j.1365-2486.2003.00569.x
Sato H., Itoh A., Kohyama T. 2007. SEIB–DGVM: A new Dynamic Global Vegetation Model using a spatially explicit individual-based approach. – Ecol. Model. 200(3–4): 279–307. https://doi.org/https://doi.org/10.1016/j.ecolmodel.2006.09.006
Beringer T., Lucht W., Schaphoff S. 2011. Bioenergy production potential of global biomass plantations under environmental and agricultural constraints. – GCB Bioenergy 3(4): 299–312. https://doi.org/https://doi.org/10.1111/j.1757-1707.2010.01088.x
Zhu J., Zeng X., Zhang M., Dai Y., Ji D., Li F., Zhang Q., Zhang H., Song X. 2018. Evaluation of the new dynamic global vegetation model in CAS-ESM. – Advances Atm. Sci. 35(6): 659–670. https://doi.org/https://doi.org/10.1007/s00376-017-7154-7
Комаров А.С., Зубкова Е.В., Фролов П.В. 2015. Клеточно-автоматная модель динамики популяций и сообществ кустарничков. – Сиб. лесн. журн. 3: 57–69.
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы