

Растительные ресурсы, 2019, T. 55, № 3, стр. 389-397
Сезонный ритм развития и зимостойкость некоторых видов рода Spiraea (Rosaceae) в условиях интродукции (г. Благовещенск)
Т. В. Ступникова 1, *, В. В. Семенко 2
1 Амурский филиал Ботанического сада-института ДВО РАН
г. Благовещенск, Россия
2 Дальневосточный государственный аграрный университет
г. Благовещенск, Россия
* E-mail: stupnikovat@yandex.ru
Поступила в редакцию 19.03.2019
После доработки 25.03.2019
Принята к публикации 17.04.2019
Аннотация
Приведены результаты изучения ритма сезонного развития 22 таксонов рода Spiraea L., интродуцированных в Амурском филиале Ботанического сада-института ДВО РАН. Установлено, что зимостойкость спирей зависит от синхронности окончания вегетации с окончанием вегетации аборигенными древесными породами. В озеленении Благовещенска рекомендовано использовать зимостойкие спиреи (S. alpina, S. aquilegifolia, S. betulifolia, S. elegans, S. media, S. pubescens, S. salicifolia, S. sericea, S. trilobata, S. ussuriensis) и сравнительно зимостойкие (S. × bumalda, S. japonica, S. longigemmis, S. × vanhouttei).
Значительный интерес в озеленении представляет обширный род спирея Spiraea L., насчитывающий по разным литературным данным от 90 до 120 видов, а также значительное число спонтанных межвидовых гибридов, сортов и форм [1, 2]. Спиреи неприхотливы к почвенным условиям, быстро растут, цветут и плодоносят на третий год жизни, достаточно долговечны: в культуре в среднем живут 20 лет, а при надлежащем уходе – до 40 лет [3]. К сожалению, этот род недооценен и практически не находит своего применения в ландшафтном дизайне нашего города [4]. Изредка в озеленении придомовых территорий Благовещенска используются S. media Fr. Schmidt. и S. salicifolia L. В последнее время в скверах и парках города появились посадки S. japonica L., реализацию которой осуществляет частный питомник “Биопарк”.
В Амурском филиале Ботанического сада-института ДВО РАН (АФ БСИ ДВО РАН) начиная с 2002 г. интродукционное испытание прошли более 40 таксонов спирей, около 30 из которых успешно введены в культуру. Коллекционный фонд сформирован из семенного и живого растительного материала, собранного в природных популяциях или полученного по обмену с отечественными и зарубежными ботаническими садами.
Исследования показали, что основным ограничивающим фактором для интродукции некоторых спирей (S. blumei G.Don, S. formosana Hayata, S. hayatana H.L. Li, S. mollifolia Rehder., S. miyabei Koidz., S. rosthornii E. Pritz. ex Diels, S. × rubella Dippel.) на юг Амурской обл. является их низкая зимостойкость. Известно, что перспективность вида для интродукции помимо зимостойкости определяется также степенью синхронности сроков начала и окончания вегетации растений с местными природно-климатическими условиями [5]. В литературе имеются данные о результатах первичного интродукционного испытания восточноазиатских видов спирей в АФ БСИ ДВО РАН [6] и зимостойкости отдельных представителей [7], однако ритм сезонного развития интродуцированных спирей на юге Амурской обл. ранее не исследовался.
Цель работы – изучить ритм сезонного развития спирей в сравнении с их зимостойкостью для определения перспективных видов, сортов и форм при дальнейшей интродукции и использовании в озеленении на юге Амурской обл.
МАТЕРИАЛ И МЕТОДЫ
Исследования проводили в 2012–2018 гг. в коллекции генетических ресурсов растений АФ БСИ ДВО РАН в окрестностях г. Благовещенск (50°16′46″ с.ш., 127°32′25″ в.д.). Климат Благовещенска резко континентальный с муссонной циркуляцией воздушных масс. Среднегодовая температура воздуха 0 °С, что на 5.4 °С ниже соответствующей на данной широте. Зима и лето продолжительные, а переходные сезоны короткие, с резкими и быстрыми изменениями метеорологических показателей. Продолжительность безморозного периода составляет около 160 дней, период активных температур 134 дня. Среднегодовое количество атмосферных осадков 575 мм, из которых на холодный период (с ноября по март) приходится всего 8% годовой нормы, на теплый (с апреля по октябрь) – 92%. В течение теплого сезона осадки распределены также неравномерно: весна и первая половина лета, как правило, засушливые, во второй половине лета преимущественно в виде ливневых дождей выпадает около 50% годовой нормы. Средняя высота снежного покрова составляет около 15 см, почва промерзает на глубину 2–2.5 м [8]. Согласно данным метеостанции Благовещенска за последние 20 лет наблюдается увеличение безморозного периода.
Объекты исследования – 22 таксона спирей.
1. S. alpina Pall. Кустарник около 1 м выс., с раскидистой кроной, листья узколанцетные до 2.5 см длиной, цветки белые в щитковидных соцветиях диам. около 2 см. Родина – Сибирь, горы Центральной Азии.
2. S. aquilegifolia Pall. Кустарник около 1.2 м выс., с раскидистой кроной, листья трехлопастные до 1.7 см длиной, цветки белые в зонтиковидных соцветиях диам. около 2 см. Родина – Забайкалье, Монголия, Северный Китай.
3. S. × arguta Zab. Гибрид (S. × multiflora Zab.× S. thunbergii Siebold ex Blume). Кустарник около 1.5 м выс., с каскадной кроной, листья широкоэллиптические до 4.0 см длиной, цветки белые в плотных щитковидных соцветиях диам. около 3 см.
4. S. betulifolia Pall. Кустарник около 0.8 м выс., с подушковидной кроной, листья эллиптические до 7 см длиной, цветки белые или розовые в щитковидных соцветиях диам. 3–9 см. Родина – Восточная Сибирь, Дальний Восток, Китай (Маньчжурия).
5. S. × bumalda Burv. Гибрид (S. japonica L. × S. albiflora (Miq.) Zab.). Кустарник около 0.8 м выс., с шаровидной кроной, листья яйцевидно-ланцетные около 8 см длиной, цветки розовые в щитковидных соцветиях диам. 10–15 см.
6. S. × bumalda f. crispa. Кустарник 0.5 м выс., с шаровидной кроной, листья продолговато-яйцевидные с глубоко надрезанным краем около 5 см длиной, цветки ярко-розовые в щитковидных соцветиях диам. около 7 см.
7. S. crenata L. Кустарник около 1.5 м выс., с раскидистой кроной, листья узкоэллиптические около 2.5 см длиной, цветки белые в плотных щитках диам. около 3 см. Родина – Восточная Европа, Кавказ, Средняя Азия, Западная Сибирь (юг).
8. S. elegans Pojark. Кустарник около 1.0 м выс., с ажурной раскидистой кроной, листья узкоэллиптические около 3.0 см длиной, цветки белые в рыхлых щитках диам. около 2.5 см. Родина – Восточная Сибирь, Дальний Восток, Монголия.
9. S. fritschiana C.K. Schneid. Кустарник около 1.0 м выс., с шаровидной кроной, листья яйцевидно-продолговатые около 8 см длиной, цветки розовые в щитковидных соцветиях диам. около 12 см. Родина – Центральный Китай.
10. S. japonica L. Кустарник около 1.0 м выс. с шаровидной кроной, листья яйцевидно-ланцетные 5–7 см длиной, цветки ярко-розовые в щитковидных соцветиях диам. около 12 см. Родина – Япония, Корея, Китай.
11. S. japonica f. alpina (Maxim.) Koidz. Кустарник около 0.5 м выс., с компактной шаровидной кроной, листья около 3 см длиной, цветки розовые в густых щитковидных соцветиях диам. 3–4 см.
12. S. japonica ‘Golden princess’. Кустарник около 30 см выс., с шаровидной кроной. Листья с сезонной изменчивостью окраски: весной – ярко-желтого цвета, летом окрашены в ярко-зеленые тона, осенью приобретают карминный оттенок. Цветки розовые в щитковидных соцветиях диам. около 5 см.
13. S. longigemmis Maxim. Кустарник около 1.0 м выс., с шаровидной кроной, листья яйцевидно-ланцетные до 7 см длиной, цветки белые в щитковидных соцветиях диам. 10–15 см. Родина – Центральный Китай (горные районы).
14. S. media Fr. Schmidt. Кустарник около 1.5 м выс., с раскидистой кроной, листья эллиптические около 3 см длиной, цветки белые в щитковидных соцветиях диам. 2–3 см. Родина – Европа, Сибирь, Дальний Восток, Монголия, Китай (северо-восток).
15. S. myrtilloides Rehder (syn. S. virgata Franch.). Кустарник около 2.5 м высотой, с каскадной кроной, листья широкоэллиптические около 3.0 см длиной, цветки белые в щитковидных соцветиях диам. около 3 см. Родина – Центральный Китай (горные районы).
16. S. nipponica Maxim. Кустарник около 2.0 м высотой, с горизонтально-направленными ветвями, листья широкоэллиптические около 3.0 см длиной, цветки белые в плотных щитковидных соцветиях диам. около 3 см. Родина – Япония.
17. S. pubescens Turcz. Кустарник около 1.5 м высотой, с колоновидной рыхлой кроной, листья продолговато-ромбические около 4.0 см длиной, цветки белые в зонтиковидных соцветиях диам. около 3 см. Родина – Восточная Сибирь (юг), Приморье (юг), Китай (Маньчжурия), Корея.
18. S. salicifolia L. Кустарник около 2.0 м высотой, с колоновидной кроной, листья ланцетные около 5.0 см длиной, цветки розовые в густых пирамидальных метелках диам. около 10 см. Родина – Европа, Сибирь, Дальний Восток, Северная Монголия, Северный Китай, Корея.
19. S. sericea Turcz. Кустарник около 1.5 м выс., с раскидистой кроной, листья шелковисто-опушенные, эллиптические около 3 см длиной, цветки белые в щитковидных соцветиях диам. 2–3 см. Родина – Восточная Сибирь, Дальний Восток, Северная Монголия, Китай (Маньчжурия).
20. S. trilobata L. Сильноветвящийся кустарник около 1.0 м выс. с зигзагообразно-изогнутыми побегами, листья обратнояйцевидные, трехлопастные около 2 см длиной, цветки белые в щитковидных соцветиях диам. около 3 см. Родина – Западная Сибирь (юг), Центральная Азия.
21. S. ussuriensis Pojark. Кустарник около 1.5 м выс., с раскидистой кроной, листья широкоэллиптические около 3 см длиной, цветки белые в щитковидных соцветиях диам. около 3 см. Родина – Дальний Восток, Северная Монголия, Китай (Маньчжурия).
22. S. × vanhauttei (Briot) Zab. Гибрид (S. trilobata × S. cantoniensis Lour.). Кустарник около 1.5 м выс., с раскидистой кроной, листья округлые, трехлопастные около 3 см длиной, цветки белые в щитковидных соцветиях диам. около 3.5 см.
S. crenata, S. japonica, S. japonica f. alpina, S. japonica ‘Golden princess’, S. longigemmis, S. media, S. myrtilloides, S. pubescens, S. salicifolia, S. sericea, S. ussuriensis представлены в коллекции в количестве 10–20 экземпляров, остальные спиреи – в количестве 3–5 экземпляров.
Таблица 1.
Сезонное развитие спирей в Амурском филиале БСИ ДВО РАН Table 1. Seasonal development of Spiraea in Amur branch of Botanical Garden-Institute FEB RAS
| Таксон Taxon |
Дата наступления фенофазы Date of onset of phenological stage |
Продолжительность, дни Duration, days |
Зимостойкость Winter hardiness |
||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Начало вегетации Beginning of vegetation |
Цветение Flowering |
Плодоношение Fruiting |
Конец вегетации End of vegetation |
Цветения Flowering |
Созревания плодов Ripening of fruit |
Вегетации Vegetation |
|||||
| Начало Beginning |
Конец End |
Массовое Full |
Формирование Formation |
Зрелые плоды Fruit maturity |
|||||||
| S. alpina | 20 IV ± ± 4.2 | 24 V ± ± 4.6 | 14 VI ± ± 4.2 | 29 V−03 VI | 04 VI ± ± 1.7 | 23 VIII ± ± 6.0 | 24 IX ± ± 2.3 | 22 ± ± 1.3 | 81 ± ± 16.7 | 158 ± ± 3.4 | I |
| S. aquilegifolia | 19 IV ± ± 4.1 | 07 V ± ± 2.8 | 30 V ± ± 3.0 | 10−16 V | 17 V ± ± 2.4 | 15 VI ± ± 6.6 | 28 IX ± ± 2.4 | 24 ± ± 2.0 | 30 ± ± 4.7 | 163 ± ± 1 .2 | I |
| S. × arguta | 04 V ± ± 2.8 | 06 VI ± ± 1.7 | 07 VII ± ± 1.3 | 12−22 VI | 24 VI ± ± 2.8 | 26 IX ± ± 3.4 | 16 X ± ± 2.2 | 32 ± ± 3.4 | 95 ± ± 4.6 | 166 ± ± 3.8 | III |
| S. betulifolia | 07 V ± ± 1.6 | 10 VI ± ± 4.2 | 12 VII ± ± 3.0 | 18−27 VI | 20 VI ± ± 4.2 | 30 IX ± ± 6.0 | 30 IX ± ± 3.2 | 33 ± ± 2.4 | 103 ± ± 6.2 | 147 ± ± 1.3 | I |
| S. × bumalda | 24 V ± ± 5.9 | 06 VII ± ± 2.3 | 31 VII ± ± 4.2 | 11−22 VII | 20 VII ± ± 4.2 | 02 X ± ± 6.6 | 05 X ± ± 4.2 | 26 ± ± 4.6 | 75 ± ± 2.3 | 135 ± 4.6 | II |
| S. × bumalda f. ‘crispa’ | 16 V ± ± 4.6 | 14 VII ± ± 8.3 | 09 X ± ± 5.8 | No data | 30 VII ± ± 6.3 | No data | 16 X ± ± 6.2 | 88 ± ± 10.8 | No data | 154 ± ± 4.8 | III |
| S. crenata | 02 V ± ± 4.2 | 23 V ± ± 3.3 | 25 VI ± ± 2.4 | 28 V−04 VI | 02 VI ± ± 3.2 | 05 VIII ± ± 2.7 | 16 X ± ± 2.4 | 34 ± ± 2.4 | 65 ± 2± .7 | 168 ± ± 3.0 | III |
| S. elegans | 18 IV ± ± 1.3 | 12 V ± ± 2.8 | 02 VI ± ± 2.3 | 16−23 V | 18 V ± ± 2.2 | 02 VIII ± ± 4.3 | 26 IX ± ± 2.3 | 22 ± ± 1.4 | 77 ± ± 2.4 | 162 ± ± 2.3 | I |
| S. fritchiana | 05 V ± ± 4.2 | 13 VI ± ± 4.0 | 08 VII ± ± 2.9 | 19−28 VI | 26 VI ± ± 2.3 | No data | 16 X ± ± 3.3 | 26 ± ± 2.3 | No data | 165 ± ± 3.2 | III |
| S. japonica | 10 V ± ± 2.3 | 26 VI ± ± 2.8 | 18 IX ± ± 4.6 | 30 VI – 22 VII | 08 VII± ± 2.0 | 03 X ± ± 6.0 | 05 X ± ± 2.3 | 85 ± ± 3.4 | 88 ± ± 3.0 | 149 ± ± 2.8 | II |
| S. japonica f. alpina | 14 V ± ± 2.0 | 02 VII ± ± 2.3 | 09 X ± ± 3.3 | No data | 18 VII ± ± 3.3 | 16 X ± ± 4.3 | 16 X ± ± 4.2 | 100 ± ± 5.1 | 91 ± ± 6.1 | 156 ± ± 2.3 | III |
| S. japonica ‘Golden princess’ | 14 V ± ± 2.3 | 05 VII ± ± 2.3 | 09 X ± ± 2.0 | No data | 24 VII ± ± 2.7 | 16 X ± ± 6.2 | 16 X ± ± 2.0 | 97 ± ± 6.3 | 85 ± ± 5.3 | 156 ± ± 2.3 | III |
| S. longigemmis | 03 V ± ± 2.8 | 07 VI± ± 3.0 | 16 VII ± ± 2.6 | 12 –27 VI | 15 VI ± ± 3.0 | 02 X ± ± 2.3 | 05 X ± ± 2.6 | 40 ± ± 2.3 | 110 ± ± 4.3 | 156 ± ± 2.3 | II |
| S. media | 18 IV ± ± 1.8 | 16 V ± ± 2.0 | 11 VI ± ± 2.3 | 21–28 V | 24 V ± ± 1.3 | 10 VIII ± ± 6.3 | 17 IX ± ± 1.3 | 27 ± ± 2.0 | 78 ± ± 5.3 | 153 ± ± 1.8 | I |
| S. myrtilloides | 13 V ± ± 3.3 | 06 VI ± ± 2.3 | 04 VII ± ± 2.0 | 10–22 VI | 12 VI ± ± 2.3 | 16 X ± ± 4.2 | 16 X ± ± 2.0 | 29 ± ± 2.3 | 127 ± ± 4.3 | 157 ± ± 2.6 | III |
| S. nipponica | 11 V ± ± 3.2 | 05 VI ± ± 2.0 | 07 VII ± ± 2.0 | 10–22 VI | 12 VI ± ± 3.3 | 16 X ± ± 4.3 | 16 X ± ± 2.3 | 33 ± ± 3.0 | 127 ± ± 3.8 | 159 ± ± 2.1 | III |
| S. pubescens | 23 IV ± ± 2.0 | 27 V ± ± 2.6 | 18 VI ± ± 2.3 | 30 V–05 VI | 02 VI ± ± 2.1 | 05 X ± ± 2.6 | 05 X ± ± 2.2 | 23 ± ± 2.0 | 125 ± ± 3.3 | 166 ± ± 2.0 | II |
| S. salicifolia | 25 IV ± ± 1.8 | 03 VII ± ± 2.8 | 20 IX ± ± 2.3 | No data | 20 VII ± ± 2.0 | 02 X ± ± 2.4 | 30 IX ± ± 1.3 | 80 ± ± 2.6 | 74 ± ± 3.0 | 159 ± ± 2.3 | I |
| S. sericea | 18 IV ± ± 1.3 | 15 V ± ± 2.3 | 06 VI ± ± 2.1 | 21–28 V | 24 V ± ± 1.3 | 10 VIII ± ± 5.0 | 17 IX ± ± 1.3 | 23 ± ± 2.1 | 78 ± ± 5.3 | 153± ± 1.8 | I |
| S. trilobata | 16 V ± ± 2.0 | 10 VI ± ± 2.3 | 07 VII ± ± 2.6 | 14–23 VI | 18 VI ± ± 2.7 | 30 IX ± ± 2.3 | 30 IX ± ± 2.1 | 28 ± ± 2.1 | 104 ± ± 2.8 | 138 ± ± 2.6 | I |
| S. ussuriensis | 18 IV ± ± 1.8 | 17 V ± ± 2.0 | 18 VI ± ± 2.0 | 20–30 V | 24 V ± ± 2.3 | 30 IX ± ± 2.4 | 27 IX ± ± 1.8 | 33 ± ± 2.3 | 130 ± ± 3.3 | 162 ± ± 2.0 | I |
| S. × vanhouttei | 15 V ± ± 3.0 | 10 VI ± ± 2.3 | 08 VII ± ± 2.4 | 17–25 VI | 20 VI ± ± 2.7 | 28 IX ± ± 2.8 | 05 X ± ± 2.0 | 29 ± ± 2.2 | 101 ± ± 3.3 | 153± ± 2.6 | II |
Фенологические наблюдения проводили стандартными методами [9, 10]. За начало вегетации принимали дату распускания почек (почечные чешуи разошлись, виден конус зеленых листьев), за окончание – дату листопада. Обработку данных фенологических наблюдений осуществляли по рекомендациям В.Н. Нилова [11]. Зимостойкость оценивали по шкале Н.Е. Булыгина [12]: I – вполне зимостойкие древесные растения (не повреждаются зимними морозами или при неблагоприятных условиях повреждаются незначительно, что не сказывается на их росте и развитии, цветении и плодоношении, декоративности); II – сравнительно зимостойкие древесные растения (морозами повреждаются лишь в отдельные годы, но полученные повреждения не оказывают влияния на дальнейший рост и развитие); III – сравнительно незимостойкие древесные растения (морозами повреждаются регулярно, но даже при значительном обмерзании габитус сохраняется); IV – незимостойкие древесные растения (обмерзают ежегодно и сильно, от чего жизненная форма изменяется); V – совершенно незимостойкие (полное вымерзание растения). Наблюдения проводили дважды за сезон: в начале активной вегетации (май), когда хорошо заметны зимние повреждения, и в середине лета (июль), когда можно установить степень восстановления утраченных частей.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Результаты наблюдения за сезонным развитием спирей представлены в таблице и в виде фенологических спектров на рис. 1. Сроки начала (18 IV–24 V) и окончания (17 IX–9 X) вегетационного периода у спирей в Благовещенске за годы наблюдений приходятся в среднем на первую декады мая и октября соответственно, а средняя продолжительность этого периода составляет 156 дней. Из представленных материалов следует, что высокой зимостойкостью (I балл) характеризуются спиреи с ранним началом и ранним окончанием вегетации (РР). К ним относятся виды, естественный ареал которых охватывает территорию Дальнего Востока, Сибири, Монголии и северных районов Китая. Вегетация у видов группы РР начинается в третьей декаде апреля, что соответствует переходу суточной температуры через 5 °С, а окончание вегетации – до наступления сильных заморозков. Листопад у спирей этой группы происходит синхронно с древесными породами аборигенной флоры (Fraxinus mandshurica Rupr., Betula davurica Pall., B. platyphylla Sukacz.) во второй–третьей декадах сентября. Перспективна в интродукции S. trilobata – вид с поздним началом и ранним окончанием вегетации.
Рис. 1.
Фенологические спектры спирей, интродуцированных в Амурском филиале БСИ ДВО РАН (средние многолетние показатели за 2012–2018 гг.). a – период вегетации, b – период цветения, c – период созревания плодов. По шкале ординат указаны таксоны спирей: I – S. alpina, II – S. aquilegifolia, III – S. × arguta, IV – S. betulifolia, V – S. × bumalda, VI – S. × × bumalda f. crispa, VII – S. crenata, VIII – S. elegans, IX – S. fritschiana, X – S. japonica, XI – S. japonica f. alpina, XII – S. japonica ‘Golden princess’, XIII – S. longigemmis, XIV – S. media, XV – S. myrtilloides, XVI – S. nipponica, XVII – S. pubescens, XVIII – S. salicifolia, XIX – S. sericea, XX – S. trilobata, XXI – S. ussuriensis, XXII – S. × vanhouttei. Fig 1. Phenological ranges of spireas cultivated in the Amur branch of Botanical Garden-Institute FEB RAS (long-term annual average for 2012–2018). a – vegetation period, b – flowering period, c – full maturity period. Taxa of Spireas is given on the ordinates: I – S. alpina, II – S. aquilegifolia, III – S. × arguta, IV – S. betulifolia, V – S. × bumalda, VI – S. × bumalda f. crispa, VII – S. crenata, VIII – S. elegans, IX – S. fritschiana, X – S. japonica, XI – S. japonica f. alpina, XII – S. japonica ‘Golden princess’, XIII – S. longigemmis, XIV – S. media, XV – S. myrtilloides, XVI – S. nipponica, XVII – S. pubescens, XVIII – S. salicifolia, XIX – S. sericea, XX – S. trilobata, XXI – S. ussuriensis, XXII – S. × vanhouttei.
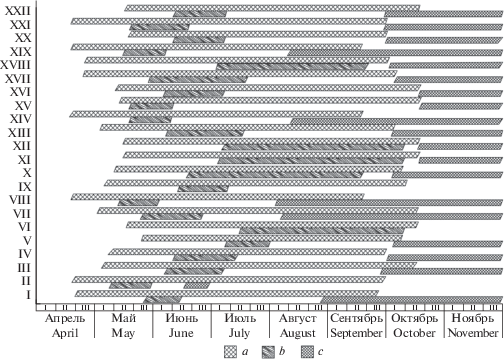
Относительно зимостойкие S. × bumalda, S. japonica, S. longigemmis, S. pubescens, S. × × vanhоuttei начинают вегетацию в разные сроки, а заканчивают вегетацию несколько позже местных древесных растений (в первой декаде октября), поэтому обмерзают лишь в неблагоприятные годы. Менее перспективны (зимостойкость III балла) спиреи со средним и поздним началом вегетации и с поздним ее окончанием (СП, ПП). В эту группу относятся виды из западных и более южных относительно пункта интродукции районов (европейская часть России, Япония, Корея, Центральный и Южный Китай), а также некоторые культурные формы. Начало вегетации у спирей этих групп происходит в мае; а окончание вегетации – во второй–третьей декадах октября, когда в Благовещенске устанавливаются устойчивые отрицательные температуры. По нашим наблюдениям, рост побегов у S. × arguta, S. crenata, S. myrtilloides, S. nipponica продолжается до конца вегетационного периода, что приводит к неполному вызреванию древесины. Опадание листьев у спирей групп СП и ПП происходит под воздействием отрицательных температур, а у некоторых представителей (S. × bumalda f. crispa, S. japonica f. alpina, S. japonica ‘Golden princess’) листья на побегах сохраняются до следующего вегетационного периода. Спиреи групп СП и ПП в агроклиматических условиях юга Амурской обл. не успевают завершить свою вегетацию, вследствие чего морозами повреждаются ежегодно. Однако, несмотря на ежегодное обмерзание побегов, все спиреи сохраняют присущую им жизненную форму.
Цветение у спирей происходит в разные сроки: со второй декады мая и до окончания вегетации. Зимостойкие виды относятся в основном к группе весенне-летнего цветения, период их массового цветения по времени совпадает с развертыванием почек на дубе монгольском (Quercus mongolica Fisch. ex Ledeb.). К весеннецветущей группе относится S. aquilegifolia. Цветение этого вида начинается в первой декаде мая, до фазы “появления свободного листа”. Позже, в первой декаде июня зацветают менее зимостойкие спиреи: S. × arguta, S. myrtilloides, S. nipponica, S. × vanhouttei. Их более позднее в сравнении с видами аборигенной флоры цветение обусловлено более поздней вегетацией в нашем регионе. В среднем начало цветения у спирей, цветочные почки которых закладываются на побегах прошлого года, наблюдается спустя месяц от начала вегетации. Высокой зимостойкостью характеризуются также некоторые представители летнецветущих спирей (S. betulifolia, S. longigemmis) и S. salicifolia – вид, относящийся к группе летне-осеннего цветения. У большинства спирей, представленных в коллекции, отмечено регулярное цветение. Нерегулярно цвели S. crenata и S. nipponica, что связано с сильным обмерзанием кроны зимой 2011 и 2016 гг. Цветение S. × bumalda f. crispa наблюдалось за весь период наблюдений, несмотря на ежегодное обмерзание кроны до уровня снегового покрова.
Продолжительность цветения значительно варьирует, достигая 80–100 дней у ремонтантных видов и форм спирей. Интересен тот факт, что продолжительность цветения у S. japonica и S. salicifolia, интродуцированных на юг Амурской обл., в среднем в 2 раза длиннее, чем у аналогичных видов спирей, культивируемых в степной зоне Южного Урала [13].
Период созревания плодов отмечается от начала формирования до растрескивания листовок. Продолжительность этого периода у разных представителей спирей колеблется от 30 до 130 дней, в большей степени зависит от их биологических особенностей. У большинства зимостойких видов местной флоры, а также некоторых инорайонных (S. alpina, S. crenata, S. longigemmis, S. trilobata) растрескивание листовок и диссеминация происходят до окончания вегетации. В условиях засушливой весны и начала лета (2014, 2015 гг.), а также в годы с ранним началом вегетации (2018 г.) плодоношение у S. elegans, S. media, S. sericea наступало в первой декаде июля, что почти на 60 дней раньше, чем в другие годы наблюдений. Следует отметить, что у S. aquilegifolia в отличие от всех других спирей листовки опадают в течение 2–3 недель после растрескивания и последующей диссеминации. Видимо, с этой особенностью связано неверное указание об отсутствии плодоношения этого вида в культуре АФ БСИ ДВО РАН [14].
Из 22 таксонов спирей регулярно плодоносят 10 видов и 1 представитель гибридного происхождения (S. × bumalda). Нерегулярное плодоношение отмечено у 9 таксонов спирей. Помимо некоторых зимостойких видов (S. pubescens, S. trilobata) сюда относятся те спиреи, для которых характерно прекращение ростовых процессов под воздействием отрицательных температур воздуха. Как правило, формирование зрелых плодов и выполненных семян у спирей с нерегулярным плодоношением (S. × arguta, S. myrtilloides, S. nipponica, S. japonica f. alpina, S. japonica ‘Golden princess’) отмечалось нами в наиболее оптимальные по погодным условиям годы наблюдений.
Однако большинство спирей в условиях культуры АФ БСИ ДВО РАН продуцируют семена высокого качества [15]. Незатрудненный тип прорастания семян в сочетании с высокой продуктивностью растений и достаточно высокими показателями грунтовой всхожести семян (30–60%) позволяет в короткие сроки получить потомство в количествах, необходимых для масштабного озеленения города.
По предварительным результатам проведенных исследований к наиболее устойчивым в агроклиматических условиях юга Амурской обл. мы относим 14 спирей: S. alpina, S. aquilegifolia, S. betulifolia, S. × bumalda, S. elegans, S. japonica, S. longigemmis, S. media, S. pubescens, S. salicifolia, S. sericea, S. trilobata, S. ussuriensis, S. × vanhouttei. Помимо высокой зимостойкости, регулярного цветения и формирования выполненных семян эти спиреи отличаются разной формой кроны и высотой куста, разными сроками и продолжительностью цветения, разной окраской цветка, что позволяет существенно обогатить ассортимент декоративных кустарников, используемых в садово-парковом строительстве города. Для спирей, которые не завязывают семян (S. fritschiana, S. × bumalda f. crispa), необходима разработка рекомендаций по вегетативному размножению, в том числе выявление сроков и способов черенкования.
В литературе указано [16], что сезонный ритм роста и развития многих интродуцированных спирей лабильный и в новых условиях изменяется соответственно характеру сезонных изменений пункта интродукции. Поэтому целесообразным является продолжение опыта интродукции гибридных форм и инорайонных видов спирей, которые за годы наблюдений проявили себя как недостаточно устойчивые. Важным направлением дальнейшей интродукционной работы является обогащение коллекции спирей новыми устойчивыми формами, сортами и экотипами.
ЗАКЛЮЧЕНИЕ
Изучение ритма роста и сезонного развития у 22 таксонов рода Spiraea L., интродуцированных в Амурском филиале Ботанического сада-института ДВО РАН, проведенное в 2012–2018 гг., показало, что сроки окончания вегетации в значительной степени влияют на зимостойкость спирей. S. alpina, S. aquilegifolia, S. betulifolia, S. elegans, S. media, S. salicifolia, S. sericea, S. trilobata, S. ussuriensis оканчивают вегетацию синхронно с местными древесными породами, что приводит к отсутствию обмерзания и позволяет отнести их к зимостойким видам. S. pubescens, S. × bumalda, S. japonica, S. ongigemmis, S. × vanhouttei оканчивают вегетацию до повторных сильных заморозков, обмерзают в неблагоприятные годы, поэтому являются сравнительно зимостойкими. Сравнительно незимостойкие спиреи S. × arguta, S. × bumalda f. crispa, S. crenata, S. ritschiana, S. japonica f. alpina, S. japonica ‘Golden princess’, S. myrtilloides, S. nipponica вегетируют до наступления отрицательных температур, поэтому ежегодно обмерзают, но затем восстанавливаются. Большинство спирей регулярно цветут и плодоносят. Не плодоносят S. × bumalda f. crispa и S. fritschiana. В озеленении Благовещенска рекомендовано использовать зимостойкие и сравнительно зимостойкие спиреи, незимостойкие спиреи в ландшафтном дизайне следует использовать ограниченно.
Список литературы
Гладкова В.Н. 1985. Систематический обзор дикорастущих и широко культивируемых видов рода Spiraea L. (Rosaceae) флоры Европейской части СССР. – Новости сист. высш. раст. 22: 126–133.
Koropachinskiy I.Yu. 2015. North Asian Woody Plants. V. 1. Taxaceae – Rosaceae. P. 495–519.
Александрова М.С. 2000. Аристократы сада: красивоцветущие кустарники. М. С. 138–148.
Тимченко Н.А., Старченко В.М., Дарман Г.Ф. 2017. Атлас деревьев, кустарников и лиан Благовещенска Амурской области: научный справочник. Благовещенск. 254 с., илл.
Лапин П.И., Сиднева С.В. 1973. Оценка перспективности интродукции древесных растений по данным визуальных наблюдений. Опыт интродукции древесных растений. М. С. 7–67.
Ступникова Т.В. 2014. Интродукция восточноазиатских видов рода Spiraea L. в Амурском филиале Ботанического сада-института ДВО РАН. – Плодоводство и ягодоводство России. 40(2): 215–219.
Ступникова Т.В., Семенко В.В. 2018. Оценка коллекции спирей (Spiraea L.) на зимостойкость в агроклиматических условиях юга Амурской области. В сб: “Экология города”: Материалы региональной науч.-практ. конф. Благовещенск. С. 95–100.
Коротаев Г.В. 1991. Особенности климата г. Благовещенска. Благовещенск. 29 с.
Лапин П.И. 1974. Интродукция древесных растений в средней полосе Европейской части СССР. Научные основы, методы, результаты. Л. 135 с.
Булыгин Н.Е. 1979. Фенологические наблюдения над древесными растениями. Л. 97 с.
Нилов В.Н. 1980. К методике статистической обработки материалов фенологических наблюдений. – Бот. журн. 65(2): 282–284.
Булыгин Н.Е. 1987. Дендрология. Методические указания к проведению учебной практики с элементами научных исследований для студентов лесохозяйственного факультета. Л. 48 с.
Кудряшова Н.А. 2005. Биологические особенности декоративных кустарников рода Spiraea L. в условиях степной зоны Южного Урала: Дис. … канд. биол. наук. Оренбург. 148 с.
Костикова В.А., Ступникова Т.В., Воробьева А.Н. 2013. Спиреи на Амуре. – Цветоводство. 4(4): 43–45.
Ступникова Т.В. 2016. Биология семян видов рода Spiraea L., интродуцированных на юг Амурской области. – Плодоводство и ягодоводство России. 47: 321–330.
Шарловская Л.П. 1984. Интродукция видов рода Spiraea L. в Центральный Казахстан: Дис. … канд. биол. наук. Алма-Ата. 221 с.
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы