

Растительные ресурсы, 2019, T. 55, № 3, стр. 409-421
Видовой состав и запасы фитомассы напочвенного покрова в антропогенно нарушенных сосняках Красноярской лесостепи
И. А. Гончарова 1, *, Л. Н. Скрипальщикова 1, А. П. Барченков 1
1 Институт леса им. В.Н. Сукачева СО РАН, обособленное подразделение
Федерального исследовательского центра “Красноярский научный центр СО РАН”
г. Красноярск, Россия
* E-mail: iagoncharova007@mail.ru
Поступила в редакцию 14.05.2018
После доработки 10.10.2018
Принята к публикации 17.04.2019
Аннотация
Изучено современное состояние напочвенного покрова антропогенно нарушенных сосняков Красноярской лесостепи и дана оценка степени и характера его изменения за 15-летний период, прошедший с момента последних исследований. В настоящее время концентрации токсичных элементов в растениях не достигают избыточных значений, при которых происходит нарушение гомеостаза. В 2017 г. наибольший запас фитомассы живого напочвенного покрова был отмечен на контрольной пробной площади (357.88 ± 70.31 г/м2), и за исследуемый 15-летний период он существенно не изменился. На участках, подверженных антропогенной нагрузке, в настоящее время запас фитомассы в 1.4–3 раза ниже, чем в контрольном фитоценозе, однако за 15-летний период он увеличился. На основе анализа изменения видового разнообразия, количественного соотношения эколого-ценотических групп, структуры и абсолютного значения запаса фитомассы определены стадии деградации напочвенного покрова.
Проблема возрастающего антропогенного воздействия на экосистемы вызывает интерес во всем мире. Многочисленные исследования посвящены обсуждению различных аспектов данной проблемы [1–8]. Отмечено, что основными антропогенными факторами, воздействующими на лесные экосистемы в зоне влияния крупных городов, являются техногенное загрязнение и интенсивная рекреационная нагрузка.
Вопросу влияния антропогенных факторов на растительный покров посвящен целый ряд исследований и обзоров [9–14]. Отмечается [15, 16], что техногенные загрязнения в течение короткого времени (20–30 лет) приводят к уменьшению флористического богатства и преобразованию различных лесных сообществ в однотипные рудеральные. Под влиянием рекреационных нагрузок лесные экосистемы проходят несколько стадий деградации. Упрощается их структура, из состава нижних ярусов исчезают лесные виды, в результате чего формируется более устойчивый к вытаптыванию злаково-разнотравный покров [17].
Антропогенные факторы постоянно изменяются во времени как по источникам воздействия, так и по степени их влияния [18–20]. В связи с этим возникает необходимость мониторинга состояния напочвенного покрова, что определяет актуальность данного исследования.
Цель работы – изучить современное состояние напочвенного покрова антропогенно нарушенных сосняков Красноярской лесостепи, а также оценить степень и характер его изменения за 15-летний период, прошедший с момента последних исследований.
ОБЪЕКТЫ И МЕТОДЫ
Исследования проводились на четырех мониторинговых пробных площадях (ПП), заложенных в типичных для Красноярской лесостепи сосновых насаждениях (Pinus sylvestris L.) (рис. 1). По данным Государственных докладов (2017, 2018), в 2016 и 2017 г. в г. Красноярске и на прилегающих территориях от стационарных источников в атмосферу поступило 196.9 тыс. т загрязняющих веществ, от передвижных источников – 127.3 тыс. т. В загрязняющих атмосферу выбросах преобладают твердые (тяжелые металлы, фториды) и газообразные вещества (оксиды серы, азота и углерода).
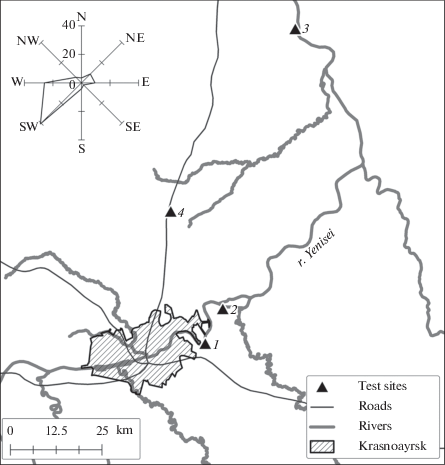
ПП 1 (Березовский бор) заложена в 10 км на юго-восток от г. Красноярска в сосняке осочково-разнотравном. Подлесок представлен как ярус с проективным покрытием 30% и образован Cotoneaster melanocarpus Fisch. ex A.Blytt, Rosa acicularis Lindl., Swida alba (L.) Opiz, Sorbus aucuparia subsp. sibirica (Hedl.) Krylov. и др. Проективное покрытие травяно-кустарничкового яруса 70%. Доминантами являются Carex macroura Meinsh., Polygonatum odoratum (Mill.) Druce, Thalictrum minus L., Calamagrostis arundinacea (L.) Roth, Vicia unijuga A. Br. Моховой покров в виде отдельных пятен Pleurozium schreberi (Willd. ex Brid.) Mitt.
ПП 2 (Есаульский бор) располагается в 30 км от г. Красноярска в северо-восточном направлении в сосняке разнотравно-осочково-зеленомошном. Подлесок представлен как ярус (проективное покрытие 80%), равномерно распределен по площади, образован Cotoneaster melanocarpus, Viburnum opulus L., Swida alba, Salix caprea L., Crataegus sanguinea Pall., Rosa acicularis и др. Проективное покрытие травяно-кустарничкового яруса составляет 60%. Доминантами и содоминантами являются Carex macroura, Rubus saxatilis L., Geranium sylvaticum L., Thalictrum minus, Polygonatum odoratum, Phlomoides tuberosa (L.) Moench. Проективное покрытие мхов 70%. Доминантом является Pleurozium schreberi. Присутствуют Hylocomium splendens (Hedw.) Schimp., Rhytidiadelphus triquetrus (Hedw.) Warnst., Helodium blandowii (F. Weber & D. Mohr) Warnst.
Березовский и Есаульский боры (ПП 1, 2) располагаются в направлении основного переноса промышленных выбросов города. Фитоценозы длительное время подвергались значительным техногенным и рекреационным нагрузкам.
ПП 3 (Юксеевский бор) располагается в 100 км от г. Красноярска в сосняке мелкотравно-зеленомошном. Подлесок равномерно распределен, занимает 10–15% площади, представлен Sorbus aucuparia subsp. sibirica, Padus avium, Rosa acicularis, Salix caprea и др. Проективное покрытие травяно-кустарничкового яруса составляет 90%. Доминанты и содоминанты травяно-кустарничкового яруса: Carex macroura, Pyrola rotundifolia L., Rubus saxatilis, Linnaea borealis L., Poa annua L., Carum carvi L., Elytrigia repens (L.) Nevski, Cimicifuga foetida L. Общее проективное покрытие мхов составляет 60%: Hylocomium splendens (50%) и Pleurozium schreberi (10%).
ПП 4 (Погорельский бор) заложена в сосняке разнотравно-зеленомошном, находящемся на расстоянии 40 км от города (контроль). Подлесок как ярус не выражен, представлен отдельными экземплярами Rosa acicularis, Padus avium, Sorbus aucuparia subsp. sibirica и др. Проективное покрытие травяно-кустарничкового яруса составляет 60%. Доминантами и содоминантами являются Calamagrostis arundinacea (L.) Roth, Vaccinium vitis-idaea L., Carex macroura, Rubus saxatilis. Проективное покрытие мохового покрова 90%. Доминантом является Pleurozium schreberi. Присутствуют в примеси Hylocomium splendens, Rhytidiadelphus triquetrus, Ptilium crista-castrensis, Dicranum polysetum.
Юксеевский и Погорельский боры (ПП 3 и 4) расположены вне основного направления переноса загрязняющих веществ. ПП 3 испытывает рекреационную и пасквальную нагрузки. На ПП 4 антропогенное воздействие носит спорадический характер.
Таксационная характеристика древесного яруса в изученных сообществах приведена в табл. 1.
Таблица 1.
Таксационные характеристики древостоев на пробных площадях Table 1. Mensurational characteristics of forest stands on sample plots
| Пробные
площади Sample plots |
Состав древостоя Stand composition | Средние Average |
Класс
бонитета Quality class |
Класс
возраста Age class |
Полнота
древостоя Stand density |
|
|---|---|---|---|---|---|---|
| Высота, м Height, m |
Диаметр, см Diameter, cm |
|||||
| ПП 1 (Березовка) Sample plot 1 (Berezovka) |
10С 10P |
16.1 | 19.8 | IV | V–VI | 0.6–0.8 |
| ПП 2 (Есаулово) Sample plot 2 (Esaulovo) |
10С 10P |
16.3 | 33.2 | III | VI | 0.8–1.0 |
| ПП 3 (Юксеево) Sample plot 3 (Yukseyevo) |
10С 10P |
29.8 | 35.8 | I-а | IV | 0.8 |
| ПП 4 (Погорелка) Sample plot 4 (Pogorelka) |
10СедЛ,Б 10P with sporadically larch and birch |
27.3 | 35.8 | I | VI | 0.9 |
Для изучения напочвенного покрова в августе 2017 г. в сосновых фитоценозах в пределах каждой ПП было заложено по 30 учетных площадок размером 1 м2, где проводили оценку видового состава, проективного покрытия видов и общего проективного покрытия травяно-кустарничкового яруса и мохового покрова по общепринятым методикам [21]. Классификация эколого-ценотических групп (ЭЦГ) видов проведена по Д.И. Назимовой и др. [22] и Т.Н. Буториной [23]. Для сравнительной оценки флористических списков применен коэффициент Сёренсена–Чекановского (Кsc). Степень видового разнообразия оценена с помощью индекса Шеннона [24].
Степень рекреационной трансформации живого напочвенного покрова определялась с использованием индекса синантропизации (X), который рассчитывался по формуле:
Для учета запаса фитомассы травяно-кустарничкового покрова на каждой пробной площади взяты укосы с 10 учетных площадок размером 20 × 25 см. Растения срезали на уровне подстилки, разбирали по видам, высушивали до абсолютно сухого состояния и взвешивали.
Влияние рекреационных нагрузок изучалось с применением оценочных шкал дигрессии [25, 26].
Степень техногенной нагрузки определялась по содержанию токсичных ингредиентов (цинк, свинец, кадмий, фтор) в хвое сосны и растениях живого напочвенного покрова (средний образец растений травяно-кустарничкового и мохово-лишайникового ярусов) в аккредитованной лаборатории (Аккредитованная испытательная лаборатория ФГБУ ГЦАС “Красноярский”) с использованием сертифицированных методик [27, 28].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Проведенные ранее исследования показали [29], что в зонах промышленного загрязнения в растениях напочвенного покрова накапливается в 10–100 раз больше микроэлементов, чем в фоновых районах. С начала 2000-х гг. на АО РусАЛ “Красноярский алюминиевый завод” начался переход на новую технологию (“Экологический Содерберг”), позволившую снизить объемы вредных для окружающей среды выбросов [30]. Исследования 2017 г. зафиксировали значительное снижение техногенных нагрузок (табл. 2). Превышение содержания свинца в растительном материале по сравнению с контрольными (ПП 4) значениями на ПП 1 в настоящее время составляет 4.8 крат, на ПП 2 – 27 крат. Содержание фтора в растительном материале на ПП 1 и 2 превышено соответственно в 3 и 2.6 раза по сравнению с контрольными величинами. Сравнение с литературными данными показало, что на данный момент концентрации токсичных элементов в растениях не достигают избыточных значений, при которых происходит нарушение гомеостаза. Это позволяет предположить, что изменения в напочвенном покрове, отмеченные в 2017 г., в большей мере обусловлены рекреационной, нежели техногенной нагрузкой.
Таблица 2.
Концентрации элементов в растительном материале (мг/кг сухой массы). (Данные анализа смешанного образца растений напочвенного покрова 2017 г.) Table 2. The elements content in plant matter (mg/kg of dry weight). Mixed plants sample of 2017
| Элемент Element |
Концентрация элементов в растительном
материале в исследуемых ценозах The elements content in plant matter of the sample plots |
Содержание элементов [35] The elements content [35] | ||||
|---|---|---|---|---|---|---|
| ПП 1 Sample plot 1 |
ПП 2 Sample plot 2 |
ПП 3 Sample plot 3 |
ПП 4 Sample plot 4 |
Достаточное Sufficient |
Избыточное Excessive |
|
| Zn | 20.8 ± 4.2 | 48.5 ± 9.7 | 32.1 ± 6.4 | 34.8 ± 7.0 | 27–150 | 100–400 |
| Pb | 2.08 ± 0.52 | 11.7 ± 2.9 | 0.76 ± 0.19 | 0.43 ± 0.11 | 5–10 | 30–300 |
| Cd | <0.05 | 0.22 ± 0.11 | 0.062 ± 0.031 | 0.073 ± 0.037 | 0.05–0.2 | 5–30 |
| F | 14.6 ± 1.5 | 12.6 ± 1.3 | 1.56 ± 0.23 | 4.83 ± 0.72 | 5–30 | 50–500 |
В 2017 г. в изучаемых ценозах зафиксировано 3 древесных вида, 13 видов кустарников, 82 вида травяно-кустарничкового яруса и 6 видов мхов. В 2002 г. было отмечено 3 древесных вида, 13 видов кустарников, 74 вида травяно-кустарничкового яруса и 6 видов мхов.
Месторасположение пробных площадей в лесостепной зоне определило эколого-ценотическую структуру флоры, в которой преобладают группы лугового и лугово-лесного разнотравья и злаков (33%), лугово-лесного крупнотравья (13%), лесостепные виды (12%). Подавляющее большинство видов относятся к светлохвойной (37.5–56.5%) и лесостепной (10.3–43.7%) поясно-зональным группам. Вклад синантропных (пасквальных, рудеральных, сегетальных) видов в флористический состав исследованных сосновых фитоценозов достаточно велик и составляет 13.8%.
В 2017 г. значение коэффициента сходства Сёренсена–Чекановского между флористическими списками всех изученных пробных площадей варьирует от 0.44 до 0.57. Наибольшие изменения в видовом составе живого напочвенного покрова за 15 лет отмечены на ПП 1 (Ksc = 0.55), наименьшие – на ПП 4, практически не подвергавшейся антропогенному воздействию (Ksc = 0.91). На ПП 2 и ПП 3 изменения видового состава живого напочвенного покрова носили умеренный характер (Ksc = 0.72 и Ksc = 0.75 соответственно) (табл. 3).
Таблица 3.
Характеристика напочвенного покрова в 2002 г. (в числителе) и 2017 г. (в знаменателе) Table 3. The characteristics of ground cover in 2002 (in the numerator) and 2017 (the denominator)
| Показатель Index |
Пробные площади Sample plots | |||
|---|---|---|---|---|
| ПП 1 Sample plot 1 |
ПП 2 Sample plot 2 |
ПП 3 Sample plot 3 |
ПП 4 Sample plot 4 |
|
| Индекс Шеннона Shannon index |
$~\frac{{1.41}}{{2.59}}$ | $\frac{{1.79}}{{2.06}}$ | $\frac{{3.02}}{{3.27}}~$ | $\frac{{3.0}}{{3.11}}$ |
| Коэффициент Сёренсена–Чекановского* Czekanowski–Sørensen index* |
0.55 | 0.72 | 0.75 | 0.91 |
| Индекс синантропизации Synantropization index |
$\frac{{33.3}}{{14.71}}$ | $\frac{{9.5}}{{8.0}}$ | $\frac{{9.09}}{{19.15}}$ | $\frac{{2.27}}{0}$ |
| Стадия деградации напочвенного покрова Stage of ground cover degradation |
$\frac{{{\text{III}}}}{{{\text{II}}}}$ | $\frac{{{\text{II}}}}{{{\text{II}}}}$ | $\frac{{\text{I}}}{{{\text{II}}}}$ | $\frac{{\text{I}}}{{\text{I}}}$ |
Установлено, что индекс видового разнообразия (индекс Шеннона) на ПП 3 и 4 на 17–37% выше, чем на ПП 1 и 2 (табл. 3). Следует отметить, что за 15 лет, прошедших с момента предыдущего исследования, индекс Шеннона возрос на всех пробных площадях. Однако на ПП 3 и 4 это увеличение было незначительным (на 7.6 и 3.5% соответственно), тогда как на ПП 2 индекс Шеннона возрос на 13.1%, а на ПП 1 увеличение индекса было наибольшим – в 1.8 раз.
На ПП 1 за 15 лет, прошедших со времени предыдущего исследования, значительно возросло (P < 0.05) видовое разнообразие подлеска (на 60%). Состав доминантов травяно-кустарничкового яруса и проективное покрытие доминантных видов, относительно устойчивых к вытаптыванию – Calamagrostis arundinacea, Carex macroura [31], не изменились, однако заметно возросло (с 10 до 25%) проективное покрытие видов, малоустойчивых к вытаптыванию – Vicia unijuga и Polygonatum odoratum. Снизилось на 4–7% проективное покрытие видов, относящихся к эколого-ценотической группе сорных: Chenopodium album L., Atriplex sagittata Borkh., Geum rivale L., полностью исчезли из видового состава такие синантропные виды, как Urtica dioica L., Trifolium repens L., Glechoma hederacea L., Cirsium setosum (Willd.) Besser ex M.Bieb. Наряду с этим появились виды следующих эколого-ценотических групп: лесостепные (Origanum vulgare L., Adenophora liliifolia (L.) A. DC.), бореальное мелкотравье (Orthilia secunda (L.) House, Viola biflora L.), луговое и лугово-лесное разнотравье и злаки (Brachypodium pinnatum (L.) P.Beauv., Sanguisorba officinalis L.) и лугово-лесное крупнотравье (Cacalia hastata L.).
На ПП 2 в 2017 г. по сравнению с 2002 г. состав доминантов травяно-кустарничкового яруса не изменился. Увеличение индекса видового разнообразия произошло за счет появления видов, относящихся к эколого-ценотическим группам лугового и лугово-лесного разнотравья и злаков (Fragaria vesca L., Galium boreale L., Sanguisorba officinalis), лугово-лесного крупнотравья (Thalictrum minus), бореального мелкотравья (Maianthemum bifolium (L.) F.W. Schmidt, Neottianthe cucullata (L.) Schltr.).
Отмеченные на ПП 1 и 2 изменения можно объяснить снижением техногенной (табл. 2) и рекреационной нагрузки. В настоящее время на этих территориях, хотя и наблюдаются следы рекреационного воздействия, оно стало заметно меньше, чем было 15 лет назад в связи с закрытием пионерлагерей, организацией обустроенных мест отдыха горожан.
На ПП 3 за истекший период сократилось (с 10–15 до 1–5%) проективное покрытие или произошло исчезновение видов, относящихся к водно-болотной эколого-ценотической группе (Parnassia palustris L.) и к группе лугово-лесного крупнотравья (Heracleum dissectum Ledeb., Lilium martagon L., Pleurospermum uralense Hoffm, Angelica sylvestris L., Chamaenerion angustifolium (L.) Scop. и др.). Наряду с этим появились виды, не отмеченные ранее: Carum carvi, Achillea millefolium L., Glechoma hederacea, Poa annua L. и др. и относящиеся в основном к группе синантропных [3]. Проективное покрытие Trifolium repens увеличилось с 1 до 7%. Известно [20], что клевер ползучий служит индикатором перевыпаса и повышение его проективного покрытия характерно для начальной стадии пастбищной дигрессии.
Наблюдаемые изменения видового состава напочвенного покрова на ПП 3 свидетельствуют об усилении рекреационной и пасквальной нагрузки, что, в частности, приводит к уплотнению и иссушению почвы [17].
На ПП 4 состав доминантов напочвенного покрова не изменился. В настоящее время на данной пробной площади не произрастают адвентивные виды (табл. 3). Единственный синантропный вид (Achillea millefolium), произраставший ранее, в 2017 г. не отмечен.
Из изученных сосновых фитоценозов наибольший индекс синантропизации (19%) в настоящее время отмечен на ПП 3, где он за истекший период увеличился почти в 2 раза (табл. 3). На остальных пробных площадях зафиксировано снижение индекса синантропизации за 15 лет, причем на ПП 1 это снижение было наибольшим (в 2 раза).
Одним из ведущих показателей, характеризующих состояние лесного фитоценоза, является продуктивность напочвенного покрова. В 2017 г. наибольший запас фитомассы живого напочвенного покрова отмечен в контроле на ПП 4 (357.88 ± 70.31 г/м2), за 15-летний период он достоверно не изменился (рис. 2). Запасы фитомассы напочвенного покрова на ПП 1 составляют 121 ± 21 г/м2, на ПП 3 138 ± 18 г/м2, что в 2.6–3 раза ниже, чем соответствующие значения в контрольном фитоценозе. За истекший период на этих пробных площадях запасы фитомассы живого напочвенного покрова увеличились на 43.5 и 31.8% соответственно. На ПП 2 в 2017 г. запас фитомассы живого напочвенного покрова составил 261.4 ± 24.65 г/м2, что в 1.4 раза ниже, чем в контроле. За 15‑летний период запас фитомассы на ПП 2 увеличился наиболее значительно – в 3.7 раз.
Рис. 2.
Общий запас фитомассы напочвенного покрова в сосновых ценозах: 1 – 2002 г., 2 – 2017 г. По горизонтали – пробные площади; по вертикали – фитомасса, г/м2. Fig. 2. Total ground layer phytomass in pine phytocenoses in: 1 – 2002, 2 – 2017. X-axis – sample plots; y-axis – phytomass, g/m2.
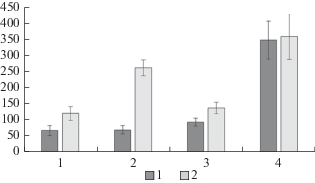
В литературе нередко отмечается тенденция снижения запаса фитомассы живого напочвенного покрова при увеличении антропогенного воздействия [4]. Однако известно, что при длительном воздействии антропогенного фактора в фитоценозе наступает фаза адаптации, когда растения могут увеличивать биомассу, интенсивность роста и размножения [32], что, по-видимому, и наблюдается в нашем случае на ПП, подверженных антропогенной нагрузке.
Сопоставление данных по структуре фитомассы в исследованных фитоценозах показало наличие существенных различий. На контрольной ПП 4 более 80% общего запаса фитомассы составляют мхи. Вклад мхов в запас фитомассы напочвенного покрова на ПП 3, испытывающей рекреационную и пасквальную нагрузки, составляет 57%, тогда как на пробных площадях, подвергавшихся в прошлом техногенному воздействию и подвергающихся рекреационному воздействию в настоящее время, не превышает 1–3%.
Сравнительный анализ данных 2002 и 2017 гг. показал, что за 15-летний период структура фитомассы на всех ПП изменилась (табл. 4). При предыдущем исследовании на ПП 1 наибольший вклад (61.6%) в состав надземной фитомассы вносила осока большехвостая (Carex macroura), на данный момент ее доля снизилась до 47%, хотя абсолютные значения фитомассы увеличились с 42.07 ± 1.03 до 56.91 ± 2.01 г/м2. Та же картина наблюдается в отношении других устойчивых и относительно устойчивых к уплотнению почвы видам (Calamagrostis arundinacea, Poa annua, Thalictrum minus): снижение процентного вклада при одновременном увеличении абсолютного значения запаса фитомассы. Аналогичные изменения отмечены и на ПП 2, где за 15-летний период уменьшился процентный вклад, но возросла фитомасса видов, устойчивых и относительно устойчивых к уплотнению почвы (Carex macroura, Poa annua, Thalictrum minus). Эти результаты согласуется с приведенным выше мнением [32] о возможности увеличения запаса фитомассы некоторых видов при хроническом умеренном антропогенном воздействии. Низкие значения фитомассы зеленых мхов и их доли в общем запасе растений напочвенного покрова на ПП 1 и 2 (табл. 4), возможно, являются остаточным следствием более высокого в прошлом уровня техногенного загрязнения на этих территориях.
Таблица 4.
Фитомасса наиболее продуктивных видов растений напочвенного покрова (г/м2) и их доля (%) от общей фитомассы напочвенного покрова в 2002 г. (в числителе) и 2017 г. (в знаменателе) Table 4. Phytomass of the most productive ground layer plant species (g/m2) and their percentage of the total ground layer phytomass (%) in 2002 (in the numerator) and 2017 (in the denominator)
| Вид Species |
ПП 1 Sample plot 1 |
ПП 2 Sample plot 2 |
ПП 3 Sample plot 3 |
ПП 4 Sample plot 4 |
||||
|---|---|---|---|---|---|---|---|---|
| Фитомасса Phytomass | Доля Percentage |
Фитомасса Phytomass | Доля Percentage |
Фитомасса Phytomass | Доля Percentage |
Фитомасса Phytomass | Доля Percentage |
|
| Calamagrostis arundinacea | $\frac{{4.99}}{{5.48}}$ | $\frac{{7.3}}{{4.5}}$ | $\frac{{4.1}}{{{\text{13}}{\text{.38}}}}$ | $\frac{{5.8}}{{5.1}}$ | $\frac{{3.98}}{{6.20}}$ | $\frac{{4.2}}{{4.5}}$ | $\frac{{3.47}}{{2.37}}$ | $\frac{{1.0}}{{0.7}}$ |
| Carex macroura | $\frac{{42.07}}{{56.91}}$ | $\frac{{61.6}}{{47.0}}$ | $\frac{{28.58}}{{40.92}}$ | $\frac{{40.7}}{{15.7}}$ | $\frac{{0.14}}{{7.02}}$ | $\frac{{0.2}}{{5.1}}$ | $\frac{{7.05}}{{5.32}}$ | $\frac{{2.0}}{{1.5}}$ |
| Fragaria vesca | $\frac{{0.53}}{{0.17}}$ | $\frac{{0.8}}{{0.1}}$ | $\frac{{1.56}}{{9.24}}$ | $\frac{{2.2}}{{3.5}}$ | $\frac{{3.87}}{{0.48}}$ | $\frac{{4.1}}{{0.3}}$ | – | – |
| Galium boreale | $\frac{{0.09}}{{3.48}}$ | $\frac{{0.1}}{{2.9}}$ | $\frac{{0.13}}{{1.95}}$ | $\frac{{0.2}}{{0.7}}$ | $\frac{{0.10}}{{0.17}}$ | $\frac{{0.1}}{{0.1}}$ | $\frac{{0.38}}{{0.21}}$ | $\frac{{0.1}}{{0.1}}$ |
| Geranium sylvaticum | $\frac{{0.35}}{{2.26}}$ | $\frac{{0.5}}{{1.9}}$ | $\frac{{0.39}}{{3.51}}$ | $\frac{{0.6}}{{1.3}}$ | $\frac{{3.86}}{{1.10}}$ | $\frac{{4.1}}{{0.8}}$ | $\frac{{7.46}}{{8.29}}$ | $\frac{{2.1}}{{2.3}}$ |
| Lathyrus humilis | $\frac{{0.01}}{{1.57}}$ | $\frac{{0.01}}{{1.2}}$ | $\frac{{1.23}}{{4.79}}$ | $\frac{{1.8}}{{1.8}}$ | – | – | $\frac{{5.22}}{{1.25}}$ | $\frac{{1.5}}{{0.4}}$ |
| Maianthemum bifolium | – | – | $\frac{{0.66}}{{0.45}}$ | $\frac{{0.9}}{{0.2}}$ | $\frac{{1.38}}{{1.12}}$ | $\frac{{1.5}}{{0.8}}$ | $\frac{{2.72}}{{0.53}}$ | $\frac{{0.8}}{{0.1}}$ |
| Orthilia secunda | – | – | $\frac{{0.12}}{{0.39}}$ | $\frac{{0.2}}{{0.2}}$ | $\frac{{1.13}}{{2.55}}$ | $\frac{{1.2}}{{1.9}}$ | $\frac{{3.54}}{{2.22}}$ | $\frac{{1.0}}{{0.6}}$ |
| Poa annua | $\frac{{6.10}}{{7.62}}$ | $\frac{{8.9}}{{6.3}}$ | $\frac{{0.81}}{{1.93}}$ | $\frac{{1.1}}{{0.7}}$ | – | – | – | – |
| Polygonatum odoratum | $\frac{{0.99}}{{12.26}}$ | $\frac{{1.4}}{{10.1}}$ | $\frac{{0.27}}{{5.15}}$ | $\frac{{0.4}}{{1.9}}$ | – | – | – | – |
| Pulmonaria mollis | – | – | – | – | $\frac{{7.35}}{{0.28}}$ | $\frac{{7.8}}{{0.2}}$ | $\frac{{0.53}}{{0.61}}$ | $\frac{{0.2}}{{0.2}}$ |
| Pyrola rotundifolia | – | – | $\frac{{7.68}}{{23.84}}$ | $\frac{{10.9}}{{9.1}}$ | $\frac{{5.63}}{{9.04}}$ | $\frac{{6.0}}{{6.6}}$ | – | – |
| Rubus saxatilis | $\frac{{3.00}}{{9.18}}$ | $\frac{{4.4}}{{7.6}}$ | $\frac{{3.53}}{{14.21}}$ | $\frac{{5.0}}{{5.4}}$ | $\frac{{4.85}}{{4.09}}$ | $\frac{{5.2}}{{2.9}}$ | $\frac{{4.86}}{{4.54}}$ | $\frac{{1.4}}{{1.27}}$ |
| Sanguisorba officinalis | – | – | $\frac{{1.16}}{{3.29}}$ | $\frac{{1.7}}{{1.3}}$ | – | – | $\frac{{3.47}}{{3.02}}$ | $\frac{{1.0}}{{0.8}}$ |
| Thalictrum minus | $\frac{{8.00}}{{9.92}}$ | $\frac{{11.7}}{{8.2}}$ | $\frac{{9.77}}{{20.86}}$ | $\frac{{13.9}}{{7.9}}$ | – | – | $\frac{{6.25}}{{8.50}}$ | $\frac{{1.8}}{{2.4}}$ |
| Vaccinium vitis-idaea | – | – | – | – | $\frac{{1.05}}{{1.81}}$ | $\frac{{1.1}}{{1.3}}$ | $\frac{{5.90}}{{6.81}}$ | $\frac{{1.7}}{{1.9}}$ |
| Viola biflora | $\frac{{0.12}}{{2.26}}$ | $\frac{{0.2}}{{1.9}}$ | – | – | $\frac{{4.98}}{{0.14}}$ | $\frac{{5.3}}{{0.1}}$ | $\frac{{0.35}}{{1.36}}$ | $\frac{{0.1}}{{0.4}}$ |
| Hylocomium splendens | – | – | – | – | $\frac{{34.82}}{{58.20}}$ | $\frac{{37.1}}{{42.3}}$ | $\frac{{63.18}}{{74.63}}$ | $\frac{{18.2}}{{20.9}}$ |
| Pleurozium schreberi | $\frac{{1.43}}{{1.45}}$ | $\frac{{2.1}}{{1.2}}$ | $\frac{{5.05}}{{5.75}}$ | $\frac{{7.2}}{{2.2}}$ | $\frac{{18.21}}{{19.01}}$ | $\frac{{19.4}}{{13.8}}$ | $\frac{{137.8}}{{143.9}}$ | $\frac{{39.7}}{{40.2}}$ |
На ПП 3 в оба срока наблюдений основной вклад в запас фитомассы живого напочвенного покрова вносят мхи (56–57%). Следует отметить, что за исследуемый период изменилась структура фитомассы травяно-кустарничкового яруса: достоверно снизился вклад видов, малоустойчивых к уплотнению почвы (Geranium sylvaticum, Fragaria vesca, Rubus saxatilis, Pulmonaria mollis), при одновременном увеличении доли Carex macroura – вида, относительно устойчивого к уплотнению почвы. Это обстоятельство может быть объяснено возросшей рекреационной нагрузкой.
На ПП 4 за 15-летний период структура фитомассы достоверно не изменилась. Наибольший вклад в нее вносят мхи (более 80%). Из видов травяно-кустарничкового яруса помимо видов-доминантов (Calamagrostis arundinacea, Vaccinium vitis-idaea, Carex macroura, Rubus saxatilis), на долю которых приходится 5–6% от общего запаса фитомассы, существенной является доля (5–5.5%) таких лугово-лесных и лесных видов, как Thalictrum minus, Geranium sylvaticum, Sanguisorba officinalis.
Согласно шкале рекреационной трансформации растительных сообществ [20], в настоящее время ПП 1, 2, 3 находятся на стадии слабой рекреационной трансформации (II стадия). В ходе исследований 2002 г. было установлено [33], что на ПП 3 рекреационные нагрузки были минимальны (I стадия деградации), ПП 2 находилась на II стадии деградации [34] и в большей степени подвергалась техногенным нагрузкам, а ПП 1 – на III стадии деградации из-за дополнительных рекреационных нагрузок. Таким образом, в настоящее время вследствие уменьшения объемов выбросов загрязняющих веществ и снижения рекреационной нагрузки состояние напочвенного покрова сосновых фитоценозов на ПП 1 улучшилось, на ПП 2 осталось без существенного изменения, а на ПП 3, напротив, ухудшилось в связи с усилением рекреационной нагрузки.
ЗАКЛЮЧЕНИЕ
В результате сравнительной оценки состояния напочвенного покрова антропогенно нарушенных сосняков Красноярской лесостепи в 2002 и 2017 гг. установлено, что в связи с существенным снижением объемов промышленных выбросов изменения, произошедшие за 15-летний период, обусловлены преимущественно рекреационной, а не техногенной нагрузкой.
Анализ параметров видового разнообразия, соотношения эколого-ценотических групп видов, величины и структуры запаса фитомассы напочвенного покрова показал, что наиболее существенные изменения произошли на территории Березовского бора. В результате снижения рекреационной нагрузки в сосняке осочково-разнотравном повысилось видовое разнообразие, в 2 раза снизилось участие синантропных видов по числу, сократилась их доля в общем запасе фитомассы, стадия деградации изменилась с III на II. На территории Есаульского бора в сосняке разнотравно-осочково-зеленомошном изменения носили аналогичный характер, но проявились в значительно более слабой степени, в связи с чем сохраняется II стадия деградации напочвенного покрова. На территории Юксеевского бора в сосняке мелкотравно-зеленомошном вследствие возросшей рекреационной нагрузки, отмечено 2-кратное увеличение доли синантропных видов, существенное увеличение их фитомассы и доли в общем запасе фитомассы, снижение участия лугово-лесного крупнотравья; усиление деградации напочвенного покрова (произошла смена I стадии на II). На контрольной пробной площади (Погорельский бор) в сосняке разнотравно-зеленомошном за 15-летний период видовой состав живого напочвенного покрова, его структура и запас фитомассы существенно не изменились.
Список литературы
Brown J.H., Kalisz S.P., Wright W.R. 1977. Effects of recreational use on forested sites. – Environmental Management. 1(5): 425–431. https://doi.org/10.1007/BF01866999
Weaver T., Dale D. 1978. Trampling effects of hikers, motorcycles and horses in meadows and forests. – J. Appl. Ecol. 15(2): 451–457. https://doi.org/10.2307/2402604
Горчаковский П.Л. 1999. Антропогенная трансформация и восстановление продуктивности луговых фитоценозов. Екатеринбург. 156 с.
Полякова Г.А., Малышева Т.В., Флеров В.А. 1981. Антропогенное влияние на сосновые леса Подмосковья. М. 144 с.
Природные аспекты рекреационного использования леса. 1987. М. 167 с.
Влияние рекреации на лесные экосистемы и их компоненты. 2004. Пущино. 302 с.
Динамика и устойчивость рекреационных лесов. 2006. М. 165 с.
Wittig R., König K., Schmidt M., Szarzynski J. 2007. A study of climate change and anthropogenic impacts in West Africa. – Environ. Sci. Pollut. R. 14(3): 182–189. https://doi.org/10.1065/espr2007.02.388
Антропогенная трансформация растительного покрова Западной Сибири. 1992. Новосибирск. 152 с.
Rodenkirchen H. 1992. Effects of acidic precipitation, fertilization and liming on the ground vegetation in coniferous forests of southern Germany. – Water, Air, Soil Poll. 61(3–4): 279–294. https://doi.org/10.1007/BF00482611
Blake L., Goulding K.W.T. 2002. Effects of atmospheric deposition, soil pH and acidification on heavy metal contents in soils and vegetation of semi-natural ecosystems at Rothamsted Experimental Station, UK. – Plant Soil. 240(2): 235–251. https://doi.org/10.1023/A:1015731530498
Chandrashekara U.M., Muraleedharan P.K., Sibichan V. 2006. Anthropogenic pressure on structure and composition of a shola forest in Kerala, India. – J. Mt. Sci. 3(1): 58–70. https://doi.org/10.1007/s11629-006-0058-0
Меланхолин П.Н. 2006. Изменение видового разнообразия травяно-кустарничкового яруса при различных антропогенных нагрузках на лесные экосистемы. – Лесоведение. 6: 52–58.
Gao W., Yongfen D., Gao S., Ingels J., Wang D. 2016. Heavy metal accumulation reflecting natural sedimentary processes and anthropogenic activities in two contrasting coastal wetland ecosystems, eastern China. – J. Soils Sediment. 16(3): 1093–1108. https://doi.org/10.1007/s11368-015-1314-0
МакКленахен Дж.Р. 1982. Изменения в лесном сообществе в связи с загрязнением воздуха. – В кн.: Взаимодействие лесных экосистем и атмосферных загрязнителей. Таллин. Ч. 1. С. 38–42.
Кузьмина Г.П., Яновский В.М. 1999. Антропогенная трансформация лесных экосистем Средней Сибири. – В сб.: Леса и лесообразовательный процесс на Дальнем Востоке: Материалы Междун. конф. Владивосток. С. 239–240.
Мониторинг рекреационных лесов. 2003. М.; Пущино. 167 с.
Полякова Г.А., Меланхолин П.Н. 2007. Изменение антропогенной нарушенности лесопарков Москвы. – В сб.: Актуальные проблемы рекреационного лесопользования. Международ. науч. конф. М. С. 60–62.
Allen C.D. 2009. Monitoring environmental impact in the Upper Sonoran lifestyle: A new tool for rapid ecological assessment. – Environmental Management. 43(2): 346–356. https://doi.org/10.1007/s00267-008-9212-5
Прокопьев Е.П., Рыбина Т.А. 2010. Опыт мониторинга синантропизации и антропогенной трансформации растительного покрова особо охраняемых природных территорий г. Томска. – Вестн. Томского гос. ун-та. Биология. 3(11): 109–118. http://journals.tsu.ru/biology/&journal_page=archive&id=750&article_id=15984
Воронов А.Г. 1973. Геоботаника. М. 384 с.
Назимова Д.И., Дробушевская О.В., Данилина Д.М., Коновалова М.Е., Кофман Г.Б., Бугаева К.С. 2012. Биоразнообразие и динамика низкогорных лесов Саян: региональный и локальный уровни. – В кн.: Разнообразие и динамика лесных экосистем России. М. Кн. 1. С. 131–173.
Буторина Т.Н. 1963. Эколого-ценотический анализ кустарничково-травяного яруса лесных ассоциаций. – В кн.: Типы лесов Сибири. М. С. 31–52.
Шмидт В.М. 1984. Математические методы в ботанике. Л. 288 с.
Кузьмина Г.П. 1982. Влияние рекреации на новые леса зеленой зоны г. Красноярска: Автореф. дис. … канд. биол. наук. Красноярск. 24 с.
Таран И.В. 1985. Рекреационные леса Западной Сибири. Новосибирск. 228 с.
Методические указания по ионометрическому определению содержания фтора в растительной продукции, кормах и комбикормах. 1995. МСК.
ПНД Ф 16.1:2.3:3.11-98. Количественный химический анализ почв. Методика выполнения измерений содержания металлов в твердых объектах методом спектрометрии с индуктивно-связанной плазмой.
Татаринцев А.И., Скрипальщикова Л.Н. 2003. Сосновые фитоценозы в зоне многолетнего воздействия антропогенных нагрузок. – География и природ. ресурсы. 3: 53–57.
Реализация проекта “Экологический Содерберг”. http://tehne.com/assets/i/upload/event/RUSAL-Krasnoiarsk.pdf.
Таран И.В., Спиридонов В.Н. 1977. Устойчивость рекреационных лесов. Новосибирск. 179 с.
Рысин Л.П., Рысин С.Л. 2012. Урболесоведение. М. 240 с.
Экологическое состояние пригородных лесов Красноярска. 2009. Новосибирск. 179 с.
Полякова Г.А. 1980. Деградация сосняков Подмосковья под влиянием рекреации. — Лесоведение. 5: 62–69.
Кабата-Пендиас А., Пендиас X. 1989. Микроэлементы в почвах и растениях. М. 439 с.
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы